- Биохимия крови. Функции крови

Содержание
- 2. Функции крови Дыхательная Питательная Выделительная Защитная Регуляторная Транспортная
- 4. РОЭ (СОЭ) мм/час Лошади – 64 Овцы – 0,8 Коровы – 0,58 Свиньи – 30 Кролики
- 5. Физико-химические показатели крови Плотность: 1,050 – 1,060. Вязкость: в 4,5 – 6 раз больше, чем воды.
- 6. Осмотическое давление крови разных животных Депрессия (С°)
- 7. Химический состав сыворотки крови
- 8. Капиллярная система Наиболее полно обмен веществ между кровью и тканями протекает в капиллярной системе. Это обеспечивается
- 9. Вязкость крови
- 10. Зависимость вязкости от количества эритроцитов
- 11. Кислотность крови Все процессы в организме животных могут происходить при строго определенных концентрациях водородных ионов. Даже
- 12. Кислотность крови разных животных
- 13. Ацидоз Понижение щелочного резерва крови называют ацидозом, что наблюдается при кетозах, сердечной недостаточности, патологических изменениях в
- 14. Алкалоз При противоположном ацидозу состоянии – алкалозе – в крови повышается содержание гидрокарбонатов, избыток которых выделяется
- 15. Буферные системы крови
- 16. Гидрокарбонатная буферная система Гидрокарбонатная буферная система крови состоит из сочетания СО2 , Н2СО3 , NaHCO3. Второй
- 17. А. Нормальные эритроциты в форме двояковогнутого диска; Б. Сморщенные эритроциты в гипертоническом солевом растворе.
- 18. Гемоглобиновая буферная система Гемоглобиновая буферная система — самая мощная буферная система крови. Она в 9 раз
- 19. Буферные свойства гемоглобина Буферные свойства гемоглобина прежде всего обусловлены возможностью взаимодействия кислореагирующих соединений с калиевой солью
- 20. Гемоглобиновый буфер
- 21. Белковая буферная система Белковая буферная система имеет меньшее значение для поддержания кислотно-основного равновесия в плазме крови,
- 22. Буферное действие белков Буферное действие белков плазмы невелико (10% буферной емкости крови ), тогда как гемоглобин
- 23. Фосфатная буферная система Фосфатная буферная система представляет собой сопряженную кислотно-основную пару, состоящую из иона Н2РО4- (донор
- 24. Белковый состав плазмы Белки неоднородные по своему составу, физико-химическим и биологическим свойствам. Методом электрофореза на бумаге
- 25. Содержание белков в плазме крови животных
- 26. Электрофолеграмма сыворотки крови человека. А. Схема прибора для электрофореза на бумаге б.
- 27. Электрофорез белков сыворотки
- 28. Белковые фракции плазмы крови человека
- 29. Роль белков крови Создают онкотическое давление. Участвуют в свертывании крови. Создают определенную вязкость крови, ответственны за
- 30. Специальные белки плазмы. Гаптоглобин. Гаптоглобин входит в состав a2-глобулиновой фракции. Этот белок обладает способностью соединяться с
- 31. Ингибиторы трипсина Ингибиторы трипсина обнаруживаются при электрофорезе белков плазмы крови в зоне α1 и α2-глобулинов; они
- 32. Трансферрин Трансферрин относится к β-глобулинам и обладает способностью соединяться с железом. Его комплекс с железом окрашен
- 33. Церулоплазмин Церулоплазмин имеет голубоватый цвет, обусловленный наличием в его составе 0,32% меди. Обладает слабой каталитической активностью,
- 34. С-реактивный белок С-реактивный белок получил свое название в результате способности вступать в реакцию преципитации с С-полисахаридом
- 35. Криоглобулин Криоглобулин в сыворотке крови здоровых людей также отсутствует и появляется в ней при патологических состояниях.
- 36. Интерферон Интерферон — специфический белок, синтезируемый в клетках организма в результате воздействия вирусов. В свою очередь
- 37. Ферменты плазмы Ферменты, которые обнаруживаются в норме в плазме или сыворотке крови, условно можно разделить на
- 38. Ферменты плазмы Экскреторные ферменты синтезируются главным образом в печени (лецинаминопентидаза, щелочная фосфатаза и др.). Эти ферменты
- 39. Ферменты крови Ферменты крови. В плазме и сыворотки крови всегда имеется некоторое количество ферментов, причем одни
- 40. Ферменты крови Так, при ряде заболеваний в крови резко повышается активность амилазы (при поражении поджелудочной железы),
- 41. Нарушения содержания белков Гипопротеинемия – снижение концентрации белков в крови (при белковом голодании, приеме больших количеств
- 42. Нарушения содержания белков Диспротеинемия – нарушение соотношения концентраций отдельных фракций белков крови при неизменной общей концентрации
- 43. Содержание азотистых веществ в плазме крови животных, мг %.
- 44. Мочевина Мочевина – главный компонент остаточного азота – составляет 40-50%, а по некоторым другим данным –
- 45. Мочевая кислота Мочевая кислота является главным конечным продуктом белкового обмена у птиц и её содержание в
- 46. Аминокислоты Аминокислоты в количественном отношении стоят на втором месте после мочевины среди составных частей остаточного азота.
- 47. Содержание безазотистых веществ
- 48. Содержание глюкозы Содержание углеводов (глюкозы) в крови относительно постоянно для каждого вида животных. Это постоянство поддерживается
- 49. Содержание лактата и пвк Постоянная составная часть крови — молочная кислота. Ее количество может увеличиваться в
- 50. Содержание липидов Липидов в крови содержится от 0,5 до 0,9 %, а в некоторых случаях достигает
- 51. Липопротеины плазмы крови
- 52. Общая характеристика липопротеинов Липиды в водной среде (а значит, и в крови) нерастворимы, поэтому для транспорта
- 53. Липопротеины В организме синтезируются следующие типы липопротеинов: хиломикроны (ХМ), липопротеины очень низкой плотности (ЛПОНП), липопротеины промежуточной
- 54. Свойства липопротеинов ЛП хорошо растворимы в крови, не опалесцируют, так как имеют небольшой размер и отрицательный
- 55. Гиперхиломикронемия, гипертриглицеронемия После приёма пищи, содержащей жиры, развивается физиологическая гипертриглицеронемия и, соответственно, гиперхиломикронемия, которая может продолжаться
- 56. Атерогенность липопротеинов ЛПНП – атерогенные ЛПОНП – атерогенные ЛПВП - неатерогенные
- 58. Скачать презентацию

Функции крови
Дыхательная
Питательная
Выделительная
Защитная
Регуляторная
Транспортная
Функции крови
Дыхательная
Питательная
Выделительная
Защитная
Регуляторная
Транспортная
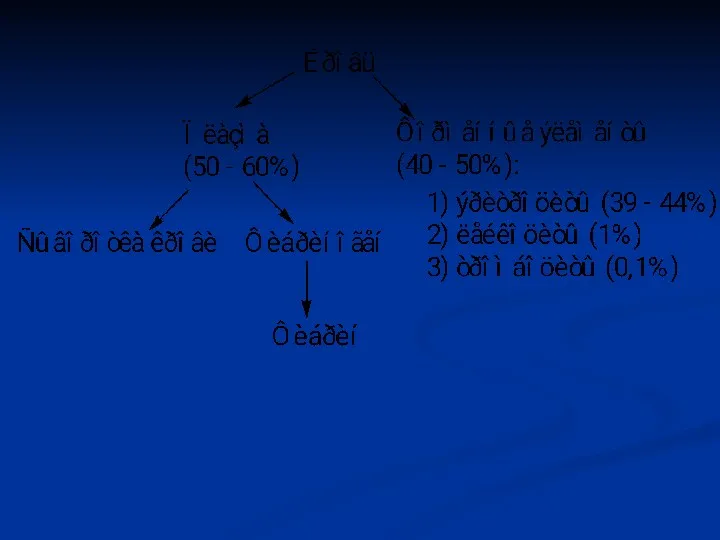

РОЭ (СОЭ) мм/час
Лошади – 64
Овцы – 0,8
Коровы – 0,58
Свиньи – 30
РОЭ (СОЭ) мм/час
Лошади – 64
Овцы – 0,8
Коровы – 0,58
Свиньи – 30

Физико-химические показатели крови
Плотность: 1,050 – 1,060.
Вязкость: в 4,5 – 6 раз
Физико-химические показатели крови
Плотность: 1,050 – 1,060.
Вязкость: в 4,5 – 6 раз
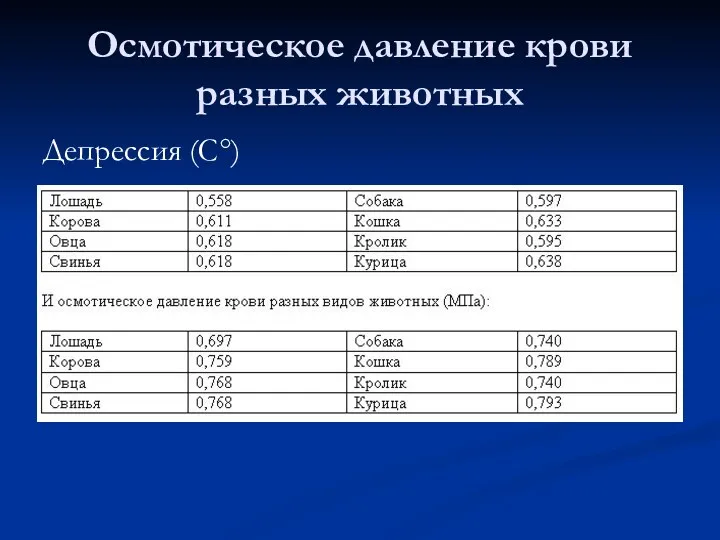
Осмотическое давление крови разных животных
Депрессия (С°)
Осмотическое давление крови разных животных
Депрессия (С°)
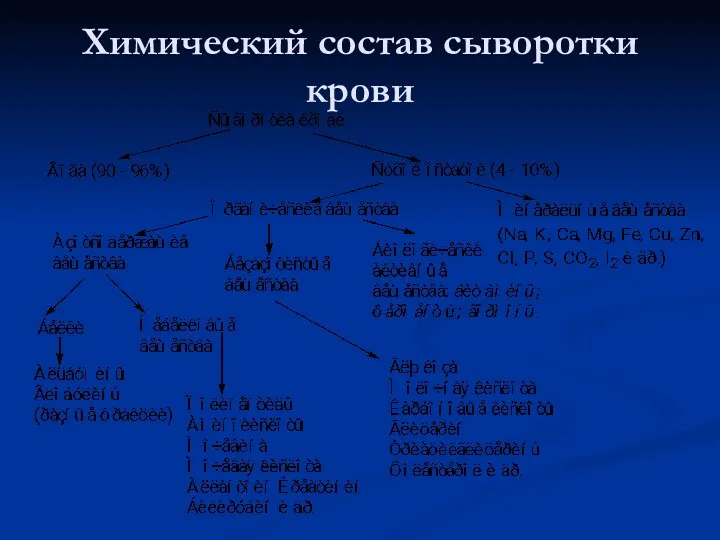
Химический состав сыворотки крови
Химический состав сыворотки крови

Капиллярная система
Наиболее полно обмен веществ между кровью и тканями протекает в
Капиллярная система
Наиболее полно обмен веществ между кровью и тканями протекает в
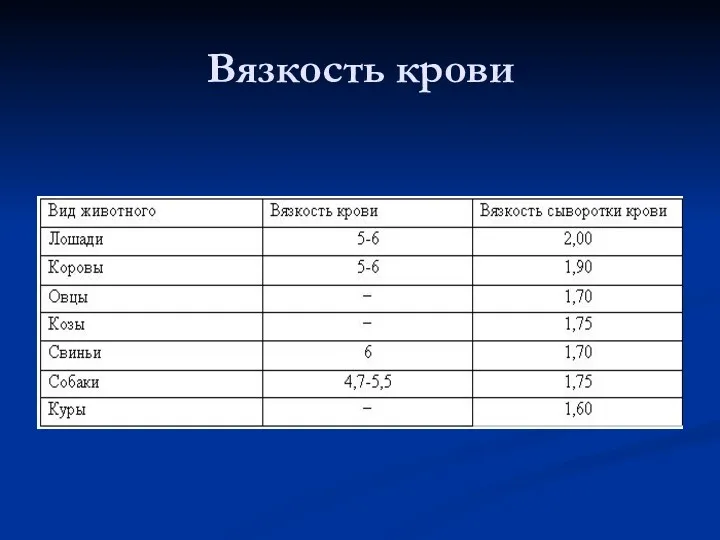
Вязкость крови
Вязкость крови
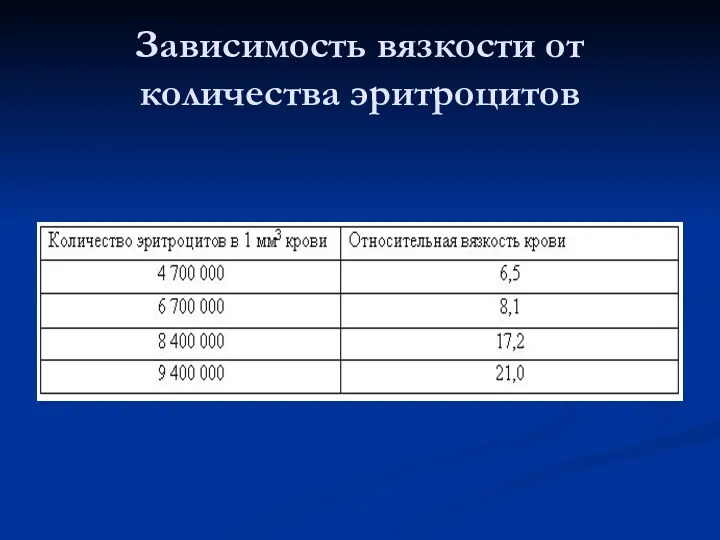
Зависимость вязкости от количества эритроцитов
Зависимость вязкости от количества эритроцитов

Кислотность крови
Все процессы в организме животных могут происходить при строго определенных
Кислотность крови
Все процессы в организме животных могут происходить при строго определенных
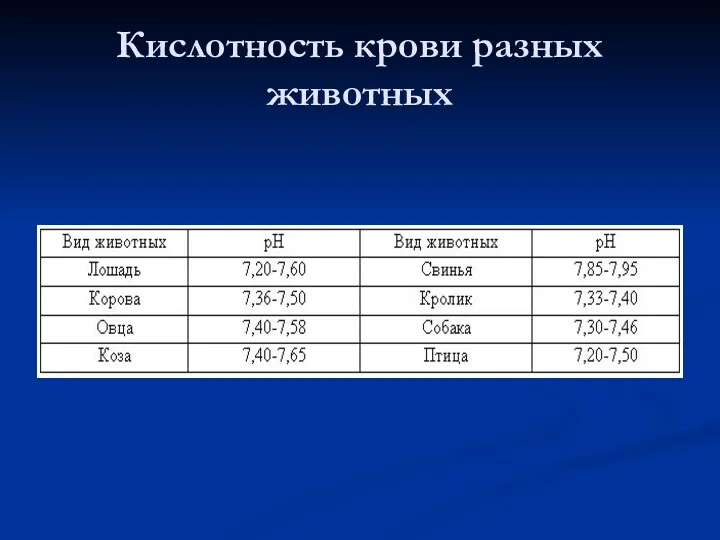
Кислотность крови разных животных
Кислотность крови разных животных

Ацидоз
Понижение щелочного резерва крови называют ацидозом, что наблюдается при кетозах, сердечной
Ацидоз
Понижение щелочного резерва крови называют ацидозом, что наблюдается при кетозах, сердечной

Алкалоз
При противоположном ацидозу состоянии – алкалозе – в крови повышается содержание
Алкалоз
При противоположном ацидозу состоянии – алкалозе – в крови повышается содержание

Буферные системы крови
Буферные системы крови

Гидрокарбонатная буферная система
Гидрокарбонатная буферная система крови состоит из сочетания СО2 ,
Гидрокарбонатная буферная система
Гидрокарбонатная буферная система крови состоит из сочетания СО2 ,
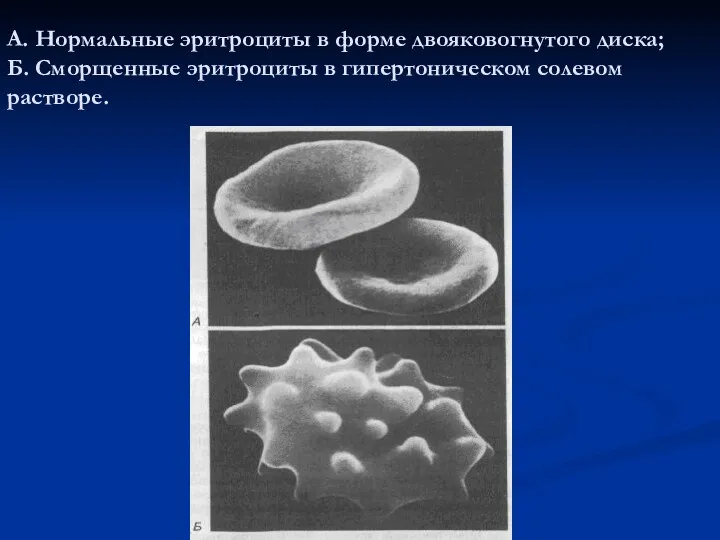
А. Нормальные эритроциты в форме двояковогнутого диска;
Б. Сморщенные эритроциты в
А. Нормальные эритроциты в форме двояковогнутого диска; Б. Сморщенные эритроциты в

Гемоглобиновая буферная система
Гемоглобиновая буферная система — самая мощная буферная система крови.
Гемоглобиновая буферная система
Гемоглобиновая буферная система — самая мощная буферная система крови.

Буферные свойства гемоглобина
Буферные свойства гемоглобина прежде всего обусловлены возможностью взаимодействия кислореагирующих
Буферные свойства гемоглобина
Буферные свойства гемоглобина прежде всего обусловлены возможностью взаимодействия кислореагирующих

Гемоглобиновый буфер
Гемоглобиновый буфер

Белковая буферная система
Белковая буферная система имеет меньшее значение для поддержания кислотно-основного
Белковая буферная система
Белковая буферная система имеет меньшее значение для поддержания кислотно-основного

Буферное действие белков
Буферное действие белков плазмы невелико (10% буферной емкости крови
Буферное действие белков
Буферное действие белков плазмы невелико (10% буферной емкости крови

Фосфатная буферная система
Фосфатная буферная система представляет собой сопряженную кислотно-основную пару, состоящую
Фосфатная буферная система
Фосфатная буферная система представляет собой сопряженную кислотно-основную пару, состоящую

Белковый состав плазмы
Белки неоднородные по своему составу, физико-химическим и биологическим свойствам.
Белковый состав плазмы
Белки неоднородные по своему составу, физико-химическим и биологическим свойствам.
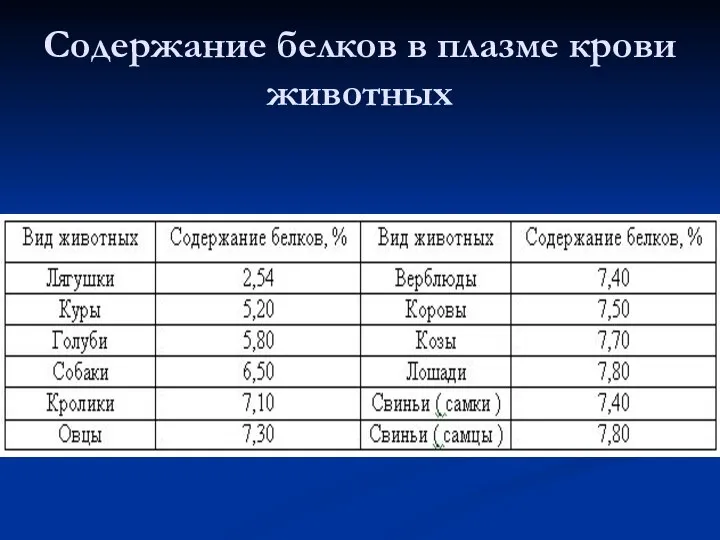
Содержание белков в плазме крови животных
Содержание белков в плазме крови животных
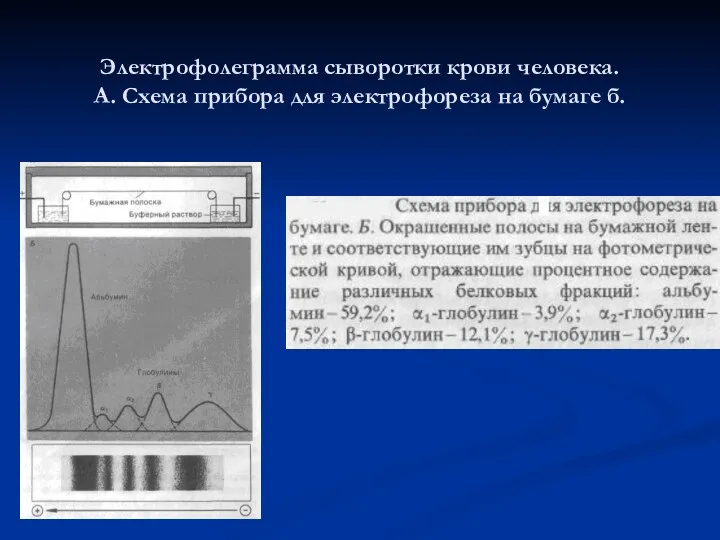
Электрофолеграмма сыворотки крови человека.
А. Схема прибора для электрофореза на бумаге
Электрофолеграмма сыворотки крови человека. А. Схема прибора для электрофореза на бумаге

Электрофорез белков сыворотки
Электрофорез белков сыворотки
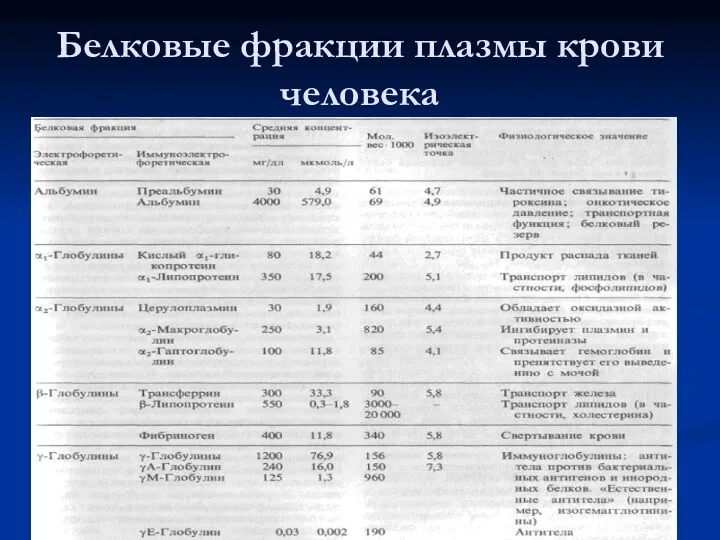
Белковые фракции плазмы крови человека
Белковые фракции плазмы крови человека

Роль белков крови
Создают онкотическое давление.
Участвуют в свертывании крови.
Создают определенную вязкость крови,
Роль белков крови
Создают онкотическое давление.
Участвуют в свертывании крови.
Создают определенную вязкость крови,

Специальные белки плазмы. Гаптоглобин.
Гаптоглобин входит в состав a2-глобулиновой фракции. Этот белок
Специальные белки плазмы. Гаптоглобин.
Гаптоглобин входит в состав a2-глобулиновой фракции. Этот белок

Ингибиторы трипсина
Ингибиторы трипсина обнаруживаются при электрофорезе белков плазмы крови в зоне
Ингибиторы трипсина
Ингибиторы трипсина обнаруживаются при электрофорезе белков плазмы крови в зоне

Трансферрин
Трансферрин относится к β-глобулинам и обладает способностью соединяться с железом. Его
Трансферрин
Трансферрин относится к β-глобулинам и обладает способностью соединяться с железом. Его

Церулоплазмин
Церулоплазмин имеет голубоватый цвет, обусловленный наличием в его составе 0,32% меди.
Церулоплазмин
Церулоплазмин имеет голубоватый цвет, обусловленный наличием в его составе 0,32% меди.

С-реактивный белок
С-реактивный белок получил свое название в результате способности вступать в
С-реактивный белок
С-реактивный белок получил свое название в результате способности вступать в

Криоглобулин
Криоглобулин в сыворотке крови здоровых людей также отсутствует и появляется в
Криоглобулин
Криоглобулин в сыворотке крови здоровых людей также отсутствует и появляется в

Интерферон
Интерферон — специфический белок, синтезируемый в клетках организма в результате воздействия
Интерферон
Интерферон — специфический белок, синтезируемый в клетках организма в результате воздействия

Ферменты плазмы
Ферменты, которые обнаруживаются в норме в плазме или сыворотке крови,
Ферменты плазмы
Ферменты, которые обнаруживаются в норме в плазме или сыворотке крови,

Ферменты плазмы
Экскреторные ферменты синтезируются главным образом в печени (лецинаминопентидаза, щелочная фосфатаза
Ферменты плазмы
Экскреторные ферменты синтезируются главным образом в печени (лецинаминопентидаза, щелочная фосфатаза

Ферменты крови
Ферменты крови. В плазме и сыворотки крови всегда имеется некоторое
Ферменты крови
Ферменты крови. В плазме и сыворотки крови всегда имеется некоторое

Ферменты крови
Так, при ряде заболеваний в крови резко повышается активность амилазы
Ферменты крови
Так, при ряде заболеваний в крови резко повышается активность амилазы

Нарушения содержания белков
Гипопротеинемия – снижение концентрации белков в крови (при белковом
Нарушения содержания белков
Гипопротеинемия – снижение концентрации белков в крови (при белковом

Нарушения содержания белков
Диспротеинемия – нарушение соотношения концентраций отдельных фракций белков крови
Нарушения содержания белков
Диспротеинемия – нарушение соотношения концентраций отдельных фракций белков крови

Содержание азотистых веществ в плазме крови животных, мг %.
Содержание азотистых веществ в плазме крови животных, мг %.

Мочевина
Мочевина – главный компонент остаточного азота – составляет 40-50%, а по
Мочевина
Мочевина – главный компонент остаточного азота – составляет 40-50%, а по

Мочевая кислота
Мочевая кислота является главным конечным продуктом белкового обмена у птиц
Мочевая кислота
Мочевая кислота является главным конечным продуктом белкового обмена у птиц

Аминокислоты
Аминокислоты в количественном отношении стоят на втором месте после мочевины среди
Аминокислоты
Аминокислоты в количественном отношении стоят на втором месте после мочевины среди
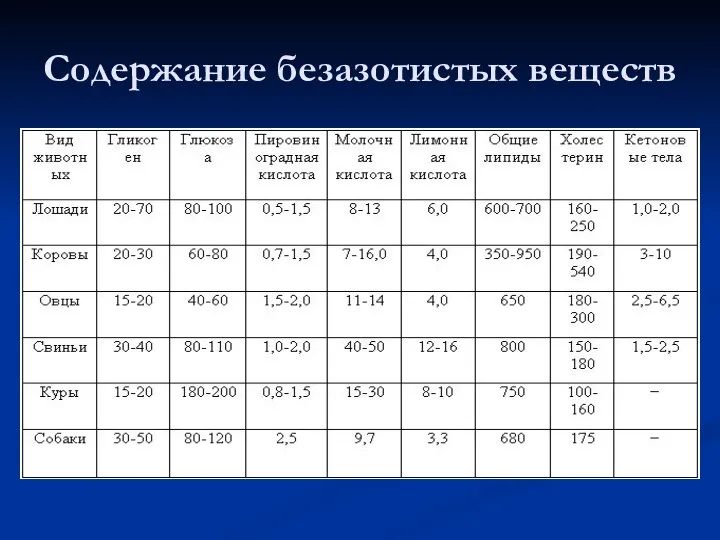
Содержание безазотистых веществ
Содержание безазотистых веществ

Содержание глюкозы
Содержание углеводов (глюкозы) в крови относительно постоянно для каждого вида
Содержание глюкозы
Содержание углеводов (глюкозы) в крови относительно постоянно для каждого вида

Содержание лактата и пвк
Постоянная составная часть крови — молочная кислота. Ее
Содержание лактата и пвк
Постоянная составная часть крови — молочная кислота. Ее

Содержание липидов
Липидов в крови содержится от 0,5 до 0,9 %, а
Содержание липидов
Липидов в крови содержится от 0,5 до 0,9 %, а
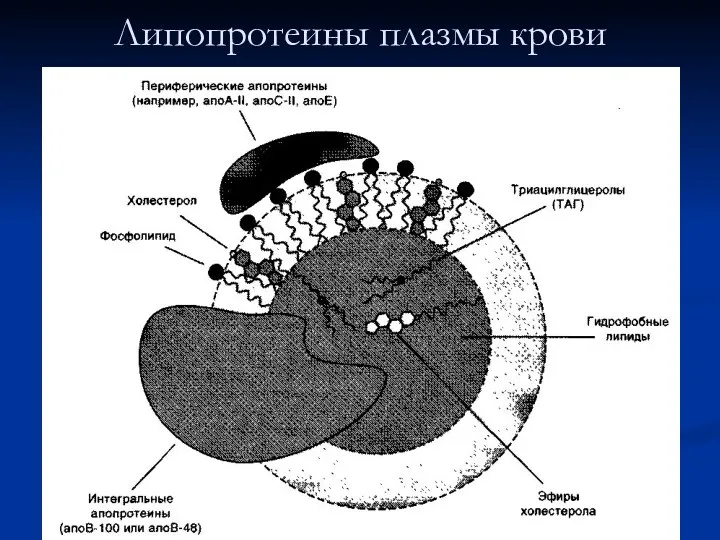
Липопротеины плазмы крови
Липопротеины плазмы крови

Общая характеристика липопротеинов
Липиды в водной среде (а значит, и в крови)
Общая характеристика липопротеинов
Липиды в водной среде (а значит, и в крови)

Липопротеины
В организме синтезируются следующие типы липопротеинов: хиломикроны (ХМ), липопротеины очень низкой
Липопротеины
В организме синтезируются следующие типы липопротеинов: хиломикроны (ХМ), липопротеины очень низкой

Свойства липопротеинов
ЛП хорошо растворимы в крови, не опалесцируют, так как имеют
Свойства липопротеинов
ЛП хорошо растворимы в крови, не опалесцируют, так как имеют

Гиперхиломикронемия, гипертриглицеронемия
После приёма пищи, содержащей жиры, развивается физиологическая гипертриглицеронемия и, соответственно,
Гиперхиломикронемия, гипертриглицеронемия
После приёма пищи, содержащей жиры, развивается физиологическая гипертриглицеронемия и, соответственно,

Атерогенность липопротеинов
ЛПНП – атерогенные
ЛПОНП – атерогенные
ЛПВП - неатерогенные
Атерогенность липопротеинов
ЛПНП – атерогенные
ЛПОНП – атерогенные
ЛПВП - неатерогенные
 Презентация к уроку биологии 6 класс
Презентация к уроку биологии 6 класс Мышечная система человека
Мышечная система человека Зона степей России. Животный и растительный мир
Зона степей России. Животный и растительный мир Строение спинного мозга человека
Строение спинного мозга человека Мимические мышцы лица
Мимические мышцы лица Биосфера
Биосфера Мир растений
Мир растений возникновение жизни главная
возникновение жизни главная Проблема индивидуальности человека в современной науке
Проблема индивидуальности человека в современной науке Фотосинтез
Фотосинтез Комнатные растения
Комнатные растения Строение и функции головного мозга
Строение и функции головного мозга Функциональные системы организма. Биотехнические системы и технологии
Функциональные системы организма. Биотехнические системы и технологии Зоология - наука о животных. 4 царства живой природы
Зоология - наука о животных. 4 царства живой природы Минеральное питание
Минеральное питание ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Тест Рыбы 7 класс.
ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Тест Рыбы 7 класс. Биохимия эмоций. Стресс
Биохимия эмоций. Стресс Животные Австралии
Животные Австралии Тип Хордовые (chordata)
Тип Хордовые (chordata) Общая характеристика грибов
Общая характеристика грибов Онтогенез нервной системы
Онтогенез нервной системы Птицы. Викторина
Птицы. Викторина Processes of Respiration
Processes of Respiration Жертвы эволюции
Жертвы эволюции строение и функции мыщц
строение и функции мыщц Естественный отбор и его формы
Естественный отбор и его формы Биологическая роль s-элементов
Биологическая роль s-элементов Ссавці. Класифікація ссавців
Ссавці. Класифікація ссавців