- Минеральное питание

Содержание
- 2. Относительное содержание элементов в растениях
- 3. Восстановление нитратов
- 4. Нитратредуктаза: схема работы, домены и структура молибдоптерина НАДН, НАД(Ф)Н (высшие растения, водоросли), НАДФН (грибы) –специфичная Nr,
- 5. Нитратредуктаза: строение
- 6. Регуляция работы нитратредуктазы: на уровне транскрипции, трансляции и регулировании активности На уровне транскрипции: индукция субстратом (50
- 7. Нитритредуктаза: строение В гетеротрофных клетках – ферредоксин за счет ПФ-шунта: Ферредоксин-НАДФ+-редуктаза Мономер, 60 – 70кДа. Активность
- 8. Как растение и бактерии «узнают» друг друга
- 9. Флавоноид + белок NodD активирует транскрипцию nod-генов
- 10. Продукт активности nod-генов - nod-фактор и инфекционный процесс NodA –NodC гены кодируют ферменты, которые обуславливают формирование
- 11. Формирование бактероидов
- 12. Электронная микрофотография клеток клубенька соевых бобов Инфициро-ванная клетка с итактными симбиосомами содержащими один или два бактероида.
- 13. Структура нитрогеназы Структура Mo-Fe-S-активного центра. Место связывания азота – предположительно… Еще одни Камикадзе… Fe-белок необратимо инактивируется
- 14. В ядре клеток растения активируются гены, необходимые для симбиоза Легоглобин – белок синтезируется растением, гем –
- 15. Включение азота в органические соединения. Система ГС – ГТС (ГОГАТ) ГОГАТ: Мв ~ 150 kDa. Пластиды.
- 16. Различные изозимы работают в разных компартментах клетки
- 17. Втория система: глутаматдегидрогеназа. Каковы ее функции? Для синтеза: Кm NH4 = 10 – 80 mM. Такой
- 18. Итак, включение азота в органические соединения Аспарагин – амид с высоким удельным содержанием азота: на четыре
- 19. «Добыча» азота растением – очень дорогой процесс… На один атом ассимилированного азота идет минимум 12 АТФ.
- 20. Транспорт азота осуществляется различными соединениями Только у травянистых растений транспорт азота может осуществляться в виде NO3
- 21. Включение азота в органические соединения при азотфиксации
- 22. Относительное содержание нитратов и других азотсодержащих соединений в ксилемном экссудате. Звездчатка Дурнишник Xhantum spp. Недотрога
- 23. Сера, в отличие от азота, «работает» в разных степенях окисления
- 24. Метаболизм серы В цитозоле и пластидах – разные задачи
- 25. Восстановление сульфатов. Основные ферменты. 1. АФС-сурфурилаза: SO42- + ATP = APS + PPi Реакция термодинамически не
- 26. Обмен серы в растении
- 27. Поглощение ионов корнем
- 28. Проницаемость биологических мембран
- 29. Поступление в клетку. Перенос через плазмалемму. Электрофизиологический подход Кинетический подход Молекулярно биологический подход
- 30. Электрофизиологический подход Потенциал Нернста может быть использован, чтобы установить тип транспорта: активный или пассивный? μ =
- 31. Варианты транспортных процессов через мембрану Три класса мембранных транспортных белков: каналы, переносчики и насосы. Каналы и
- 32. Модель первично-активного транспорта. Насосы (помпы) – используют для транспорта АТФ «напрямую»
- 33. Модель вторично-активного транспорта. Переносчики (транспортеры). Используют энергию протонного (или Na+) градиента
- 34. Принципиальная схема транспортных процессов растительной клетки
- 35. Если бы все транспортировалось пассивно… Измеренные и предсказанные как равновесные концентрации ионов (mМ) в тканях корней
- 36. Схема транспортных систем растительной клетки Насосы Каналы Транспортеры
- 37. Три типа АТФ-аз растительной клетки: F-, p- и V-типа
- 38. Структура белка Н-АТФазы плазмалеммы Один полипептид 100 – 106 кДа Домен связывания Mg-АТФ Е1 связывает Н
- 39. Свойства и регулирование активности АТФ-зы р-типа Существует более десятка изоферментов АТФ-зы р-типа Один большой полипептид 100
- 40. 3 каталитических центра связывания АТФ, 6-9 субъединиц, связывающих Н+ Многомерная структура 70 кДа, 60 кДа, 16
- 41. Стехиометрия переноса: 1 пирофосфат : 1 Н+ H+-пирофосфатаза (дифосфатаза) тонопласта 64 – 67 кДа катионзависимая (стимулируется
- 42. Са2+-АТФ-за – еще один «насос» плазмалеммы и тонопласта, но совсем с другими функциями Са2+АТФазы принадлежат к
- 43. Транспорт катионов. Типы калиевых каналов растительной клетки. IRK - Inwardly-Rectifying К+ 1 белок TWIK или TPC
- 44. Калиевые каналы Shaker-типа могут регулируются мембранным потенциалом или циклическими нуклеотидами, работают в виде тетрамеров
- 45. Транспорт калия через мембраны идет не только через каналы, но и через переносчики (транспортеры). Схема AtKUP1
- 46. «Дерево» всех К+-транспортеров A. thaliana с 5 основными «веточками» a) KUP/HAK/KT transporters (13 genes), b) Trk/HKT
- 47. Общая схема транспортеров MFS (Major Facilitator Super family) В геноме арабидопсиса найдено около 600 мембранных транспортеров
- 48. Сульфатные транспортеры Высокого сродства Sultr1 или ST1 (Km порядка 10 мкмолей) Низкого сродства Sultr2 или ST2
- 49. Клонированные NO3-/ NO2- транспортеры NNP (nitrate –nitrite porters) и PTR (peptide transporter)
- 50. Три типа NO3-/ NO2- транспортеров NNP-семейства
- 51. Аммонийные транспортеры AtMT1. Унипортер, Кm = 65мкМ. AtMT2 SAT1 – для транспорта из бактероидов
- 52. Схема строения сульфатных и фосфатных транспортеров
- 53. Схема строения и регулирования высокоафинного фосфатного транспортера
- 54. Гены фосфатных транспортеров арабидопсиса TPT – Триозофосфат - Pi – Транспортер РРТ – Фосфоенолпируват - Pi
- 55. Нозерн-блот двух фосфатных транспортеров из корней и листьев помидоров.
- 56. Везикулярно-арбускулярная микориза необходима прежде всего для поглощения фосфатов
- 57. Катион/протон антипортеры, обнаруженные у Arabidopsis
- 58. Схема строения транспортеров ZIP-семейства
- 59. «Дерево» ZIP-транспортеров A. tailana Семейство ZIP-транспортеров (Zinc and Iron regulated transporter Proteins) – транспортеры металлов, могут
- 60. ABC-транспортеры Транспортные белки, которые используют энергию гидролиза АТФ для транспорта через мембраны самых разных химических агентов
- 61. Схема транспортных систем клеток корня
- 62. Две стратегии поглощения железа растениями Стратегия I: двудольные, незлаковые однодольные, а также дрожжи Стратегия II -
- 63. Стратегия I. Fe3+ восстанавливается, расположенной на ПМ Fe3+ хелат-редуктазой (FRO2 феррик редуктаза) (Fe2+) переносится через мембрану
- 64. Восстановление Fe(III) и Cu(II) вдоль поверхности первичных боковых корней 14-дневных растений гороха. Fe(III) Cu(II) Усиление при
- 65. Феррохелатаза
- 66. Предполагаемая структура белка FRO2 феррик редуктазы плазмалеммы арабидопсис
- 67. Стратегия II, обеспечивает поглощение Fe, за счет образования специальных хелатирующих соединений – фитосидерофоров. Фитосидерофоры – низкомолекулярные
- 68. Структура фитосидерофоров
- 70. Скачать презентацию
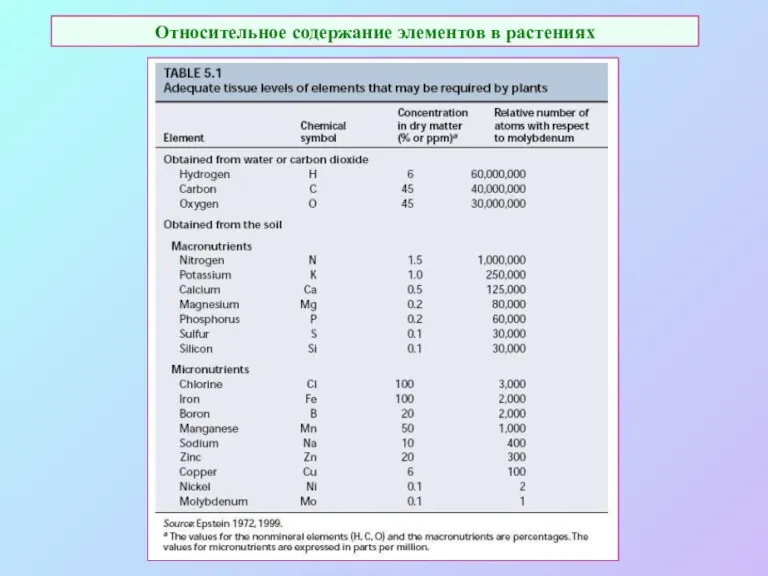
Относительное содержание элементов в растениях
Относительное содержание элементов в растениях
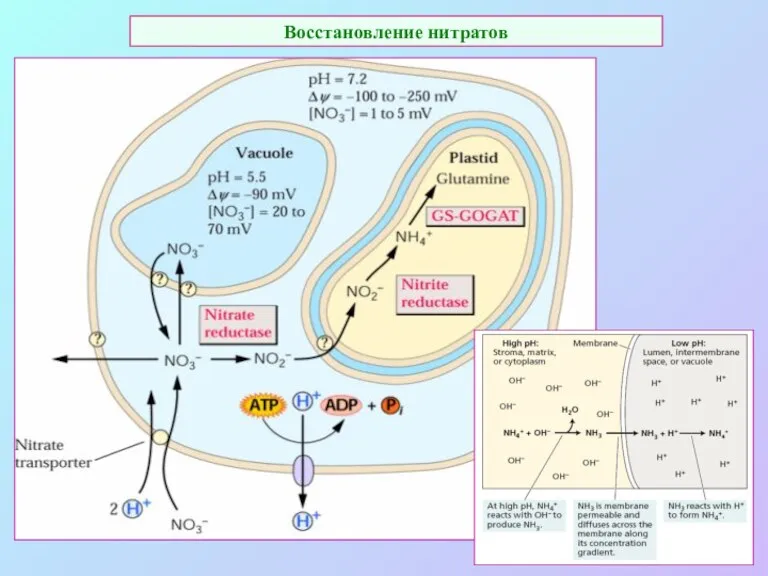
Восстановление нитратов
Восстановление нитратов
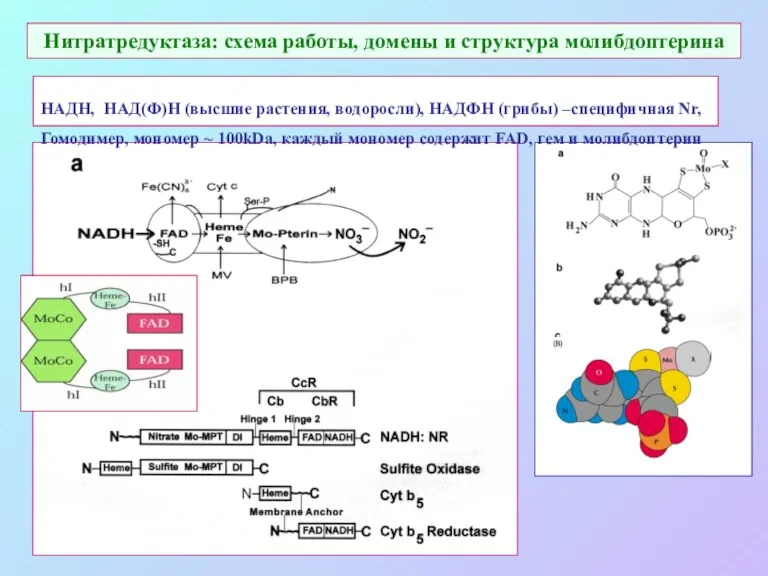
Нитратредуктаза: схема работы, домены и структура молибдоптерина
НАДН, НАД(Ф)Н (высшие растения, водоросли),
Нитратредуктаза: схема работы, домены и структура молибдоптерина
НАДН, НАД(Ф)Н (высшие растения, водоросли),
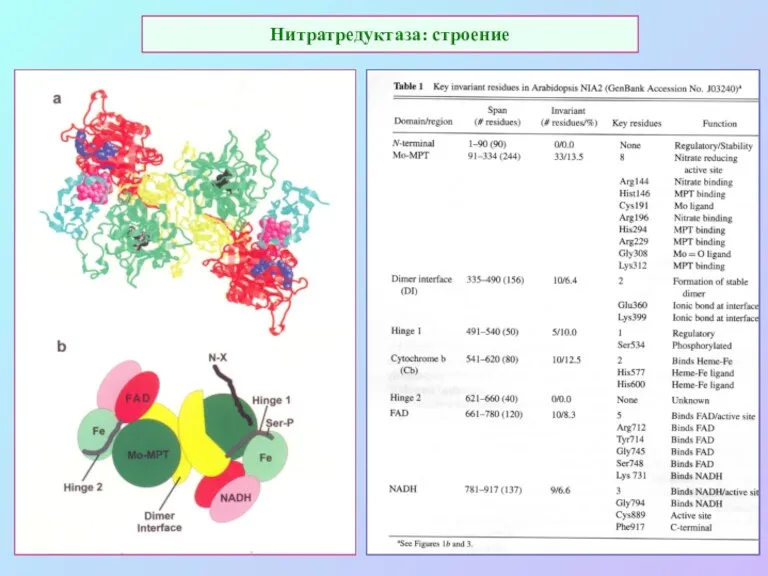
Нитратредуктаза: строение
Нитратредуктаза: строение
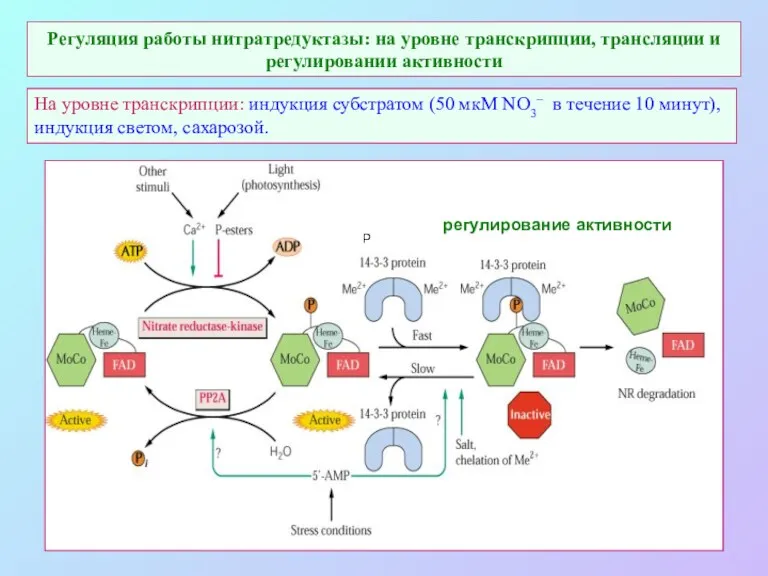
Регуляция работы нитратредуктазы: на уровне транскрипции, трансляции и регулировании активности
На уровне
Регуляция работы нитратредуктазы: на уровне транскрипции, трансляции и регулировании активности
На уровне
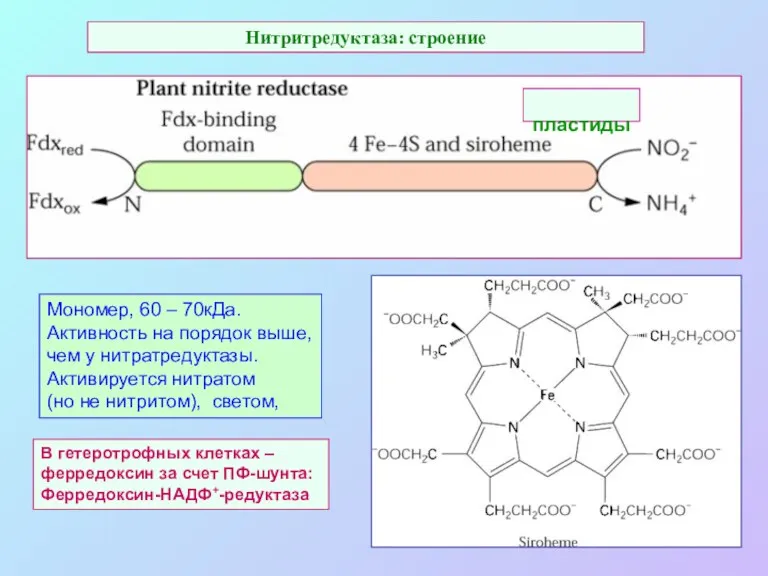
Нитритредуктаза: строение
В гетеротрофных клетках –
ферредоксин за счет ПФ-шунта:
Ферредоксин-НАДФ+-редуктаза
Мономер, 60 –
Нитритредуктаза: строение
В гетеротрофных клетках –
ферредоксин за счет ПФ-шунта:
Ферредоксин-НАДФ+-редуктаза
Мономер, 60 –
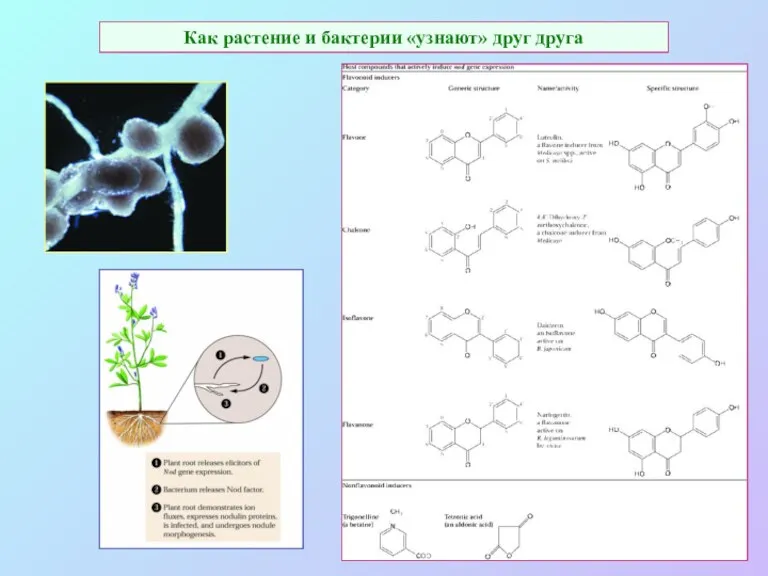
Как растение и бактерии «узнают» друг друга
Как растение и бактерии «узнают» друг друга
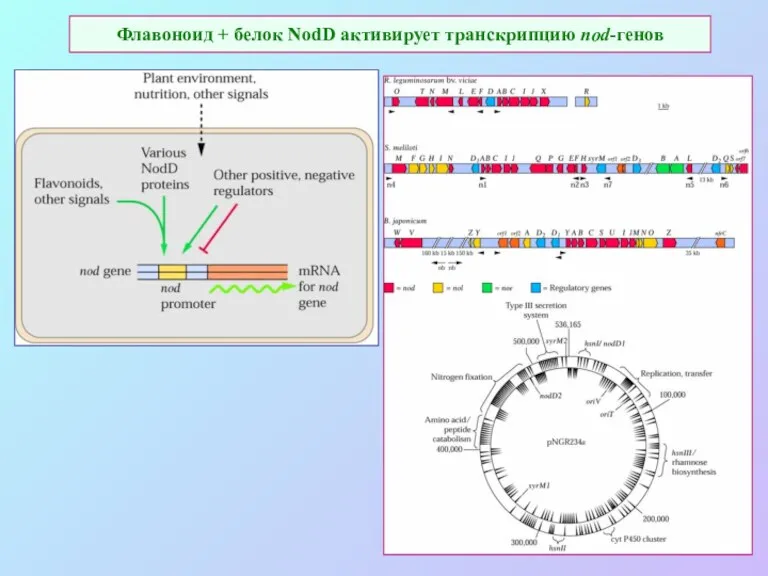
Флавоноид + белок NodD активирует транскрипцию nod-генов
Флавоноид + белок NodD активирует транскрипцию nod-генов
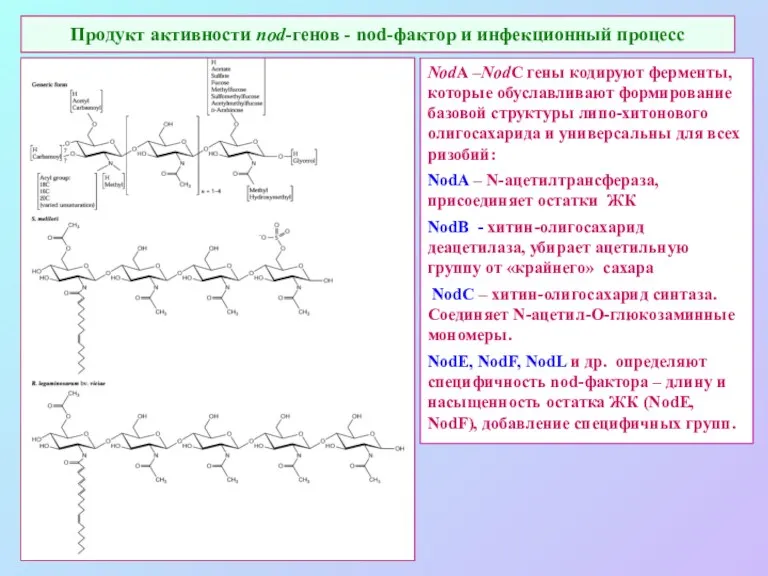
Продукт активности nod-генов - nod-фактор и инфекционный процесс
NodA –NodC гены кодируют
Продукт активности nod-генов - nod-фактор и инфекционный процесс
NodA –NodC гены кодируют
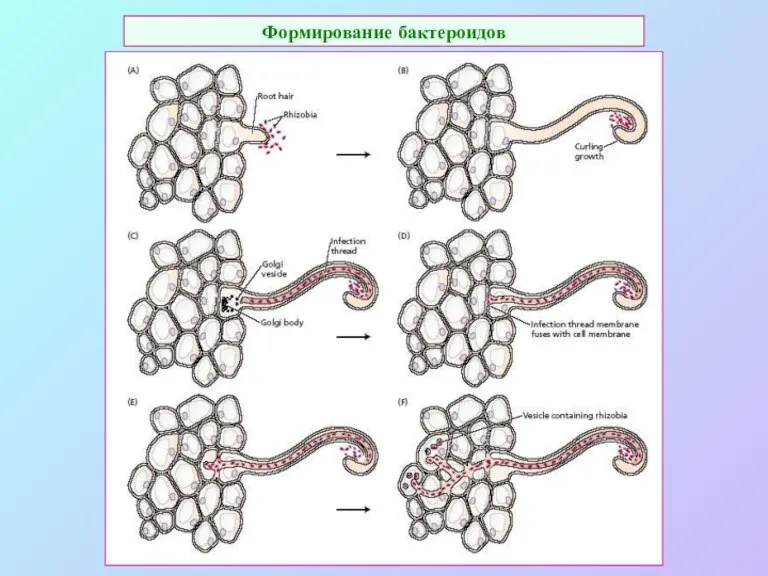
Формирование бактероидов
Формирование бактероидов
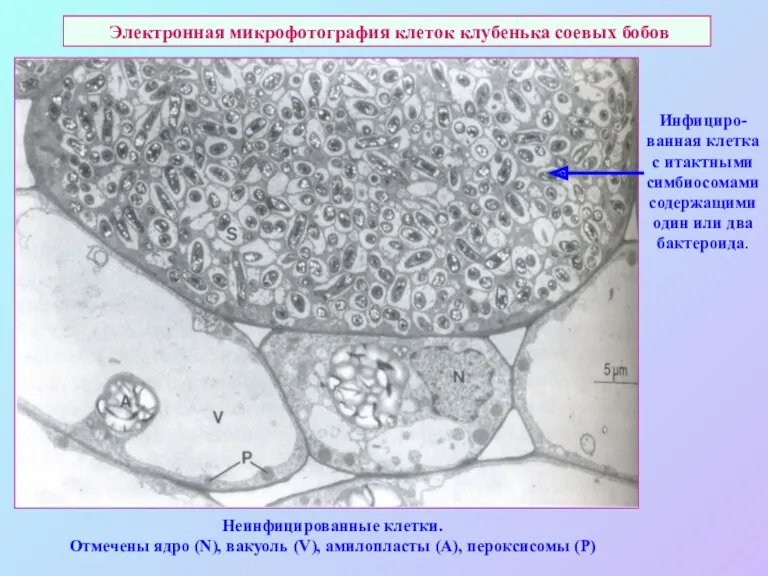
Электронная микрофотография клеток клубенька соевых бобов
Инфициро-ванная клетка с итактными симбиосомами
Электронная микрофотография клеток клубенька соевых бобов
Инфициро-ванная клетка с итактными симбиосомами
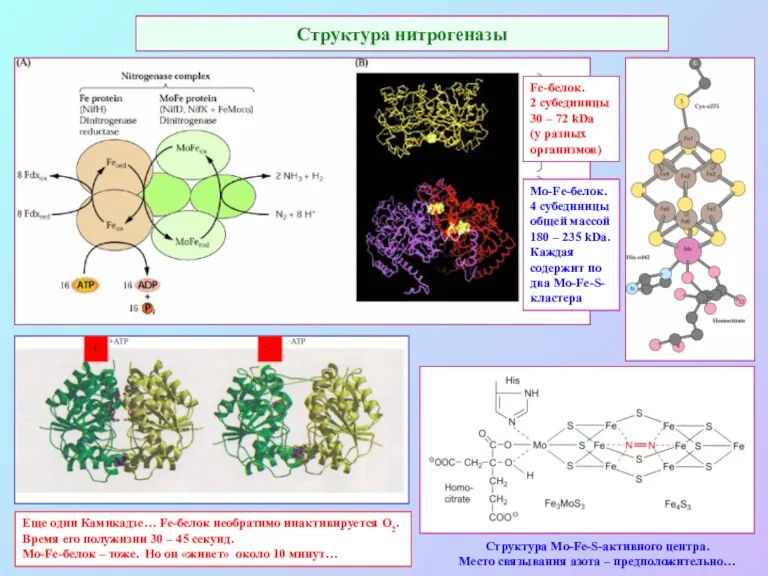
Структура нитрогеназы
Структура Mo-Fe-S-активного центра.
Место связывания азота – предположительно…
Еще одни Камикадзе…
Структура нитрогеназы
Структура Mo-Fe-S-активного центра.
Место связывания азота – предположительно…
Еще одни Камикадзе…
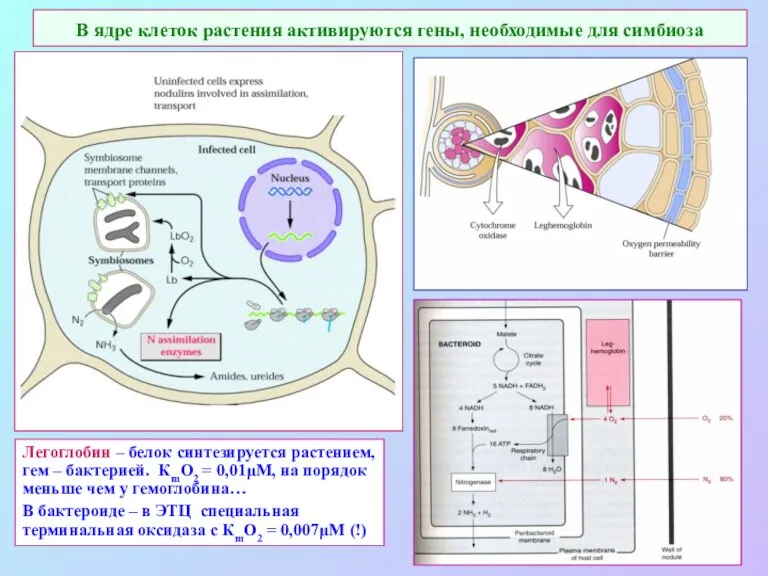
В ядре клеток растения активируются гены, необходимые для симбиоза
Легоглобин –
В ядре клеток растения активируются гены, необходимые для симбиоза
Легоглобин –
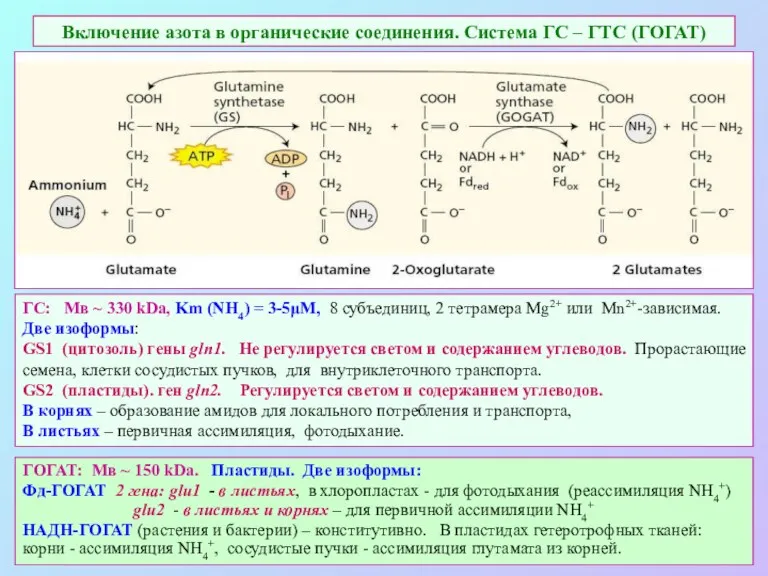
Включение азота в органические соединения. Система ГС – ГТС (ГОГАТ)
ГОГАТ:
Включение азота в органические соединения. Система ГС – ГТС (ГОГАТ)
ГОГАТ:
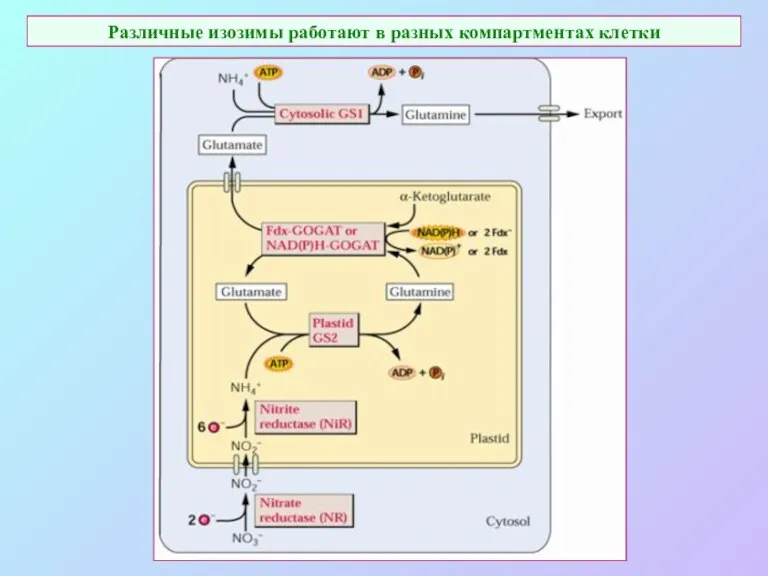
Различные изозимы работают в разных компартментах клетки
Различные изозимы работают в разных компартментах клетки
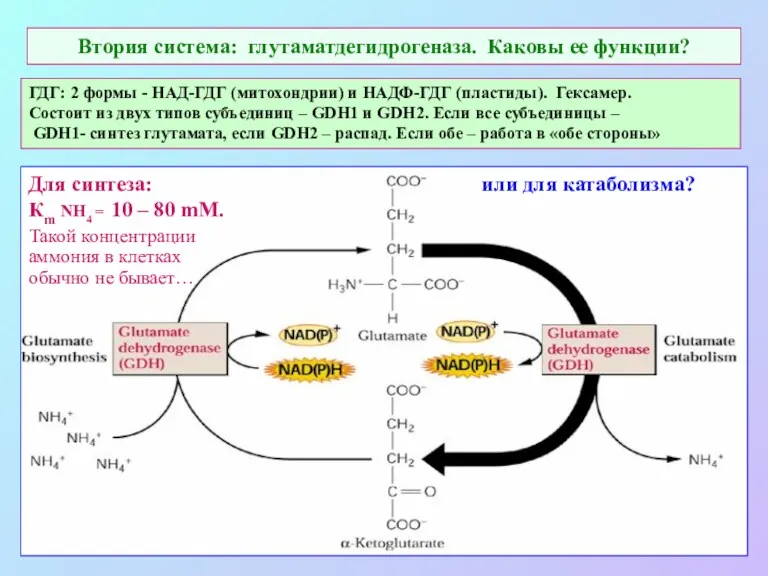
Втория система: глутаматдегидрогеназа. Каковы ее функции?
Для синтеза:
Кm NH4 = 10
Втория система: глутаматдегидрогеназа. Каковы ее функции?
Для синтеза:
Кm NH4 = 10
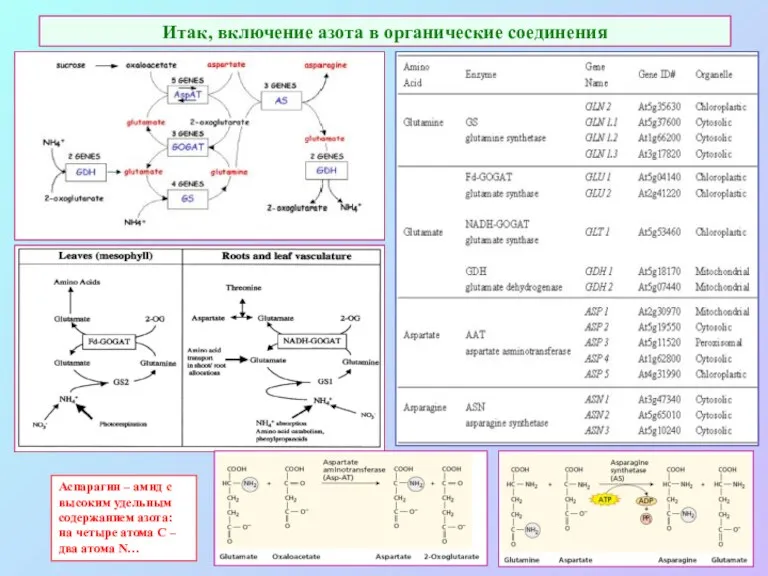
Итак, включение азота в органические соединения
Аспарагин – амид с высоким удельным
Итак, включение азота в органические соединения
Аспарагин – амид с высоким удельным
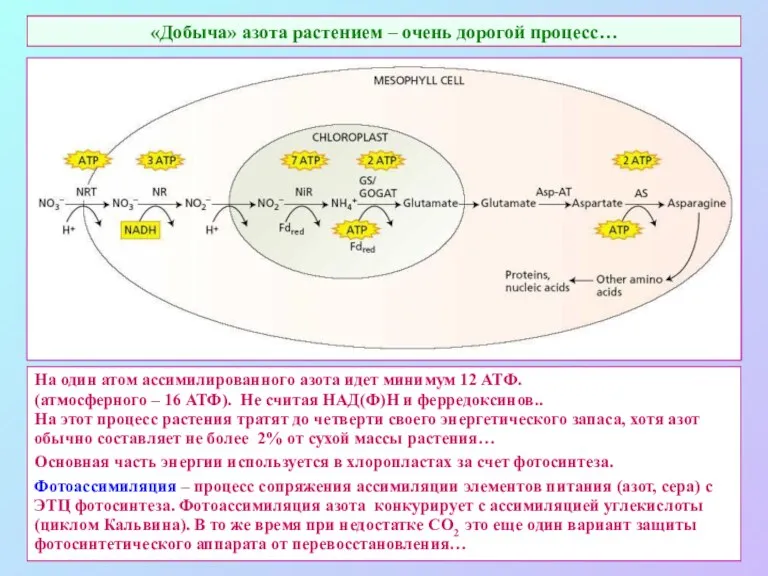
«Добыча» азота растением – очень дорогой процесс…
На один атом ассимилированного азота
«Добыча» азота растением – очень дорогой процесс…
На один атом ассимилированного азота
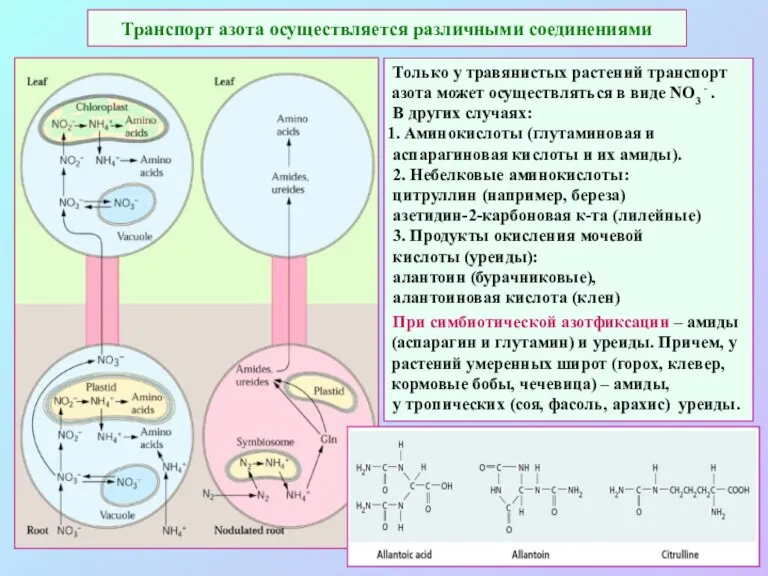
Транспорт азота осуществляется различными соединениями
Только у травянистых растений транспорт азота может
Транспорт азота осуществляется различными соединениями
Только у травянистых растений транспорт азота может
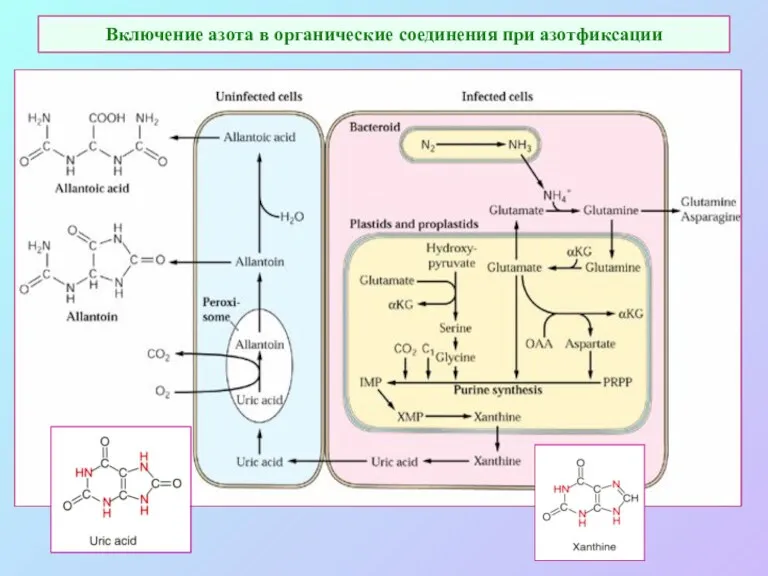
Включение азота в органические соединения при азотфиксации
Включение азота в органические соединения при азотфиксации
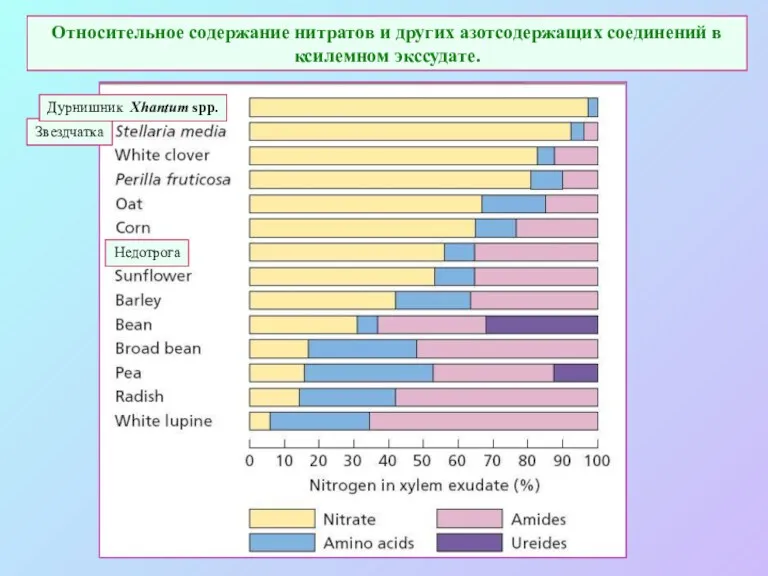
Относительное содержание нитратов и других азотсодержащих соединений в ксилемном экссудате.
Звездчатка
Дурнишник Xhantum
Относительное содержание нитратов и других азотсодержащих соединений в ксилемном экссудате.
Звездчатка
Дурнишник Xhantum
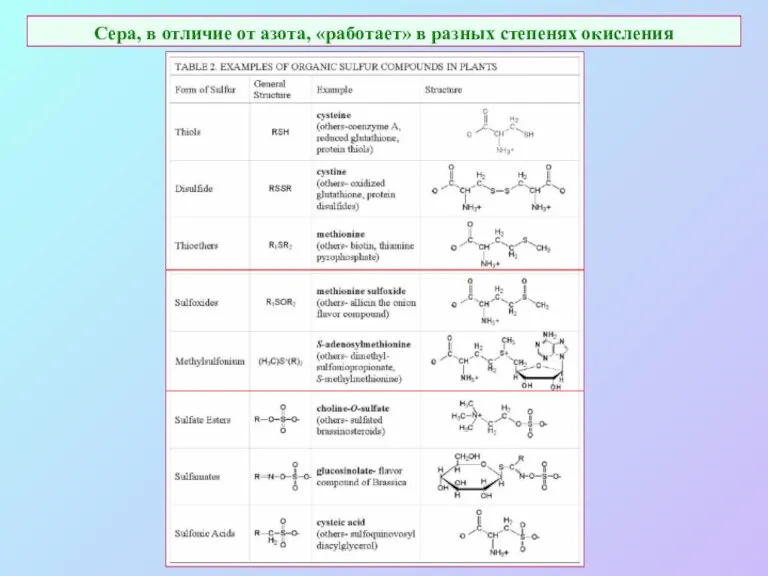
Сера, в отличие от азота, «работает» в разных степенях окисления
Сера, в отличие от азота, «работает» в разных степенях окисления
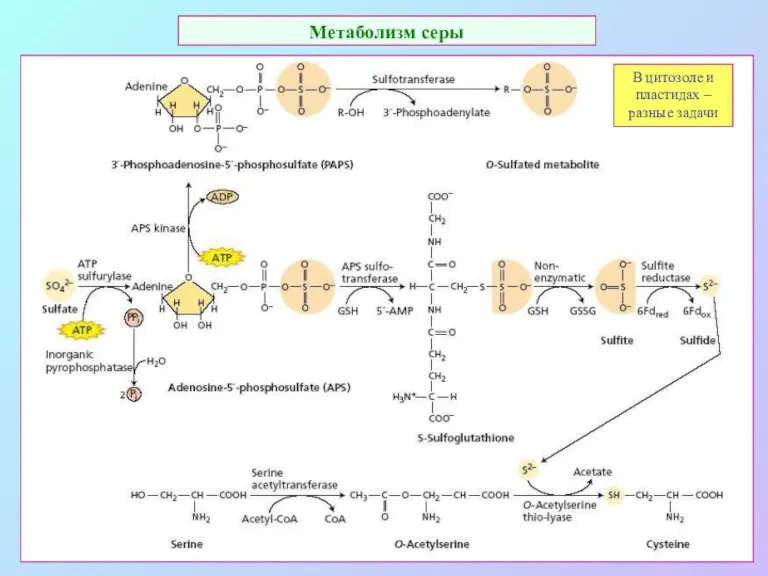
Метаболизм серы
В цитозоле и пластидах – разные задачи
Метаболизм серы
В цитозоле и пластидах – разные задачи
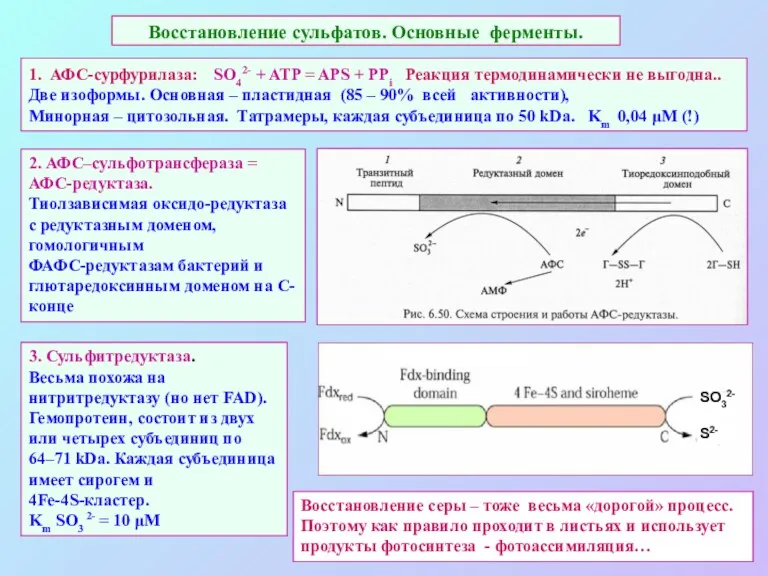
Восстановление сульфатов. Основные ферменты.
1. АФС-сурфурилаза: SO42- + ATP = APS
Восстановление сульфатов. Основные ферменты.
1. АФС-сурфурилаза: SO42- + ATP = APS
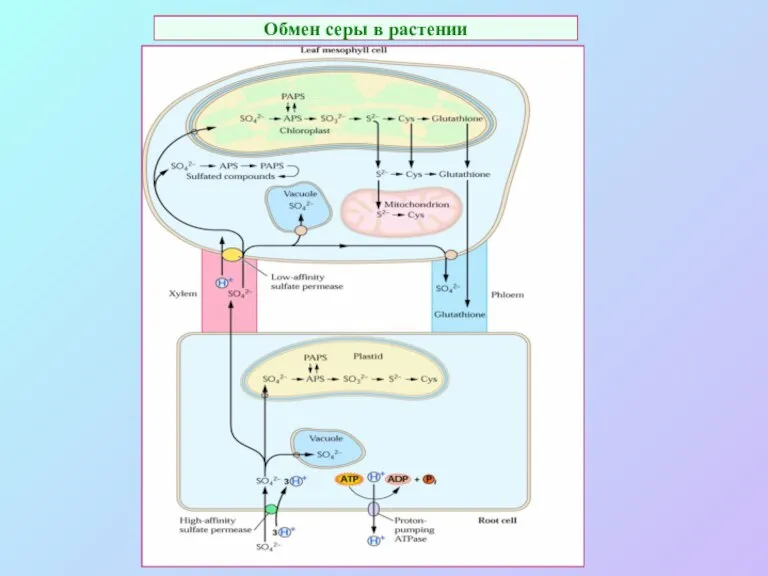
Обмен серы в растении
Обмен серы в растении
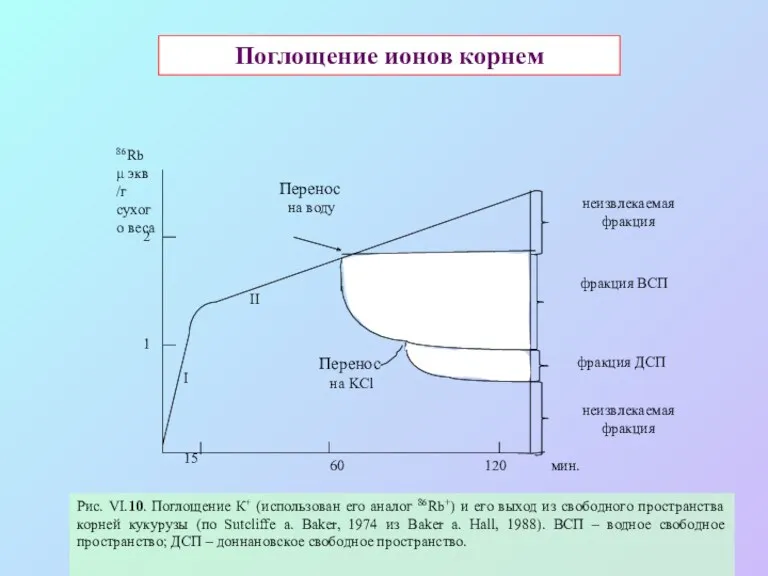
Поглощение ионов корнем
Поглощение ионов корнем
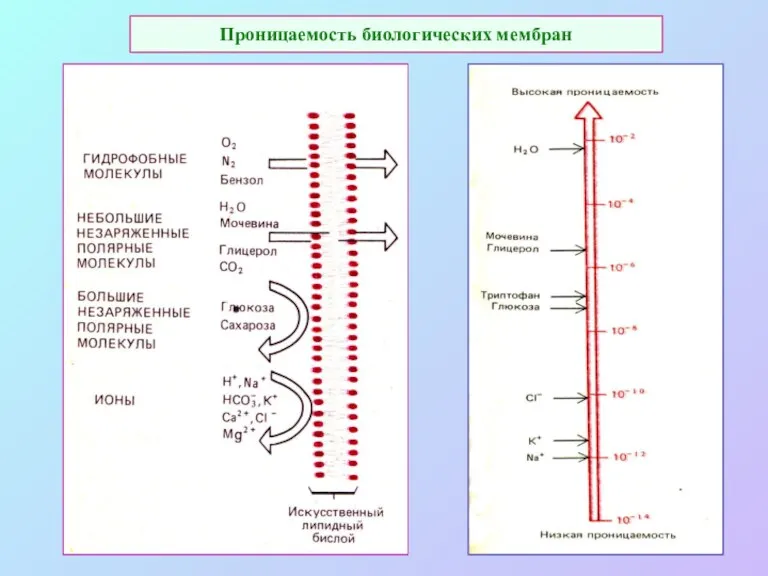
Проницаемость биологических мембран
Проницаемость биологических мембран

Поступление в клетку.
Перенос через плазмалемму.
Электрофизиологический подход
Кинетический подход
Молекулярно биологический подход
Поступление в клетку.
Перенос через плазмалемму.
Электрофизиологический подход
Кинетический подход
Молекулярно биологический подход

Электрофизиологический подход
Потенциал Нернста может быть использован, чтобы установить тип транспорта:
Электрофизиологический подход
Потенциал Нернста может быть использован, чтобы установить тип транспорта:
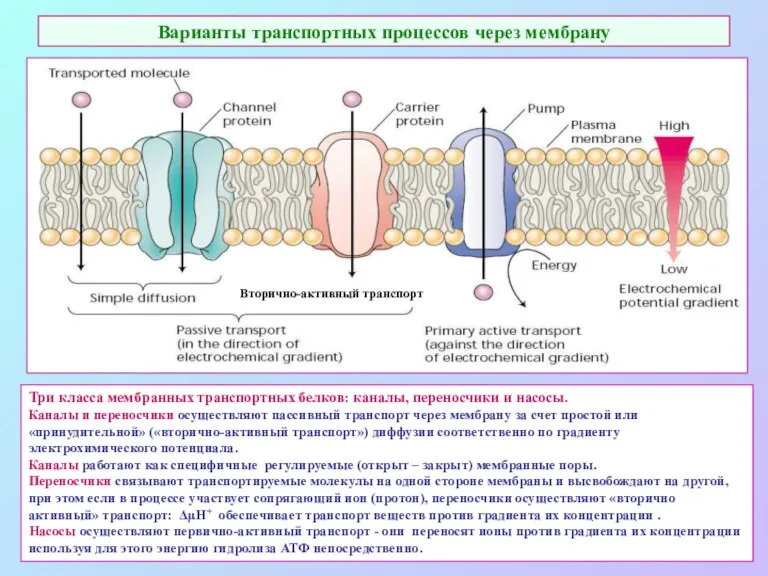
Варианты транспортных процессов через мембрану
Три класса мембранных транспортных белков: каналы, переносчики
Варианты транспортных процессов через мембрану
Три класса мембранных транспортных белков: каналы, переносчики
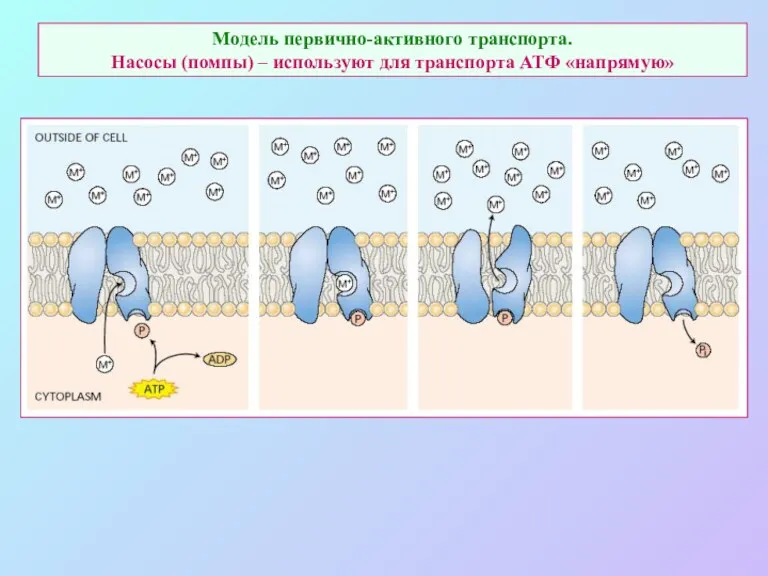
Модель первично-активного транспорта.
Насосы (помпы) – используют для транспорта АТФ «напрямую»
Модель первично-активного транспорта.
Насосы (помпы) – используют для транспорта АТФ «напрямую»
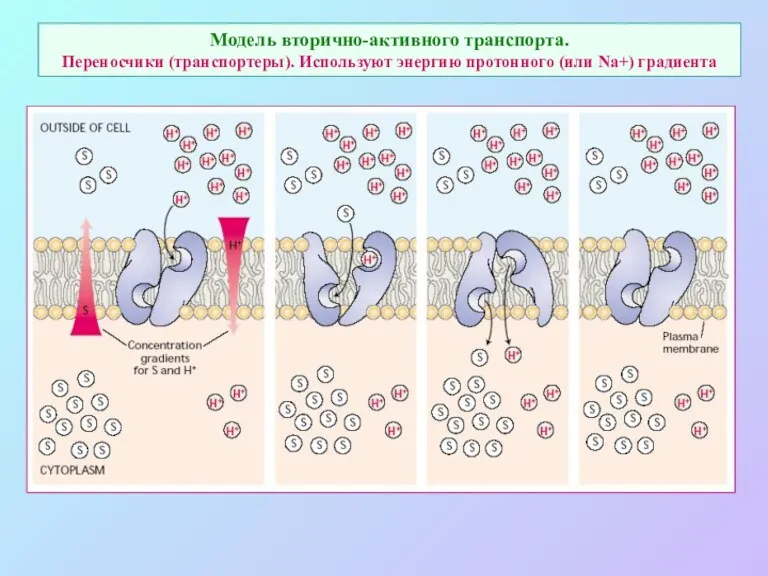
Модель вторично-активного транспорта.
Переносчики (транспортеры). Используют энергию протонного (или Na+) градиента
Модель вторично-активного транспорта.
Переносчики (транспортеры). Используют энергию протонного (или Na+) градиента
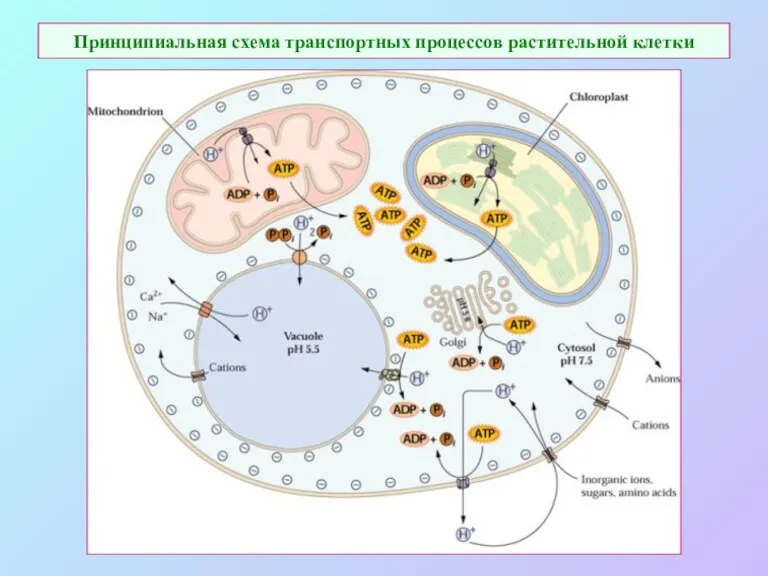
Принципиальная схема транспортных процессов растительной клетки
Принципиальная схема транспортных процессов растительной клетки
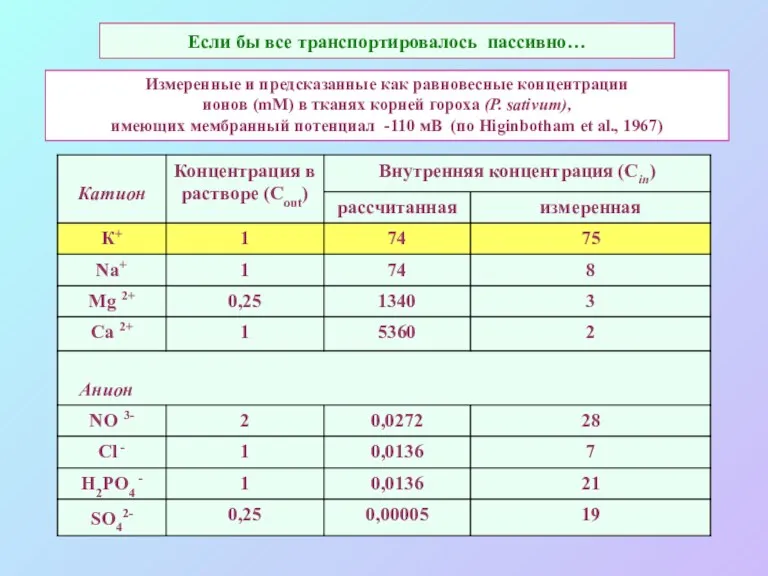
Если бы все транспортировалось пассивно…
Измеренные и предсказанные как равновесные концентрации
ионов
Если бы все транспортировалось пассивно…
Измеренные и предсказанные как равновесные концентрации
ионов
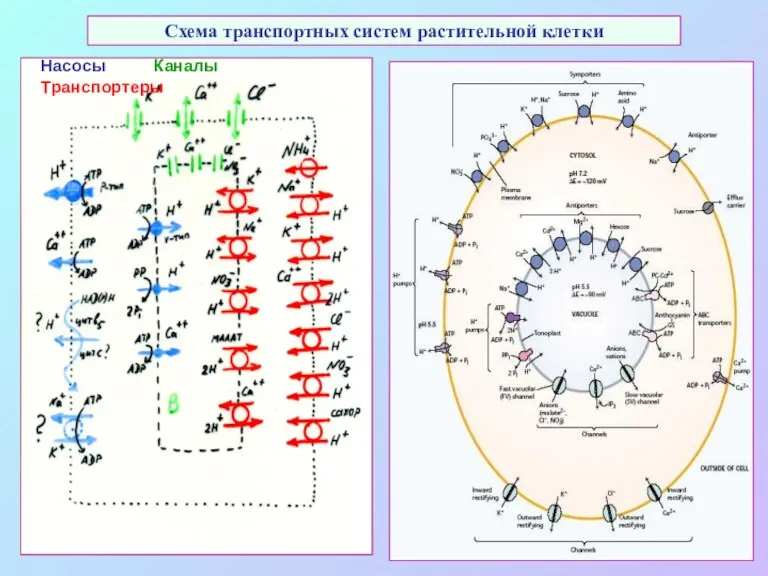
Схема транспортных систем растительной клетки
Насосы Каналы Транспортеры
Схема транспортных систем растительной клетки
Насосы Каналы Транспортеры
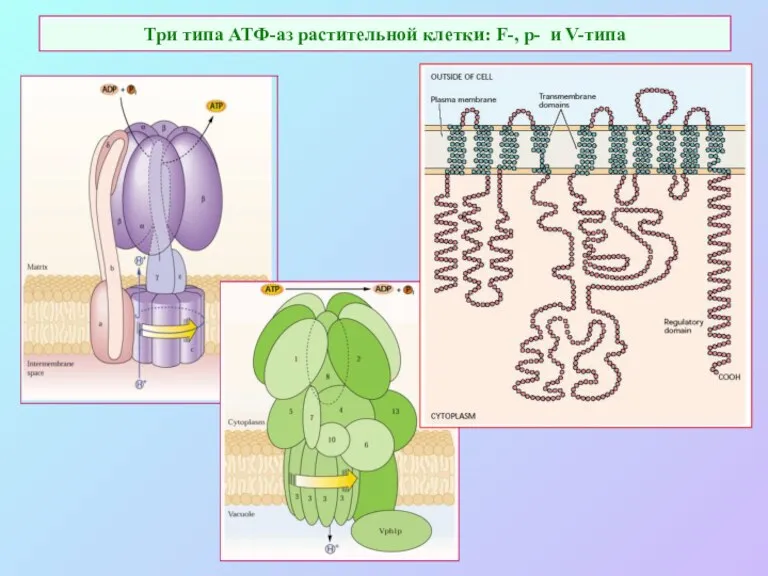
Три типа АТФ-аз растительной клетки: F-, p- и V-типа
Три типа АТФ-аз растительной клетки: F-, p- и V-типа
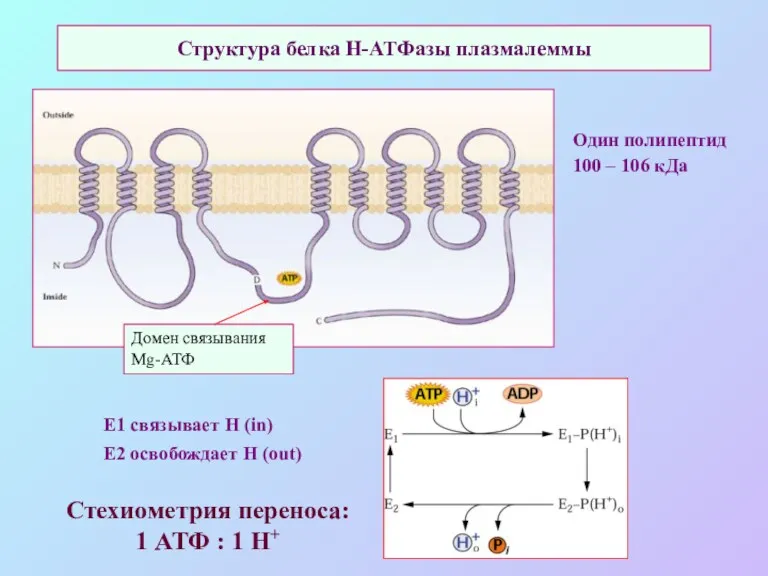
Структура белка Н-АТФазы плазмалеммы
Один полипептид
100 – 106 кДа
Домен связывания Mg-АТФ
Е1 связывает
Структура белка Н-АТФазы плазмалеммы
Один полипептид
100 – 106 кДа
Домен связывания Mg-АТФ
Е1 связывает
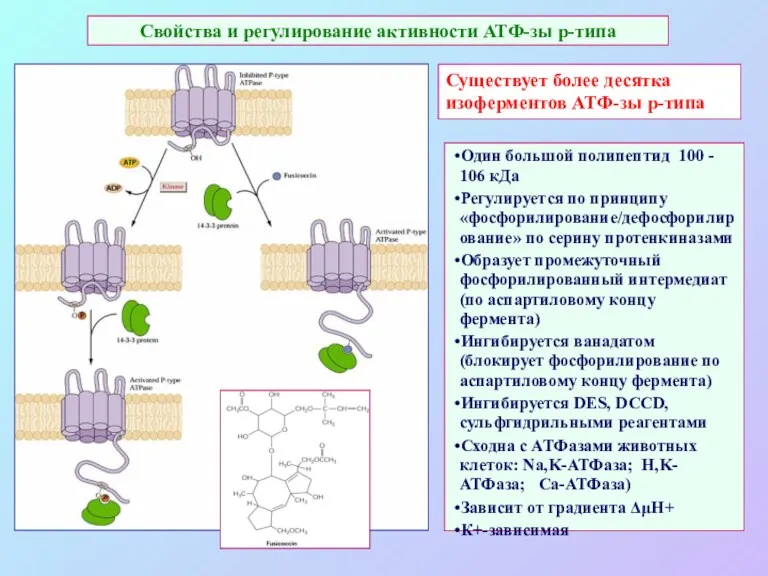
Свойства и регулирование активности АТФ-зы р-типа
Существует более десятка изоферментов АТФ-зы р-типа
Один
Свойства и регулирование активности АТФ-зы р-типа
Существует более десятка изоферментов АТФ-зы р-типа
Один
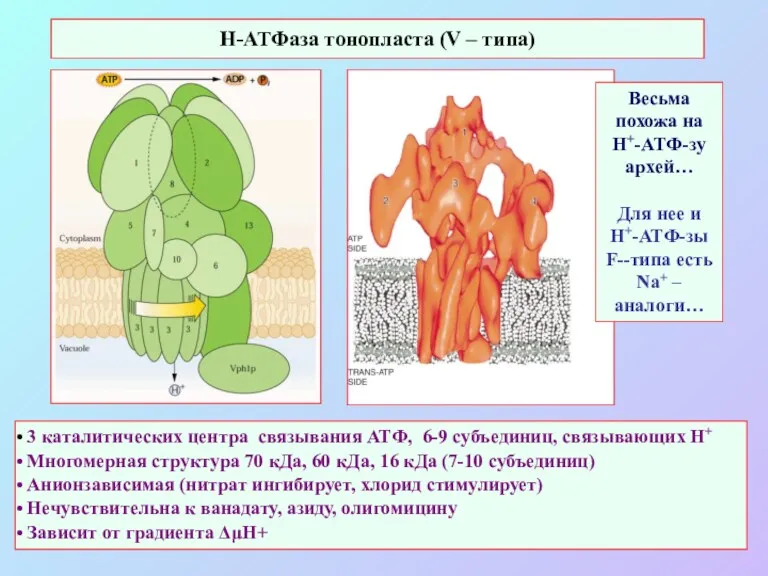
3 каталитических центра связывания АТФ, 6-9 субъединиц, связывающих Н+
Многомерная
3 каталитических центра связывания АТФ, 6-9 субъединиц, связывающих Н+
Многомерная
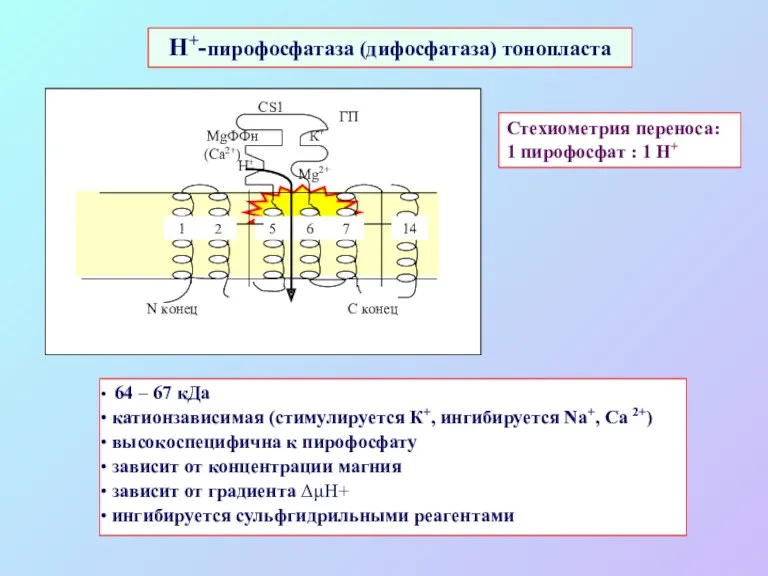
Стехиометрия переноса:
1 пирофосфат : 1 Н+
H+-пирофосфатаза (дифосфатаза) тонопласта
64 – 67
Стехиометрия переноса:
1 пирофосфат : 1 Н+
H+-пирофосфатаза (дифосфатаза) тонопласта
64 – 67
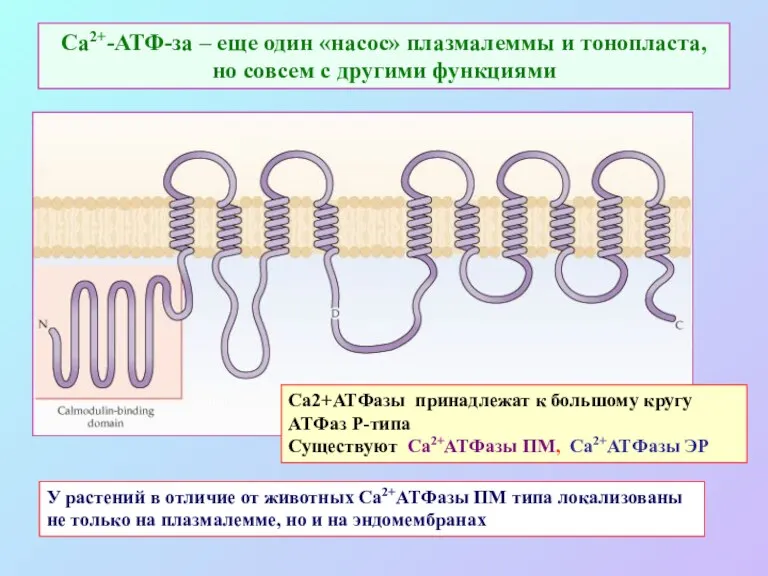
Са2+-АТФ-за – еще один «насос» плазмалеммы и тонопласта, но совсем с
Са2+-АТФ-за – еще один «насос» плазмалеммы и тонопласта, но совсем с
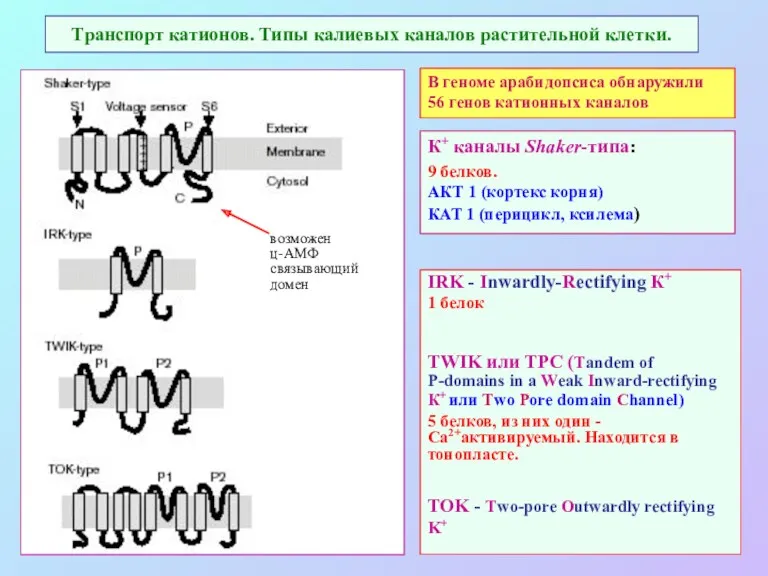
Транспорт катионов. Типы калиевых каналов растительной клетки.
IRK - Inwardly-Rectifying К+
1 белок
TWIK
Транспорт катионов. Типы калиевых каналов растительной клетки.
IRK - Inwardly-Rectifying К+
1 белок
TWIK
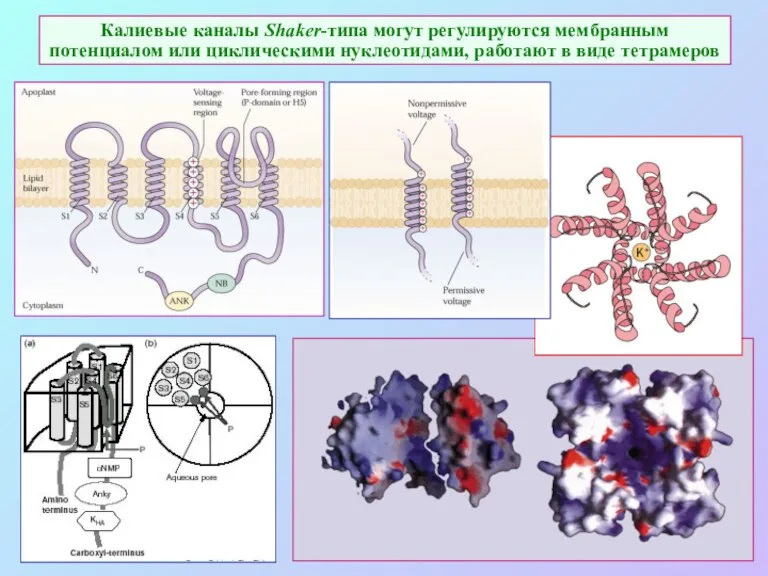
Калиевые каналы Shaker-типа могут регулируются мембранным потенциалом или циклическими нуклеотидами, работают
Калиевые каналы Shaker-типа могут регулируются мембранным потенциалом или циклическими нуклеотидами, работают

Транспорт калия через мембраны идет не только через каналы, но и
Транспорт калия через мембраны идет не только через каналы, но и
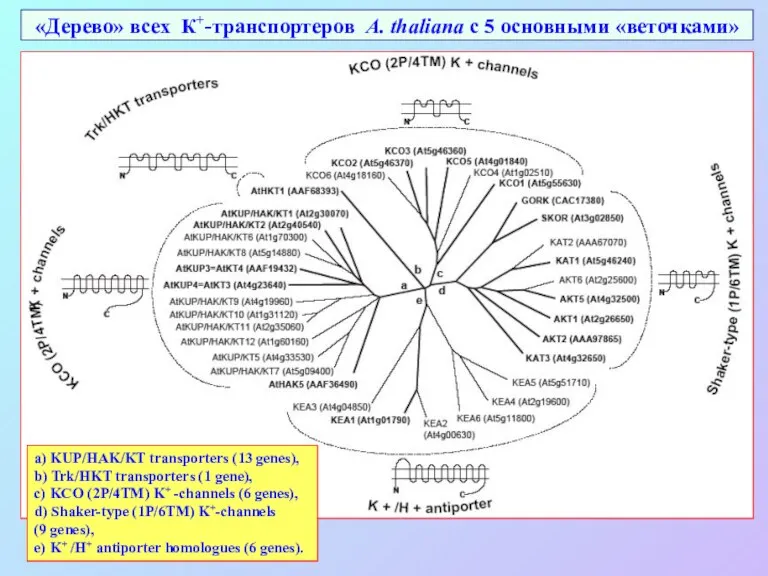
«Дерево» всех К+-транспортеров A. thaliana с 5 основными «веточками»
a) KUP/HAK/KT transporters
«Дерево» всех К+-транспортеров A. thaliana с 5 основными «веточками»
a) KUP/HAK/KT transporters
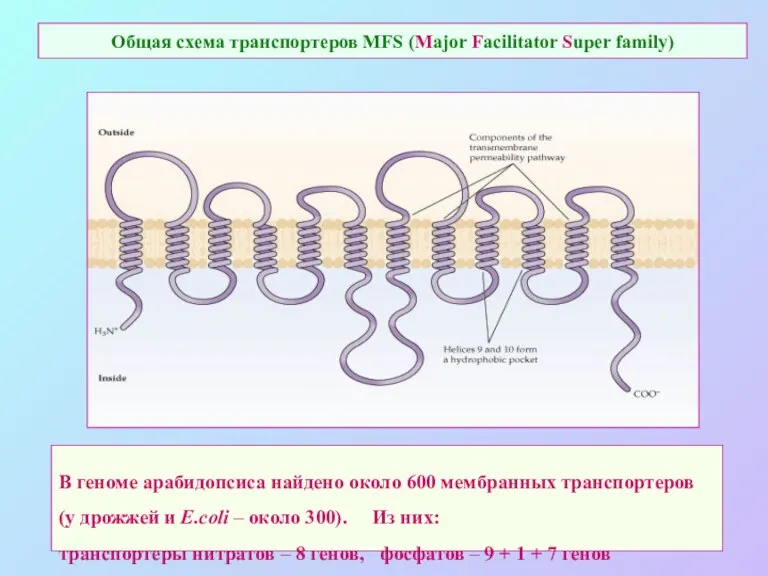
Общая схема транспортеров MFS (Major Facilitator Super family)
В геноме арабидопсиса найдено
Общая схема транспортеров MFS (Major Facilitator Super family)
В геноме арабидопсиса найдено

Сульфатные транспортеры
Высокого сродства Sultr1 или ST1 (Km порядка 10 мкмолей)
Низкого
Сульфатные транспортеры
Высокого сродства Sultr1 или ST1 (Km порядка 10 мкмолей)
Низкого
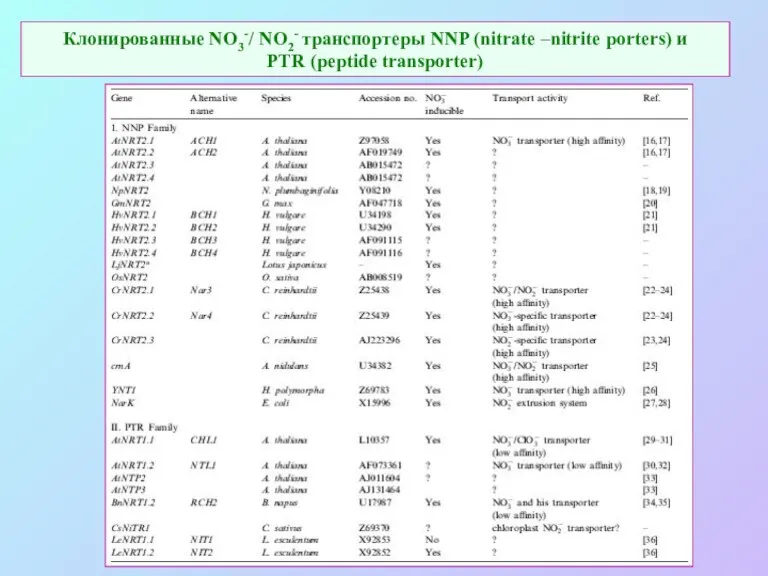
Клонированные NO3-/ NO2- транспортеры NNP (nitrate –nitrite porters) и
PTR (peptide
Клонированные NO3-/ NO2- транспортеры NNP (nitrate –nitrite porters) и PTR (peptide
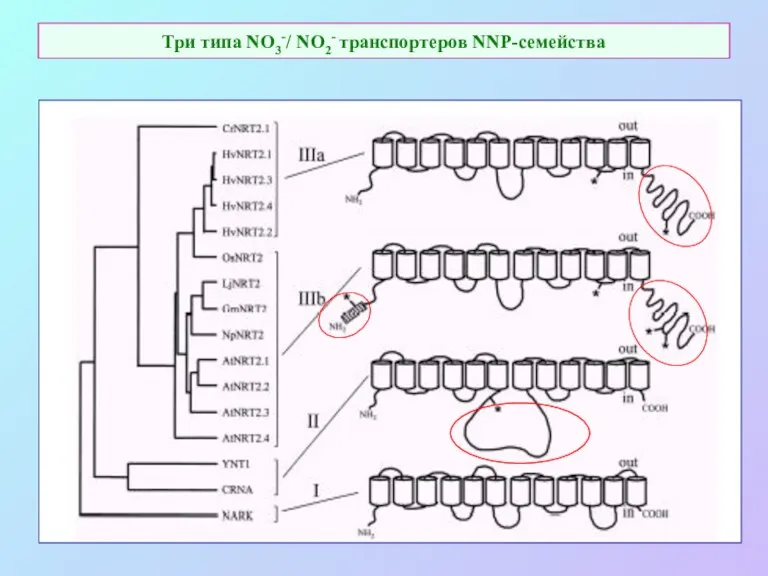
Три типа NO3-/ NO2- транспортеров NNP-семейства
Три типа NO3-/ NO2- транспортеров NNP-семейства
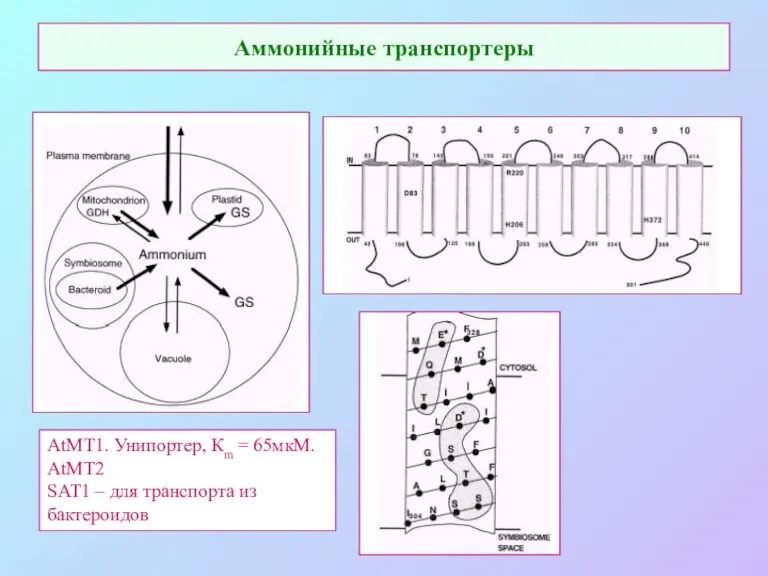
Аммонийные транспортеры
AtMT1. Унипортер, Кm = 65мкМ.
AtMT2
SAT1 – для транспорта из бактероидов
Аммонийные транспортеры
AtMT1. Унипортер, Кm = 65мкМ.
AtMT2
SAT1 – для транспорта из бактероидов
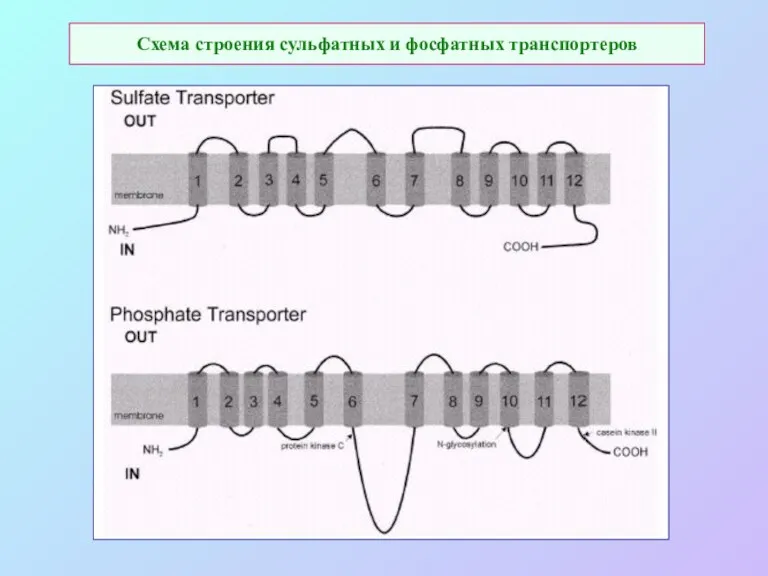
Схема строения сульфатных и фосфатных транспортеров
Схема строения сульфатных и фосфатных транспортеров
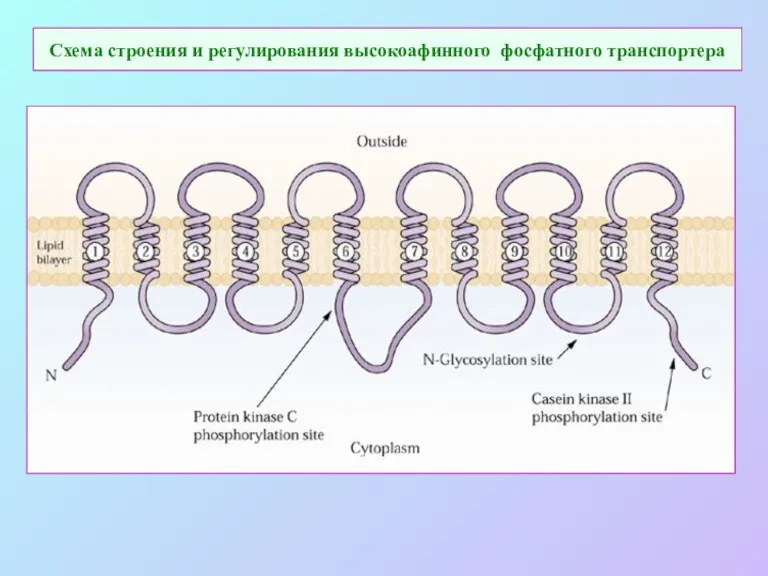
Схема строения и регулирования высокоафинного фосфатного транспортера
Схема строения и регулирования высокоафинного фосфатного транспортера
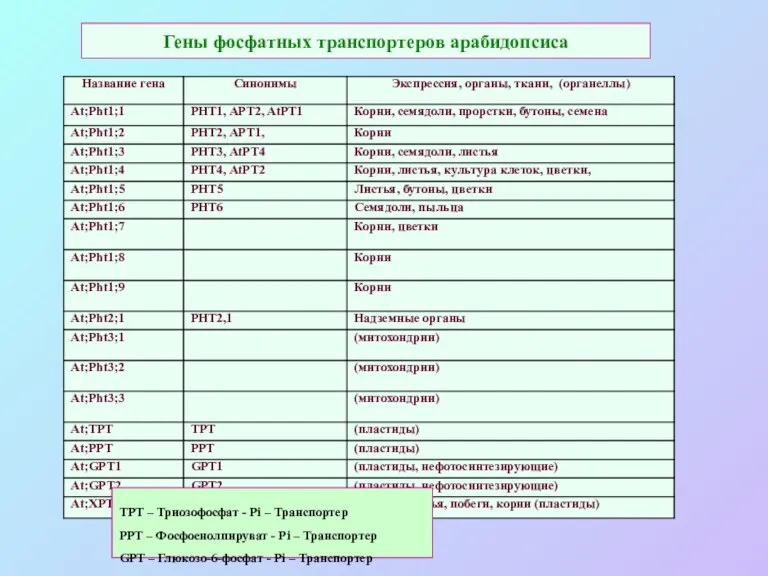
Гены фосфатных транспортеров арабидопсиса
TPT – Триозофосфат - Pi – Транспортер
РРТ –
Гены фосфатных транспортеров арабидопсиса
TPT – Триозофосфат - Pi – Транспортер
РРТ –
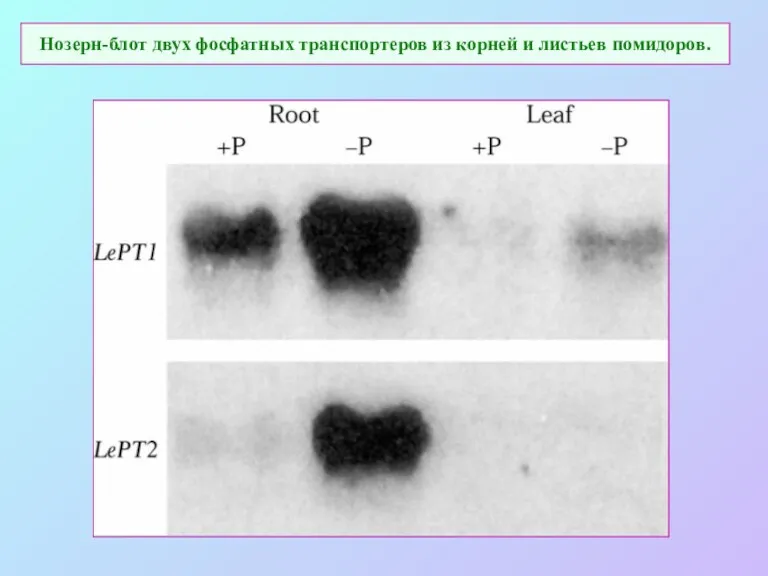
Нозерн-блот двух фосфатных транспортеров из корней и листьев помидоров.
Нозерн-блот двух фосфатных транспортеров из корней и листьев помидоров.
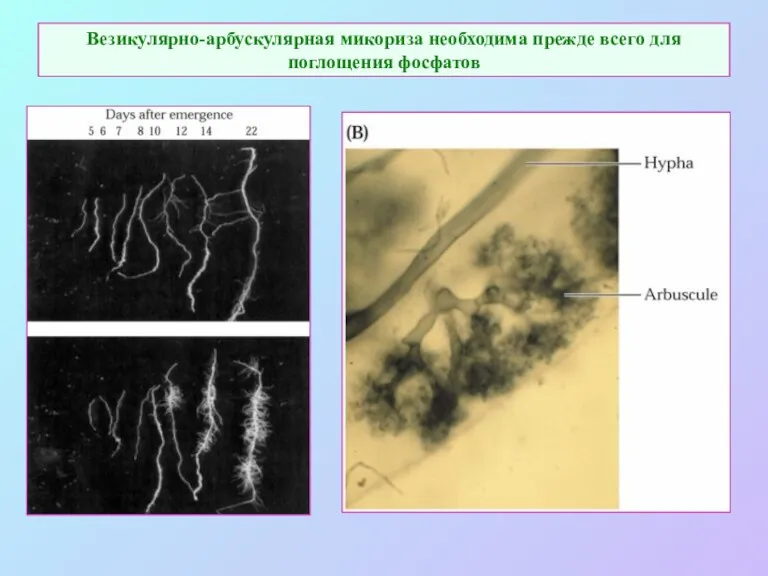
Везикулярно-арбускулярная микориза необходима прежде всего для поглощения фосфатов
Везикулярно-арбускулярная микориза необходима прежде всего для поглощения фосфатов
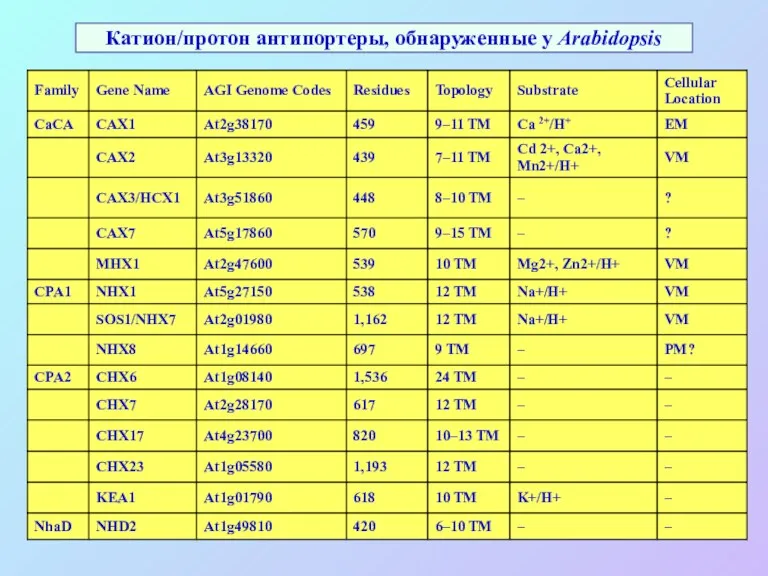
Катион/протон антипортеры, обнаруженные у Arabidopsis
Катион/протон антипортеры, обнаруженные у Arabidopsis
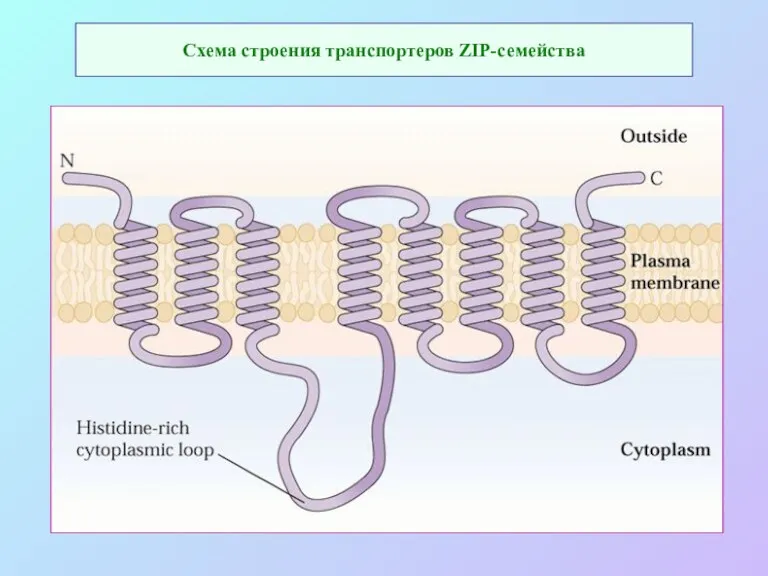
Схема строения транспортеров ZIP-семейства
Схема строения транспортеров ZIP-семейства
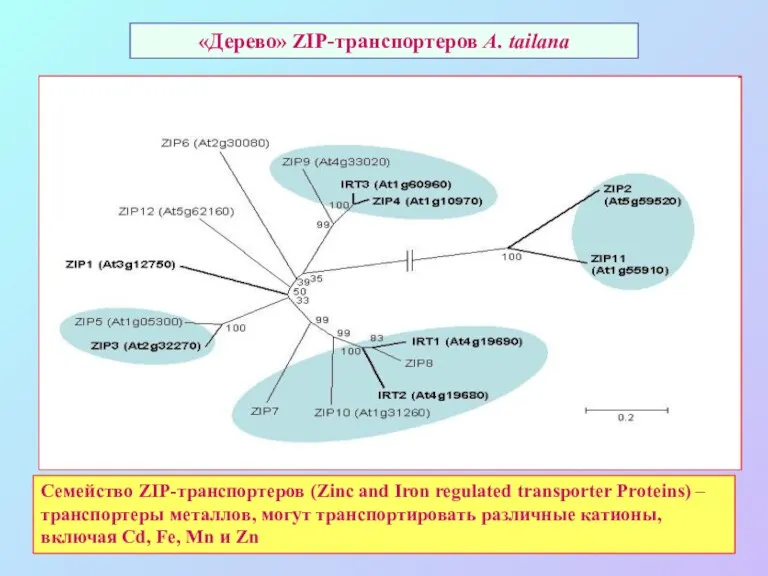
«Дерево» ZIP-транспортеров A. tailana
Семейство ZIP-транспортеров (Zinc and Iron regulated transporter Proteins)
«Дерево» ZIP-транспортеров A. tailana
Семейство ZIP-транспортеров (Zinc and Iron regulated transporter Proteins)

ABC-транспортеры
Транспортные белки, которые используют энергию гидролиза АТФ для транспорта через мембраны
ABC-транспортеры
Транспортные белки, которые используют энергию гидролиза АТФ для транспорта через мембраны
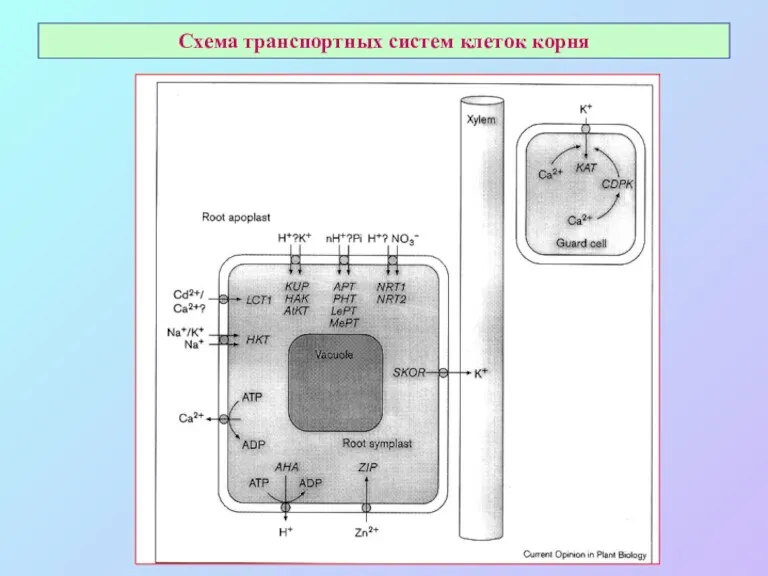
Схема транспортных систем клеток корня
Схема транспортных систем клеток корня
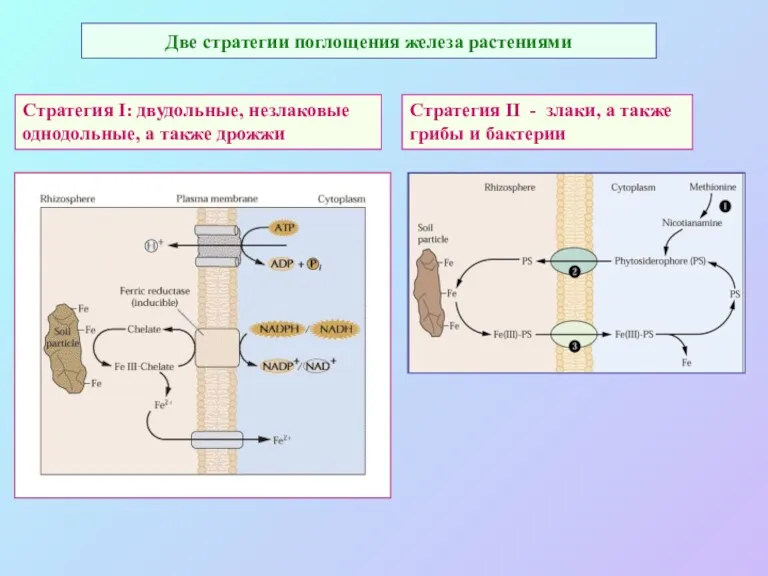
Две стратегии поглощения железа растениями
Стратегия I: двудольные, незлаковые
однодольные, а также дрожжи
Стратегия
Две стратегии поглощения железа растениями
Стратегия I: двудольные, незлаковые
однодольные, а также дрожжи
Стратегия
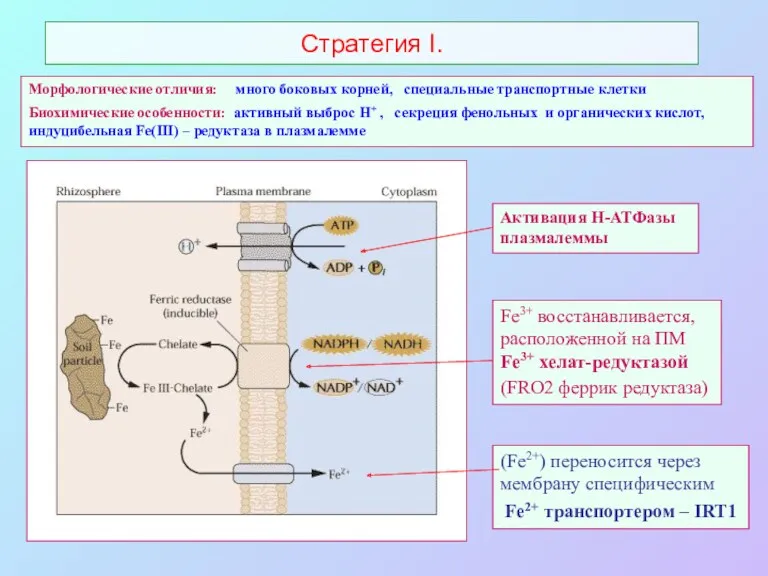
Стратегия I.
Fe3+ восстанавливается, расположенной на ПМ Fe3+ хелат-редуктазой
(FRO2 феррик редуктаза)
(Fe2+)
Стратегия I.
Fe3+ восстанавливается, расположенной на ПМ Fe3+ хелат-редуктазой
(FRO2 феррик редуктаза)
(Fe2+)
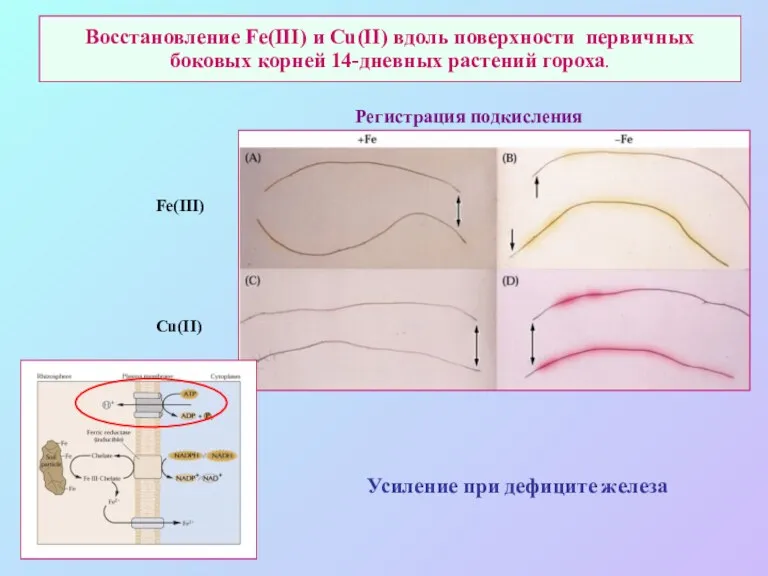
Восстановление Fe(III) и Cu(II) вдоль поверхности первичных боковых корней 14-дневных растений
Восстановление Fe(III) и Cu(II) вдоль поверхности первичных боковых корней 14-дневных растений
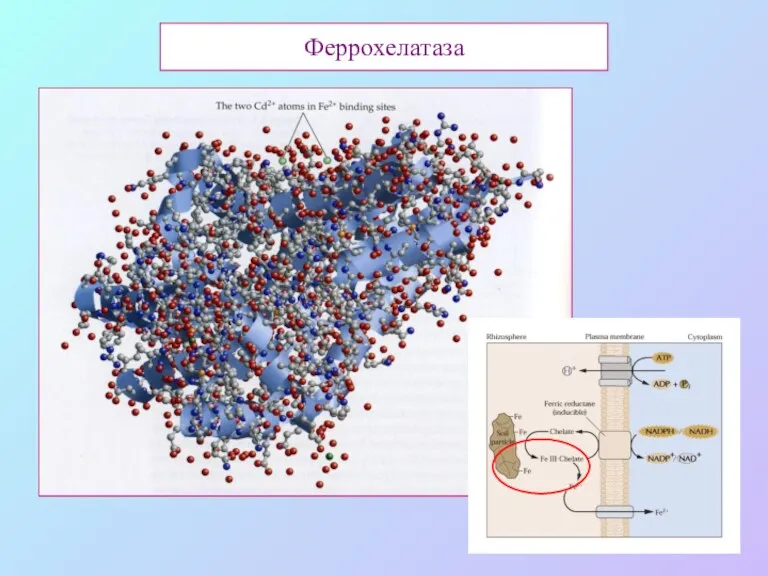
Феррохелатаза
Феррохелатаза
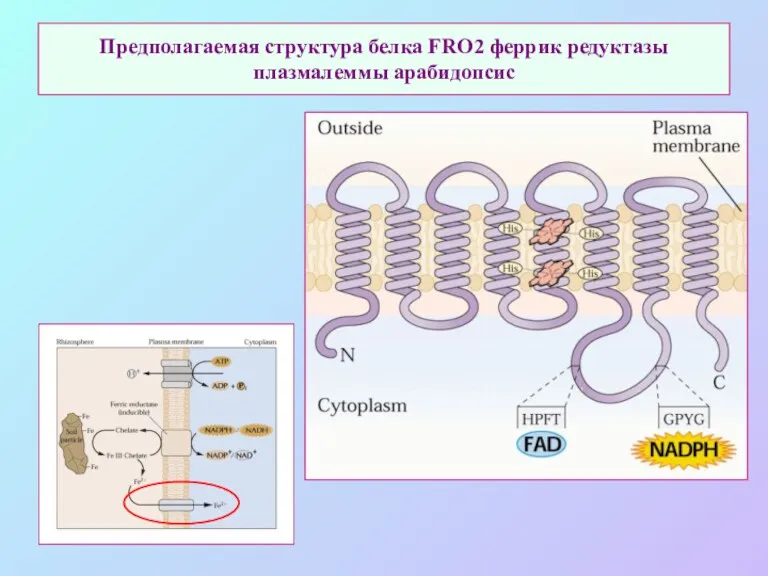
Предполагаемая структура белка FRO2 феррик редуктазы плазмалеммы арабидопсис
Предполагаемая структура белка FRO2 феррик редуктазы плазмалеммы арабидопсис
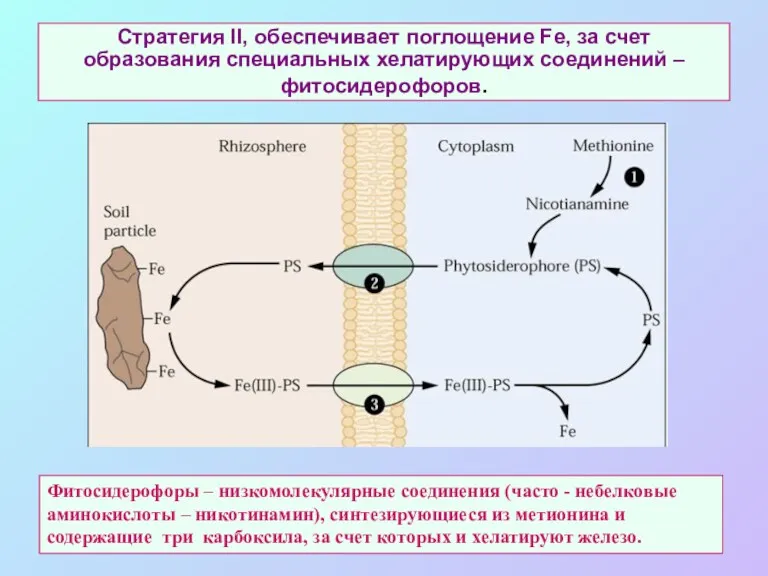
Стратегия II, обеспечивает поглощение Fe, за счет образования специальных хелатирующих соединений
Стратегия II, обеспечивает поглощение Fe, за счет образования специальных хелатирующих соединений
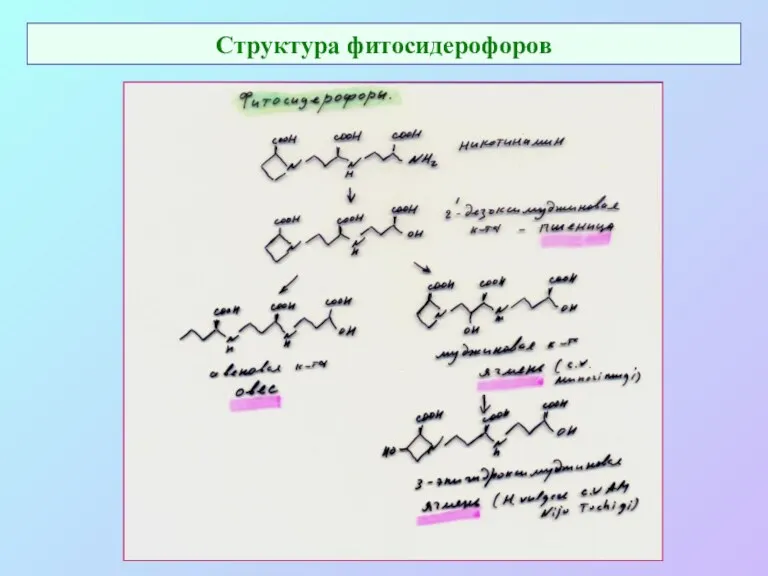
Структура фитосидерофоров
Структура фитосидерофоров
 Биологическое разнообразие грибов. Их практическое значение
Биологическое разнообразие грибов. Их практическое значение Положение человека в системе животного мира. Теория эволюции
Положение человека в системе животного мира. Теория эволюции Мышцы конечностей
Мышцы конечностей Эмбриональное развитие Drosophila. (Лекция 9)
Эмбриональное развитие Drosophila. (Лекция 9) Жасуша – тіршіліктің құрылымдық-функциялық бірлігі
Жасуша – тіршіліктің құрылымдық-функциялық бірлігі Морфофункциональная характеристика органов пищеварительной системы
Морфофункциональная характеристика органов пищеварительной системы Подцарство Одноклеточные животные (Простейшие)
Подцарство Одноклеточные животные (Простейшие) Составление тренировочных программ для мужчин различных типов сложения
Составление тренировочных программ для мужчин различных типов сложения Материальные основы наследственности
Материальные основы наследственности Як вчяться пташенята
Як вчяться пташенята Органы цветковых растений. Корень
Органы цветковых растений. Корень Полевые цветы
Полевые цветы Отрасли животноводства. Значение млекопитающих
Отрасли животноводства. Значение млекопитающих Ягоды и грибы (для дошкольников)
Ягоды и грибы (для дошкольников) Занимательный час зоологии. Презентация.
Занимательный час зоологии. Презентация. Прокариотические клетки
Прокариотические клетки Дерматозоонозы. Клещи
Дерматозоонозы. Клещи Разнообразие растений
Разнообразие растений Урок по теме Строение и значение цветка
Урок по теме Строение и значение цветка Строение сердца, сосудов
Строение сердца, сосудов Ингибиторы синтеза клеточной стенки
Ингибиторы синтеза клеточной стенки Кто живет в Африке
Кто живет в Африке Стволовые клетки
Стволовые клетки Бобовые культуры
Бобовые культуры Хижі рослини
Хижі рослини Лесничество. Проект
Лесничество. Проект Адаптационные особенности охраняемых животных северной территории Республики Коми
Адаптационные особенности охраняемых животных северной территории Республики Коми Рекордсмены животных
Рекордсмены животных