- Материальные основы наследственности

Содержание
- 2. Структура ДНК Первичная структура ДНК. Нуклеотиды соединяются ковалентной 5'-3' фосфодиэфирной связью. Вторичная структура ДНК. Цепи ДНК
- 3. Генетический код Генетический код обладает рядом особенностей. Во-первых, в коде отсутствуют «знаки препинания», то есть сигналы,
- 4. Возможность воспроизведения генов заложена в структуре ДНК – в комплементарности ее оснований – и заключается в
- 5. Ферменты репликации
- 6. Центральная догма молекулярной биологии (Ф. Крик, 1958) Линейная последовательность мономеров ДНК (дезоксирибонуклеотидов) определяет линейную последовательность мономеров
- 7. Структура и функции гена Ген в современном представлении это цистрон. Цистрон – это единица генетической функции,
- 8. Транскрипция Транскрипция – переписывания. Т.е. это процесс синтеза иРНК с ДНК. В результате транскрипции формируется проРНК
- 9. Трансляция (от лат. translatio — перевод) — процесс синтеза белка из аминокислот на матрице информационной (матричной)
- 10. Принципиальная схема реализации генетической информации у про- и эукариот. ПРОКАРИОТЫ. У прокариот синтез белка рибосомой (трансляция)
- 11. Регуляция белкового синтеза регуляция синтеза белка происходит на основе кибернетического принципа обратной связи, то есть у
- 12. Структура лактозного оперона E. coli (Тарасенко Екатерина Владимировна) P - промотор (место присоединения РНК-полимеразы) О -
- 13. Механизм регуляции лактозного оперона В присутствии лактозы белок-репрессор неактивен, он не может присоединиться к оператору и
- 14. Структура триптофанового оперона E. coli Механизм регуляции триптофанового оперона При недостатке триптофана оперон включен При избытке
- 15. Регуляция на стадии транскрипции у эукариот Способы регуляции синтеза белка у эукариот более разнообразны, сложны и
- 16. Процесс регуляции синтеза белка у эукариот с помощью белков-гистонов Гистоновые и негистоновые белки – антогонисты (противоположны
- 17. Организация генетического материала
- 18. Хромосома это структура, которая содержит нуклеиновую кислоту и функция которой состоит в хранении, реализации и передаче
- 19. Строение хромосом эукариот Основу хромосомы составляет линейная макромолекула ДНК значительной длины. Например, в молекулах ДНК хромосом
- 20. Схема упаковки хроматина Упаковка ДНК в хроматин обеспечивает многократное сокращение линейных размеров ДНК, необходимое для размещения
- 21. Нуклеосома В состав гистонового кора входит 4 типа гистонов - Н2А, Н2В, Н3, Н4. На гистоновый
- 22. Морфология метафазных хромосом В митозе хромосомы преобразуются в плотно упакованные элементы, способные сопротивляться внешним воздействиям, сохранять
- 23. В зависимости от функционального состояния хроматина различают: гетерохроматин (5) и эухроматин (6). Эухроматин — генетически активные,
- 25. Скачать презентацию

Структура ДНК
Первичная структура ДНК.
Нуклеотиды соединяются ковалентной 5'-3' фосфодиэфирной связью.
Вторичная структура ДНК.
Цепи
Структура ДНК
Первичная структура ДНК.
Нуклеотиды соединяются ковалентной 5'-3' фосфодиэфирной связью.
Вторичная структура ДНК.
Цепи

Генетический код
Генетический код обладает рядом особенностей. Во-первых, в коде отсутствуют «знаки
Генетический код
Генетический код обладает рядом особенностей. Во-первых, в коде отсутствуют «знаки
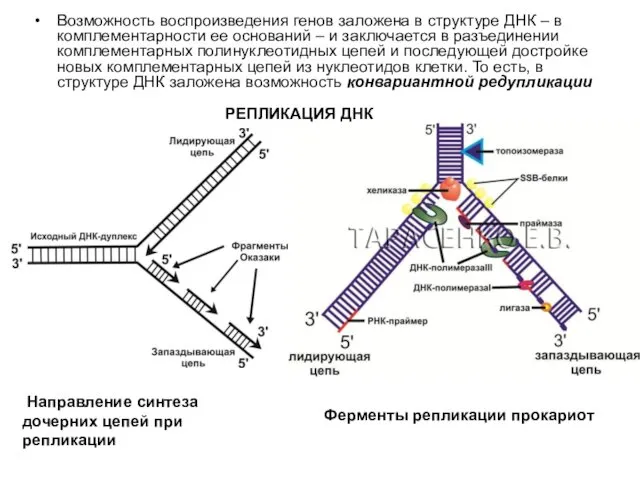
Возможность воспроизведения генов заложена в структуре ДНК – в комплементарности ее
Возможность воспроизведения генов заложена в структуре ДНК – в комплементарности ее

Ферменты репликации
Ферменты репликации

Центральная догма молекулярной биологии (Ф. Крик, 1958)
Линейная последовательность мономеров ДНК (дезоксирибонуклеотидов)
Центральная догма молекулярной биологии (Ф. Крик, 1958)
Линейная последовательность мономеров ДНК (дезоксирибонуклеотидов)
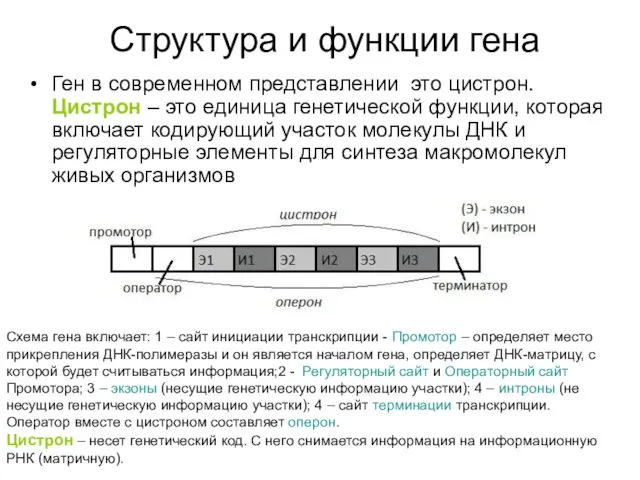
Структура и функции гена
Ген в современном представлении это цистрон. Цистрон –
Структура и функции гена
Ген в современном представлении это цистрон. Цистрон –

Транскрипция
Транскрипция – переписывания. Т.е. это процесс синтеза иРНК с ДНК.
В
Транскрипция
Транскрипция – переписывания. Т.е. это процесс синтеза иРНК с ДНК.
В

Трансляция
(от лат. translatio — перевод) — процесс синтеза белка из аминокислот на матрице информационной (матричной) РНК (иРНК, мРНК).
Общая схема трансляции.
Инициация.1. Узнавание
Трансляция
(от лат. translatio — перевод) — процесс синтеза белка из аминокислот на матрице информационной (матричной) РНК (иРНК, мРНК).
Общая схема трансляции. Инициация.1. Узнавание
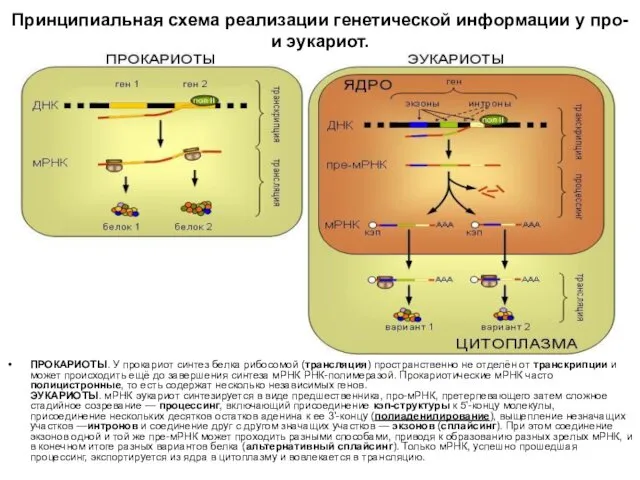
Принципиальная схема реализации генетической информации у про- и эукариот.
ПРОКАРИОТЫ. У прокариот синтез белка рибосомой (трансляция)
Принципиальная схема реализации генетической информации у про- и эукариот.
ПРОКАРИОТЫ. У прокариот синтез белка рибосомой (трансляция)

Регуляция белкового синтеза
регуляция синтеза белка происходит на основе кибернетического принципа обратной
Регуляция белкового синтеза
регуляция синтеза белка происходит на основе кибернетического принципа обратной

Структура лактозного оперона E. coli (Тарасенко Екатерина Владимировна)
P - промотор (место
Структура лактозного оперона E. coli (Тарасенко Екатерина Владимировна)
P - промотор (место

Механизм регуляции лактозного оперона
В присутствии лактозы белок-репрессор неактивен, он не
Механизм регуляции лактозного оперона
В присутствии лактозы белок-репрессор неактивен, он не

Структура триптофанового оперона E. coli
Механизм регуляции триптофанового оперона
При недостатке
Структура триптофанового оперона E. coli
Механизм регуляции триптофанового оперона
При недостатке

Регуляция на стадии транскрипции у эукариот
Способы регуляции синтеза белка у эукариот
Регуляция на стадии транскрипции у эукариот
Способы регуляции синтеза белка у эукариот
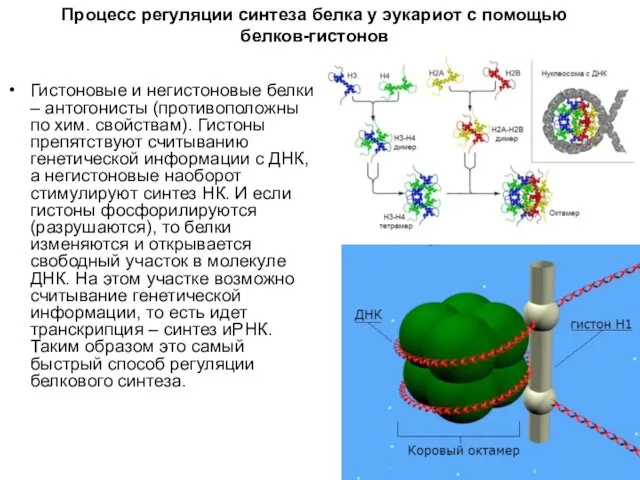
Процесс регуляции синтеза белка у эукариот с помощью белков-гистонов
Гистоновые и негистоновые
Процесс регуляции синтеза белка у эукариот с помощью белков-гистонов
Гистоновые и негистоновые

Организация генетического материала
Организация генетического материала
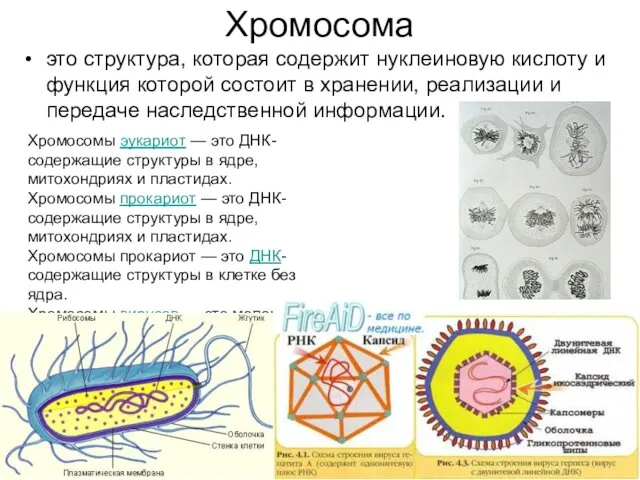
Хромосома
это структура, которая содержит нуклеиновую кислоту и функция которой состоит в хранении, реализации
Хромосома
это структура, которая содержит нуклеиновую кислоту и функция которой состоит в хранении, реализации

Строение хромосом эукариот
Основу хромосомы составляет линейная макромолекула ДНК значительной длины. Например,
Строение хромосом эукариот
Основу хромосомы составляет линейная макромолекула ДНК значительной длины. Например,

Схема упаковки хроматина
Упаковка ДНК в хроматин обеспечивает многократное сокращение линейных размеров ДНК,
Схема упаковки хроматина
Упаковка ДНК в хроматин обеспечивает многократное сокращение линейных размеров ДНК,
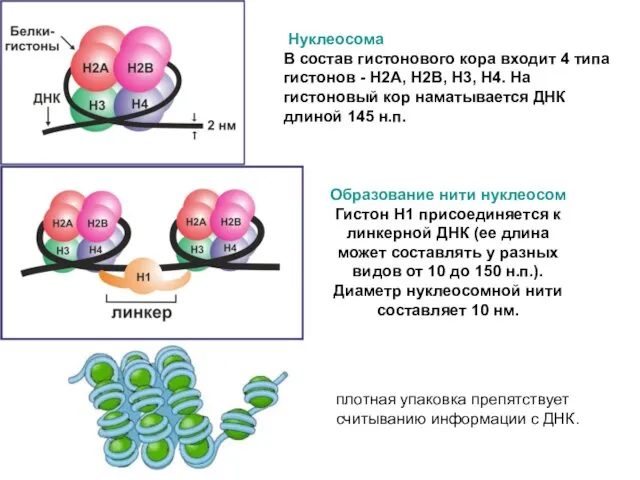
Нуклеосома
В состав гистонового кора входит 4 типа гистонов - Н2А, Н2В,
Нуклеосома
В состав гистонового кора входит 4 типа гистонов - Н2А, Н2В,

Морфология метафазных хромосом
В митозе хромосомы преобразуются в плотно упакованные элементы, способные
Морфология метафазных хромосом
В митозе хромосомы преобразуются в плотно упакованные элементы, способные

В зависимости от функционального состояния хроматина различают: гетерохроматин (5) и эухроматин (6).
Эухроматин — генетически
В зависимости от функционального состояния хроматина различают: гетерохроматин (5) и эухроматин (6).
Эухроматин — генетически
 Нормальная анатомия легких, плевры и средостения
Нормальная анатомия легких, плевры и средостения Черенкование комнатных растений
Черенкование комнатных растений Такие разные жуки. Коллекции. Занятие № 2. Консультация. Планирование деятельности
Такие разные жуки. Коллекции. Занятие № 2. Консультация. Планирование деятельности Введение в системную биологию
Введение в системную биологию Разнообразие животных
Разнообразие животных Болезни органов дыхания и их предупреждение. Гигиена дыхания
Болезни органов дыхания и их предупреждение. Гигиена дыхания Развитие низших хордовых. Амфибии
Развитие низших хордовых. Амфибии Специфика энергетических систем растительной клетки
Специфика энергетических систем растительной клетки Летучие мыши
Летучие мыши Анатомо-физиологические особенности сердечно сосудистой системы
Анатомо-физиологические особенности сердечно сосудистой системы Презентация у уроку Строение и функции пищеварительной системы человека 8 класс
Презентация у уроку Строение и функции пищеварительной системы человека 8 класс Вопросы экзамена по нервной системе
Вопросы экзамена по нервной системе Устройство речевого аппарата
Устройство речевого аппарата Жизнь и научная деятельность Ж.Б.Ламарка (1744-1829)
Жизнь и научная деятельность Ж.Б.Ламарка (1744-1829) Мышцы шеи
Мышцы шеи Половое размножение у покрытосеменных растений
Половое размножение у покрытосеменных растений Матричные биосинтезы
Матричные биосинтезы Мендель Грегор Иоганн
Мендель Грегор Иоганн Растения-рекордсмены
Растения-рекордсмены Выделительная система человека
Выделительная система человека Онтогенез. Лекция 14
Онтогенез. Лекция 14 Биологиялық тыңайтқыштар
Биологиялық тыңайтқыштар Все о грибах
Все о грибах Биологический метод исследования. Определение патогенности и вирулентности микроорганизмов
Биологический метод исследования. Определение патогенности и вирулентности микроорганизмов Научно-исследовательский проект Удивительные свойства сахара
Научно-исследовательский проект Удивительные свойства сахара Процеси гниття. Хімізм процесу гниття
Процеси гниття. Хімізм процесу гниття Ұлпаның және ағзаның электростимуляциясы
Ұлпаның және ағзаның электростимуляциясы Лабораторная диагностика стафилококковой инфекции
Лабораторная диагностика стафилококковой инфекции