- Репарация ДНК

Содержание
- 2. Типы повреждений ДНК. Классификация систем репарации ДНК. Фотореактивация. Эксцизионная репарация. Пострепликативная репарация. SOS-репарация. Репарация однонитевых и
- 3. Ежедневно у человека возникает около 50 тыс. однонитевых разрывов, более 8 тыс. окисленных и алкилированных оснований,
- 4. Повреждения ДНК Повреждения ДНК Репарация ДНК Генетическая стабильность Неполная репарация ДНК Генетическая нестабильность Рак, наследственные заболевания
- 5. Физические факторы – все виды радиации, ультрафиолетовый свет (УФ-свет), СВЧ, температура. Химические факторы – полициклические и
- 6. Для ДНК характерно: Наличие большого числа репарационных систем. В клетках имеются белки, специально «патрулирующие» ДНК и
- 7. 1. Изменение структуры азотистых оснований – алкилирование (чаще всего метилирование с образованием 7-метилгуанина, 1-метиладенина, 6-О-метилгуанина, а
- 8. Репарируемые и нерепарируемые. Спонтанные и индуцированные. Индуцируемые экзогенными факторами. Индуцируемые эндогенными факторами. Репарируемые повреждения удаляются собственными
- 9. Образование АП-сайта Образование АП-сайт ов 2. Спонтанные повреждения возникают без каких либо направленных воздействий, а индуцированные
- 10. К спонтанным повреждениям ДНК относится также дезаминирование азотистых оснований: цитозин урацил 3 Ежедневно в каждой клетке
- 11. Гуанин Цитозин О 6 -метилгуаинин Тимин метилирование и репликация 1.Алкилирование
- 12. Тимин Тимин Тимин гликоль При взаимодействии с активными формами кислорода и гидроперекисями образуется тимин, гидроксилированный по
- 13. Изменение структуры оснований: окисление Образование 8-оксо-2`-деоксигуанозина Образование 8-оксо-2`-деоксигуанозина (8-ОГ): Образуется в результате окисления гуанина Приводит к
- 14. Окисление может привести к разрыву колец оснований
- 15. 3. Гидролиз Образование АП-сайта
- 16. Образование димеров вызывается УФ-светом (УФ-С –длина волны менее 280 нм) и УФ-В (длина волны 280-320 нм)
- 18. 5. 5. Разрывы цепей одиночные (а) и двойные (б) (а) (б)
- 19. 5. 5. Разрывы цепей одиночные и двойные Возникновение двуцепочечного разрыва Начало репликации: Ku-антиген узнает двухцепочечные концы
- 20. 7. 6. Образование межнитевых сшивок ДНК Ковалентные связи между основаниями двух разных нитей ДНК Количество межнитевых
- 22. 8. 7. Образование ковалентных связей между основанием ДНК и белком Возникают при образовании топоизомеразных ковалентных связей
- 23. Образование тройной нити ДНК при взаимодействии с циануровой кислотой Циануровая кислота – дезинфектант, используемый в бассейнах,
- 24. Повреждения ДНК, вызываемые табачным дымом Наличие в дыме частиц (так называемых реактивных окислительных метаболитов), генерирующих окисление
- 25. Репарация ДНК
- 26. Прямая репарация ДНК. Фотореактивация. Эксцизионная репарация ДНК. Пострепликативная репарация ДНК. SOS-репарация. Репарация, склонная к ошибкам. Репарация
- 27. Схема репарации с участием контрпараллельной нити
- 28. У прокариотических организмов: Фотореактивация Эксцизионная репарация Пострепликативная репарация Репарация, склонная к ошибкам SOS-репарация У эукариотических организмов:
- 29. В репарации ДНКучаствуют более 150 генов . Нарушение репарации ДНК является одной из причин возникновения ряда
- 30. Прямая репарация ДНК
- 31. В минуту в клетке Е. coli может синтезироваться порядка 100 молекул метилтрансфераз. Примерами метилтрансфераз могут быть:
- 32. Фотореактивация открыта в 1948 И. Ф. Ковалевым (СССР), А. Келнером и Р. Дульбекко (США) в опытах
- 33. Восстановительный эффект при фотореактивации связан с действием фермента — фотолиазы (дезоксирибопиримидинфотолиазы), представляющей собой полипептид, ассоциированный с
- 34. За 1 минуту фотолиаза может расщепить 2,4 пиримидиновых димера. У Е. coli система фотореактивации удаляет до
- 35. Видимый свет Разрыв циклобутанового мостика Восстановление нативной структуры ДНК UV Фотореактивация Тиминовый димер 5` 3` TT
- 36. Репарация АР -сайтов Образовавшиеся в результате удаления пурина АР-сайты могут быть залечены ферментами инсертазами (от англ.
- 37. Эксцизионная репарация нуклеотидов Удаляет: химические аддукты; димеры пиримидинов. Для эксцизионной репарации необходима интактная неповрежденная комплементараная нить
- 38. Эксцизионная репарация В настоящее время известно два типа эксцизионной репарации: Эксцизия азотистых оснований с помощью специальных
- 39. Эксцизионная репарация представляет собой многоэтапный процесс и включает: ) «Узнавание» тиминового димера. ) Инцизию – надрезание
- 40. Таким способом репарируются следующие повреждения ДНК, включение: урацила вместо тимина; гипоксантина; формамидопиримидина; 5,6 тимина гидрата; 8-окси-гуанина;
- 41. Эксцизия азотистых оснований Удаляет специфические повреждения в азотистых основаниях ДНК. Основной фермент – гликозилаза. Имеется несколько
- 42. Пример действия ДНК-гликозилазы
- 43. UvrA recognizes bulky lesions 2. ДНК-хеликаза UvrD Белок uvrA-узнает тиминовый димер Тиминовый димер UvrB и uvrC
- 44. Некоторые наследственные заболевания человека связаны с дефектом эксцизионной репарации ДНК: Пигментная ксеродерма, Cиндром Кокэйна, триходистрофия и
- 45. Base excision repair pathway (BER). (a) A DNA glycosylase recognizes a damaged base and cleaves between
- 46. Известно 3 типа пигментной ксеродермы: XPI, XPII, XPvar, общими симптомами которой служит повышенная чувствительность к солнечному
- 47. Репарация ошибок репликации. Поскольку ошибки возникают на дочерней цепи, система репарации только на ней должна проводить
- 49. Репарация неспаренных оснований Перед репликацией ДНК находится в метилированной форме, вновь синтезированная цепь – неметилирована Упрощенная
- 50. Репарация неспаренных оснований связывается белок MutS, с которым затем связываются белки MutL и MutH. Образуется репарационный
- 51. Белок MutH разрезает неметилированную нить ДНК по сайту GATC, который может располагаться по любую сторону от
- 52. Затем ДНК-хеликаза II (MutU=UvrD) расплетает надрезанную нить ДНК между надрезом и неспаренным основанием (включая его) и
- 53. Экзонуклеаза I (если это 3'-конец) или экзонуклеаза VII (если это 5'-конец) удаляет вытесненную нить. Этот процесс
- 54. Затем образовавшаяся брешь застраивается ДНК-полимеразой III в присутствии SSB- белка. Наконец, ДНК-лигаза восстанавливает фосфодиэфирную связь.
- 55. Экзонуклеаза I – 3’-5’- активность Экзонуклеаза VII – 5’-3’- активность Хеликаза II = UvrD=MutU dam, mutH,
- 56. Пострепликативная репарация происходит когда в ДНК возникает так много повреждений, что в ходе эксцизионной репарации клетка
- 57. Пиримидиновый димер задерживает продвижение ДНК-полимеразы в ходе репликации. Она останавливается. Продолжение репликации происходит с участием фрагментов
- 58. Пострепликативная репарация Повреждение возникает в ДНК еще до начала репликации Участок, комплементарный поврежденному ДНК-полимераза обходит тиминовый
- 59. Основание в одной нити ДНК повреждено Происходит репликация молекулы ДНК Пробел перемещается в другую молекулу ДНК
- 60. Механизм пострепликативной репарации
- 61. Существование этой системы впервые постулировал М. Радман в 1974 г. Он же дал название этому механизму,
- 62. Наиболее изучена SOS-репарация у Е. coli, главными участниками которой являются белки, кодируемые генами Rec A и
- 64. Начало SOS- ответа определяется взаимодействием белка RecA с белком репрессором LexA. Ответ клетки на повреждающее воздействие
- 65. Одной из функций белка RecA является включение генов umuD и umuC, которые способны замедлять процесс синтеза
- 66. Система рестрикции-модификации Система рестрикции-модификации — ферментативная система бактерий, разрушающая попавшую в клетку чужеродную ДНК. Основная еѐ
- 67. Системы рестрикции-модификации были открыты в результате изучения молекулярных механизмов явления, называемого «ограничение, контролируемое хозяином» (англ. host-controlled
- 68. В 1978 году В. Арбер, Д. Натанс и Х. Смит были удостоены Нобелевской премии «За обнаружение
- 69. Система рестрикции-модификации специфична по отношению к определѐнным последовательностям нуклеотидов в ДНК, называемых сайтах рестрикции. Эффективность действия
- 71. Скачать презентацию

Типы повреждений ДНК.
Классификация систем репарации ДНК.
Фотореактивация.
Эксцизионная репарация.
Пострепликативная репарация.
SOS-репарация.
Репарация однонитевых и двухнитевых
Типы повреждений ДНК.
Классификация систем репарации ДНК.
Фотореактивация.
Эксцизионная репарация.
Пострепликативная репарация.
SOS-репарация.
Репарация однонитевых и двухнитевых

Ежедневно у человека возникает около 50 тыс. однонитевых разрывов, более 8
Ежедневно у человека возникает около 50 тыс. однонитевых разрывов, более 8
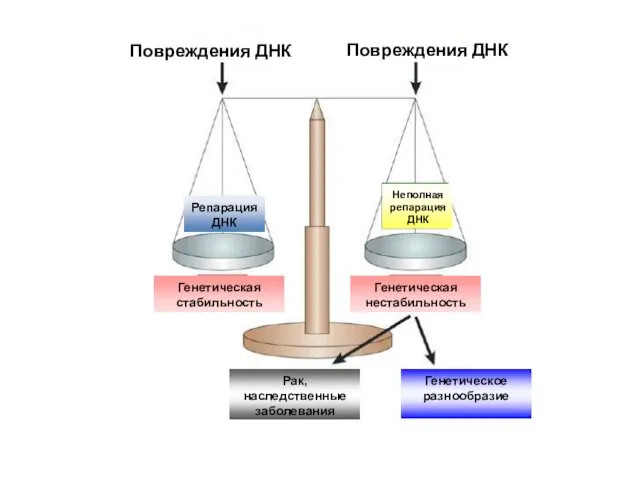
Повреждения ДНК
Повреждения ДНК
Репарация ДНК
Генетическая стабильность
Неполная репарация ДНК
Генетическая нестабильность
Рак, наследственные заболевания
Генетическое разнообразие
Повреждения ДНК
Повреждения ДНК
Репарация ДНК
Генетическая стабильность
Неполная репарация ДНК
Генетическая нестабильность
Рак, наследственные заболевания
Генетическое разнообразие

Физические факторы – все виды радиации, ультрафиолетовый свет (УФ-свет), СВЧ, температура.
Химические
Физические факторы – все виды радиации, ультрафиолетовый свет (УФ-свет), СВЧ, температура.
Химические

Для ДНК характерно:
Наличие большого числа репарационных систем.
В клетках имеются белки,
Для ДНК характерно:
Наличие большого числа репарационных систем.
В клетках имеются белки,

1. Изменение структуры азотистых оснований – алкилирование (чаще всего метилирование с
1. Изменение структуры азотистых оснований – алкилирование (чаще всего метилирование с

Репарируемые и нерепарируемые.
Спонтанные и индуцированные.
Индуцируемые экзогенными факторами.
Индуцируемые
Репарируемые и нерепарируемые.
Спонтанные и индуцированные.
Индуцируемые экзогенными факторами.
Индуцируемые
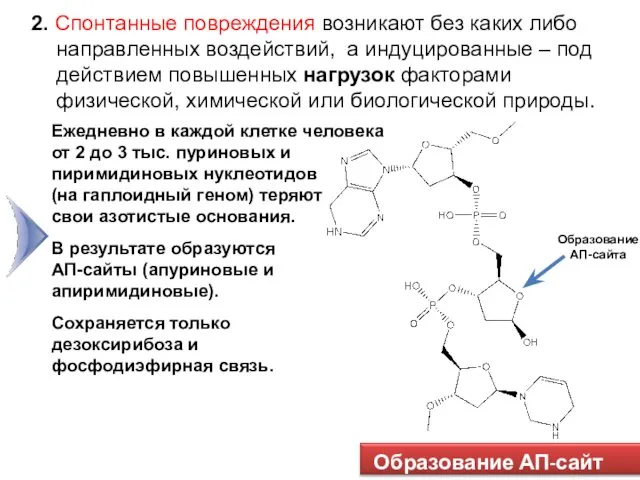
Образование АП-сайта
Образование АП-сайт ов
2. Спонтанные повреждения возникают без каких либо направленных
Образование АП-сайта
Образование АП-сайт ов
2. Спонтанные повреждения возникают без каких либо направленных
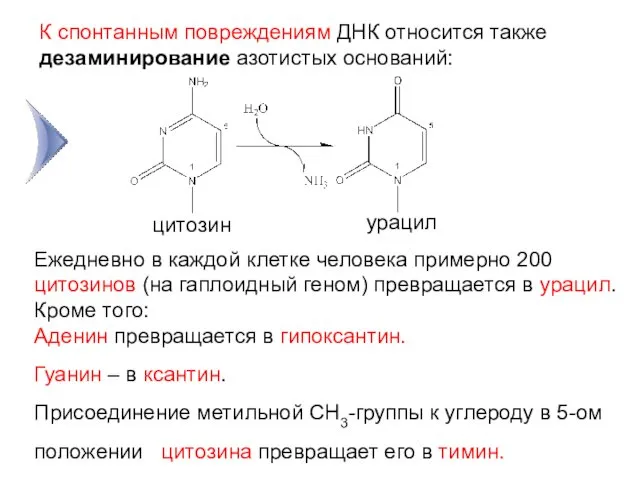
К спонтанным повреждениям ДНК относится также дезаминирование азотистых оснований:
цитозин
урацил
3
Ежедневно в каждой
К спонтанным повреждениям ДНК относится также дезаминирование азотистых оснований:
цитозин
урацил
3
Ежедневно в каждой
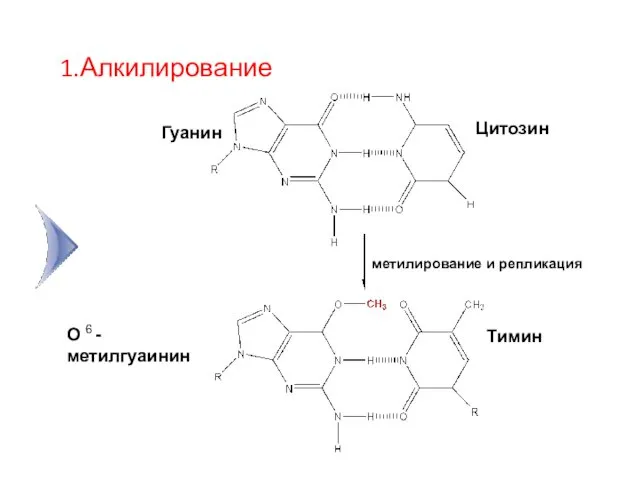
Гуанин
Цитозин
О 6 -метилгуаинин
Тимин
метилирование и репликация
1.Алкилирование
Гуанин
Цитозин
О 6 -метилгуаинин
Тимин
метилирование и репликация
1.Алкилирование
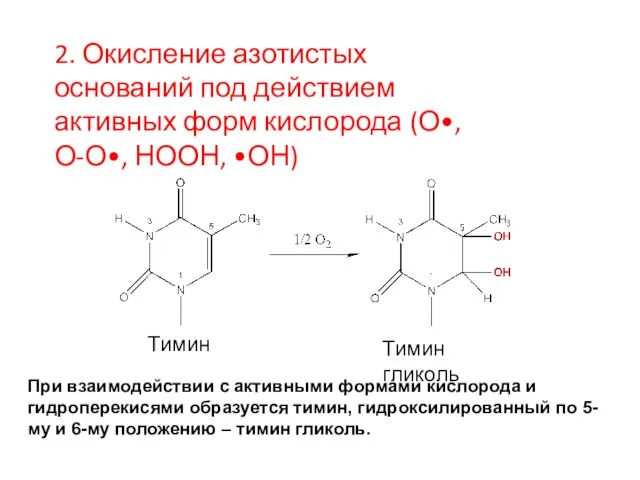
Тимин
Тимин
Тимин гликоль
При взаимодействии с активными формами кислорода и гидроперекисями образуется тимин,
Тимин
Тимин
Тимин гликоль
При взаимодействии с активными формами кислорода и гидроперекисями образуется тимин,
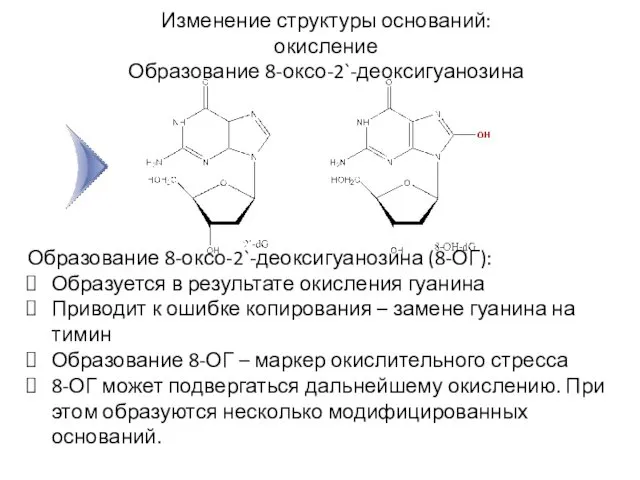
Изменение структуры оснований: окисление
Образование 8-оксо-2`-деоксигуанозина
Образование 8-оксо-2`-деоксигуанозина (8-ОГ):
Образуется в результате окисления гуанина
Приводит
Изменение структуры оснований: окисление
Образование 8-оксо-2`-деоксигуанозина
Образование 8-оксо-2`-деоксигуанозина (8-ОГ):
Образуется в результате окисления гуанина
Приводит
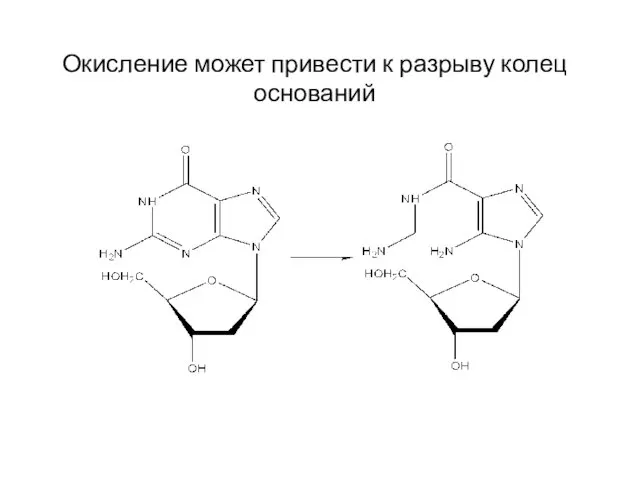
Окисление может привести к разрыву колец оснований
Окисление может привести к разрыву колец оснований
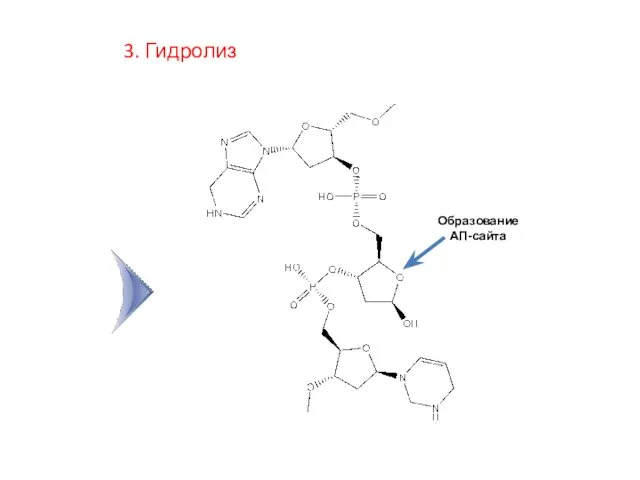
3. Гидролиз
Образование АП-сайта
3. Гидролиз
Образование АП-сайта
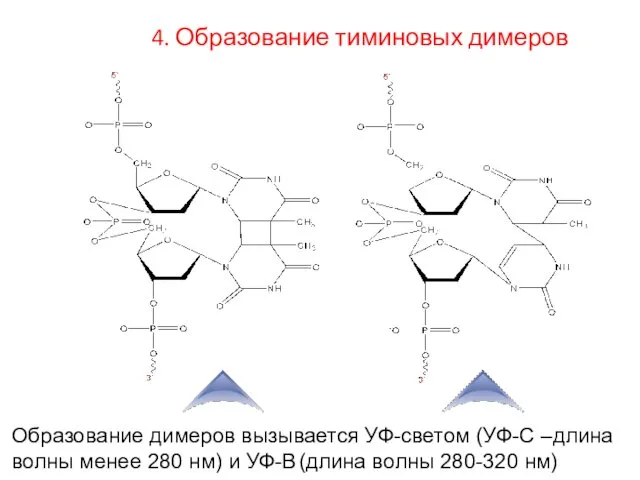
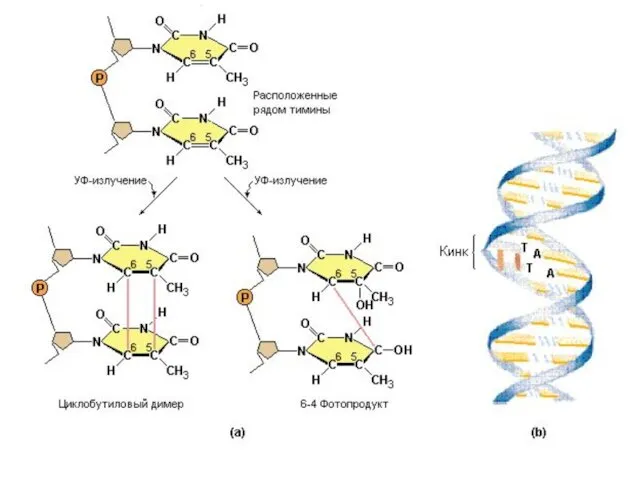
Образование димеров вызывается УФ-светом (УФ-С –длина волны менее 280 нм) и
Образование димеров вызывается УФ-светом (УФ-С –длина волны менее 280 нм) и
5.
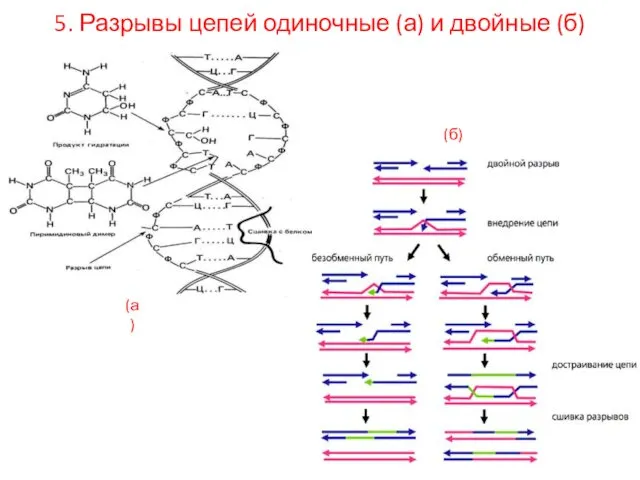
5. Разрывы цепей одиночные (а) и двойные (б)
(а)
(б)
5.
5. Разрывы цепей одиночные (а) и двойные (б)
(а)
(б)
5.
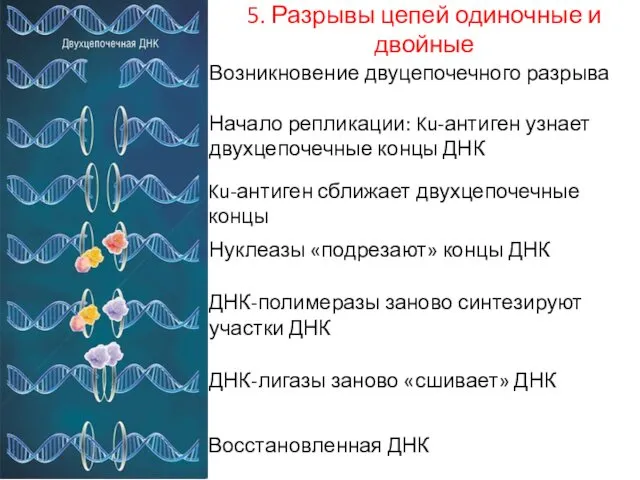
5. Разрывы цепей одиночные и двойные
Возникновение двуцепочечного разрыва
Начало репликации: Ku-антиген узнает
5.
5. Разрывы цепей одиночные и двойные
Возникновение двуцепочечного разрыва
Начало репликации: Ku-антиген узнает
7.
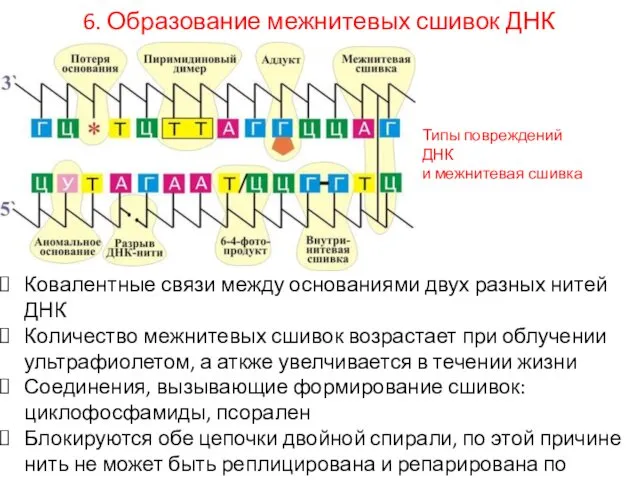
6. Образование межнитевых сшивок ДНК
Ковалентные связи между основаниями двух разных нитей
7.
6. Образование межнитевых сшивок ДНК
Ковалентные связи между основаниями двух разных нитей

8.
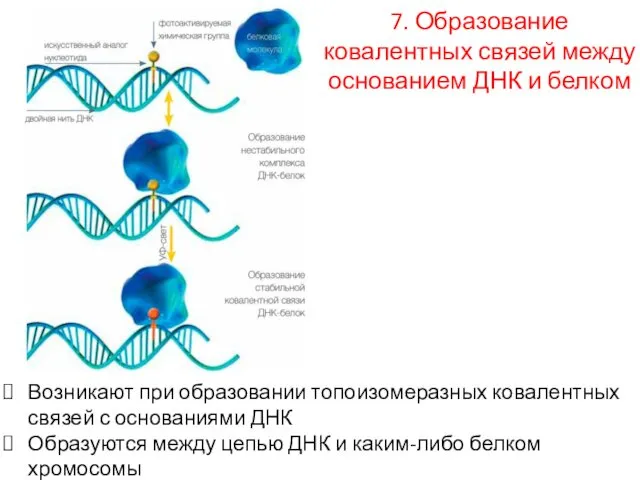
7. Образование ковалентных связей между основанием ДНК и белком
Возникают при образовании
8.
7. Образование ковалентных связей между основанием ДНК и белком
Возникают при образовании
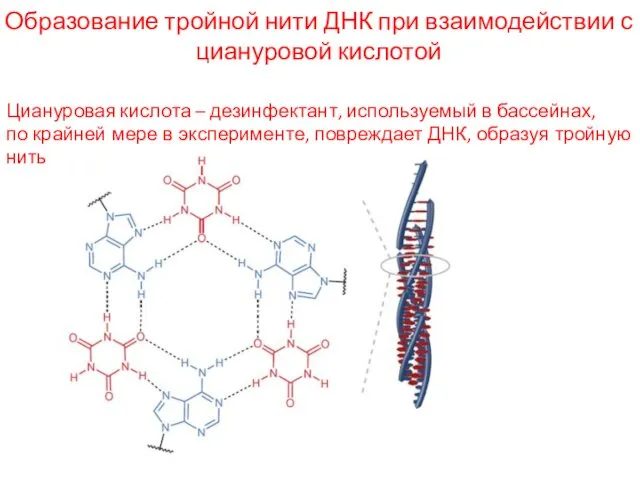
Образование тройной нити ДНК при взаимодействии с циануровой кислотой
Циануровая кислота –
Образование тройной нити ДНК при взаимодействии с циануровой кислотой
Циануровая кислота –

Повреждения ДНК, вызываемые табачным дымом
Наличие в дыме частиц (так называемых реактивных окислительных
Повреждения ДНК, вызываемые табачным дымом
Наличие в дыме частиц (так называемых реактивных окислительных

Репарация ДНК
Репарация ДНК

Прямая репарация ДНК.
Фотореактивация.
Эксцизионная репарация ДНК.
Пострепликативная репарация ДНК.
SOS-репарация.
Репарация, склонная к ошибкам.
Репарация
Прямая репарация ДНК.
Фотореактивация.
Эксцизионная репарация ДНК.
Пострепликативная репарация ДНК.
SOS-репарация.
Репарация, склонная к ошибкам.
Репарация
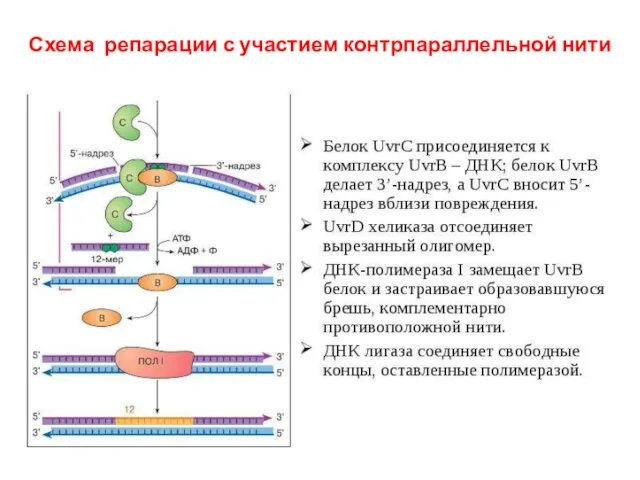
Схема репарации с участием контрпараллельной нити
Схема репарации с участием контрпараллельной нити

У прокариотических организмов:
Фотореактивация
Эксцизионная репарация
Пострепликативная репарация
Репарация, склонная к ошибкам
SOS-репарация
У эукариотических организмов:
Эксцизионная репарация
– Пострепликативная репарация
– Репарация, склонная
У прокариотических организмов:
Фотореактивация
Эксцизионная репарация
Пострепликативная репарация
Репарация, склонная к ошибкам
SOS-репарация
У эукариотических организмов:
Эксцизионная репарация
– Пострепликативная репарация
– Репарация, склонная

В репарации ДНКучаствуют более 150 генов .
Нарушение репарации ДНК является одной
В репарации ДНКучаствуют более 150 генов .
Нарушение репарации ДНК является одной
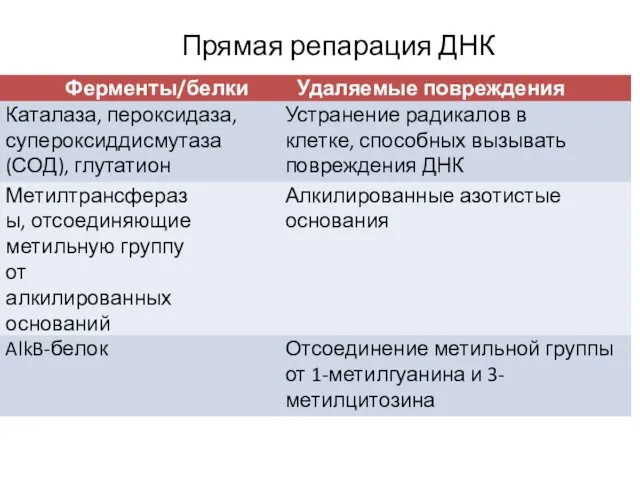
Прямая репарация ДНК
Прямая репарация ДНК
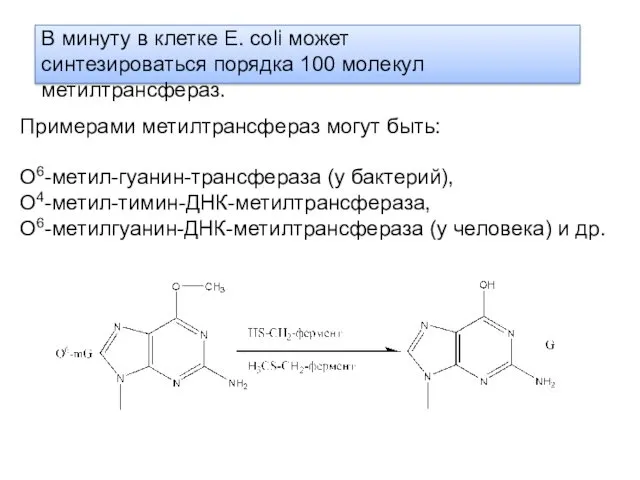
В минуту в клетке Е. coli может синтезироваться порядка 100 молекул
В минуту в клетке Е. coli может синтезироваться порядка 100 молекул

Фотореактивация открыта в 1948 И. Ф. Ковалевым (СССР), А. Келнером и
Фотореактивация открыта в 1948 И. Ф. Ковалевым (СССР), А. Келнером и

Восстановительный эффект при фотореактивации связан с действием фермента — фотолиазы (дезоксирибопиримидинфотолиазы),
Восстановительный эффект при фотореактивации связан с действием фермента — фотолиазы (дезоксирибопиримидинфотолиазы),

За 1 минуту фотолиаза может расщепить 2,4 пиримидиновых димера. У Е.
За 1 минуту фотолиаза может расщепить 2,4 пиримидиновых димера. У Е.
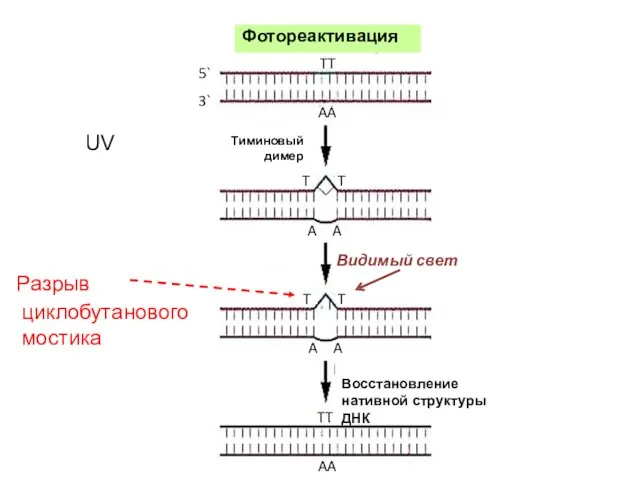
Видимый свет
Разрыв
циклобутанового
мостика
Восстановление нативной структуры ДНК
UV
Фотореактивация
Тиминовый
димер
5`
3`
TT
TT
AA
AA
A
A
A
A
T
T
T
T
Видимый свет
Разрыв
циклобутанового
мостика
Восстановление нативной структуры ДНК
UV
Фотореактивация
Тиминовый
димер
5`
3`
TT
TT
AA
AA
A
A
A
A
T
T
T
T

Репарация АР -сайтов
Образовавшиеся в результате удаления пурина АР-сайты могут быть залечены
Репарация АР -сайтов
Образовавшиеся в результате удаления пурина АР-сайты могут быть залечены

Эксцизионная репарация нуклеотидов
Удаляет:
химические аддукты;
димеры пиримидинов.
Для эксцизионной репарации необходима интактная неповрежденная комплементараная
Эксцизионная репарация нуклеотидов
Удаляет:
химические аддукты;
димеры пиримидинов.
Для эксцизионной репарации необходима интактная неповрежденная комплементараная

Эксцизионная репарация
В настоящее время известно два типа эксцизионной репарации:
Эксцизия азотистых оснований
Эксцизионная репарация
В настоящее время известно два типа эксцизионной репарации:
Эксцизия азотистых оснований

Эксцизионная репарация представляет собой многоэтапный процесс и включает:
) «Узнавание» тиминового димера.
)
Эксцизионная репарация представляет собой многоэтапный процесс и включает:
) «Узнавание» тиминового димера.
)

Таким способом репарируются
следующие повреждения ДНК, включение:
урацила вместо тимина;
гипоксантина;
формамидопиримидина;
5,6 тимина гидрата;
8-окси-гуанина;
5-метил-цитозина;
алкил-аденина;
3-метил-аденина;
7-метил-гуанина.
Таким способом репарируются
следующие повреждения ДНК, включение:
урацила вместо тимина;
гипоксантина;
формамидопиримидина;
5,6 тимина гидрата;
8-окси-гуанина;
5-метил-цитозина;
алкил-аденина;
3-метил-аденина;
7-метил-гуанина.

Эксцизия азотистых оснований
Удаляет специфические повреждения в азотистых основаниях ДНК. Основной фермент – гликозилаза.
Имеется
Эксцизия азотистых оснований
Удаляет специфические повреждения в азотистых основаниях ДНК. Основной фермент – гликозилаза.
Имеется
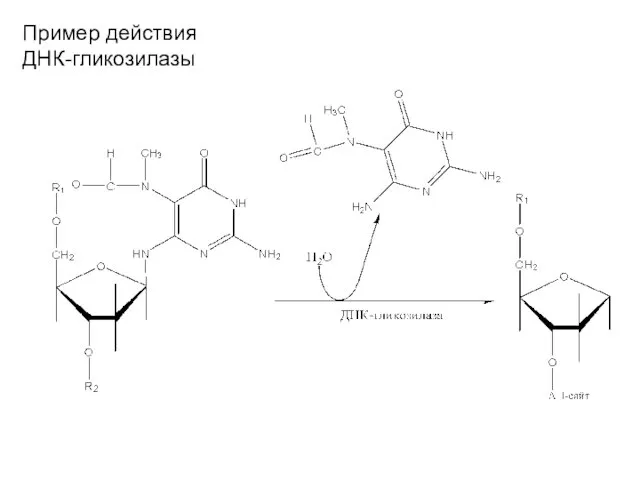
Пример действия ДНК-гликозилазы
Пример действия ДНК-гликозилазы
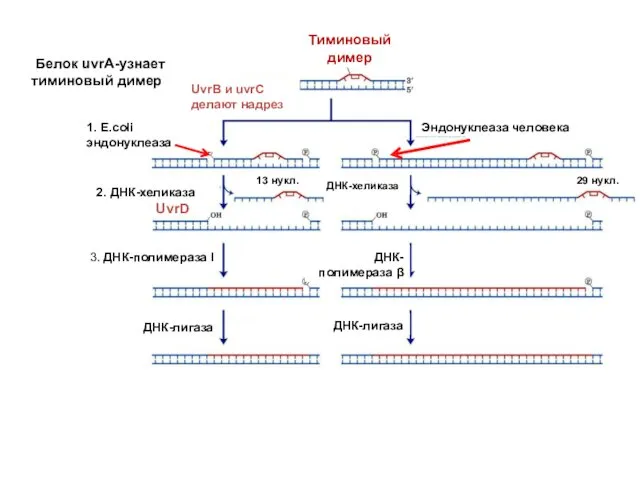
UvrA recognizes bulky lesions
2. ДНК-хеликаза
UvrD
Белок uvrA-узнает тиминовый димер
Тиминовый димер
UvrB и uvrC
делают
UvrA recognizes bulky lesions
2. ДНК-хеликаза
UvrD
Белок uvrA-узнает тиминовый димер
Тиминовый димер
UvrB и uvrC
делают

Некоторые наследственные заболевания человека связаны с дефектом эксцизионной репарации ДНК:
Пигментная ксеродерма,
Некоторые наследственные заболевания человека связаны с дефектом эксцизионной репарации ДНК:
Пигментная ксеродерма,
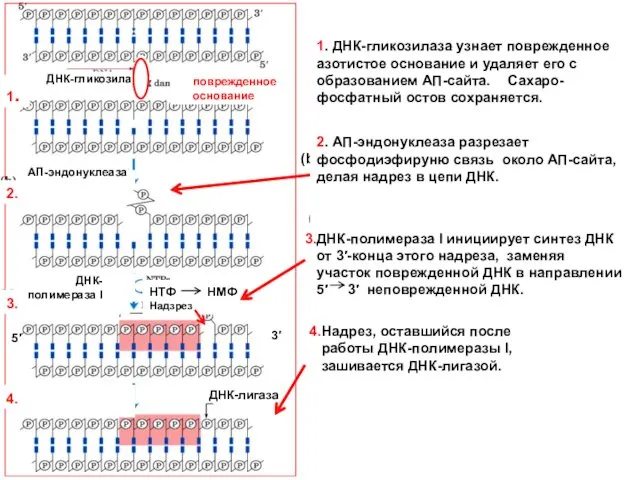
Base excision repair pathway (BER).
(a) A DNA glycosylase recognizes a damaged
Base excision repair pathway (BER).
(a) A DNA glycosylase recognizes a damaged

Известно 3 типа пигментной ксеродермы: XPI, XPII, XPvar, общими симптомами которой
Известно 3 типа пигментной ксеродермы: XPI, XPII, XPvar, общими симптомами которой

Репарация ошибок репликации.
Поскольку ошибки возникают на дочерней цепи, система репарации только
Репарация ошибок репликации. Поскольку ошибки возникают на дочерней цепи, система репарации только
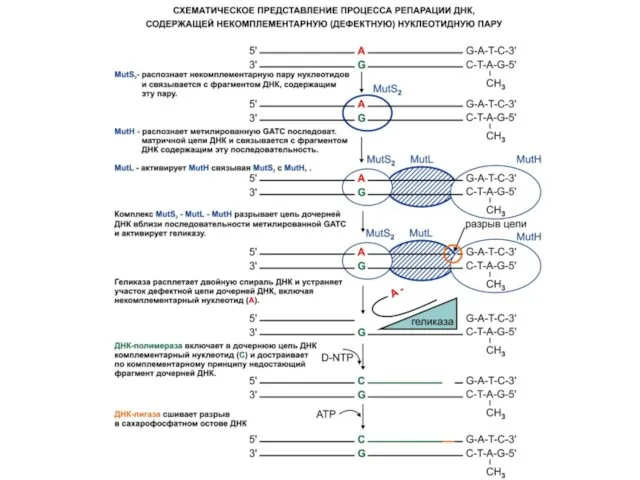
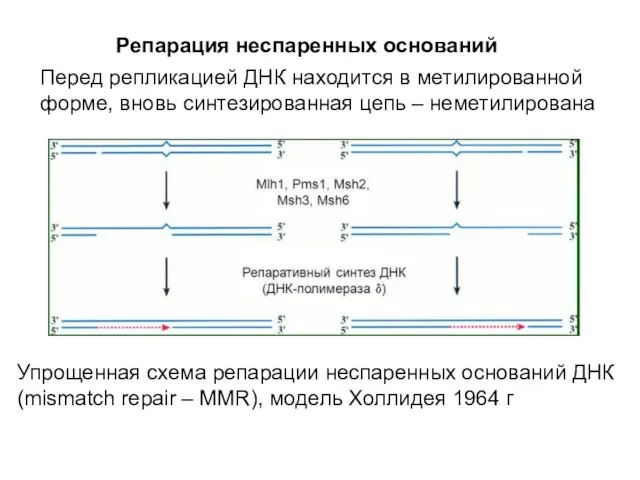
Репарация неспаренных оснований
Перед репликацией ДНК находится в метилированной форме, вновь синтезированная
Репарация неспаренных оснований
Перед репликацией ДНК находится в метилированной форме, вновь синтезированная
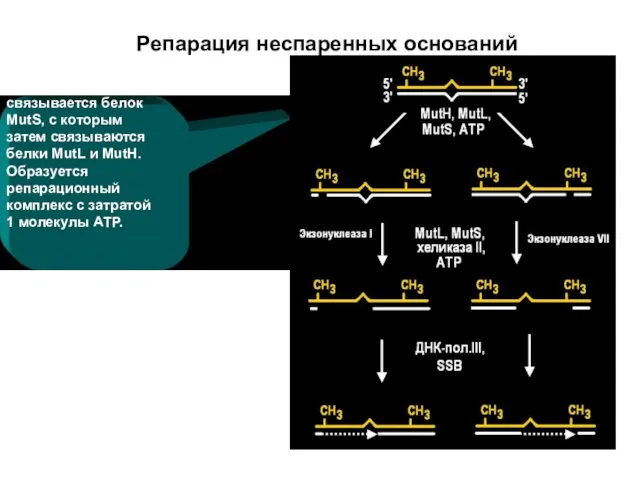
Репарация неспаренных оснований
связывается белок MutS, с которым затем связываются белки MutL
Репарация неспаренных оснований
связывается белок MutS, с которым затем связываются белки MutL
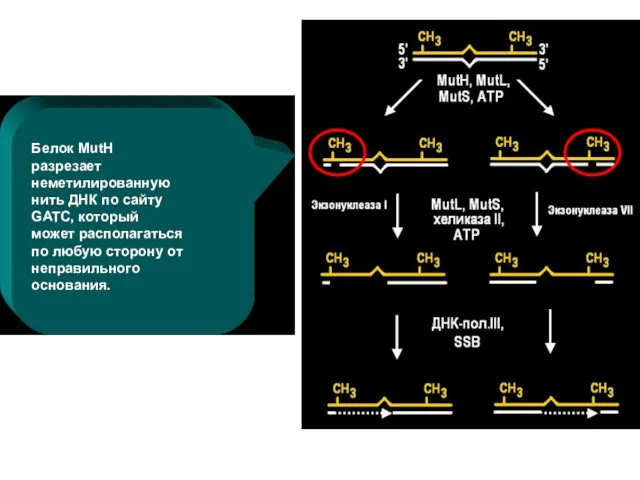
Белок MutH разрезает неметилированную нить ДНК по сайту GATC, который может
Белок MutH разрезает неметилированную нить ДНК по сайту GATC, который может
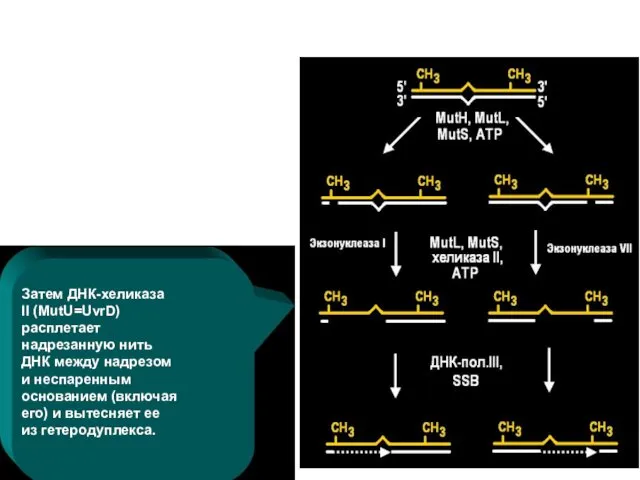
Затем ДНК-хеликаза II (MutU=UvrD) расплетает надрезанную нить ДНК между надрезом и
Затем ДНК-хеликаза II (MutU=UvrD) расплетает надрезанную нить ДНК между надрезом и
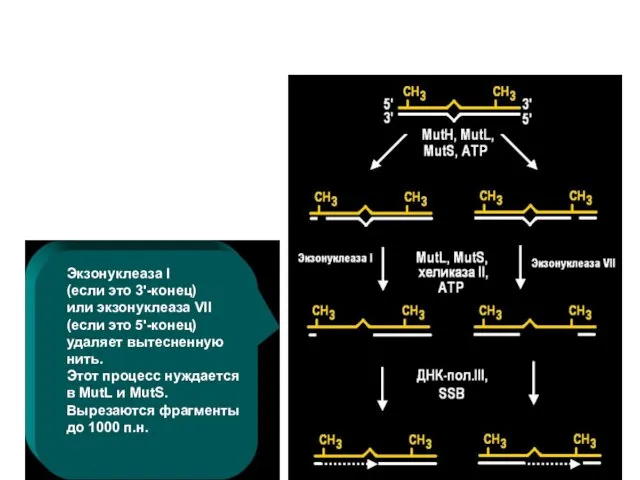
Экзонуклеаза I (если это 3'-конец) или экзонуклеаза VII (если это 5'-конец)
удаляет
Экзонуклеаза I (если это 3'-конец) или экзонуклеаза VII (если это 5'-конец)
удаляет
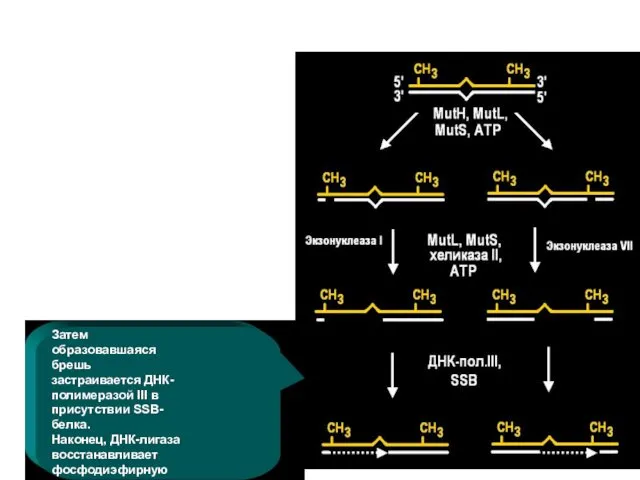
Затем образовавшаяся брешь застраивается ДНК-полимеразой III в присутствии SSB- белка.
Наконец, ДНК-лигаза
Затем образовавшаяся брешь застраивается ДНК-полимеразой III в присутствии SSB- белка.
Наконец, ДНК-лигаза
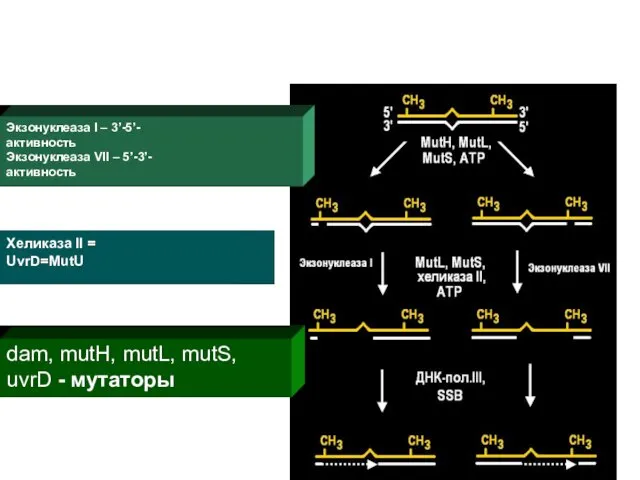
Экзонуклеаза I – 3’-5’- активность Экзонуклеаза VII – 5’-3’- активность
Хеликаза II
Экзонуклеаза I – 3’-5’- активность Экзонуклеаза VII – 5’-3’- активность
Хеликаза II

Пострепликативная репарация происходит когда в ДНК возникает так много повреждений, что
Пострепликативная репарация происходит когда в ДНК возникает так много повреждений, что
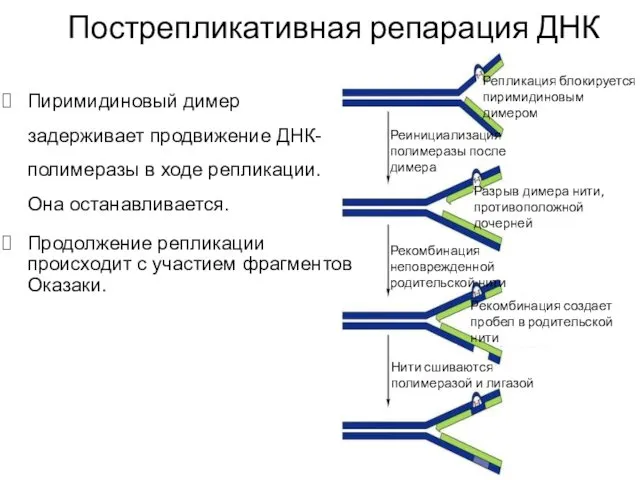
Пиримидиновый димер задерживает продвижение ДНК-полимеразы в ходе репликации. Она останавливается.
Продолжение репликации
Пиримидиновый димер задерживает продвижение ДНК-полимеразы в ходе репликации. Она останавливается.
Продолжение репликации
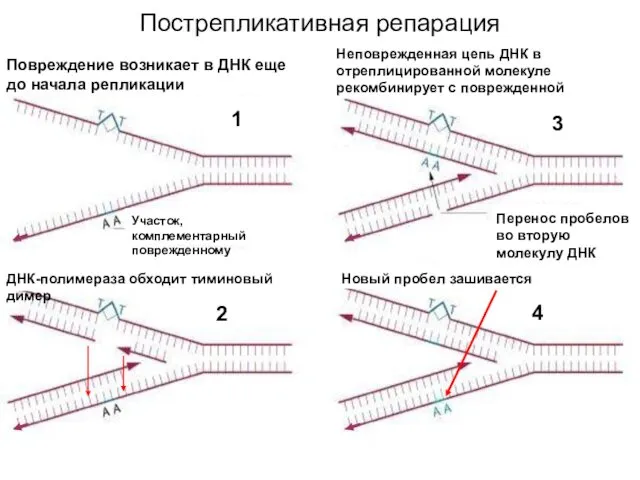
Пострепликативная репарация
Повреждение возникает в ДНК еще до начала репликации
Участок, комплементарный поврежденному
ДНК-полимераза
Пострепликативная репарация
Повреждение возникает в ДНК еще до начала репликации
Участок, комплементарный поврежденному
ДНК-полимераза
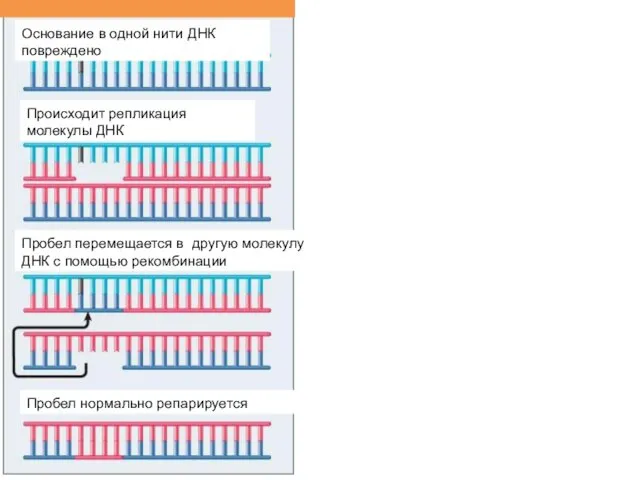
Основание в одной нити ДНК повреждено
Происходит репликация молекулы ДНК
Пробел перемещается в
Основание в одной нити ДНК повреждено
Происходит репликация молекулы ДНК
Пробел перемещается в
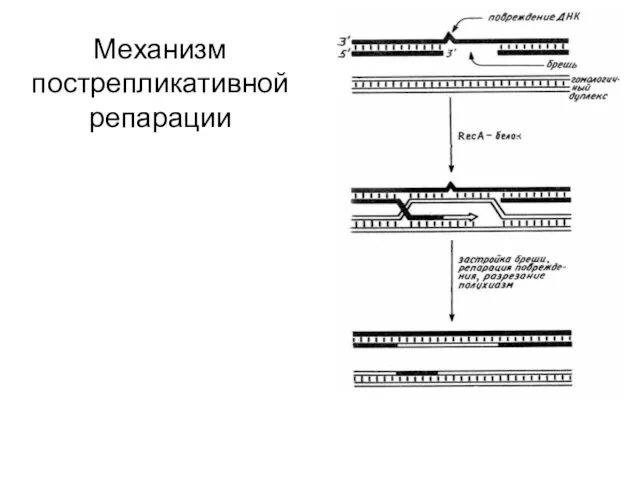
Механизм пострепликативной репарации
Механизм пострепликативной репарации

Существование этой системы впервые постулировал
М. Радман в 1974 г. Он же
Существование этой системы впервые постулировал
М. Радман в 1974 г. Он же

Наиболее изучена SOS-репарация у Е. coli, главными участниками которой являются белки,
Наиболее изучена SOS-репарация у Е. coli, главными участниками которой являются белки,


Начало SOS- ответа определяется взаимодействием белка RecA с белком репрессором LexA.
Начало SOS- ответа определяется взаимодействием белка RecA с белком репрессором LexA.

Одной из функций белка RecA является включение генов umuD и umuC,
Одной из функций белка RecA является включение генов umuD и umuC,

Система рестрикции-модификации
Система рестрикции-модификации — ферментативная система бактерий, разрушающая попавшую в клетку
Система рестрикции-модификации
Система рестрикции-модификации — ферментативная система бактерий, разрушающая попавшую в клетку

Системы рестрикции-модификации были открыты в результате изучения молекулярных механизмов явления, называемого
Системы рестрикции-модификации были открыты в результате изучения молекулярных механизмов явления, называемого
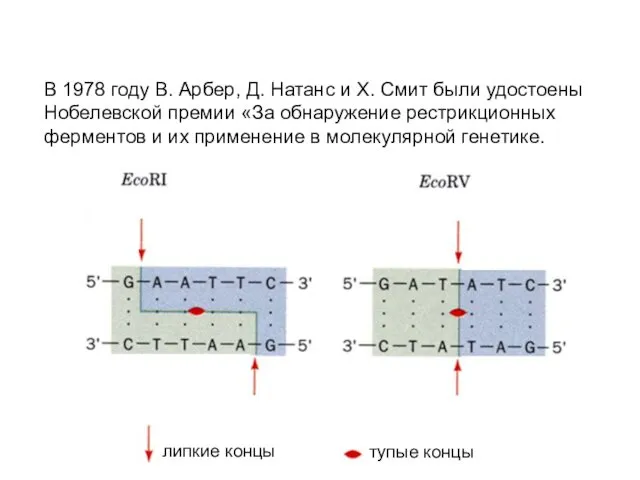
В 1978 году В. Арбер, Д. Натанс и Х. Смит были
В 1978 году В. Арбер, Д. Натанс и Х. Смит были

Система рестрикции-модификации специфична по отношению к определѐнным последовательностям нуклеотидов в ДНК,
Система рестрикции-модификации специфична по отношению к определѐнным последовательностям нуклеотидов в ДНК,
 Биоэнергетика. Биологическое окисление. Биологические виды энергии
Биоэнергетика. Биологическое окисление. Биологические виды энергии Поверхностное и коренное улучшение сенокосов и пастбищ
Поверхностное и коренное улучшение сенокосов и пастбищ Класс млекопитающие или звери
Класс млекопитающие или звери Соцветие. Виды соцветий
Соцветие. Виды соцветий Як бачать тварини. 7 клас
Як бачать тварини. 7 клас Игра В мире животных
Игра В мире животных Викторина про насекомых
Викторина про насекомых Пингвины (фото и картинки)
Пингвины (фото и картинки) Растения Красной книги Воронежской области
Растения Красной книги Воронежской области Органы осязания, обоняния и вкуса
Органы осязания, обоняния и вкуса Презентация Насекомые Красной Книги
Презентация Насекомые Красной Книги Зарождение человека. Процесс оплодотворения. Эмбриональное развитие плода
Зарождение человека. Процесс оплодотворения. Эмбриональное развитие плода Презентация к уроку на тему:Бесполое размножение 10 класс
Презентация к уроку на тему:Бесполое размножение 10 класс Решение задач на моногибридное скрещивание и взаимодействие аллельных генов
Решение задач на моногибридное скрещивание и взаимодействие аллельных генов Nucleic acids
Nucleic acids Интерактивная игра Экологическое Ассорти
Интерактивная игра Экологическое Ассорти Будова, роль та значення полісахаридів у метаболізмі рослинної клітини
Будова, роль та значення полісахаридів у метаболізмі рослинної клітини Птицы из Красной книги Югры
Птицы из Красной книги Югры Устройство увеличительных приборов
Устройство увеличительных приборов Генетические механизмы контроля формирования пола
Генетические механизмы контроля формирования пола Тип круглые черви — Нематоды. 7 класс
Тип круглые черви — Нематоды. 7 класс Особливості реалізації стійкості рослин
Особливості реалізації стійкості рослин Класс паукообразные
Класс паукообразные Жасуша құрылысы
Жасуша құрылысы Өсімдіктердің генетикалық полиморфизмін талдау
Өсімдіктердің генетикалық полиморфизмін талдау презентация Пушистый доктор
презентация Пушистый доктор Методы исследования генетики человека
Методы исследования генетики человека Многообразие грибов. Значение грибов в природе и жизни человека
Многообразие грибов. Значение грибов в природе и жизни человека