- Биологические мембраны

Содержание
- 2. ОПРЕДЕЛЕНИЕ И ОБЩАЯ ХАРАКТЕРИСТИКА БИОЛОГИЧЕСКИХ МЕМБРАН Биологические мембраны – сложные высокоорганизованные липопротеиновые структуры, окружающие клетку и
- 3. Рис. Мозаичная структура мембран
- 4. Рис. Структура липидного бислоя. Мембранные фосфолипиды расположены в форме бислоя, который образует жидкокристаллическую матрицу. Каждый фосфолипидный
- 5. Отдельные молекулы фосфолипидов могут перемещаться между наружным и внутренним монослоем мембраны. В этом процессе участвуют особые
- 6. Рис. Перенос молекулы фосфолипида из внешнего монослоя во внутренний с помощью флиппазы (схема): 1 - молекула
- 7. ЛИПИДНЫЙ БИСЛОЙ Образно говоря, липиды биологических мембран представляют собой море, в котором плавают мембранные белки. Благодаря
- 8. Мембранные фосфолипиды Пространственная структура и размер полярной области фосфолипидов влияют на плотность и пространственное расположение в
- 9. Ассимметрия фосфолипидов в мембране Биологические мембраны характеризуются структурной асимметрией, которая обусловлена различным распределением липидов между внутренними
- 10. Ряд фосфолипидов обнаружены в мембранах внутриклеточных структур. Например, кардиолипин локализован главным образом во внутренней мембране митохондрий,
- 11. Мембранные белки На состояние мембранных липидов влияет и их белковый состав. В зависимости от локализации и
- 12. Рис. Мембранные белки
- 13. Интегральные белки могут прошивать всю толщу мембраны (в этом случае их называют трансмембранными) или же быть
- 14. Рис. Расположение трансмембранных доменов пол и пептидной цепи β2-адренорецептора в плазматической мембране клетки (схема)
- 15. Некоторые липиды являются эффективными аллостерическими регуляторами мембранных ферментов. Например, фосфатидилсерин способствует связыванию протеинкиназы С с внутренним
- 16. Мембранные липиды образуют вокруг мембраносвязанных белков гидрофобную область, которая способствует взаимодействию мембранных ферментов со своими субстратами.
- 17. Латеральная неоднородность (гетерогенность) биологических мембран Гетерогенность липидов в плоскости мембраны является следствием образования различных ассоциаций липидных
- 18. ФУНКЦИИ МЕМБРАН Мембраны выполняют разнообразные функции, которые определяются типом клеток и той ролью, которую они играют
- 19. ОБЩАЯ ХАРАКТЕРИСТИКА Главной функцией биомембран является избирательный транспорт различных веществ и ионов. Обмен между клеткой и
- 20. Рис. Анионный канал мембраны эритроцитов. Хлорид- и бикарбонат-ионы двигаются в противоположных направлениях по градиентам концентраций.
- 21. Другой вид трансмембранного переноса веществ против градиента концентраций совершается за счет внешнего источника энергии, которым является
- 22. Каталитический цикл начинается со связывания трех ионов натрия с Na+,К+-АТФазой на цитоплазматической стороне мембраны. Далее происходит
- 23. Особенности функционирования мембранных транспортеров Унипорт - перенос любых веществ (в том числе ионов) с одной стороны
- 24. ГЛЮТ1 связывает молекулу глюкозы на внешней стороне плазматической мембраны. В результате последующей серии конформационных изменений ГЛЮТ1
- 25. Рис. Совместный транспорт глюкозы и ионов натрия в клетку. Поскольку ионы натрия все время выкачиваются из
- 26. Рис. Система совместного транспорта глюкозы и ионов натрия Подобный механизм функционирует и в случае Na+-зависимого всасывания
- 27. Антипорт — совместный трансмембранный перенос двух веществ и более в противоположном направлении. Примером антипорта, который осуществляется
- 28. Каналы – важный класс систем пассивного транспорта. В их образовании могут участвовать белки, специфичные для определенного
- 29. У человека обнаружено не менее 11 аквапориноподобных белков. Физиологическая важность аквапоринов наиболее заметна в почках, где
- 30. ТРАНСМЕМБРАННЫЙ ПЕРЕНОС МАКРОМОЛЕКУЛ, НАДМОЛЕКУЛЯРНЫХ КОМПЛЕКСОВ И ЧАСТИЦ Помимо транспорта низкомолекулярных веществ, биологические мембраны осуществляют перенос различных
- 31. Выделяют несколько вариантов эндоцитоза: рецептор – опосредованный эндоцитоз, фагоцитоз и пиноцитоз. Рецептор-опосредованный эндоцитоз - специфический способ
- 32. Фагоцитоз Захват и поглощение инородных частиц фагоцитирующими клетками или фагоцитами называют фагоцитозом. К фагоцитам относят нейтрофилы
- 33. Пиноцитоз Процесс поглощения клеткой жидкости ( с растворенными в ней веществами) из окружающей среды называют пиноцитозом.
- 34. Экзоцитоз — выведение веществ из клетки. В ходе этого процесса экспортируемые молекулы упаковываются в секреторные везикулы
- 35. ПАРАКЛЕТОЧНЫЙ ТРАНСПОРТ Помимо трансклеточного транспорта веществ, в котором задействованы различные клеточные поры, каналы и переносчики, электролиты,
- 38. Скачать презентацию

ОПРЕДЕЛЕНИЕ И ОБЩАЯ ХАРАКТЕРИСТИКА
БИОЛОГИЧЕСКИХ МЕМБРАН
Биологические мембраны – сложные высокоорганизованные липопротеиновые структуры,
ОПРЕДЕЛЕНИЕ И ОБЩАЯ ХАРАКТЕРИСТИКА
БИОЛОГИЧЕСКИХ МЕМБРАН
Биологические мембраны – сложные высокоорганизованные липопротеиновые структуры,
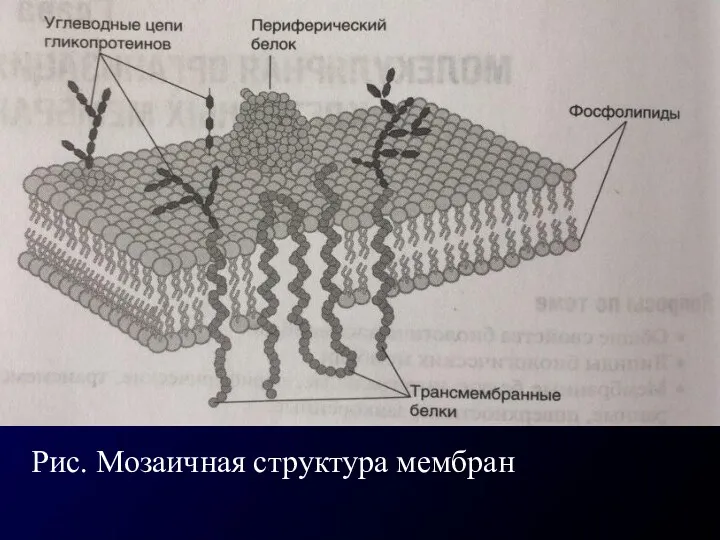
Рис. Мозаичная структура мембран
Рис. Мозаичная структура мембран
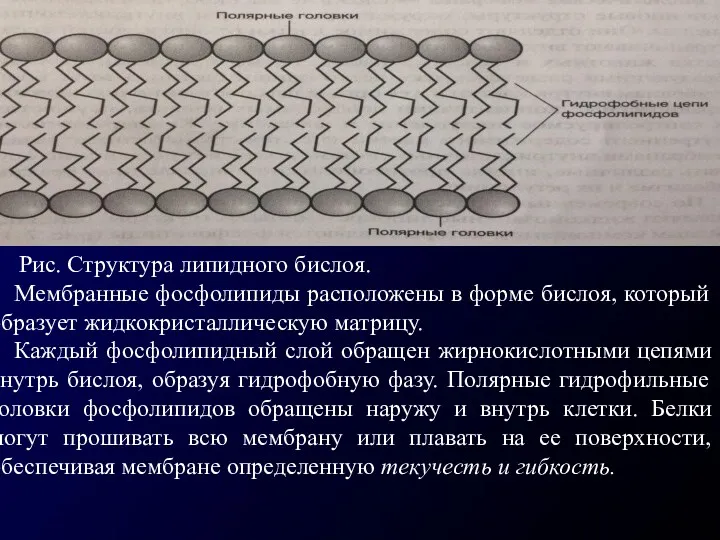
Рис. Структура липидного бислоя.
Мембранные фосфолипиды расположены в форме бислоя,
Рис. Структура липидного бислоя.
Мембранные фосфолипиды расположены в форме бислоя,

Отдельные молекулы фосфолипидов могут перемещаться между наружным и внутренним монослоем мембраны.
Отдельные молекулы фосфолипидов могут перемещаться между наружным и внутренним монослоем мембраны.

Рис. Перенос молекулы фосфолипида из внешнего монослоя во внутренний с помощью
Рис. Перенос молекулы фосфолипида из внешнего монослоя во внутренний с помощью

ЛИПИДНЫЙ БИСЛОЙ
Образно говоря, липиды биологических мембран представляют собой море, в котором
ЛИПИДНЫЙ БИСЛОЙ
Образно говоря, липиды биологических мембран представляют собой море, в котором

Мембранные фосфолипиды
Пространственная структура и размер полярной области фосфолипидов влияют на
Мембранные фосфолипиды
Пространственная структура и размер полярной области фосфолипидов влияют на

Ассимметрия фосфолипидов в мембране
Биологические мембраны характеризуются структурной асимметрией, которая обусловлена
Ассимметрия фосфолипидов в мембране
Биологические мембраны характеризуются структурной асимметрией, которая обусловлена

Ряд фосфолипидов обнаружены в мембранах внутриклеточных структур. Например, кардиолипин локализован главным
Ряд фосфолипидов обнаружены в мембранах внутриклеточных структур. Например, кардиолипин локализован главным

Мембранные белки
На состояние мембранных липидов влияет и их белковый состав. В
Мембранные белки
На состояние мембранных липидов влияет и их белковый состав. В
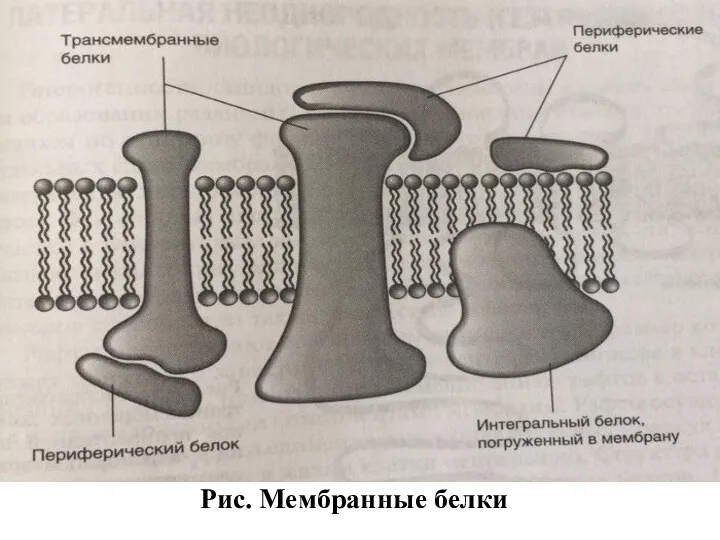
Рис. Мембранные белки
Рис. Мембранные белки

Интегральные белки могут прошивать всю толщу мембраны (в этом случае их
Интегральные белки могут прошивать всю толщу мембраны (в этом случае их
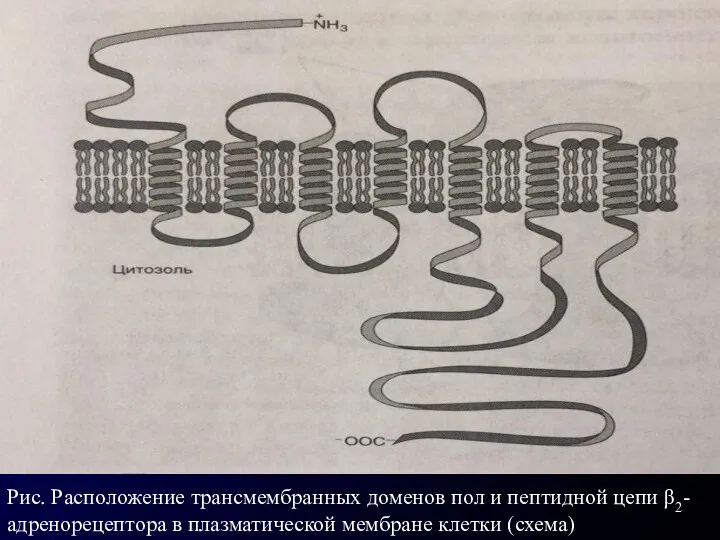
Рис. Расположение трансмембранных доменов пол и пептидной цепи β2-адренорецептора в плазматической
Рис. Расположение трансмембранных доменов пол и пептидной цепи β2-адренорецептора в плазматической

Некоторые липиды являются эффективными аллостерическими регуляторами мембранных ферментов. Например, фосфатидилсерин способствует
Некоторые липиды являются эффективными аллостерическими регуляторами мембранных ферментов. Например, фосфатидилсерин способствует

Мембранные липиды образуют вокруг мембраносвязанных белков гидрофобную область, которая способствует
Мембранные липиды образуют вокруг мембраносвязанных белков гидрофобную область, которая способствует

Латеральная неоднородность (гетерогенность) биологических мембран
Гетерогенность липидов в плоскости мембраны является следствием
Латеральная неоднородность (гетерогенность) биологических мембран
Гетерогенность липидов в плоскости мембраны является следствием

ФУНКЦИИ МЕМБРАН
Мембраны выполняют разнообразные функции, которые
определяются типом клеток
ФУНКЦИИ МЕМБРАН
Мембраны выполняют разнообразные функции, которые
определяются типом клеток

ОБЩАЯ ХАРАКТЕРИСТИКА
Главной функцией биомембран является избирательный транспорт различных веществ и
ОБЩАЯ ХАРАКТЕРИСТИКА
Главной функцией биомембран является избирательный транспорт различных веществ и
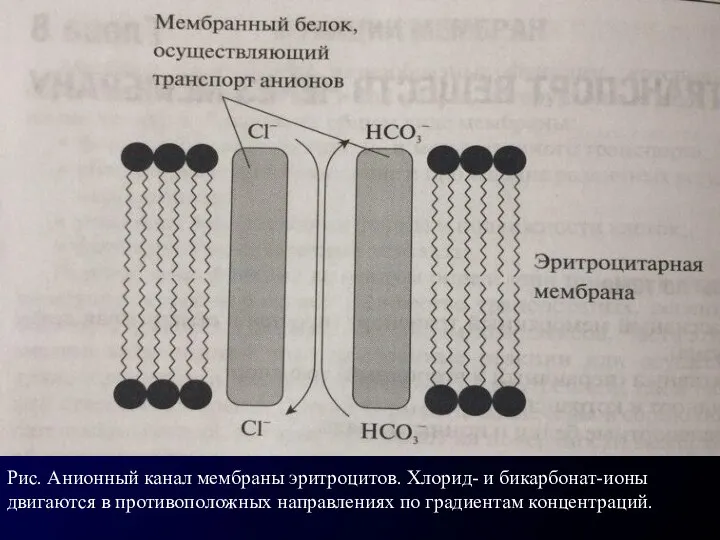
Рис. Анионный канал мембраны эритроцитов. Хлорид- и бикарбонат-ионы двигаются в противоположных
Рис. Анионный канал мембраны эритроцитов. Хлорид- и бикарбонат-ионы двигаются в противоположных

Другой вид трансмембранного переноса веществ против градиента концентраций совершается за счет
Другой вид трансмембранного переноса веществ против градиента концентраций совершается за счет
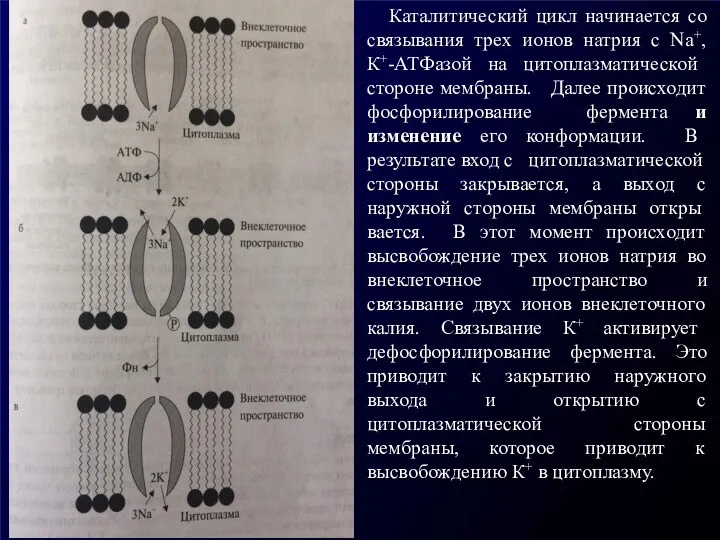
Каталитический цикл начинается со связывания трех ионов натрия с Na+,К+-АТФазой
Каталитический цикл начинается со связывания трех ионов натрия с Na+,К+-АТФазой
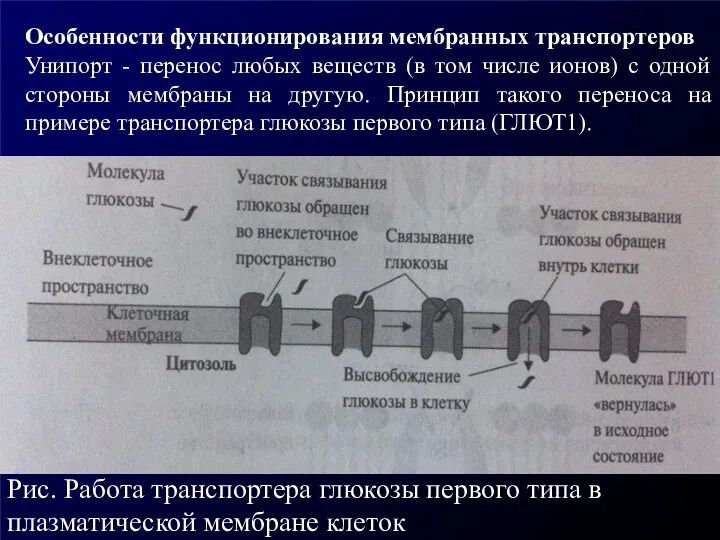
Особенности функционирования мембранных транспортеров
Унипорт - перенос любых веществ (в том числе
Особенности функционирования мембранных транспортеров
Унипорт - перенос любых веществ (в том числе

ГЛЮТ1 связывает молекулу глюкозы на внешней стороне плазматической мембраны. В результате
ГЛЮТ1 связывает молекулу глюкозы на внешней стороне плазматической мембраны. В результате

Рис. Совместный транспорт глюкозы и ионов натрия в клетку.
Поскольку ионы натрия
Рис. Совместный транспорт глюкозы и ионов натрия в клетку.
Поскольку ионы натрия
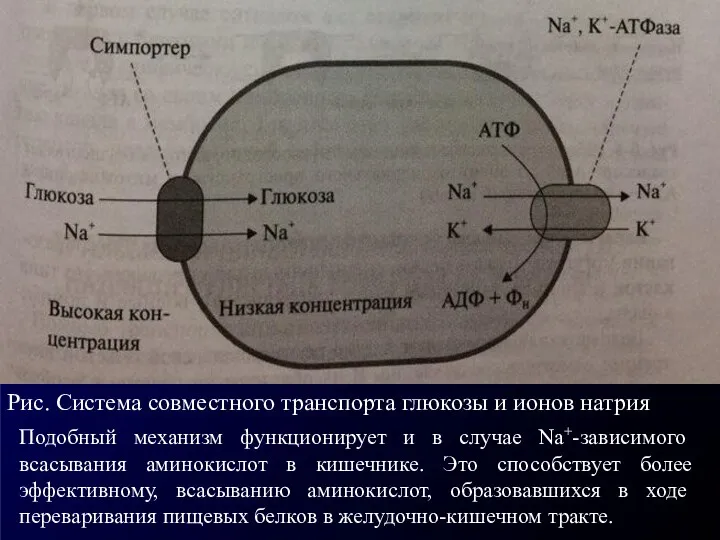
Рис. Система совместного транспорта глюкозы и ионов натрия
Подобный механизм функционирует и
Рис. Система совместного транспорта глюкозы и ионов натрия
Подобный механизм функционирует и

Антипорт — совместный трансмембранный перенос двух веществ и более в противоположном
Антипорт — совместный трансмембранный перенос двух веществ и более в противоположном
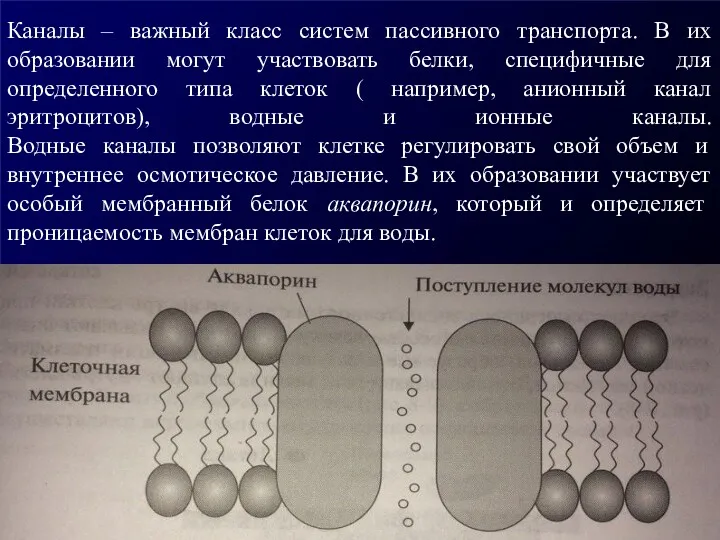
Каналы – важный класс систем пассивного транспорта. В их образовании могут
Каналы – важный класс систем пассивного транспорта. В их образовании могут

У человека обнаружено не менее 11 аквапориноподобных белков. Физиологическая важность аквапоринов
У человека обнаружено не менее 11 аквапориноподобных белков. Физиологическая важность аквапоринов
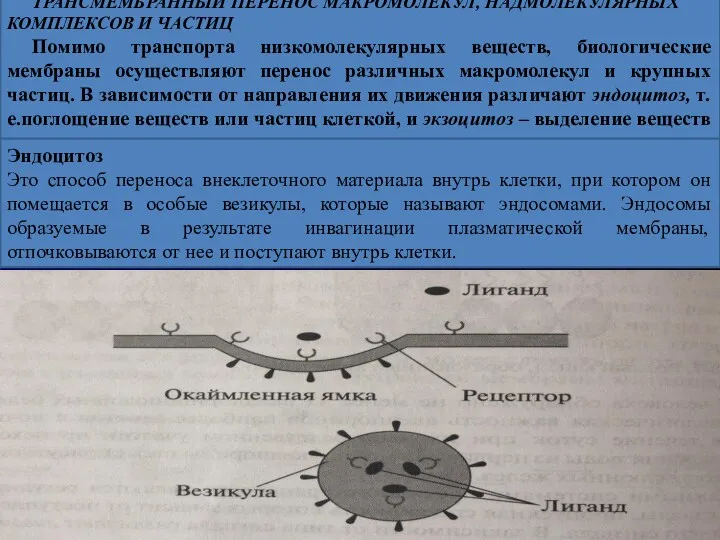
ТРАНСМЕМБРАННЫЙ ПЕРЕНОС МАКРОМОЛЕКУЛ, НАДМОЛЕКУЛЯРНЫХ КОМПЛЕКСОВ И ЧАСТИЦ
Помимо транспорта низкомолекулярных веществ, биологические
ТРАНСМЕМБРАННЫЙ ПЕРЕНОС МАКРОМОЛЕКУЛ, НАДМОЛЕКУЛЯРНЫХ КОМПЛЕКСОВ И ЧАСТИЦ
Помимо транспорта низкомолекулярных веществ, биологические

Выделяют несколько вариантов эндоцитоза: рецептор – опосредованный эндоцитоз, фагоцитоз и пиноцитоз.
Рецептор-опосредованный
Выделяют несколько вариантов эндоцитоза: рецептор – опосредованный эндоцитоз, фагоцитоз и пиноцитоз.
Рецептор-опосредованный
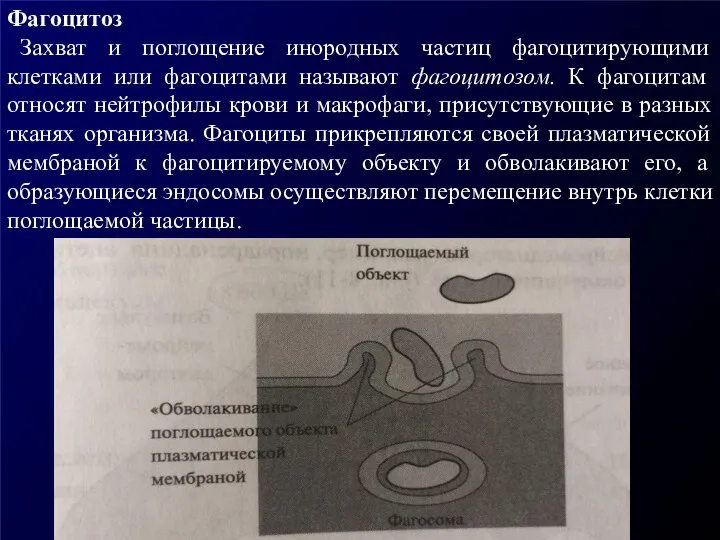
Фагоцитоз
Захват и поглощение инородных частиц фагоцитирующими клетками или фагоцитами называют
Фагоцитоз Захват и поглощение инородных частиц фагоцитирующими клетками или фагоцитами называют
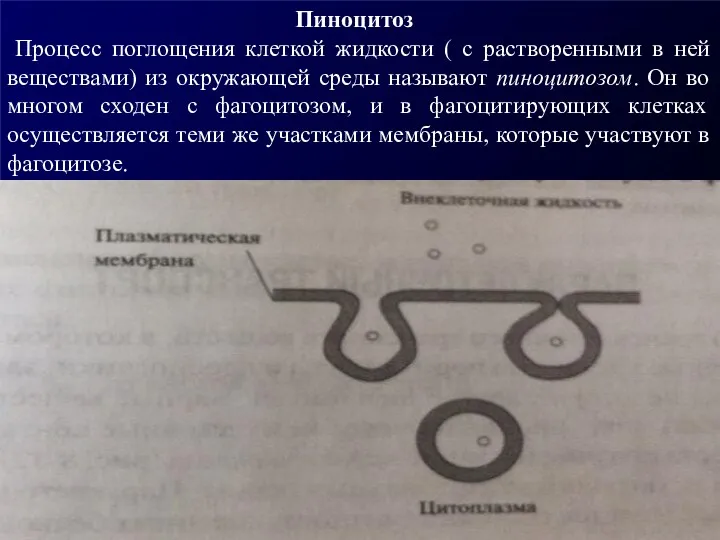
Пиноцитоз
Процесс поглощения клеткой жидкости ( с растворенными в ней
Пиноцитоз
Процесс поглощения клеткой жидкости ( с растворенными в ней
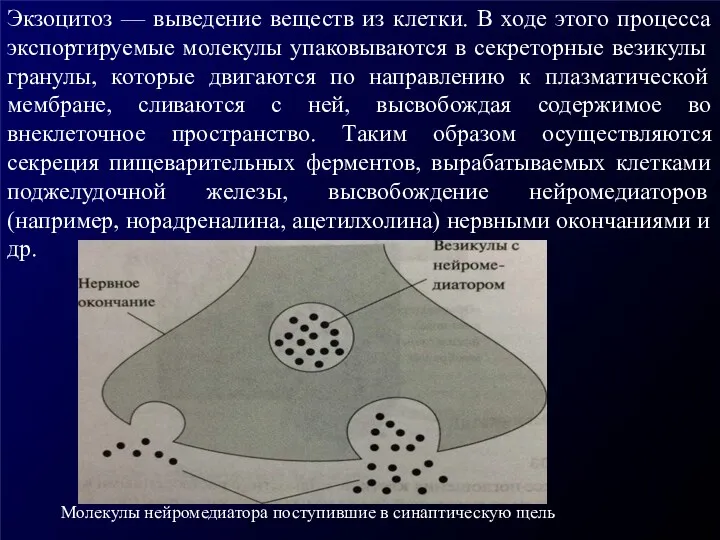
Экзоцитоз — выведение веществ из клетки. В ходе этого процесса экспортируемые
Экзоцитоз — выведение веществ из клетки. В ходе этого процесса экспортируемые

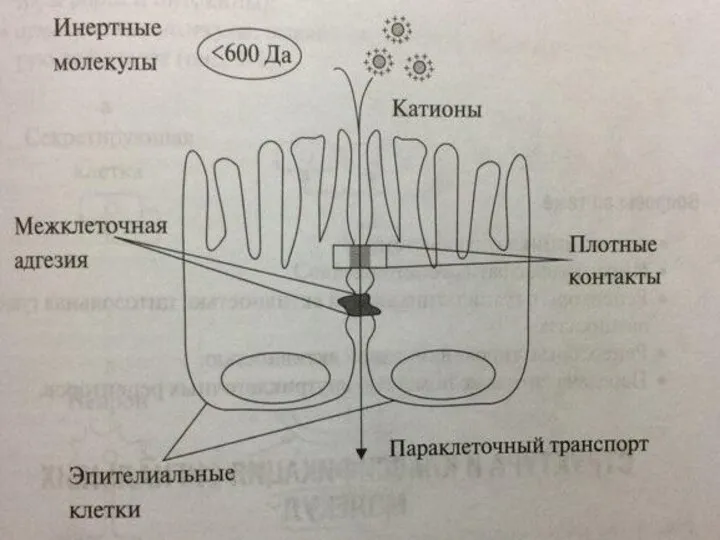
ПАРАКЛЕТОЧНЫЙ ТРАНСПОРТ
Помимо трансклеточного транспорта веществ, в котором задействованы различные клеточные поры,
ПАРАКЛЕТОЧНЫЙ ТРАНСПОРТ
Помимо трансклеточного транспорта веществ, в котором задействованы различные клеточные поры,
 Картофель. Сорт-стандарт Невский
Картофель. Сорт-стандарт Невский Презентация к уроку Дыхательные движения. Газообмен
Презентация к уроку Дыхательные движения. Газообмен f611e845-2141-4c61-a9b2-34e2049b0658
f611e845-2141-4c61-a9b2-34e2049b0658 Артерии. Вены. Лимфатическая система
Артерии. Вены. Лимфатическая система Формы естественного отбора
Формы естественного отбора Изменчивость. Формы изменчивости
Изменчивость. Формы изменчивости Царство прокариоты, подцарство бактерии
Царство прокариоты, подцарство бактерии Естественный отбор - направляющий фактор эволюции
Естественный отбор - направляющий фактор эволюции Клебсиеллы
Клебсиеллы Презентация Здоровье – ключ к успеху
Презентация Здоровье – ключ к успеху Строение и функционирование генома
Строение и функционирование генома Fiziologia_endokr_sist
Fiziologia_endokr_sist Виды осетровых и их краткое описание
Виды осетровых и их краткое описание презентация Кастовая система семьи медоносной пчелы
презентация Кастовая система семьи медоносной пчелы Эволюционная теория. Вид, его критерии. Популяция как структурная единица вида и эволюции
Эволюционная теория. Вид, его критерии. Популяция как структурная единица вида и эволюции ПРЕЗЕНТАЦИИ 5 КЛАСС 2 ЧЕТВЕРТЬ
ПРЕЗЕНТАЦИИ 5 КЛАСС 2 ЧЕТВЕРТЬ Углеводы. Глюкоза
Углеводы. Глюкоза Ямчатость стебля – обзор, физиология
Ямчатость стебля – обзор, физиология Голосовой аппарат
Голосовой аппарат урок-презентация по теме Зеленая аптека
урок-презентация по теме Зеленая аптека Жасушалық биология
Жасушалық биология История развития органического мира
История развития органического мира Вклад отечественных ученых в разработку учения о высшей нервной деятельности
Вклад отечественных ученых в разработку учения о высшей нервной деятельности Основные константы крови человека. Физико – химические свойства крови
Основные константы крови человека. Физико – химические свойства крови Всасывание, пищеварение
Всасывание, пищеварение Химический состав клетки
Химический состав клетки Химические свойства воды. Значение воды в жизни человека
Химические свойства воды. Значение воды в жизни человека Игра по окружающему миру для 5 класса
Игра по окружающему миру для 5 класса