- Ямчатость стебля – обзор, физиология

Содержание
- 2. ОБЗОР ИЗМЕНЕНИЙ В СТРОЕНИИ ДРЕВЕСИНЫ, СВЯЗАННЫХ С ЛУЧЕВЫМИ АНОМАЛИЯМИ СТРУКТУРНЫЕ ИЗМЕНЕНИЯ Рассмотренные в предыдущей главе частные
- 3. Синдром ямчатости стебля во всех случаях его проявления сопровождается вполне определенными изменениями в строении древесины. Сопоставляя
- 4. Первым признаком развития аномалии, распознаваемым с помощью светового микроскопа, является нарушение в строении лучей. Один луч
- 5. В дальнейшем паренхима в аномальной зоне может так или иначе дифференцироваться: меняется окраска и оптическая плотность
- 6. В процессе нормализации строения «ямка» на поверхности древесинного цилиндра нивелируется далеко не сразу, так как в
- 8. Наши наблюдения и анализ данных литературы показали, что такая последовательность структурных изменений справедлива для любого случая
- 9. Характерной и весьма существенной, с нашей точки зрения, особенностью аномальных участков древесины является их способность на
- 10. Наши наблюдения показали (Коровин, 1970), что во флоэме капов, в зоне возникающих придаточных почек склереиды практически
- 11. Заканчивая рассмотрение анатомических изменений, возникающих в древесине при развитии синдрома ямчатости стебля, мы приходим к выводу,
- 12. В заключение данного раздела следует добавить, что структурные неспецифические реакции древесных растений на самые различные денормализующие
- 13. Первыми возникают два взаимно связанных вопроса: какие нарушения регуляции ростовых процессов приводят к образованию аномальных лучей,
- 14. Здесь же следует добавить, что общепринятые представления о регуляции камбиальной активности ростовыми веществами отражают лишь определенный
- 15. Известно, например, что равномерно расположенные по поверхности древесинного цилиндра аномальные зоны у явора с текстурой древесины
- 16. Величина радиального годичного прироста стволовой древесины достоверно больше средней лишь в верхней части ствола молодых, интенсивно
- 17. Относительная независимость от апикального доминирования наблюдается и в процессе дифференциации производных камбия. Относительная потому, что единому
- 18. Относительно потому, что увеличение поверхности ствола или ветви, происходящее вследствие антиклинальных делений, связано прямой функциональной зависимостью
- 19. Изложенные соображения об относительной независимости ростовых процессов, связанных с камбиальной активностью, от апикального доминирования мы не
- 20. Идея о решающей роли нарушения баланса ростовых веществ при формировании аномальной древесины карельской березы находит свое
- 21. Высказывается предположение, что та же причина лежит в основе формирования ложных годичных слоев древесины, связанного с
- 22. В связи с предметом обсуждения представляется интересным тот факт, что среди древесных растений наиболее высокое содержание
- 23. Синонимом паренхиматизации во многих случаях можно назвать нарушение дифференциации. Паренхиматизация древесины может происходить за счет образования
- 24. Если принять, что скопление паренхимы, возникающее при формировании аномального луча, представляет собой определенный этап в образовании
- 25. Так что же мешает превращению аномального скопления паренхимы в настоящий очаг меристематической ткани? Вероятнее всего –
- 26. Теперь остается высказать наши соображения по поводу ранее поставленных вопросов: как контролируется частота и относительная регулярность
- 27. По мнению E. Bünning, и Sagromsky (1948) равномерное распределение лучей можно объяснить их ингибирующим действием друг
- 28. Многочисленные примеры этого были приведены ранее при рассмотрении особенностей строения конкретных аномальных древесин, они могут быть
- 29. РОЛЬ ЛУЧЕЙ В ЗАЖИВЛЕНИИ ПОВРЕЖДЕНИЙ СТЕБЛЯ И ДРУГИХ СЛУЧАЯХ ФОРМИРОВАНИЯ АНОМАЛЬНЫХ УЧАСТКОВ ДРЕВЕСИНЫ По мнению P.Блоха
- 30. Разумеется, мы далеки от отождествления понятий “аномальный рост” и “регенерация”, тем более, что регенерация – явление,
- 31. Гартиг обратил внимание на то, что наибольшей способностью к регенерации обладает камбиальная зона и паренхима древесинных
- 32. Гомогенность раневого каллуса сохраняется ограниченное время, вскоре в нем дифференцируется лучевая паренхима и короткие трахеальные элементы,
- 33. Дедифференциация клеток при нормальном развитии явление не частое. Однако, по мнению автора, она играет очень важную
- 34. Изучение обнаженных поверхностей показало, что разделение тканей коры и древесины произошло по зоне материнских клеток ксилемы
- 36. Разрастание клеток над раневой поверхностью древесины можно рассматривать как образование первичного каллюса, характерной особенностью клеток которого
- 37. При проведении подобных исследований необходимо иметь в виду, что данные по срокам развития каллюса следует рассматривать
- 40. Анатомические изменения при зарастании механических повреждений стебля мало отличаются от изменений, происходящих в стволе древесных растений
- 41. При слабом повреждении расширяющиеся лучи могут не смыкаться, между ними в этом случае продолжают дифференцироваться осевые
- 42. По нашим наблюдениям и данным литературы (Воусе, 1923, 1961), процесс зарастания на стволах следов молнии в
- 43. Древесинные лучи принимают участие в локализации и иного рода повреждений. Широко распространенным пороком деловой древесины березы
- 45. По нашим наблюдениям, аномальная паренхимная прослойка сердцевинных повторений возникает вследствие зарастания полости, проеденной личинкой, клетками ,
- 46. Обобщив результаты наших наблюдении и данные литературы, мы пришли к убеждению, что лучи помимо функций запасания
- 47. В порядке дополнительного подтверждения высказанных соображений приведем несколько примеров из известных работ М.В. Беннена (Ваnnаn, 1941,
- 50. Многие аномальные лучи, достигнув предельных размеров, расчленяются трахеидами на более мелкие, которые в свою очередь в
- 52. Скачать презентацию

ОБЗОР ИЗМЕНЕНИЙ В СТРОЕНИИ ДРЕВЕСИНЫ,
СВЯЗАННЫХ С ЛУЧЕВЫМИ АНОМАЛИЯМИ
СТРУКТУРНЫЕ ИЗМЕНЕНИЯ
Рассмотренные в предыдущей
ОБЗОР ИЗМЕНЕНИЙ В СТРОЕНИИ ДРЕВЕСИНЫ,
СВЯЗАННЫХ С ЛУЧЕВЫМИ АНОМАЛИЯМИ
СТРУКТУРНЫЕ ИЗМЕНЕНИЯ
Рассмотренные в предыдущей

Синдром ямчатости стебля во всех случаях его проявления сопровождается вполне определенными
Синдром ямчатости стебля во всех случаях его проявления сопровождается вполне определенными

Первым признаком развития аномалии, распознаваемым с помощью светового микроскопа, является нарушение
Первым признаком развития аномалии, распознаваемым с помощью светового микроскопа, является нарушение

В дальнейшем паренхима в аномальной зоне может так или иначе дифференцироваться:
В дальнейшем паренхима в аномальной зоне может так или иначе дифференцироваться:

В процессе нормализации строения «ямка» на поверхности древесинного цилиндра нивелируется далеко
В процессе нормализации строения «ямка» на поверхности древесинного цилиндра нивелируется далеко
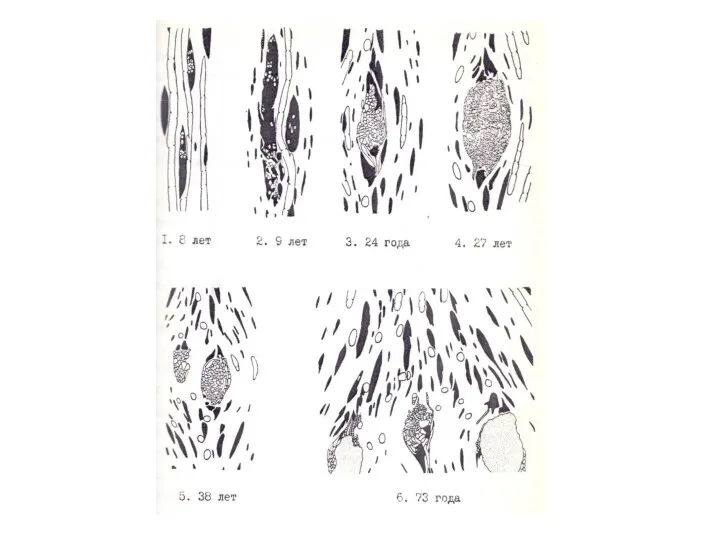

Наши наблюдения и анализ данных литературы показали, что такая последовательность структурных
Наши наблюдения и анализ данных литературы показали, что такая последовательность структурных

Характерной и весьма существенной, с нашей точки зрения, особенностью аномальных участков
Характерной и весьма существенной, с нашей точки зрения, особенностью аномальных участков

Наши наблюдения показали (Коровин, 1970), что во флоэме капов, в зоне
Наши наблюдения показали (Коровин, 1970), что во флоэме капов, в зоне

Заканчивая рассмотрение анатомических изменений, возникающих в древесине при развитии синдрома ямчатости
Заканчивая рассмотрение анатомических изменений, возникающих в древесине при развитии синдрома ямчатости

В заключение данного раздела следует добавить, что структурные неспецифические реакции древесных
В заключение данного раздела следует добавить, что структурные неспецифические реакции древесных

Первыми возникают два взаимно связанных вопроса: какие нарушения регуляции ростовых процессов
Первыми возникают два взаимно связанных вопроса: какие нарушения регуляции ростовых процессов

Здесь же следует добавить, что общепринятые представления о регуляции камбиальной активности
Здесь же следует добавить, что общепринятые представления о регуляции камбиальной активности

Известно, например, что равномерно расположенные по поверхности древесинного цилиндра аномальные зоны
Известно, например, что равномерно расположенные по поверхности древесинного цилиндра аномальные зоны

Величина радиального годичного прироста стволовой древесины достоверно больше средней лишь в
Величина радиального годичного прироста стволовой древесины достоверно больше средней лишь в

Относительная независимость от апикального доминирования наблюдается и в процессе дифференциации производных
Относительная независимость от апикального доминирования наблюдается и в процессе дифференциации производных

Относительно потому, что увеличение поверхности ствола или ветви, происходящее вследствие антиклинальных
Относительно потому, что увеличение поверхности ствола или ветви, происходящее вследствие антиклинальных

Изложенные соображения об относительной независимости ростовых процессов, связанных с камбиальной активностью,
Изложенные соображения об относительной независимости ростовых процессов, связанных с камбиальной активностью,

Идея о решающей роли нарушения баланса ростовых веществ при формировании аномальной
Идея о решающей роли нарушения баланса ростовых веществ при формировании аномальной

Высказывается предположение, что та же причина лежит в основе формирования ложных
Высказывается предположение, что та же причина лежит в основе формирования ложных

В связи с предметом обсуждения представляется интересным тот факт, что среди
В связи с предметом обсуждения представляется интересным тот факт, что среди

Синонимом паренхиматизации во многих случаях можно назвать нарушение дифференциации. Паренхиматизация древесины
Синонимом паренхиматизации во многих случаях можно назвать нарушение дифференциации. Паренхиматизация древесины

Если принять, что скопление паренхимы, возникающее при формировании аномального луча, представляет
Если принять, что скопление паренхимы, возникающее при формировании аномального луча, представляет

Так что же мешает превращению аномального скопления паренхимы в настоящий очаг
Так что же мешает превращению аномального скопления паренхимы в настоящий очаг

Теперь остается высказать наши соображения по поводу ранее поставленных вопросов: как
Теперь остается высказать наши соображения по поводу ранее поставленных вопросов: как

По мнению E. Bünning, и Sagromsky (1948) равномерное распределение лучей можно
По мнению E. Bünning, и Sagromsky (1948) равномерное распределение лучей можно

Многочисленные примеры этого были приведены ранее при рассмотрении особенностей строения конкретных
Многочисленные примеры этого были приведены ранее при рассмотрении особенностей строения конкретных

РОЛЬ ЛУЧЕЙ В ЗАЖИВЛЕНИИ ПОВРЕЖДЕНИЙ СТЕБЛЯ И ДРУГИХ СЛУЧАЯХ ФОРМИРОВАНИЯ
РОЛЬ ЛУЧЕЙ В ЗАЖИВЛЕНИИ ПОВРЕЖДЕНИЙ СТЕБЛЯ И ДРУГИХ СЛУЧАЯХ ФОРМИРОВАНИЯ

Разумеется, мы далеки от отождествления понятий “аномальный рост” и “регенерация”, тем
Разумеется, мы далеки от отождествления понятий “аномальный рост” и “регенерация”, тем

Гартиг обратил внимание на то, что наибольшей способностью к регенерации обладает
Гартиг обратил внимание на то, что наибольшей способностью к регенерации обладает

Гомогенность раневого каллуса сохраняется ограниченное время, вскоре в нем дифференцируется лучевая
Гомогенность раневого каллуса сохраняется ограниченное время, вскоре в нем дифференцируется лучевая

Дедифференциация клеток при нормальном развитии явление не частое. Однако, по мнению
Дедифференциация клеток при нормальном развитии явление не частое. Однако, по мнению

Изучение обнаженных поверхностей показало, что разделение тканей коры и древесины произошло
Изучение обнаженных поверхностей показало, что разделение тканей коры и древесины произошло


Разрастание клеток над раневой поверхностью древесины можно рассматривать как образование первичного
Разрастание клеток над раневой поверхностью древесины можно рассматривать как образование первичного

При проведении подобных исследований необходимо иметь в виду, что данные по
При проведении подобных исследований необходимо иметь в виду, что данные по



Анатомические изменения при зарастании механических повреждений стебля мало отличаются от изменений,
Анатомические изменения при зарастании механических повреждений стебля мало отличаются от изменений,

При слабом повреждении расширяющиеся лучи могут не смыкаться, между ними в
При слабом повреждении расширяющиеся лучи могут не смыкаться, между ними в

По нашим наблюдениям и данным литературы (Воусе, 1923, 1961), процесс зарастания
По нашим наблюдениям и данным литературы (Воусе, 1923, 1961), процесс зарастания

Древесинные лучи принимают участие в локализации и иного рода повреждений. Широко
Древесинные лучи принимают участие в локализации и иного рода повреждений. Широко
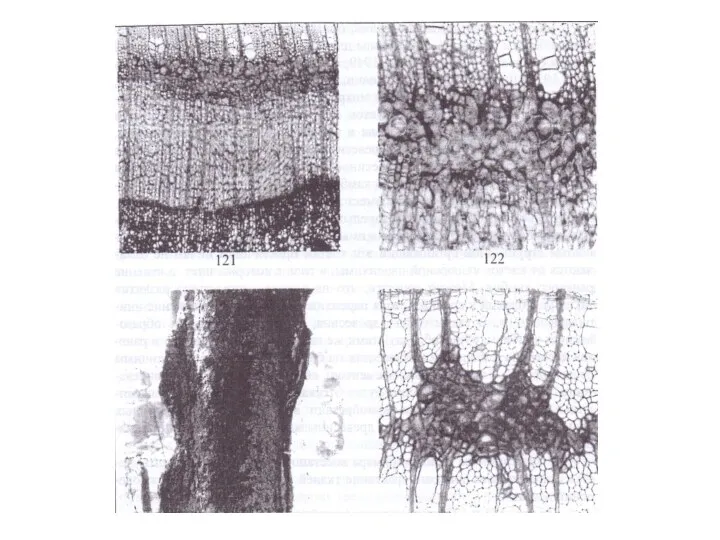

По нашим наблюдениям, аномальная паренхимная прослойка сердцевинных повторений возникает вследствие зарастания
По нашим наблюдениям, аномальная паренхимная прослойка сердцевинных повторений возникает вследствие зарастания

Обобщив результаты наших наблюдении и данные литературы, мы пришли к убеждению,
Обобщив результаты наших наблюдении и данные литературы, мы пришли к убеждению,

В порядке дополнительного подтверждения высказанных соображений приведем несколько примеров из известных
В порядке дополнительного подтверждения высказанных соображений приведем несколько примеров из известных
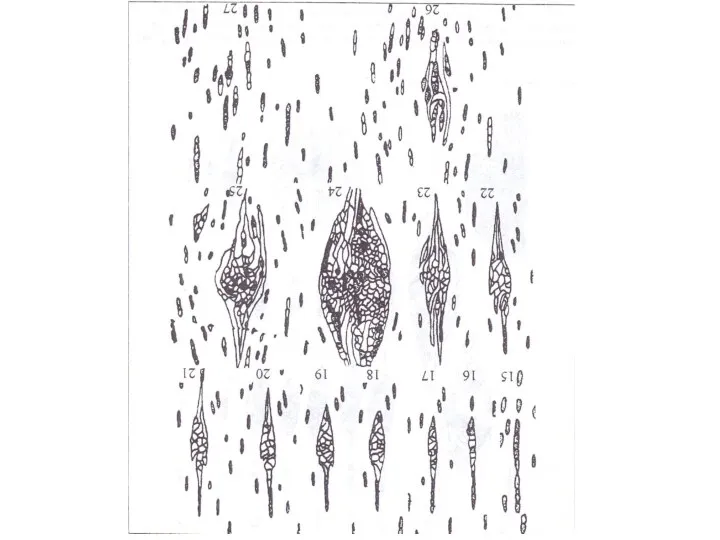


Многие аномальные лучи, достигнув предельных размеров, расчленяются трахеидами на более мелкие,
Многие аномальные лучи, достигнув предельных размеров, расчленяются трахеидами на более мелкие,
 Урок биологии в 7 классе Класс Земноводные (презентация)
Урок биологии в 7 классе Класс Земноводные (презентация) Железы организма и их функции
Железы организма и их функции Кружок Жемчужина. Воспитание бережного отношения к ресурсам растительного мира
Кружок Жемчужина. Воспитание бережного отношения к ресурсам растительного мира Физиология сенсорных систем
Физиология сенсорных систем Проявления генов в развитии. Плейотропное действие гена
Проявления генов в развитии. Плейотропное действие гена Endangered species
Endangered species Кружок Основы молекулярной генетики
Кружок Основы молекулярной генетики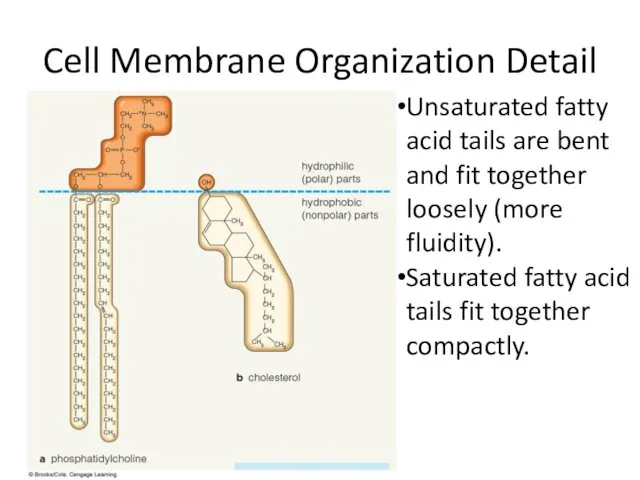 Cell Membrane Organization Detail
Cell Membrane Organization Detail Разнообразие зеленых водорослей
Разнообразие зеленых водорослей Анықталмағандық қатынасы
Анықталмағандық қатынасы Строение и многообразие бактерий
Строение и многообразие бактерий Экскурсия Разнообразие животных в природе
Экскурсия Разнообразие животных в природе Презентация курса Биология в профессии - Практическая биология
Презентация курса Биология в профессии - Практическая биология Нейрофизиология сенсорных систем. Восприятие стимулов. Сенсорная адаптация. Тема 5
Нейрофизиология сенсорных систем. Восприятие стимулов. Сенсорная адаптация. Тема 5 Популяции. Типы пространственной структуры популяций
Популяции. Типы пространственной структуры популяций Основные методы селекции растений, животных, микроорганизмов
Основные методы селекции растений, животных, микроорганизмов Регуляция системной деятельности организма. Функции коры больших полушарий. ЭЭГ
Регуляция системной деятельности организма. Функции коры больших полушарий. ЭЭГ Плоды тыквенные
Плоды тыквенные Ботаника. Салалары
Ботаника. Салалары Паразитизм. Хищничество
Паразитизм. Хищничество Современная биологическая картина мира
Современная биологическая картина мира Сорняки. Однолетние. Многолетние
Сорняки. Однолетние. Многолетние Жизнь муравьев
Жизнь муравьев Порівняння будови кровоносної системи хребетних тварин. Практична робота №4
Порівняння будови кровоносної системи хребетних тварин. Практична робота №4 Кто спит зимой
Кто спит зимой Удод – птица 2016 года
Удод – птица 2016 года Natural selection of human population. Darwin’s theory of evolution by natural selection
Natural selection of human population. Darwin’s theory of evolution by natural selection Клеточная оболочка. Оболочка животных и растительных клеток
Клеточная оболочка. Оболочка животных и растительных клеток