- Биологические мембраны. Структура и функция липидов

Содержание
- 2. - функционально активные поверхностные структуры толщиной в несколько молекулярных слоев, ограничивающие цитоплазму и большинство органелл клетки,
- 3. Биологические мембраны состоят из фосфолипидов, гликолипидов, белков и холестерина. Рассматриваются как белково-липидные комплексы. Все клетки имеют
- 4. Биологические мембраны Любая клетка имеет мембраны. И в каждой эукариотической клетке существуют различные органеллы, каждая из
- 5. отделение клетки от окружающей среды и формирование внутриклеточных компартментов(отсеков); контроль и регулирование транспорта огромного разнообразия веществ
- 6. Плазматическая мембрана Плазматическая мембрана, окружающая каждую клетку, определяет её величину, обеспечивает транспорт малых и больших молекул
- 7. Ядерная мембрана Ядерная оболочка состоит из внешней и внутренней ядерных мембран. Ядерная оболочка имеет поры, через
- 8. Мембрана эндоплазматического ретикулума (ЭР) Мембрана ЭР имеет многочисленные складки и изгибы. Она образует непрерывную поверхность, ограничивающую
- 9. Аппарат Гольджи Аппарат Гольджи - важная мембранная органелла, отвечающая за модификацию, накопление, сортировку и направление различных
- 10. Митохондриальные мембраны Митохондрии - органеллы, окружённые двойной мембраной, специализирующиеся на синтезе АТФ путём окислительного фосфорилирования. Отличительная
- 11. Мембрана лизосом Мембрана лизосом играет роль "щита" между активными ферментами (более 50), обеспечивающими реакции распада белков,
- 12. Основу мембраны составляет двойной липидный слой, в формировании которого участвуют фосфолштиды и гликолипиды. Липидный бислой образован
- 13. Липиды — жироподобные органические соединения, нерастворимые в воде, но хорошо растворимые в неполярных растворителях (эфире, бензине,
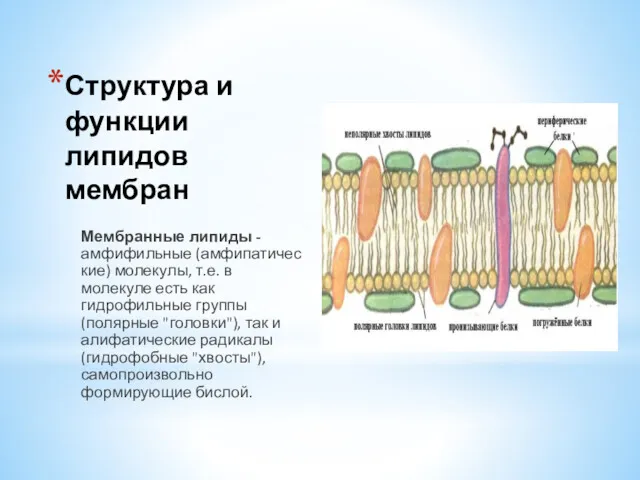
- 14. Структура и функции липидов мембран Мембранные липиды -амфифильные (амфипатические) молекулы, т.е. в молекуле есть как гидрофильные
- 15. Структура липидов
- 16. Фосфолипиды Все фосфолипиды можно разделить на 2 группы - глицерофосфолипиды и сфингофосфолипиды. Глицерофосфолипиды относят к производным
- 17. Гликолипиды В гликолипидах гидрофобная часть представлена церамидом. В зависимости от длины и строения углеводной части различают
- 18. Холестерол. Холестерол присутствует во всех мембранах животных клеток. Его молекула состоит из жёсткого гидрофобного ядра и
- 19. Для мембран характерна жидкостность (текучесть), способность липидов и белков к латеральной диффузии. Скорость перемещения молекул зависит
- 20. Классификация липидов По способности к гидролизу подавляющие большинство липидов можно отнести к омыляемым либо к неомыляемым.
- 21. ТРАНСМЕМБРАННАЯ ПЕРЕДАЧА СИГНАЛА Важное свойство мембран - способность воспринимать и передавать внутрь клетки сигналы из внешней
- 22. Участие рецепторов в трансмембранной передаче сигнала. Рецепторы: 1 - связанные с ионными каналами, например рецептор ГАМК;
- 23. Передача сигнала на внутриклеточные рецепторы. Передача сигнала липидорастворимых стероидных гормонов и тироксина возможна только при прохождении
- 24. Передача сигнала на внутриклеточные рецепторы.
- 26. Скачать презентацию
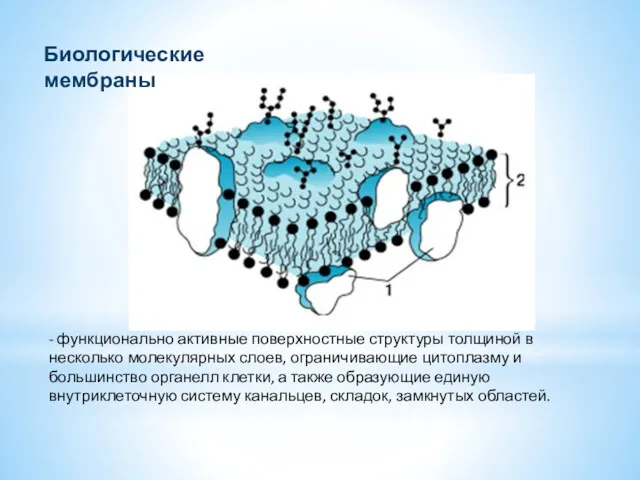
- функционально активные поверхностные структуры толщиной в несколько молекулярных слоев, ограничивающие
- функционально активные поверхностные структуры толщиной в несколько молекулярных слоев, ограничивающие
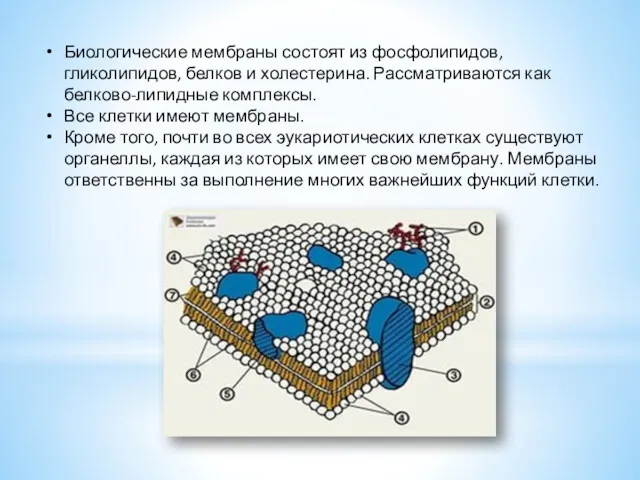
Биологические мембраны состоят из фосфолипидов, гликолипидов, белков и холестерина. Рассматриваются как
Биологические мембраны состоят из фосфолипидов, гликолипидов, белков и холестерина. Рассматриваются как

Биологические мембраны
Любая клетка имеет мембраны. И в каждой эукариотической клетке существуют
Биологические мембраны
Любая клетка имеет мембраны. И в каждой эукариотической клетке существуют

отделение клетки от окружающей среды и формирование внутриклеточных компартментов(отсеков);
контроль и регулирование
отделение клетки от окружающей среды и формирование внутриклеточных компартментов(отсеков);
контроль и регулирование

Плазматическая мембрана
Плазматическая мембрана, окружающая каждую клетку, определяет её величину, обеспечивает транспорт
Плазматическая мембрана
Плазматическая мембрана, окружающая каждую клетку, определяет её величину, обеспечивает транспорт
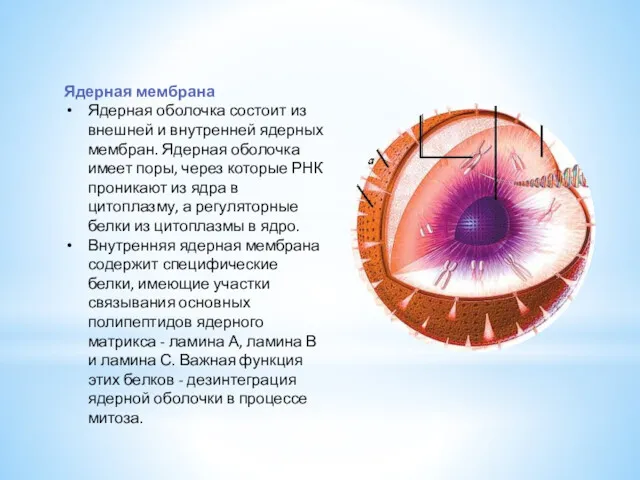
Ядерная мембрана
Ядерная оболочка состоит из внешней и внутренней ядерных мембран. Ядерная
Ядерная мембрана
Ядерная оболочка состоит из внешней и внутренней ядерных мембран. Ядерная
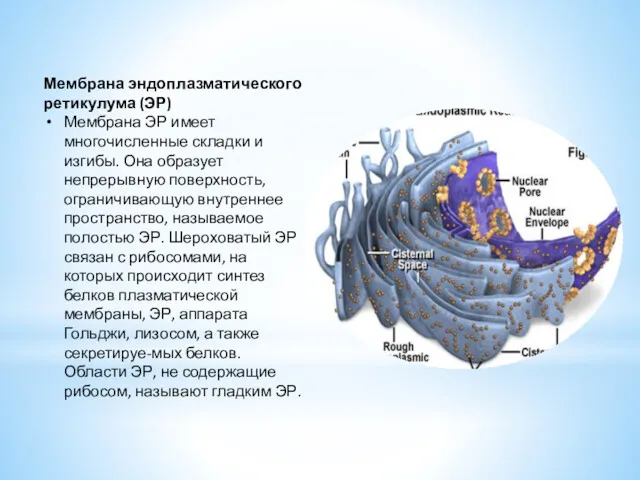
Мембрана эндоплазматического ретикулума (ЭР)
Мембрана ЭР имеет многочисленные складки и изгибы. Она
Мембрана эндоплазматического ретикулума (ЭР)
Мембрана ЭР имеет многочисленные складки и изгибы. Она

Аппарат Гольджи
Аппарат Гольджи - важная мембранная органелла, отвечающая за модификацию, накопление,
Аппарат Гольджи
Аппарат Гольджи - важная мембранная органелла, отвечающая за модификацию, накопление,
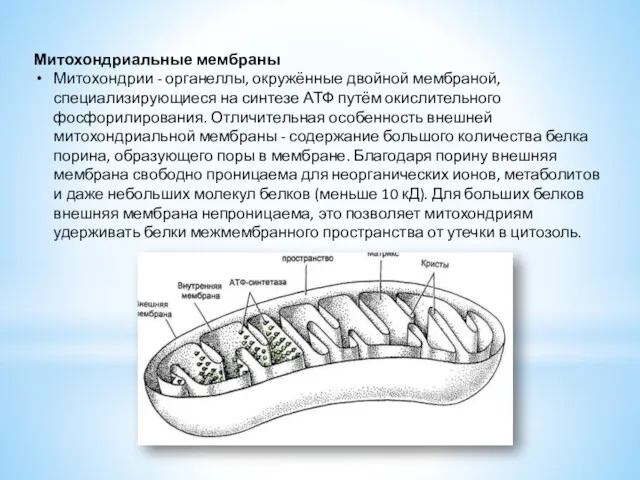
Митохондриальные мембраны
Митохондрии - органеллы, окружённые двойной мембраной, специализирующиеся на синтезе АТФ
Митохондриальные мембраны
Митохондрии - органеллы, окружённые двойной мембраной, специализирующиеся на синтезе АТФ
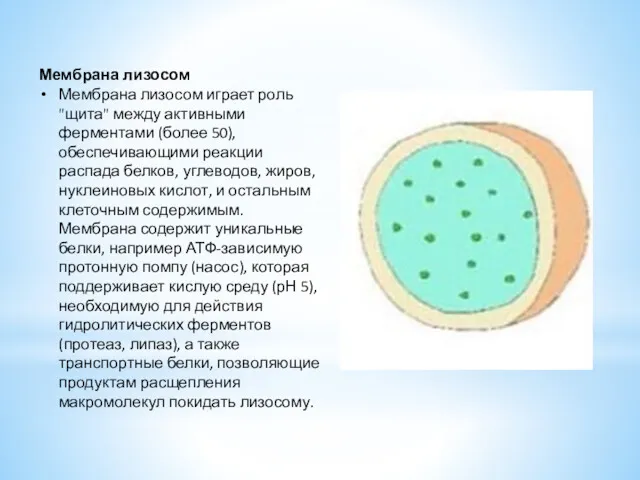
Мембрана лизосом
Мембрана лизосом играет роль "щита" между активными ферментами (более 50),
Мембрана лизосом
Мембрана лизосом играет роль "щита" между активными ферментами (более 50),
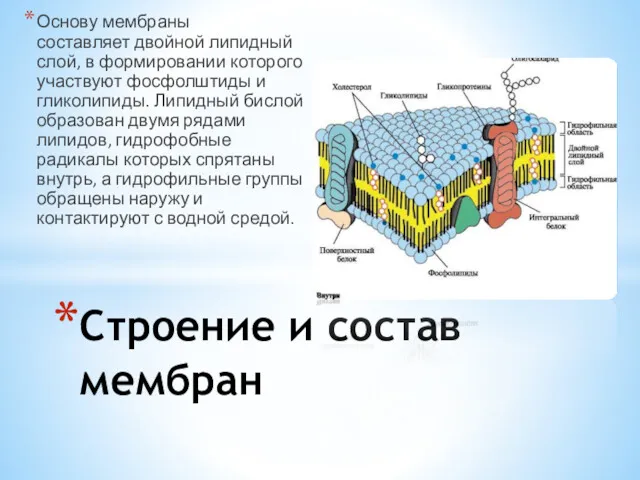
Основу мембраны составляет двойной липидный слой, в формировании которого участвуют фосфолштиды и гликолипиды.
Основу мембраны составляет двойной липидный слой, в формировании которого участвуют фосфолштиды и гликолипиды.
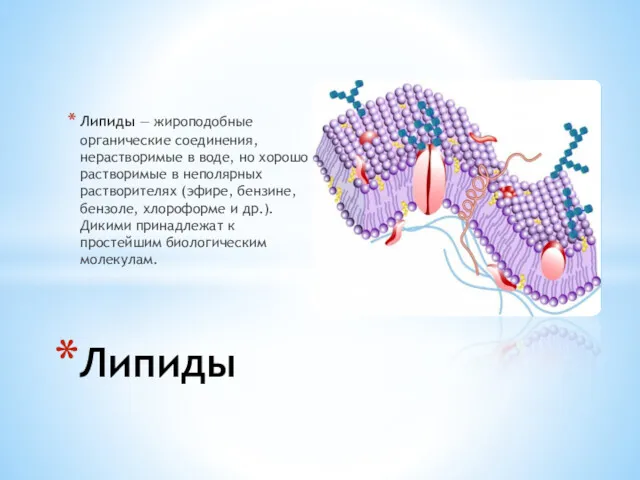
Липиды — жироподобные органические соединения, нерастворимые в воде, но хорошо растворимые в
Липиды — жироподобные органические соединения, нерастворимые в воде, но хорошо растворимые в
Структура и функции липидов мембран
Мембранные липиды -амфифильные (амфипатические) молекулы, т.е. в молекуле есть как
Структура и функции липидов мембран
Мембранные липиды -амфифильные (амфипатические) молекулы, т.е. в молекуле есть как
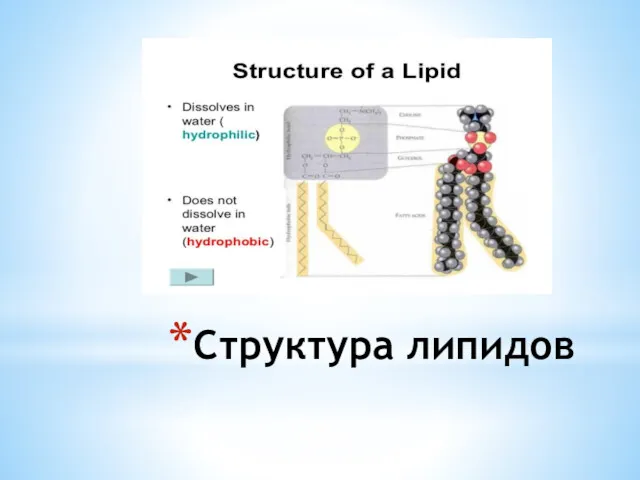
Структура липидов
Структура липидов
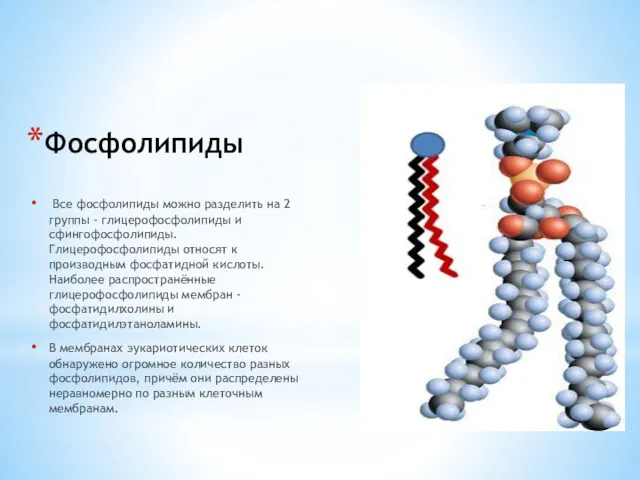
Фосфолипиды
Все фосфолипиды можно разделить на 2 группы - глицерофосфолипиды и сфингофосфолипиды.
Фосфолипиды
Все фосфолипиды можно разделить на 2 группы - глицерофосфолипиды и сфингофосфолипиды.

Гликолипиды
В гликолипидах гидрофобная часть представлена церамидом. В зависимости от длины и
Гликолипиды
В гликолипидах гидрофобная часть представлена церамидом. В зависимости от длины и

Холестерол.
Холестерол присутствует во всех мембранах животных клеток. Его молекула состоит из
Холестерол.
Холестерол присутствует во всех мембранах животных клеток. Его молекула состоит из

Для мембран характерна жидкостность (текучесть), способность липидов и белков к латеральной
Для мембран характерна жидкостность (текучесть), способность липидов и белков к латеральной

Классификация липидов
По способности к гидролизу подавляющие большинство липидов можно отнести к
Классификация липидов По способности к гидролизу подавляющие большинство липидов можно отнести к

ТРАНСМЕМБРАННАЯ ПЕРЕДАЧА СИГНАЛА
Важное свойство мембран - способность воспринимать и передавать внутрь
ТРАНСМЕМБРАННАЯ ПЕРЕДАЧА СИГНАЛА
Важное свойство мембран - способность воспринимать и передавать внутрь
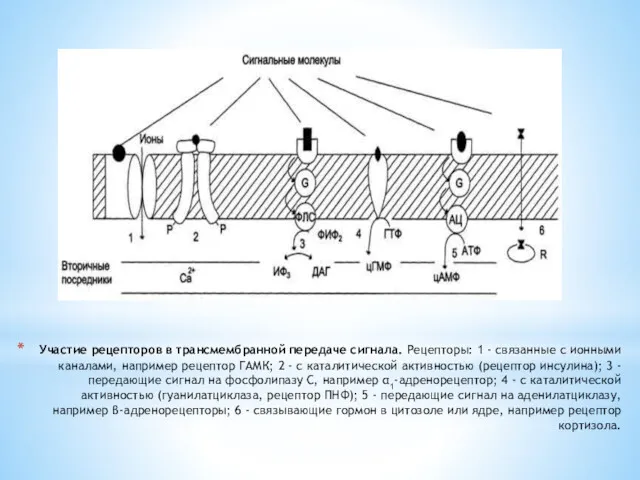
Участие рецепторов в трансмембранной передаче сигнала. Рецепторы: 1 - связанные с ионными
Участие рецепторов в трансмембранной передаче сигнала. Рецепторы: 1 - связанные с ионными

Передача сигнала на внутриклеточные рецепторы.
Передача сигнала липидорастворимых стероидных гормонов и
Передача сигнала на внутриклеточные рецепторы.
Передача сигнала липидорастворимых стероидных гормонов и
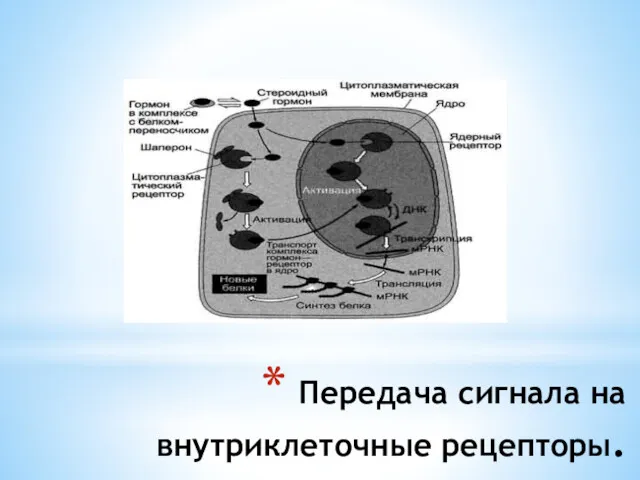
Передача сигнала на внутриклеточные рецепторы.
Передача сигнала на внутриклеточные рецепторы.
 Презентация к уроку биология 10 класс Питание клетки.
Презентация к уроку биология 10 класс Питание клетки. Царство Животные
Царство Животные Женская половая система
Женская половая система Экология сообществ (синэкология)
Экология сообществ (синэкология) Витамины, их биологическая роль и влияние на организм
Витамины, их биологическая роль и влияние на организм Биоценоз және Биогеоценоз
Биоценоз және Биогеоценоз Скелет туловища, его развитие в фило- и онтогенезе
Скелет туловища, его развитие в фило- и онтогенезе АСҚ пайдаланылу жолдары
АСҚ пайдаланылу жолдары Дигибридное скрещивание
Дигибридное скрещивание Ознакомление с фитонцидными растениями и выявление возможности их использования в интерьере. Практическая работа
Ознакомление с фитонцидными растениями и выявление возможности их использования в интерьере. Практическая работа 20230923_driopiteki
20230923_driopiteki Бактерии в организме человека
Бактерии в организме человека Ткани растений и животных
Ткани растений и животных Презентация Бактерии в нашей жизни
Презентация Бактерии в нашей жизни Наследственный аппарат клетки
Наследственный аппарат клетки Закон Менделя
Закон Менделя Регуляция пищеварения. Гигиена питания
Регуляция пищеварения. Гигиена питания Царство Растения
Царство Растения Вегетативное обеспечение мышечной работы
Вегетативное обеспечение мышечной работы Деление клетки – основа размножения, роста и развития организмов
Деление клетки – основа размножения, роста и развития организмов История развития генетики
История развития генетики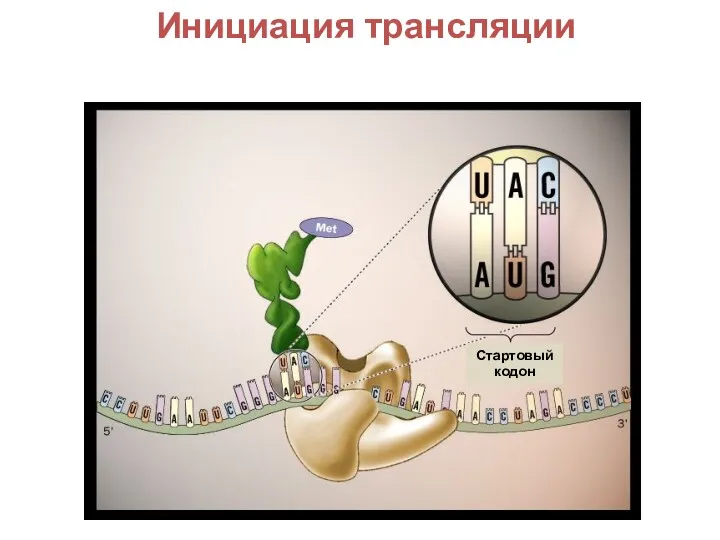 Инициация трансляции в генетике. (Лекция 18)
Инициация трансляции в генетике. (Лекция 18) Белый медведь
Белый медведь презентация к уроку биологии в 7 классе по теме Птицы
презентация к уроку биологии в 7 классе по теме Птицы Як бачать тварини
Як бачать тварини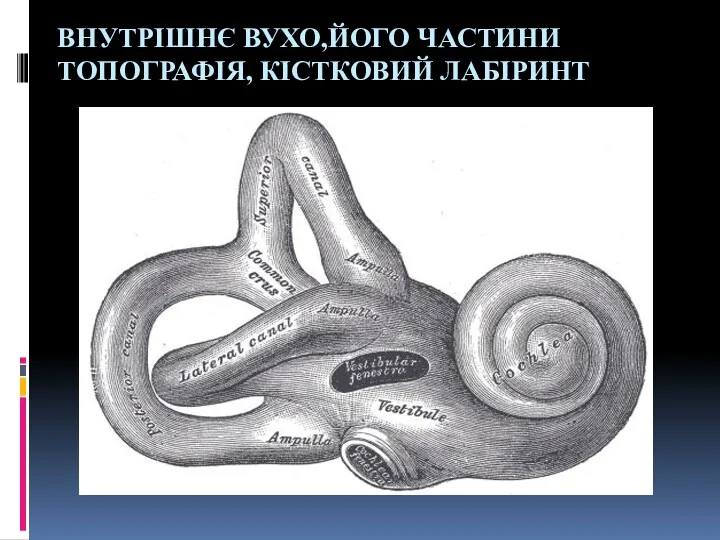 Внутрішнє вухо, його частини, топографія, кістковий лабіринт
Внутрішнє вухо, його частини, топографія, кістковий лабіринт Структура ЕГЭ по биологии
Структура ЕГЭ по биологии Обмен углеводов
Обмен углеводов