- Биотехнология. Генетические векторы для клонирования ДНК

Содержание
- 2. ГЕНЕТИЧЕСКИЕ ВЕКТОРЫ ДЛЯ КЛОНИРОВАНИЯ ДНК
- 3. В настоящее время существует два подхода для получения необходимого количества конкретного участка исследуемой ДНК: 1. Клонирование
- 4. Для клонирования ДНК in vivo нужно: ДНК-вставка (ДНК-мишень, ДНК-интереса); ДНК-вектор (генетический вектор клонирования); клетка-хозяин; набор ферментов-инструментов
- 5. В основе молекулярного клонирования лежит встраивание нужного фрагмента ДНК (вставки) в другую молекулу ДНК (вектор), которая
- 6. Вектор для переноса генетической информации должен удовлетворять ряду основных требований: 1. Способность к автономной репликации, т.е.
- 7. ПЛАЗМИДНЫЕ ВЕКТОРЫ Плазмиды – внехромосомные автономнореплицирующиеся двухцепочечные кольцевые молекулы ДНК. Плазмидные векторы создают на основе природных
- 8. Вектор клонирования pBR322 был создан Боливаром и Родригесом в 80-тые годы прошлого века – один из
- 9. Фенотипические свойства клеток, содержащих плазмиду pBR322 со вставками и без них
- 11. Трансформация – процесс введения рекомбинантной ДНК в бактериальную клетку. Клетка-хозяин должна иметь определенный фенотип: r ,
- 12. Частота трансформации – доля клеток в клеточной популяции, получивших чужеродную ДНК; выражается числом трансформантов к общему
- 13. Селективный отбор на основании присутствия –отсутствия резистентности к определенному антибиотику
- 14. Эпоха вектора pBR322, начатая Боливаром и Родригесом в самом начале 80-тых годов ХХ-го столетия, продолжается и
- 15. Группа векторов семейства pUC “unicom cloning” – обозначает наличие в структуре ДНК полилинкера, который представляет собой
- 16. Плазмидный вектор pUC19 pUC19 содержит: -ген устойчивости к ампициллину (Amp); -регулируемый сегмент гена β-галактозидазы (lacZ) лактозного
- 17. ИПТГ (изопропил-β-D-тиогалактопиранозид) – индуктор lac – оперона. В его присутствии продукт гена lacI не сможет связаться
- 19. ВЕКТОРЫ ДЛЯ КЛОНИРОВАНИЯ КРУПНЫХ ФРАГМЕНТОВ ДНК С помощью плазмидных векторов можно клонировать фрагменты ДНК до 10
- 20. Векторы на основе бактериофага λ. Два альтернативных пути развития событий при проникновении фага λ в клетки
- 21. Литический путь развития фага λ 1. При репликации кольцевой ДНК бактериофага λ образуется линейная молекула, состоящая
- 22. Литический путь развития бактериофага λ
- 23. Рекомбинантные молекулы упаковывают в головки бактериофага λ in vitro, и после добавления отростков получают инфекционные фаговые
- 24. Приготовление экстрактов для осуществления упаковки in vitro ДНК фага λ. Используют два штамма E.coli, каждый из
- 25. Упаковка in vitro ДНК фага λ
- 26. Процесс введения рекомбинантной фаговой ДНК со встроенным фрагментом чужеродной генетической информации в клетки-реципиенты основан на естественном
- 27. Космиды – один из видов гибридных векторов, которые реплицируются, используя плазмидный тип репликации, и обладают способностью
- 28. Векторные системы для клонирования очень крупных фрагментов ДНК (вставки >100 т.п.н.) имеют большую ценность при анализе
- 29. Клонирование фрагментов ДНК большого размера с помощью ВАС
- 30. ВАС - стабильный вектор, способный интегрировать вставки длиной от 150 до 350 т. п. н. Трансформация
- 31. СРЕДСТВА ДОСТАВКИ РЕКОМБИНАНТНЫХ ДНК В РЕЦИПИЕНТНЫЕ КЛЕТКИ В настоящее время известно около 40 различных способов доставки
- 32. ТРАНСФОРМАЦИЯ Трансформация в самом общем значении – это процесс введения свободной ДНК в клетку. В более
- 33. Часто используется этап подготовки компетентных клеток: получение сферопластов – клеток, частично или полностью (протопласты)лишенных наружной ригидной
- 34. Конъюгация Существуют бактериальные плазмиды (конъюгативные плазмиды), обладающие способностью создавать межклеточные контакты, через которые они и переходят
- 35. Вирусная инфекция. Для внедрения векторов на основе вирусов широко используется природный инфекционный путь заражения клетки-хозяина, который
- 36. Трансфекция Одним из первых разработанных методов эффективной трансфекции была инкубация ДНК с ДЕАЕ-декстраном. Механизм действия ДЕАЕ-декстрана
- 37. Полимерные катионы образуют с нуклеиновыми кислотами устойчивые комплексы с нейтрализованными зарядами, которые могут с высокой эффективностью
- 38. ЛИПОСОМЫ (ДОТМА (1,2-диолеил-3-N,N,N-триметиламинопропан) Фелгнером (Felgner, 1987) Структура комплекса с ДНК (а) и общая структура катионного липидного
- 39. Микроинъекция Клеточная мембрана прокалывается микроиглой и раствор, содержащий ДНК, вводится в цитоплазму клетки или напрямую в
- 40. Баллистическая трансфекция (биобаллистика, или биолистика) Обстрел клеток микросферами размером около 1-2 мкм, покрытых ДНК. Применяются микрочастицы
- 42. Скачать презентацию

ГЕНЕТИЧЕСКИЕ ВЕКТОРЫ ДЛЯ КЛОНИРОВАНИЯ ДНК
ГЕНЕТИЧЕСКИЕ ВЕКТОРЫ ДЛЯ КЛОНИРОВАНИЯ ДНК

В настоящее время существует два подхода для получения необходимого количества конкретного

Для клонирования ДНК in vivo нужно:
ДНК-вставка (ДНК-мишень, ДНК-интереса);
ДНК-вектор (генетический вектор клонирования);
клетка-хозяин;
набор
ДНК-вставка (ДНК-мишень, ДНК-интереса);
ДНК-вектор (генетический вектор клонирования);
клетка-хозяин;
набор

В основе молекулярного клонирования лежит встраивание нужного фрагмента ДНК (вставки) в
В основе молекулярного клонирования лежит встраивание нужного фрагмента ДНК (вставки) в

Вектор для переноса генетической информации должен удовлетворять ряду основных требований:
1. Способность
1. Способность

ПЛАЗМИДНЫЕ ВЕКТОРЫ
Плазмиды – внехромосомные автономнореплицирующиеся двухцепочечные кольцевые молекулы ДНК.
Плазмидные векторы создают
Плазмиды – внехромосомные автономнореплицирующиеся двухцепочечные кольцевые молекулы ДНК.
Плазмидные векторы создают
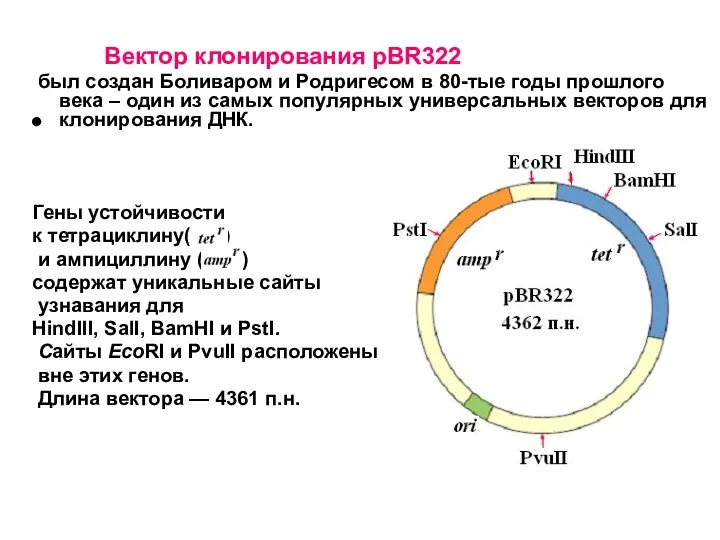
Вектор клонирования pBR322
был создан Боливаром и Родригесом в 80-тые
Вектор клонирования pBR322
был создан Боливаром и Родригесом в 80-тые
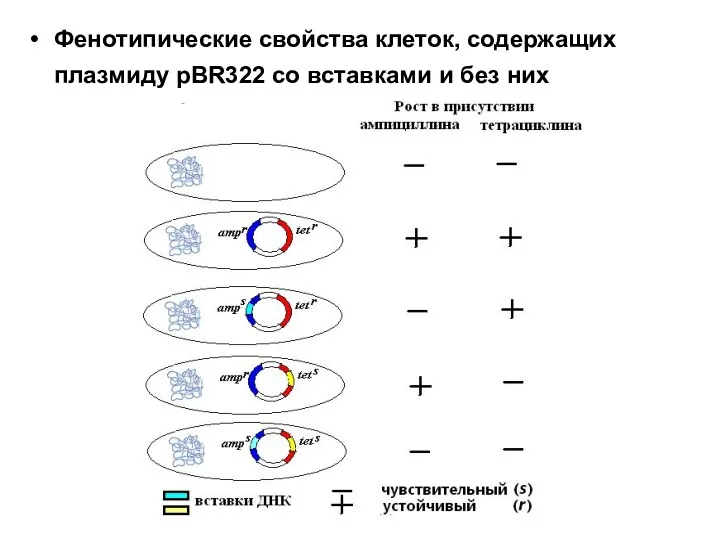
Фенотипические свойства клеток, содержащих плазмиду pBR322 со вставками и без них
Фенотипические свойства клеток, содержащих плазмиду pBR322 со вставками и без них
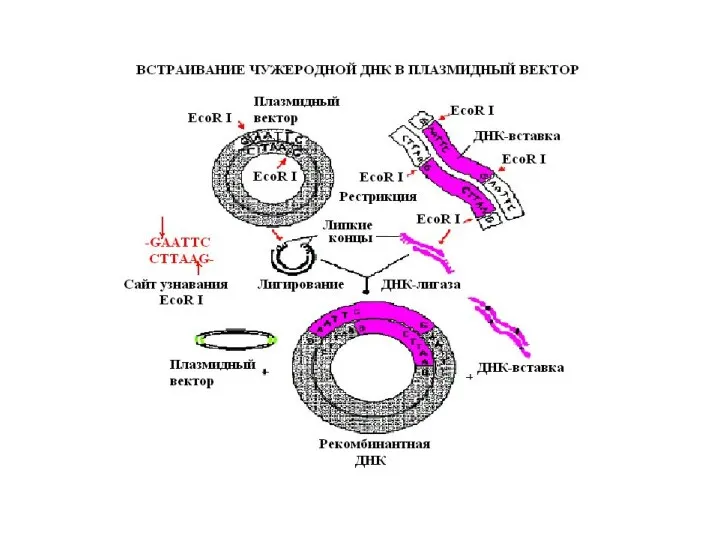

Трансформация – процесс введения рекомбинантной ДНК в бактериальную клетку.
Клетка-хозяин должна
Клетка-хозяин должна

Частота трансформации – доля клеток в клеточной популяции, получивших чужеродную ДНК;
Частота трансформации – доля клеток в клеточной популяции, получивших чужеродную ДНК;
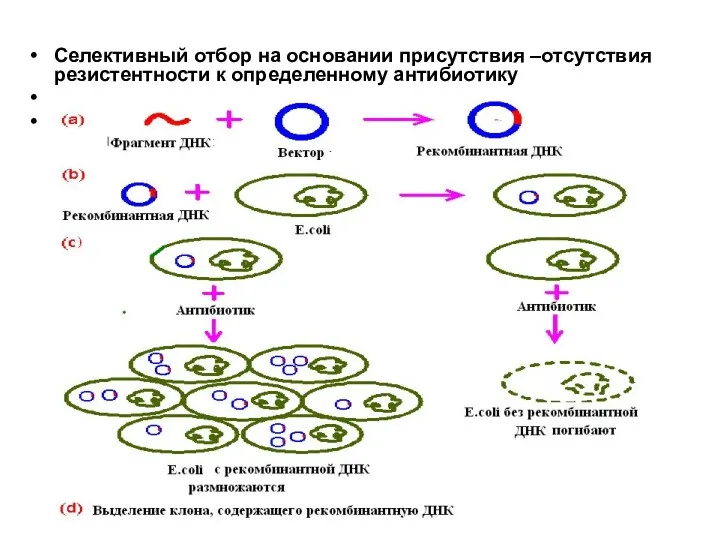
Селективный отбор на основании присутствия –отсутствия резистентности к определенному антибиотику

Эпоха вектора pBR322, начатая Боливаром и Родригесом в самом начале 80-тых
Эпоха вектора pBR322, начатая Боливаром и Родригесом в самом начале 80-тых

Группа векторов семейства pUC
“unicom cloning” – обозначает наличие в
Группа векторов семейства pUC
“unicom cloning” – обозначает наличие в
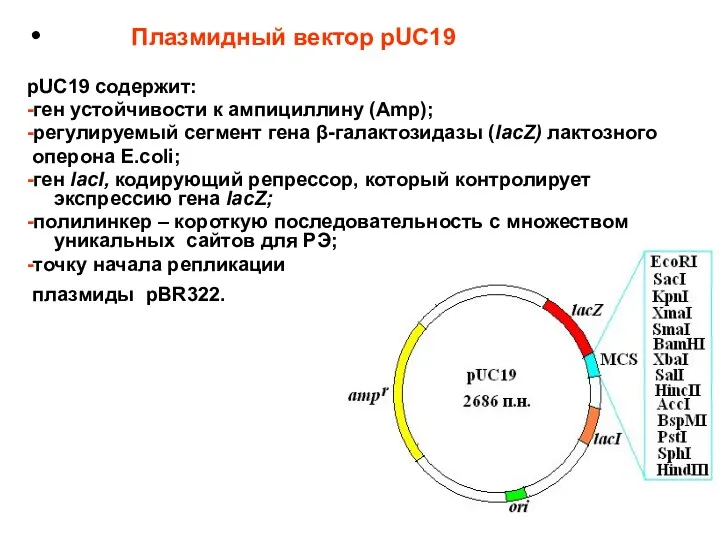
Плазмидный вектор pUC19
pUC19 содержит:
-ген устойчивости к ампициллину (Amp);
-регулируемый сегмент
Плазмидный вектор pUC19
pUC19 содержит:
-ген устойчивости к ампициллину (Amp);
-регулируемый сегмент

ИПТГ (изопропил-β-D-тиогалактопиранозид) – индуктор lac – оперона. В его присутствии продукт
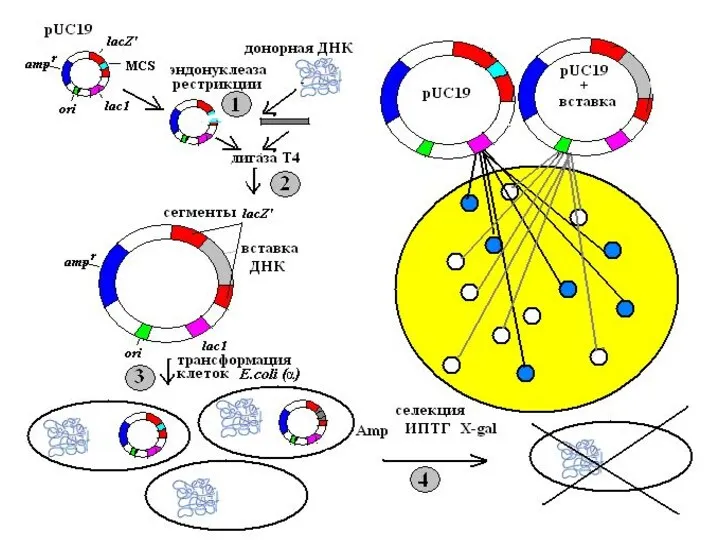

ВЕКТОРЫ ДЛЯ КЛОНИРОВАНИЯ КРУПНЫХ ФРАГМЕНТОВ ДНК
С помощью плазмидных векторов можно
С помощью плазмидных векторов можно

Векторы на основе бактериофага λ.
Два альтернативных пути развития событий при проникновении
Два альтернативных пути развития событий при проникновении

Литический путь развития фага λ
1. При репликации кольцевой
Литический путь развития фага λ
1. При репликации кольцевой
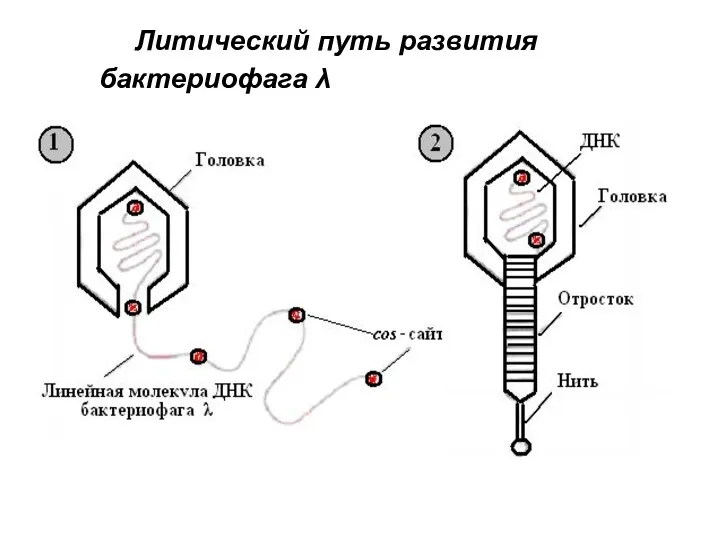
Литический путь развития бактериофага λ
Литический путь развития бактериофага λ
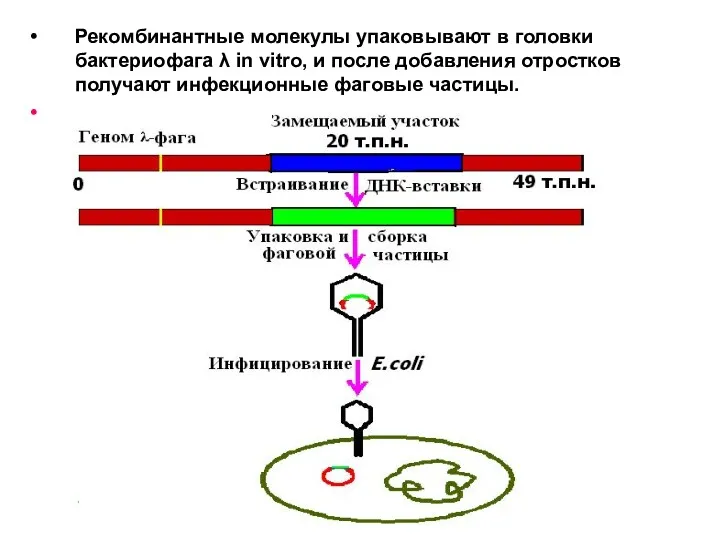
Рекомбинантные молекулы упаковывают в головки бактериофага λ in vitro, и после
Рекомбинантные молекулы упаковывают в головки бактериофага λ in vitro, и после

Приготовление экстрактов для осуществления упаковки in vitro ДНК фага λ.
Используют два
Приготовление экстрактов для осуществления упаковки in vitro ДНК фага λ.
Используют два
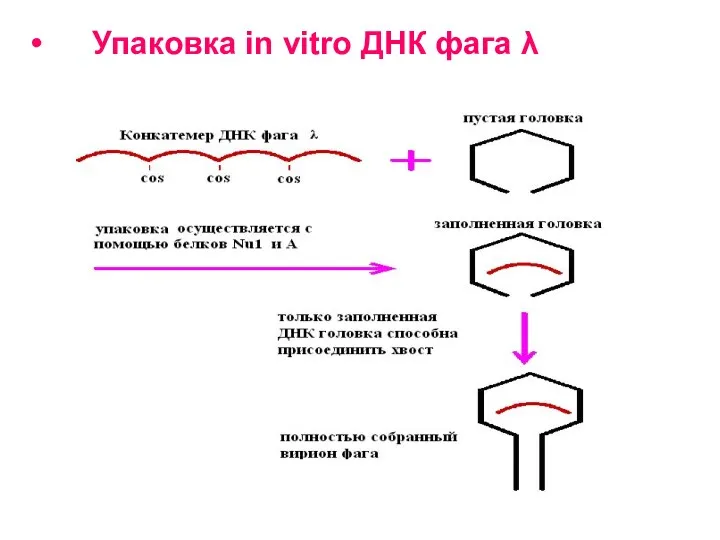
Упаковка in vitro ДНК фага λ
Упаковка in vitro ДНК фага λ

Процесс введения рекомбинантной фаговой ДНК со встроенным фрагментом чужеродной генетической информации

Космиды – один из видов гибридных векторов, которые реплицируются, используя плазмидный

Векторные системы для клонирования очень крупных фрагментов ДНК (вставки >100 т.п.н.)
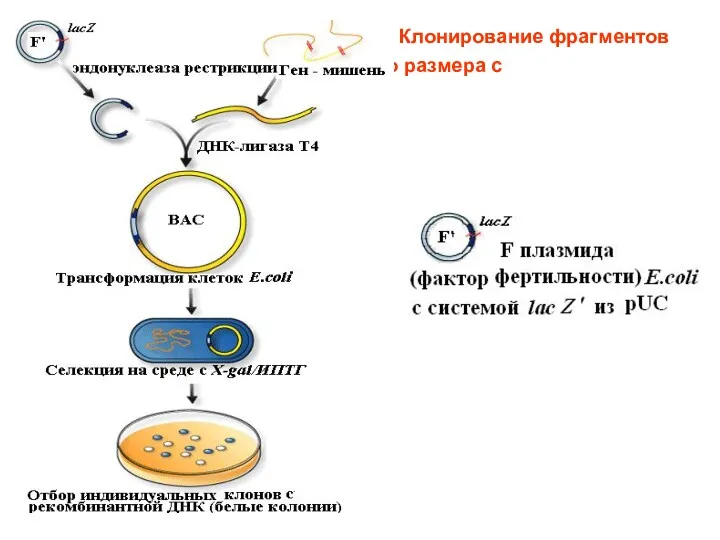
Клонирование фрагментов
ДНК большого размера с помощью ВАС
Клонирование фрагментов
ДНК большого размера с помощью ВАС

ВАС - стабильный вектор, способный интегрировать вставки длиной от 150 до
ВАС - стабильный вектор, способный интегрировать вставки длиной от 150 до

СРЕДСТВА ДОСТАВКИ РЕКОМБИНАНТНЫХ ДНК В РЕЦИПИЕНТНЫЕ КЛЕТКИ
В настоящее время известно около
СРЕДСТВА ДОСТАВКИ РЕКОМБИНАНТНЫХ ДНК В РЕЦИПИЕНТНЫЕ КЛЕТКИ
В настоящее время известно около

ТРАНСФОРМАЦИЯ
Трансформация в самом общем значении – это процесс введения свободной ДНК
Трансформация в самом общем значении – это процесс введения свободной ДНК

Часто используется этап подготовки компетентных клеток: получение сферопластов – клеток, частично
Часто используется этап подготовки компетентных клеток: получение сферопластов – клеток, частично

Конъюгация
Существуют бактериальные плазмиды (конъюгативные плазмиды), обладающие способностью создавать межклеточные контакты,
через
Конъюгация
Существуют бактериальные плазмиды (конъюгативные плазмиды), обладающие способностью создавать межклеточные контакты,
через

Вирусная инфекция.
Для внедрения векторов на основе вирусов широко используется природный инфекционный
Вирусная инфекция.
Для внедрения векторов на основе вирусов широко используется природный инфекционный
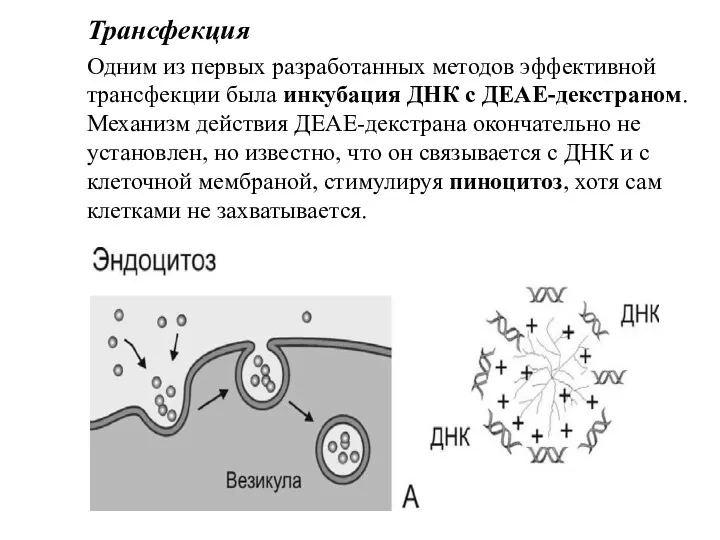
Трансфекция
Одним из первых разработанных методов эффективной трансфекции была инкубация ДНК
Трансфекция
Одним из первых разработанных методов эффективной трансфекции была инкубация ДНК
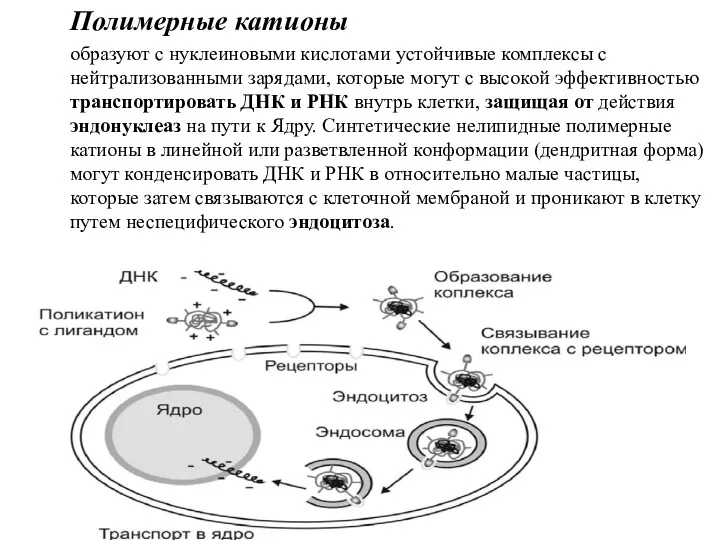
Полимерные катионы
образуют с нуклеиновыми кислотами устойчивые комплексы с нейтрализованными зарядами, которые
Полимерные катионы
образуют с нуклеиновыми кислотами устойчивые комплексы с нейтрализованными зарядами, которые
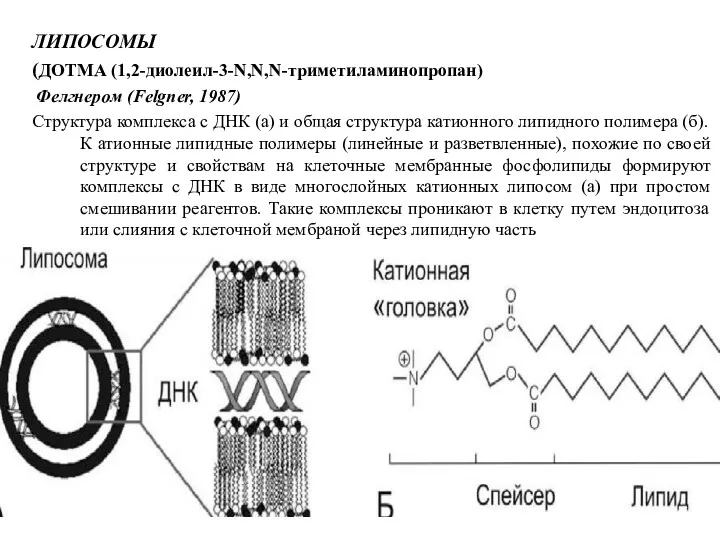
ЛИПОСОМЫ
(ДОТМА (1,2-диолеил-3-N,N,N-триметиламинопропан)
Фелгнером (Felgner, 1987)
Структура комплекса с ДНК (а) и
ЛИПОСОМЫ
(ДОТМА (1,2-диолеил-3-N,N,N-триметиламинопропан)
Фелгнером (Felgner, 1987)
Структура комплекса с ДНК (а) и
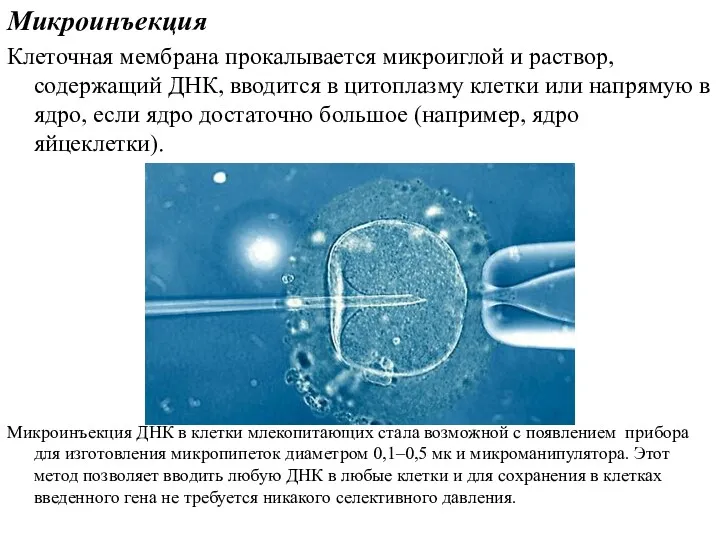
Микроинъекция
Клеточная мембрана прокалывается микроиглой и раствор, содержащий ДНК, вводится в
Микроинъекция
Клеточная мембрана прокалывается микроиглой и раствор, содержащий ДНК, вводится в
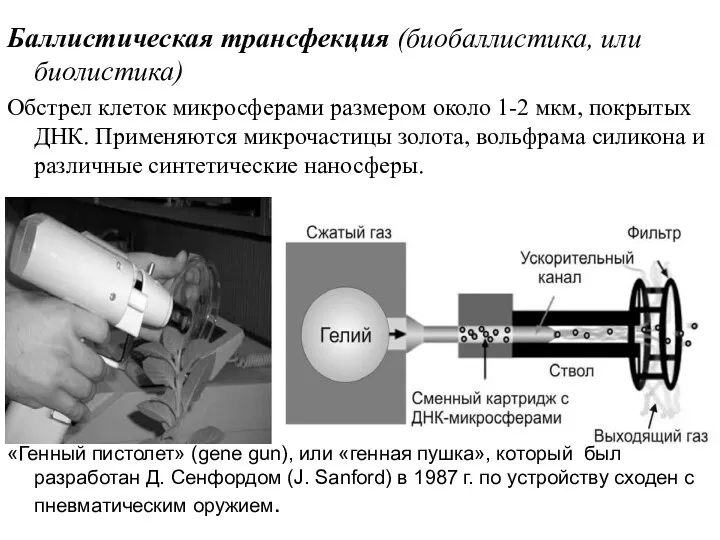
Баллистическая трансфекция (биобаллистика, или биолистика)
Обстрел клеток микросферами размером около 1-2 мкм,
Баллистическая трансфекция (биобаллистика, или биолистика)
Обстрел клеток микросферами размером около 1-2 мкм,
 Железы организма и их функции
Железы организма и их функции The Elementary Structure of the Population
The Elementary Structure of the Population Життя підводних мешканців
Життя підводних мешканців Ферментті препараттар өндіру
Ферментті препараттар өндіру Обработка почвы, основные приемы ухода за растениями
Обработка почвы, основные приемы ухода за растениями К уроку по окружающему миру в 4-м классе
К уроку по окружающему миру в 4-м классе Семейство Лилейные
Семейство Лилейные Участие прокариот в превращениях соединений биогенных элементов
Участие прокариот в превращениях соединений биогенных элементов Презентация Клеточное строение листа Диск
Презентация Клеточное строение листа Диск Развитие жизни на Земле. История Земли
Развитие жизни на Земле. История Земли игра Что Где Когда по биологии
игра Что Где Когда по биологии Влияние сельского хозяйства на биосферу
Влияние сельского хозяйства на биосферу История органического мира земли
История органического мира земли Отряд Парнокопытные
Отряд Парнокопытные Мейоз. Клетки организма
Мейоз. Клетки организма Викторина по биологии Что? Где? Когда?
Викторина по биологии Что? Где? Когда? Обмен белков (часть 1)
Обмен белков (часть 1) Бактериологиялық бояулардың түрлерімен танысу
Бактериологиялық бояулардың түрлерімен танысу Энергетика мышечного сокращения. Ферменты в толстых нитях поперечно-полосатых мышц позвоночных
Энергетика мышечного сокращения. Ферменты в толстых нитях поперечно-полосатых мышц позвоночных Интересное животное Африки - оцелот
Интересное животное Африки - оцелот растения
растения Sleep for life
Sleep for life Потенциал действия
Потенциал действия Насекомые вредные и полезные
Насекомые вредные и полезные Основы селекции. Работы Н.И. Вавилова
Основы селекции. Работы Н.И. Вавилова Размножение иразвтие организмов
Размножение иразвтие организмов Биологическое, психическое и социальное в человеке
Биологическое, психическое и социальное в человеке Вода как экологический фактор
Вода как экологический фактор