- ДНҚ молекуласының құрылымы

Содержание
- 2. ДНҚ молекуласының құрылымы; Жасушадағы ДНҚ полинуклеотидтік тізбегі мынандай нуклеотидттерден d-АМФ, d-ГМФ, d-ЦТФ, d-ТМФ, т.б. құрамында дезоксирибоза
- 3. Молекула ДНК - две комплиментарные полинуклеотидные цепи
- 5. Чаргафф ережесі: ДНҚ молекуласында: 1. Аденин саны Тиминге,Гуанин саны Цитозинге тең. ( А=Т, Г=Ц) 6-жағдайындағы кето
- 6. Генетикалық код құрылымы: 1. Триплет-ДНҚ құрамындағы полинуклеотидтік тізбектің кодтық элементі,сәйкес келетін аминқышкылдары арқылы полипеитидтік тізбекті анықтайды.
- 7. Триплетный код
- 8. ГЕН Қазіргі уақытта ген терминінің түпнегізінде ДНҚ жатыр, в котором в триплетной последовательности закодирована информация на
- 9. РНК құрылысы РНҚ-ның ДНҚ айырмашылғы көп бөлігінде жалғыз тізбекті болып келеді. РНҚ түрлері- тасымалдаушы (тРНК), рибосомалық(рРНК)
- 10. мРНК құрылысы Матрицалық ДНК нмесе ақпараттық ДНК барлық РНҚ құрамының 3-5 % құрайды. Бұл бір тізбекті
- 11. Рибосомалық РНҚ Рибосома массасының 80% р-РНҚ-дан тұрады,ядрошықта жоғары метилденген рибосомалық РНҚ-ның рибосомалық белоктармен бірге жинақталуы жүреді.
- 12. Тасымалдаушы т-РНК РНК-ның 15 пайызға жуық бөлігін алып жатыр. Т-РНК ең қысқа полинуклеотидті тізбекке ие,яғни 80
- 13. Т-РНК құрылымы Т-РНК келесі құрылымдық бөліктерден тұрады: 1.Акцепторлы иіні-жұптасқан нуклеотидтер сабағынан тұрады және ол ССА реттілігімен
- 15. ДНҚ-ның матрицалық синтезі: Днқ-ның екі еселену процессін-репликация деп атайды. (редупликация). ДНҚ-екі еселенуі ол-жіпшелердің ажырауы нәтижесінде,және әрбір
- 16. ДНҚ-ның репликациясы мен репарациясының молекулалық негізі. ДНҚ репликациясына міндетті түрде қажет: 1.Жаңа тізбекті синтездеу үшін: d-АТФ
- 17. ферменттер: 1. Топоизомераза – “репликативтiк айырдың” алдындағы ДНҚ-ның аса жоғары ширатылған жерлерiн босатады 2.Геликаза - ДНҚ
- 18. ДНҚ синтезінде мынандай қасиеттерге ие,антипаралельдік және унирполярлық. Әрбір ДНҚ тізбегі белгілі бағытқа ие болады.дезоксирибозаның 3‘-ОН тобымен
- 19. Днк плимеразаны артта қалған деп атайды себебі 3' 5‘ қозғала алады. Днқ полимераза қор ретінде және
- 20. ДНК синтезі. Днқ биосинтезі схемасы: m(dАТФ+dТТФ)+ n(dГТФ+dЦТФ)------>ДНК + (m+n)Н4Р2О7 Бұл процесстің маңызды ерекшеліктері болып табылады: 1-
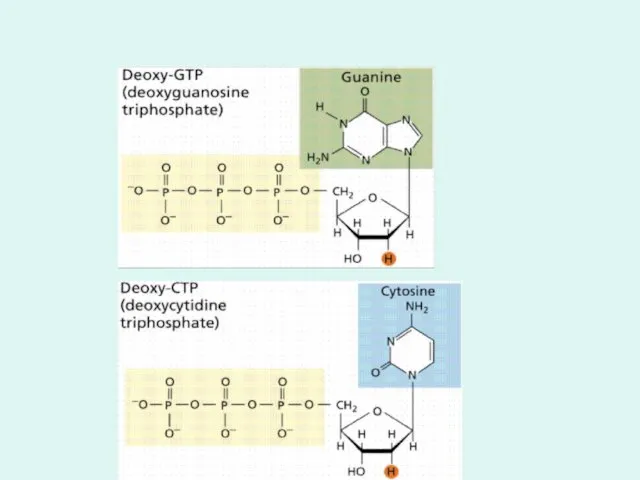
- 21. Дезокси-АТФ
- 23. Днқ қос тізбегі өте мықты шиыршықталған,оны шешу үшін ерекше белоктар ферменттер қажет. Арнайы фермент днқ-хеликаза бір
- 25. Днқ тізбегіндегі инициация процессі. Днқ полимераза днқ синтезін матрицада бастай алмайды,себебі ол тек 3‘-бар полинуклеотидті тізбекке
- 26. РНҚ праймерлерінің үзіліп түсіп қалуы,ДНҚ жіпшесінің жаңа синтезделген жіпшесі 10-20 нуклеотидке қысқарады. Осы жерде Репликацияның толық
- 27. ДНК синтезі -"праймерлер". (5` 3 `бағытындағы синтез. ) Хеликаза ферментінің әсерінен қос жіпшелер арасында сутектік байланыстар
- 29. Праймерлерге ДНҚ-полимераза арқасында сәйкес дезоксиринукеозидтер 5` 3` бағытында жалғанады. Нәтижесінде ДНҚ матрицалық полинуклеотидті тізбегіне,айырылмайтын гибридтік тізбек,яғни
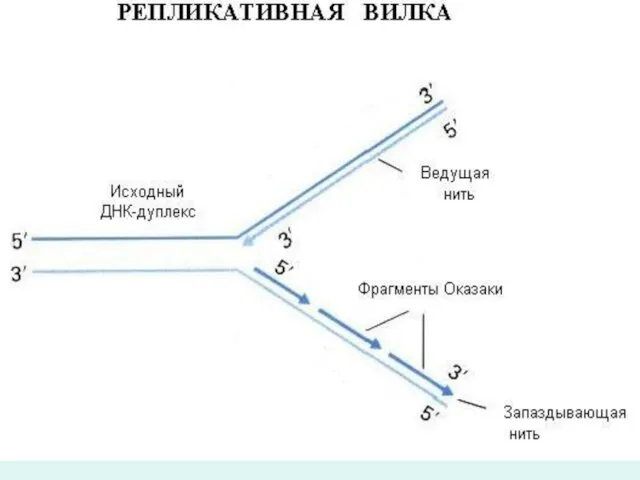
- 30. Мына бағыттағы синтез 3` 5` Оказаки фрагменті. ДНҚ –молекуласының жаңа тізбегінің біреуі матрицада үздіксіз,үзілмей репликайия айырының
- 31. Бұл реакция 2 кезеңнен тұрады: 1.ДНК-лигаза АМФ днаторы қызметін атқаратын НАД әсере етеді. Соның әсерінен -
- 34. Репликациядағы қателердің репарациясы Репликация барысында – өздігінен немесе сыртқы факторлардың әрекет етуінен(радиация,ультракүлгін сәулелену,химиялық агенттер және т.б.)қателер
- 35. Репликация қателіктері. ДНК молекуласының полинуклеотидтік тізбегінің синтезі кезінде болатын қателіктер 3 типті ДНК – полимеразасымен қалпына
- 36. Транскрипция . Оперонның функционалдық ұйымдасуы. РНК синтезіне қатсатын ДНК молекуласының гендері оперон атты бөлек функционалдық топтарға
- 37. ОПЕРОН Оперонның ақпараттық бөлімдері полипептидтік тізбектің және РНК структурасының ақпараты кодталған триплеттік ретпен цистрондармен және структуралық
- 38. Функциональная организация о п е р о н а - транскриптона ( Жакоб, Моно, 1965).
- 39. Оперон генінің қызметі. Ген промотордан транскрипция басталады. Оған транскрипцияны бастайтын белоктар (цАМФ рецепторлық протеин) және транскрипцияны
- 40. Транскрипция і фазада жүреді:инициация фазасы,элонгация фазасы,терминация фазасы. Инициация фазасында ДНКға тәуелді РНК полимераза протондық ген оперонға
- 42. Инициация фазасында РНК синтезінің бастапқы реакциясы болып 5`-3` фосфоэфирлық байланысқан АТФтың (или ГТФ) сәйкес екіні рибонуклеид
- 43. Элонгация фазасы Кейін элонгация фазасы болады – полинуклеотидтік РНК тізбегінің өсуі. РНК полимеразаның ДНК бойымен ауысуына
- 44. Терминация Терминация (аяқталу) РНК полимераза кодондарға жеткен кезде болады. БІр мезгілде арнайы белок транскрипцияны кодондармен қосылған
- 45. Процессинг В ходе процессинга удаляются неинформативные участки в пре-РНК и образуются функционально зрелые молекулы РНК. Процессинг
- 46. Трансляция негіздері Трансляция барысында 2 кезең қарасытуға болады: 1.Рекогниция немесе аминқышқылдарын тану 2.Ақуыз биосинтезі. Трансляция -
- 47. Рекогниция мехнизмі Эти ферменты (аминоацил -тРНК- синтетазы ) катализируют реакции активации аминокислот с образованием аминоацил-аденилата (а)
- 48. Ақуыз биосинтезі және трансляция факторлары Трансляцияның 2-ші кезеңі - полипептидтік тізбектің инициациясы. Бұл кезеңге қажетті компоненттер:
- 49. Рибосомалар Осы активті рибосоманың түзілуіне Ғ1, Ғ2, Ғ3 белоктық факторлар да өз үлесін қосады. Рибосоманың кіші
- 50. Рибосоманы құрастыру және рансляция фазалары. Бұл кезеңде белок синтезінің ядролық кезеңінде түзілген, белгілі бір полипептидтің, амин
- 51. . Пептидилдік центрде синтезделетін пептид тізбегі орналасса, аминоацилді центрде осы пептидтік тізбектің өсуіне қатысатын аминоацил-т-РНҚ орналасады.
- 52. Элонгация фазасы Бұл кезеңге қажетті заттар: екінші кезеңде түзілген активті рибосома; и-РНҚ-дағы кодондарға сәйкес келетін аминоацил
- 53. Терминация фазасы: Терминация яғни синтездің бітуі, аяқталу кезеңі, керекті эаттар:1/ АТФ;2/ белок синтезінің біткенін білдіруші и-РНҚ-дағы
- 54. Синтез белка
- 55. Антибиотиктер – матрицалық синтездің ингибиторлары ретінде. Белоктардың синтезі көптеген антибиотиктер әсерінен тежеуге ұшырауы мүмкін. Кейбір микроорганизмдер
- 56. Белоктар синтезінің реттелуі. Белок синтезінің реттелуі и-РНҚ-ның синтезі және трансляция /яғни белок синтезі/ кезеңінде жүреді. Бұл
- 58. Скачать презентацию

ДНҚ молекуласының құрылымы;
Жасушадағы ДНҚ полинуклеотидтік тізбегі мынандай нуклеотидттерден d-АМФ, d-ГМФ, d-ЦТФ,
ДНҚ молекуласының құрылымы;
Жасушадағы ДНҚ полинуклеотидтік тізбегі мынандай нуклеотидттерден d-АМФ, d-ГМФ, d-ЦТФ,
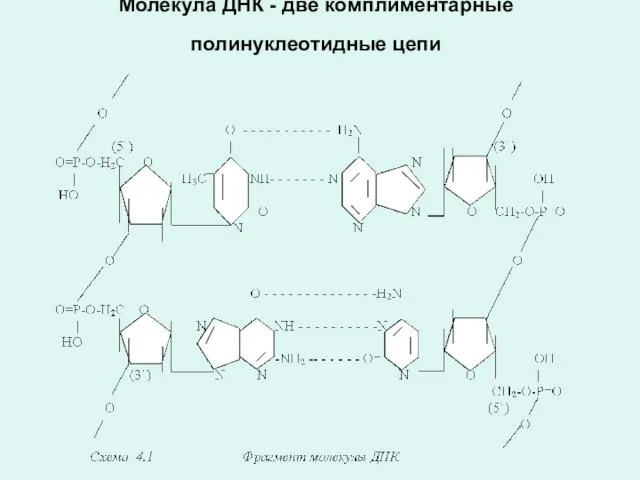
Молекула ДНК - две комплиментарные полинуклеотидные цепи
Молекула ДНК - две комплиментарные полинуклеотидные цепи
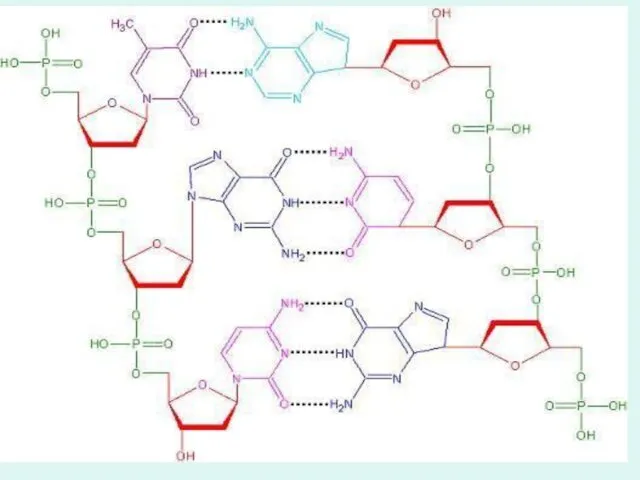

Чаргафф ережесі:
ДНҚ молекуласында:
1. Аденин саны Тиминге,Гуанин саны Цитозинге тең.
(
Чаргафф ережесі:
ДНҚ молекуласында:
1. Аденин саны Тиминге,Гуанин саны Цитозинге тең.
(

Генетикалық код құрылымы:
1. Триплет-ДНҚ құрамындағы полинуклеотидтік тізбектің кодтық элементі,сәйкес келетін аминқышкылдары
Генетикалық код құрылымы:
1. Триплет-ДНҚ құрамындағы полинуклеотидтік тізбектің кодтық элементі,сәйкес келетін аминқышкылдары

Триплетный код
Триплетный код

ГЕН
Қазіргі уақытта ген терминінің түпнегізінде ДНҚ жатыр, в котором в триплетной
ГЕН
Қазіргі уақытта ген терминінің түпнегізінде ДНҚ жатыр, в котором в триплетной

РНК құрылысы
РНҚ-ның ДНҚ айырмашылғы көп бөлігінде жалғыз тізбекті болып келеді.
РНК құрылысы
РНҚ-ның ДНҚ айырмашылғы көп бөлігінде жалғыз тізбекті болып келеді.

мРНК құрылысы
Матрицалық ДНК нмесе ақпараттық ДНК барлық РНҚ құрамының 3-5
мРНК құрылысы
Матрицалық ДНК нмесе ақпараттық ДНК барлық РНҚ құрамының 3-5

Рибосомалық РНҚ
Рибосома массасының 80% р-РНҚ-дан тұрады,ядрошықта жоғары метилденген рибосомалық РНҚ-ның рибосомалық
Рибосомалық РНҚ
Рибосома массасының 80% р-РНҚ-дан тұрады,ядрошықта жоғары метилденген рибосомалық РНҚ-ның рибосомалық

Тасымалдаушы т-РНК
РНК-ның 15 пайызға жуық бөлігін алып жатыр.
Т-РНК ең қысқа полинуклеотидті
Тасымалдаушы т-РНК
РНК-ның 15 пайызға жуық бөлігін алып жатыр.
Т-РНК ең қысқа полинуклеотидті

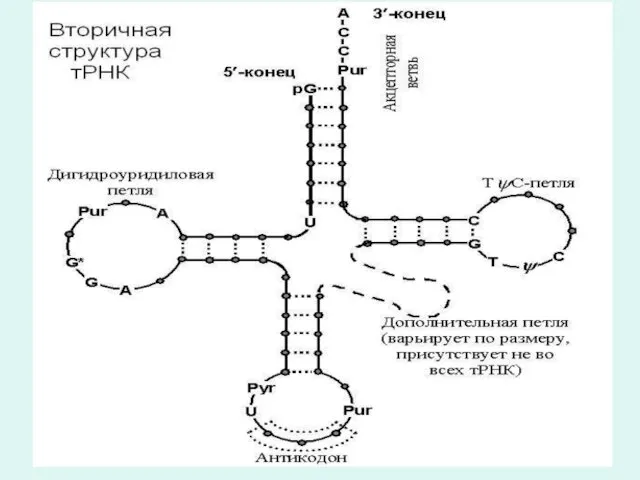
Т-РНК құрылымы
Т-РНК келесі құрылымдық бөліктерден тұрады:
1.Акцепторлы иіні-жұптасқан нуклеотидтер сабағынан тұрады және
Т-РНК құрылымы
Т-РНК келесі құрылымдық бөліктерден тұрады:
1.Акцепторлы иіні-жұптасқан нуклеотидтер сабағынан тұрады және

ДНҚ-ның матрицалық синтезі:
Днқ-ның екі еселену процессін-репликация деп атайды. (редупликация).
ДНҚ-екі еселенуі
ДНҚ-ның матрицалық синтезі:
Днқ-ның екі еселену процессін-репликация деп атайды. (редупликация).
ДНҚ-екі еселенуі

ДНҚ-ның репликациясы мен репарациясының молекулалық негізі.
ДНҚ репликациясына міндетті түрде қажет:
1.Жаңа тізбекті
ДНҚ-ның репликациясы мен репарациясының молекулалық негізі.
ДНҚ репликациясына міндетті түрде қажет:
1.Жаңа тізбекті

ферменттер:
1. Топоизомераза – “репликативтiк айырдың” алдындағы ДНҚ-ның аса жоғары ширатылған
ферменттер:
1. Топоизомераза – “репликативтiк айырдың” алдындағы ДНҚ-ның аса жоғары ширатылған

ДНҚ синтезінде мынандай қасиеттерге ие,антипаралельдік және унирполярлық.
Әрбір ДНҚ тізбегі белгілі бағытқа
ДНҚ синтезінде мынандай қасиеттерге ие,антипаралельдік және унирполярлық.
Әрбір ДНҚ тізбегі белгілі бағытқа

Днк плимеразаны артта қалған деп атайды себебі 3' 5‘ қозғала алады.
Днк плимеразаны артта қалған деп атайды себебі 3' 5‘ қозғала алады.

ДНК синтезі.
Днқ биосинтезі схемасы:
m(dАТФ+dТТФ)+ n(dГТФ+dЦТФ)------>ДНК + (m+n)Н4Р2О7
Бұл процесстің маңызды
ДНК синтезі.
Днқ биосинтезі схемасы:
m(dАТФ+dТТФ)+ n(dГТФ+dЦТФ)------>ДНК + (m+n)Н4Р2О7
Бұл процесстің маңызды
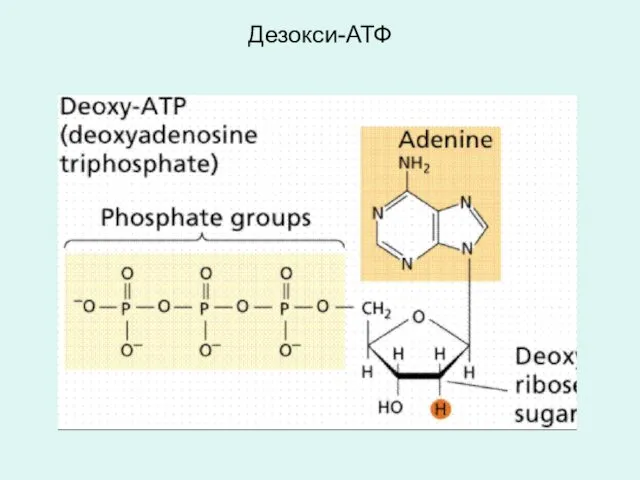
Дезокси-АТФ
Дезокси-АТФ

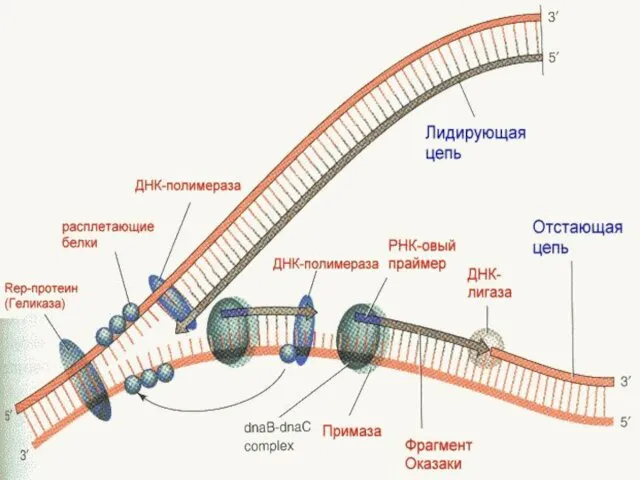
Днқ қос тізбегі өте мықты шиыршықталған,оны шешу үшін ерекше белоктар ферменттер
Днқ қос тізбегі өте мықты шиыршықталған,оны шешу үшін ерекше белоктар ферменттер

Днқ тізбегіндегі инициация процессі.
Днқ полимераза днқ синтезін матрицада бастай алмайды,себебі ол
Днқ тізбегіндегі инициация процессі.
Днқ полимераза днқ синтезін матрицада бастай алмайды,себебі ол

РНҚ праймерлерінің үзіліп түсіп қалуы,ДНҚ жіпшесінің жаңа синтезделген жіпшесі 10-20 нуклеотидке
РНҚ праймерлерінің үзіліп түсіп қалуы,ДНҚ жіпшесінің жаңа синтезделген жіпшесі 10-20 нуклеотидке

ДНК синтезі -"праймерлер".
(5` 3 `бағытындағы синтез. )
Хеликаза ферментінің әсерінен
ДНК синтезі -"праймерлер".
(5` 3 `бағытындағы синтез. )
Хеликаза ферментінің әсерінен

Праймерлерге ДНҚ-полимераза арқасында сәйкес дезоксиринукеозидтер 5` 3` бағытында жалғанады.
Нәтижесінде ДНҚ матрицалық
Праймерлерге ДНҚ-полимераза арқасында сәйкес дезоксиринукеозидтер 5` 3` бағытында жалғанады.
Нәтижесінде ДНҚ матрицалық

Мына бағыттағы синтез 3` 5` Оказаки фрагменті.
ДНҚ –молекуласының жаңа тізбегінің
Мына бағыттағы синтез 3` 5` Оказаки фрагменті.
ДНҚ –молекуласының жаңа тізбегінің

Бұл реакция 2 кезеңнен тұрады:
1.ДНК-лигаза АМФ днаторы қызметін атқаратын НАД әсере
Бұл реакция 2 кезеңнен тұрады:
1.ДНК-лигаза АМФ днаторы қызметін атқаратын НАД әсере
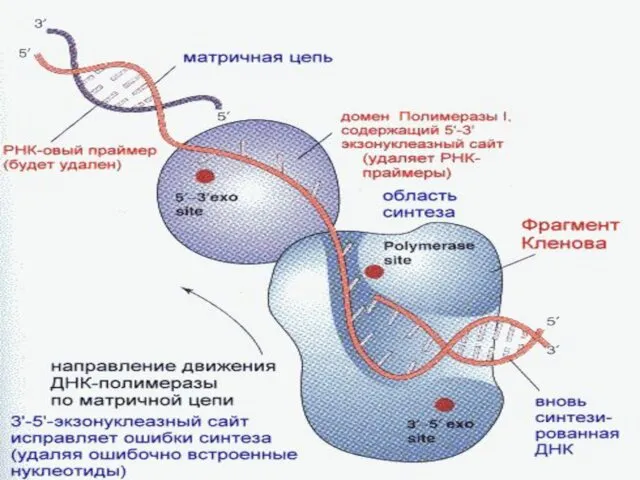

Репликациядағы қателердің репарациясы
Репликация барысында – өздігінен немесе сыртқы факторлардың әрекет етуінен(радиация,ультракүлгін
Репликациядағы қателердің репарациясы
Репликация барысында – өздігінен немесе сыртқы факторлардың әрекет етуінен(радиация,ультракүлгін

Репликация қателіктері.
ДНК молекуласының полинуклеотидтік тізбегінің синтезі кезінде болатын қателіктер 3 типті
Репликация қателіктері.
ДНК молекуласының полинуклеотидтік тізбегінің синтезі кезінде болатын қателіктер 3 типті

Транскрипция . Оперонның функционалдық ұйымдасуы.
РНК синтезіне қатсатын ДНК молекуласының гендері оперон
Транскрипция . Оперонның функционалдық ұйымдасуы.
РНК синтезіне қатсатын ДНК молекуласының гендері оперон

ОПЕРОН
Оперонның ақпараттық бөлімдері полипептидтік тізбектің және РНК структурасының ақпараты кодталған триплеттік
ОПЕРОН
Оперонның ақпараттық бөлімдері полипептидтік тізбектің және РНК структурасының ақпараты кодталған триплеттік
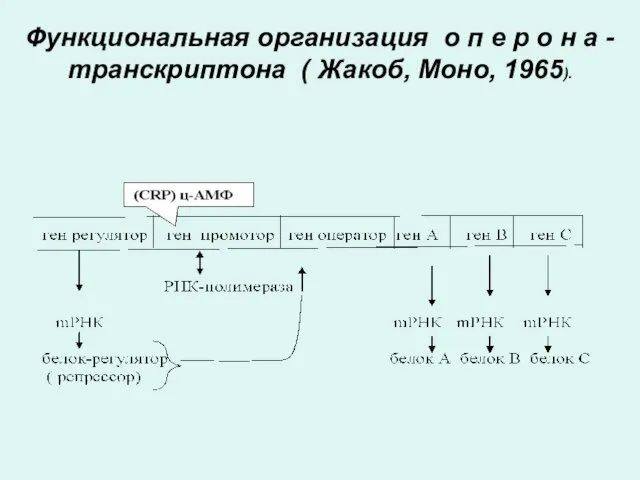
Функциональная организация о п е р о н а - транскриптона
Функциональная организация о п е р о н а - транскриптона

Оперон генінің қызметі.
Ген промотордан транскрипция басталады.
Оған транскрипцияны бастайтын белоктар (цАМФ рецепторлық
Оперон генінің қызметі.
Ген промотордан транскрипция басталады.
Оған транскрипцияны бастайтын белоктар (цАМФ рецепторлық

Транскрипция і фазада жүреді:инициация фазасы,элонгация фазасы,терминация фазасы.
Инициация фазасында ДНКға тәуелді
Инициация фазасында ДНКға тәуелді
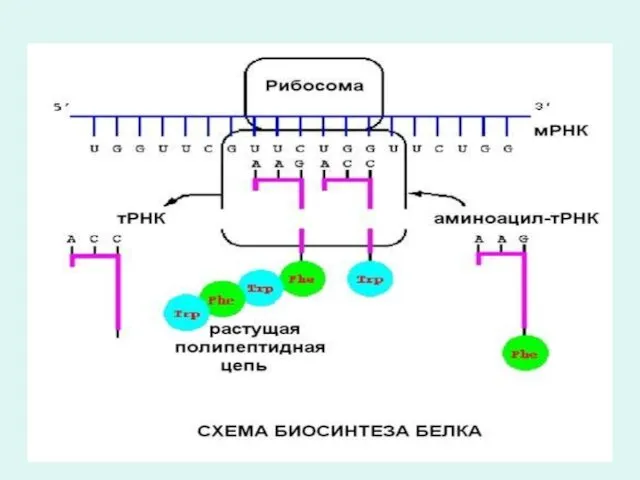

Инициация фазасында РНК синтезінің бастапқы реакциясы болып 5`-3` фосфоэфирлық байланысқан АТФтың
Инициация фазасында РНК синтезінің бастапқы реакциясы болып 5`-3` фосфоэфирлық байланысқан АТФтың

Элонгация фазасы
Кейін элонгация фазасы болады – полинуклеотидтік РНК тізбегінің өсуі.
РНК
Элонгация фазасы
Кейін элонгация фазасы болады – полинуклеотидтік РНК тізбегінің өсуі.
РНК

Терминация
Терминация (аяқталу) РНК полимераза кодондарға жеткен кезде болады.
БІр мезгілде арнайы
Терминация
Терминация (аяқталу) РНК полимераза кодондарға жеткен кезде болады.
БІр мезгілде арнайы

Процессинг
В ходе процессинга удаляются неинформативные участки в пре-РНК и образуются функционально
Процессинг
В ходе процессинга удаляются неинформативные участки в пре-РНК и образуются функционально

Трансляция негіздері
Трансляция барысында 2 кезең қарасытуға болады:
1.Рекогниция немесе аминқышқылдарын тану
2.Ақуыз биосинтезі.
Трансляция
Трансляция негіздері
Трансляция барысында 2 кезең қарасытуға болады:
1.Рекогниция немесе аминқышқылдарын тану
2.Ақуыз биосинтезі.
Трансляция
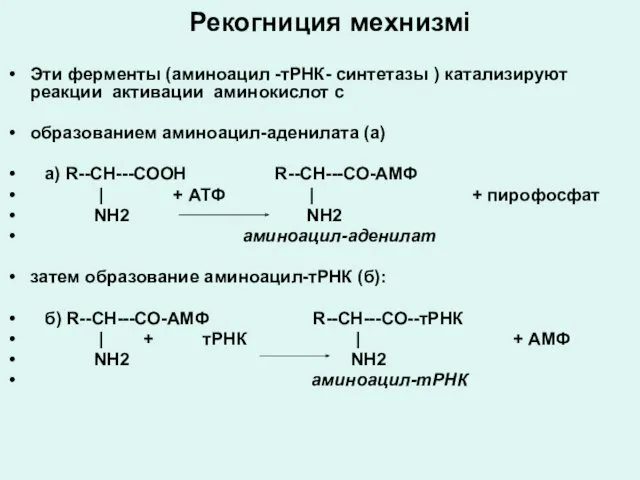
Рекогниция мехнизмі
Эти ферменты (аминоацил -тРНК- синтетазы ) катализируют реакции активации аминокислот
Рекогниция мехнизмі
Эти ферменты (аминоацил -тРНК- синтетазы ) катализируют реакции активации аминокислот

Ақуыз биосинтезі және трансляция факторлары
Трансляцияның 2-ші кезеңі - полипептидтік тізбектің
Ақуыз биосинтезі және трансляция факторлары
Трансляцияның 2-ші кезеңі - полипептидтік тізбектің

Рибосомалар
Осы активті рибосоманың түзілуіне Ғ1, Ғ2, Ғ3 белоктық факторлар да өз
Рибосомалар
Осы активті рибосоманың түзілуіне Ғ1, Ғ2, Ғ3 белоктық факторлар да өз

Рибосоманы құрастыру және рансляция фазалары.
Бұл кезеңде белок синтезінің ядролық кезеңінде түзілген,
Рибосоманы құрастыру және рансляция фазалары.
Бұл кезеңде белок синтезінің ядролық кезеңінде түзілген,

.
Пептидилдік центрде синтезделетін пептид тізбегі орналасса, аминоацилді центрде осы пептидтік тізбектің
.
Пептидилдік центрде синтезделетін пептид тізбегі орналасса, аминоацилді центрде осы пептидтік тізбектің

Элонгация фазасы
Бұл кезеңге қажетті заттар: екінші кезеңде түзілген активті рибосома;
Элонгация фазасы
Бұл кезеңге қажетті заттар: екінші кезеңде түзілген активті рибосома;

Терминация фазасы:
Терминация яғни синтездің бітуі, аяқталу кезеңі, керекті эаттар:1/ АТФ;2/ белок
Терминация фазасы:
Терминация яғни синтездің бітуі, аяқталу кезеңі, керекті эаттар:1/ АТФ;2/ белок
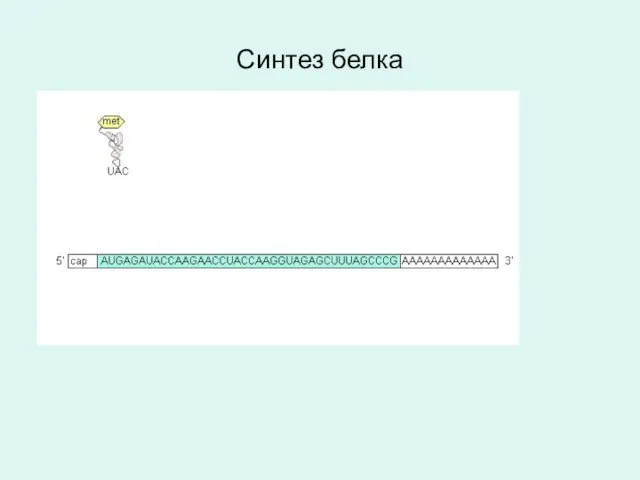
Синтез белка
Синтез белка

Антибиотиктер – матрицалық синтездің ингибиторлары ретінде.
Белоктардың синтезі көптеген антибиотиктер әсерінен тежеуге
Антибиотиктер – матрицалық синтездің ингибиторлары ретінде.
Белоктардың синтезі көптеген антибиотиктер әсерінен тежеуге

Белоктар синтезінің реттелуі. Белок синтезінің реттелуі и-РНҚ-ның синтезі және трансляция /яғни
Белоктар синтезінің реттелуі. Белок синтезінің реттелуі и-РНҚ-ның синтезі және трансляция /яғни
 Дослідження росту вегетативних органів
Дослідження росту вегетативних органів Грибы. Общая характеристика
Грибы. Общая характеристика Безусловные рефлексы
Безусловные рефлексы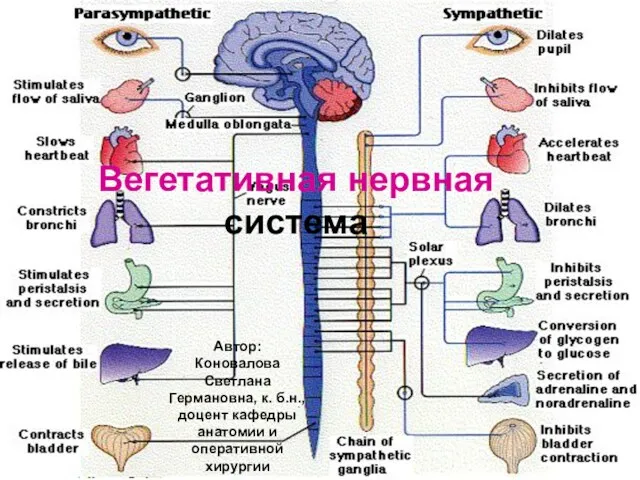 Вегетативная нервная система
Вегетативная нервная система Фізіологія виділення
Фізіологія виділення Только ли лист кормит растение? 5 класс
Только ли лист кормит растение? 5 класс Формы размножения организмов
Формы размножения организмов Ферменты. Типы ферментативных реакций
Ферменты. Типы ферментативных реакций Топырақ түсінігі
Топырақ түсінігі Грибы. Общая характеристика царства Грибы
Грибы. Общая характеристика царства Грибы Акутәрізді балықтардың системасы, отрядтары, таралуы, көлемі мен пішіндері, маңызы. Акутәрізді балықтардың алуан түрлілігі
Акутәрізді балықтардың системасы, отрядтары, таралуы, көлемі мен пішіндері, маңызы. Акутәрізді балықтардың алуан түрлілігі Physiology of Bacteria
Physiology of Bacteria Віроїди та пріони
Віроїди та пріони Растительный и животный мир Республики Удмуртия
Растительный и животный мир Республики Удмуртия Вещества, необходимые для жизнедеятельности организма. Витамины
Вещества, необходимые для жизнедеятельности организма. Витамины Биогеоценозы. Экосистемы. Строение и свойства
Биогеоценозы. Экосистемы. Строение и свойства Серце у людини
Серце у людини Особенности агрофитоценозов как антропогенно измененных НЭС
Особенности агрофитоценозов как антропогенно измененных НЭС Кожа. Функции и строение кожи
Кожа. Функции и строение кожи The 5 senses
The 5 senses Пропорциональные особенности и пластические точки лица человека
Пропорциональные особенности и пластические точки лица человека Растения и животные леса
Растения и животные леса Календула в назначениях современного доктора. Аптека в огороде
Календула в назначениях современного доктора. Аптека в огороде Рыбы реки Ирень
Рыбы реки Ирень Відходи технічних виробництв та харчові відходи
Відходи технічних виробництв та харчові відходи Генетика развития растений. Развитие листа
Генетика развития растений. Развитие листа Биохимия крови
Биохимия крови Симбиоз и симбиогенез организмов
Симбиоз и симбиогенез организмов