- Электрические свойства нейронов

Содержание
- 2. Сигнал по мембране нейрона передается в виде коротких элект-рических импульсов – потенциалов действия (ПД). Этот процесс
- 3. Потенциал покоя (ПП) нейрона – его постоянный отрицательный заряд, равный в среднем -70 мВ. Измерить ПП
- 4. Наличие ПП – результат жизнедеятельности нейрона, совместного функционирования всех биополимеров и органоидов клетки; погибший нейрон быстро
- 5. В результате работы Na+-K+-АТФазы в нейроне оказывается примерно в 10 раз меньше Na+ и в 30
- 6. Как долго идет диффузия К+ из нейрона? Очевидный вариант («до выравнивания концентраций») неверен, поскольку двигаются заряженные
- 7. ПП – это отрицате-льный заряд цито-плазмы, останавли-вающий диффузию ионов К+ в межкле-точную среду. Вальтер Нернст (Ноб.пр.
- 8. ПП = -91 мВ («равновесный потенциал» для К+) В реальной клетке ПП находится ближе к нулю
- 9. внутриклеточная среда Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх и частичной потере ПП В
- 10. Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх и частичной потере ПП (отсюда название –
- 11. Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх и частичной потере ПП (отсюда название –
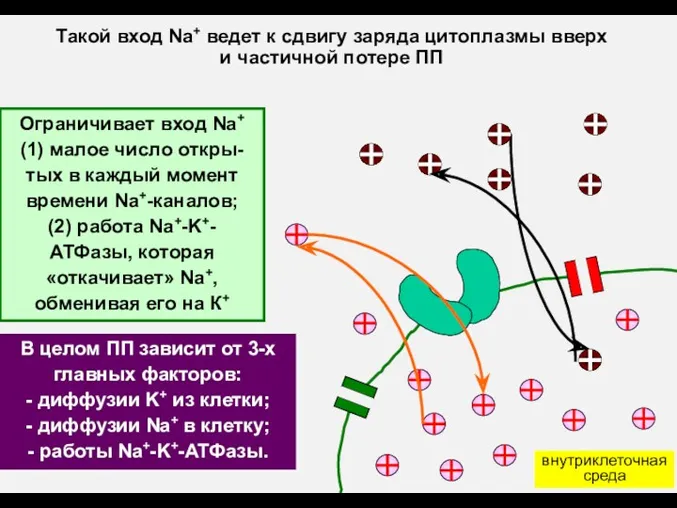
- 12. В целом ПП зависит от 3-х главных факторов: - диффузии K+ из клетки; - диффузии Na+
- 13. Работа Na+-K+-АТФазы может быть нарушена химич. веществами, например, токсином одной из тропических лиан строфантином. Строфантин: повышение
- 14. ПП вход ионов натрия АНАЛОГИЯ ПП: лодка на поверхности водоема. Уровень воды
- 15. ПД – универсальный ответ нервной клетки на стимуляцию 20 мВ: пороговый стимул при ПП= -70 мВ
- 16. При ПП=-80 мВ, пороговый стимул= …? 30 мВ При ПП=-60 мВ, пороговый стимул= …? 10 мВ
- 17. Рассмотрим ПД подробнее. Длительность ПД на схеме составляет 1 мс. По ходу ПД можно выделить восходящую
- 18. В основе этих процессов – открывание и закрывание электрочувствительных Na+- и К+-каналов. Эти каналы имеют створки,
- 19. В основе этих процессов – открывание и закрывание электрочувствительных Na+- и К+-каналов. Эти каналы имеют створки,
- 20. Для закрытия Na+-кана-лов на пике ПД служит дополнительная створка (внутриклеточная, «большая», инактива-ционная); основная створка («малая», акти-вационная)
- 21. Именно разная скорость открытия Na+-каналов и К+-каналов позволяет возникнуть сначала восходящей, а затем – нисходящей фазе
- 22. Мы познакомились с общими принципами генерации ПД. Следующие три вопроса: [1]. Что будет, если заблокировать электрочувствительные
- 23. тетродотоксин – яд рыбы фугу (аминогруппа работает как «пробка» для Na+-канала) В результате действия токсина прекра-
- 24. В результате действия токсина прекра- щается генерация и проведение ПД: сначала – по периферическим нервам («иллюзии»
- 25. ТЭА – тетраэтиламмоний: работает как «пробка» по отношению к К+-каналу. В результате восходящая фаза ПД изменяется
- 26. Этот рисунок уже был. Он иллюстрирует не только вклад Na+-K+-АТФазы в поддержание ПП, но и позволяет
- 27. РАСПРОСТРАНЕНИЕ ПД. Если ПД возник хотя бы в одной точке мембраны ней- рона – он распространяется
- 28. РАСПРОСТРАНЕНИЕ ПД. Если ПД возник хотя бы в одной точке мембраны ней- рона – он распространяется
- 29. Гигантский аксон кальмара (d=0.5-1 мм) – классичес-кий объект для изучения ПД Замена аксоплазмы на раствор, содержащий
- 30. «Радикальный» рост скорости проведения – за счет миели-низации аксонов, которая на периферии обеспечивается одним из типов
- 31. Миелиновая оболочка (несколько десятков мемб-ранных слоев) – хороший изолятор. В связи с этим связанные с ПД
- 32. А) олигодендроциты (в т.ч. шванновские клетки): электроизоляции нейронов; в ЦНС один олигодендроцит образует миелиновые оболочки на
- 33. Что еще? * нейроны-пейсмекеры * местные анестетики * батрахотоксин * электрические рыбы
- 34. Чем больше постоянно открытых Na+-каналов, тем чаще следуют ПД. Регуляция частоты разрядов идет также за счет
- 35. НОВОКАИН – гидрохлорид диэтиламиноэтилового эфира аминобензойной кислоты. «Анестезия» (греч.) – потеря чувствительности. Местные анестетики: проникают внутрь
- 36. Батрахотоксин: токсин кожи некоторых лягушек-листолазов; модифицированный стероидный гормон насекомых. Токсин проникает внутрь клетки и связывается с
- 37. У электрических рыб (например, электрического угря) имеются особые видоизмененные мышечные клетки – электроциты. Они собраны в
- 38. Генерация электрического поля (нильский слоник) + электрорецепция у рыб и млекопитающих
- 40. Скачать презентацию
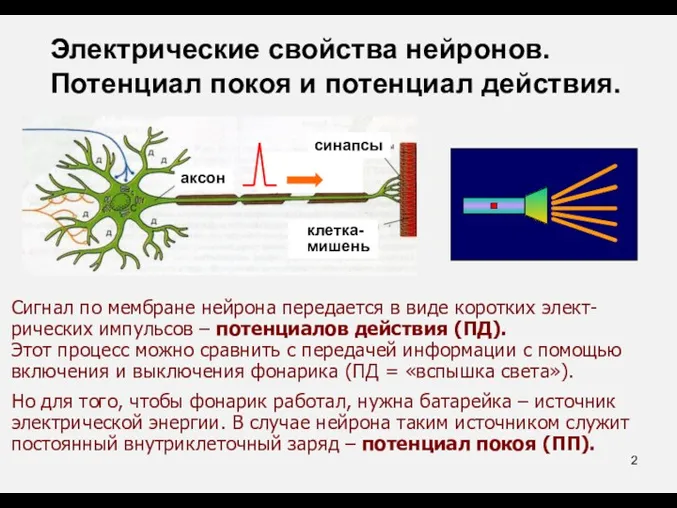
Сигнал по мембране нейрона передается в виде коротких элект-рических импульсов –
Сигнал по мембране нейрона передается в виде коротких элект-рических импульсов –
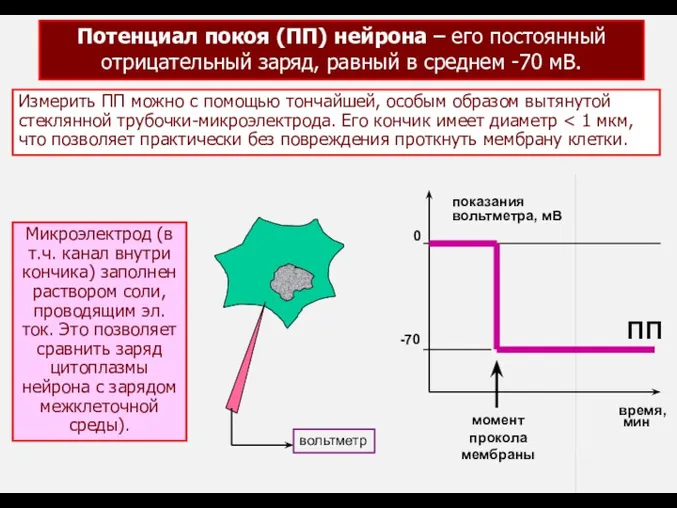
Потенциал покоя (ПП) нейрона – его постоянный
отрицательный заряд, равный в
Потенциал покоя (ПП) нейрона – его постоянный
отрицательный заряд, равный в
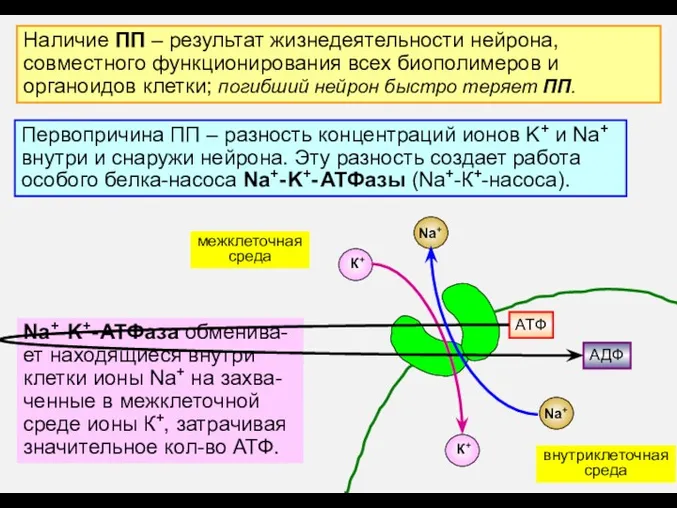
Наличие ПП – результат жизнедеятельности нейрона,
совместного функционирования всех биополимеров и
органоидов
Наличие ПП – результат жизнедеятельности нейрона,
совместного функционирования всех биополимеров и
органоидов
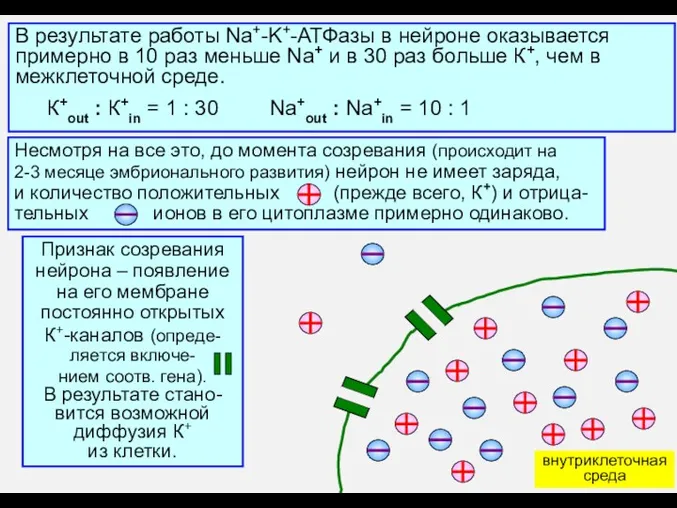
В результате работы Na+-K+-АТФазы в нейроне оказывается
примерно в 10 раз
В результате работы Na+-K+-АТФазы в нейроне оказывается
примерно в 10 раз
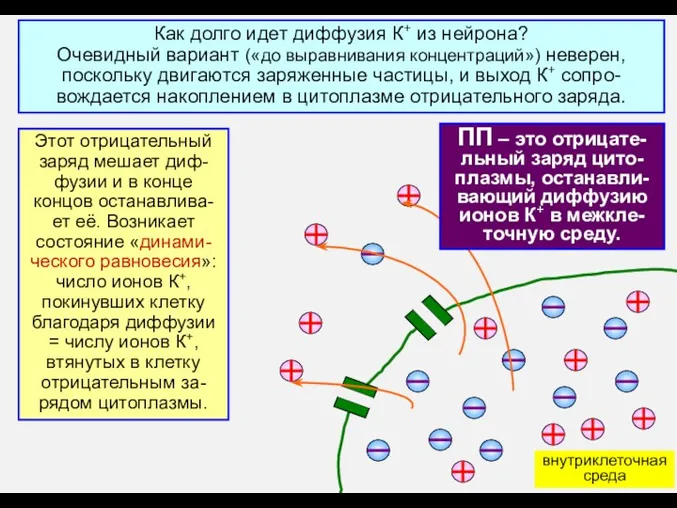
Как долго идет диффузия К+ из нейрона?
Очевидный вариант («до выравнивания концентраций»)
Как долго идет диффузия К+ из нейрона?
Очевидный вариант («до выравнивания концентраций»)
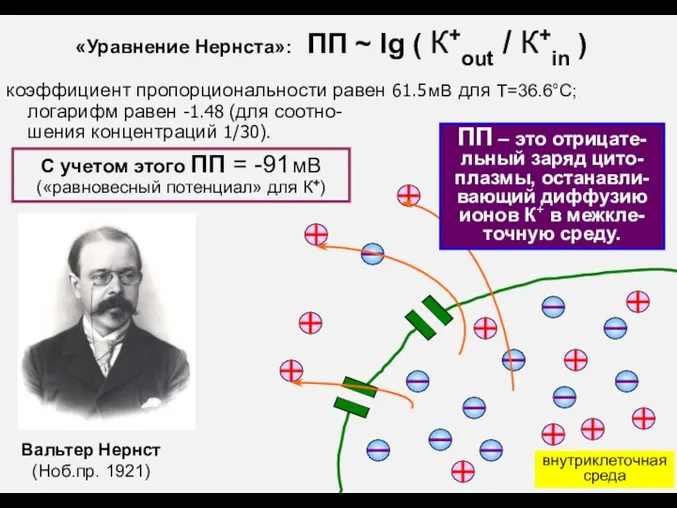
ПП – это отрицате-льный заряд цито-плазмы, останавли-вающий диффузию ионов К+ в
ПП – это отрицате-льный заряд цито-плазмы, останавли-вающий диффузию ионов К+ в
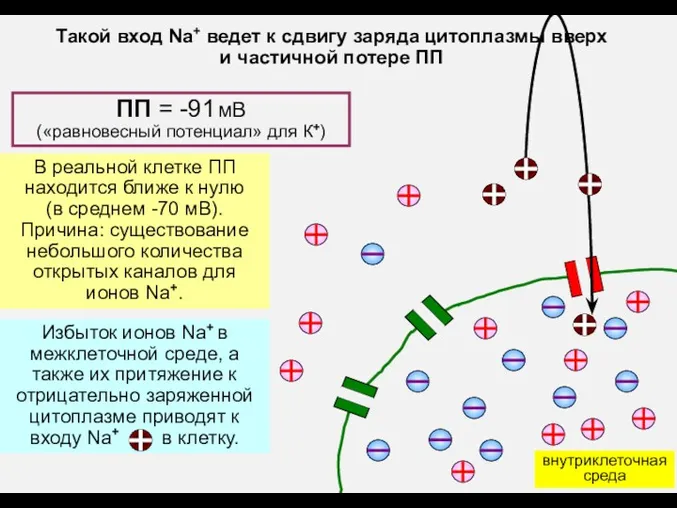
ПП = -91 мВ
(«равновесный потенциал» для К+)
В реальной клетке ПП
находится
ПП = -91 мВ
(«равновесный потенциал» для К+)
В реальной клетке ПП
находится
внутриклеточная
среда
Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх
и частичной потере
внутриклеточная
среда
Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх
и частичной потере
Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх
и частичной потере
Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх
и частичной потере
Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх
и частичной потере
Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх
и частичной потере
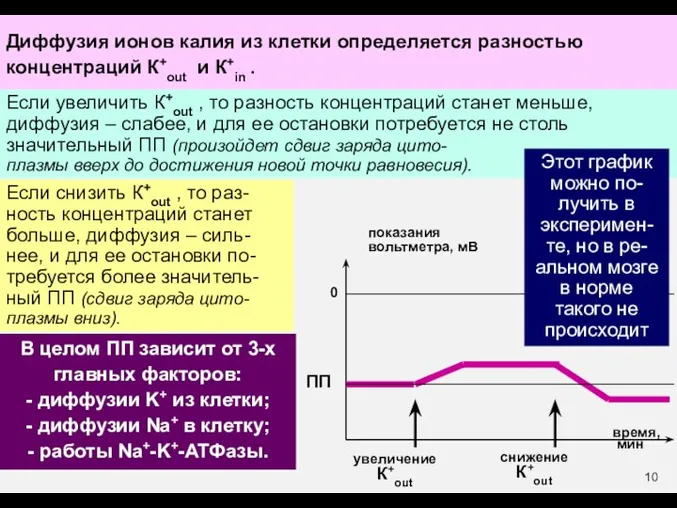
В целом ПП зависит от 3-х главных факторов:
- диффузии K+ из
В целом ПП зависит от 3-х главных факторов:
- диффузии K+ из
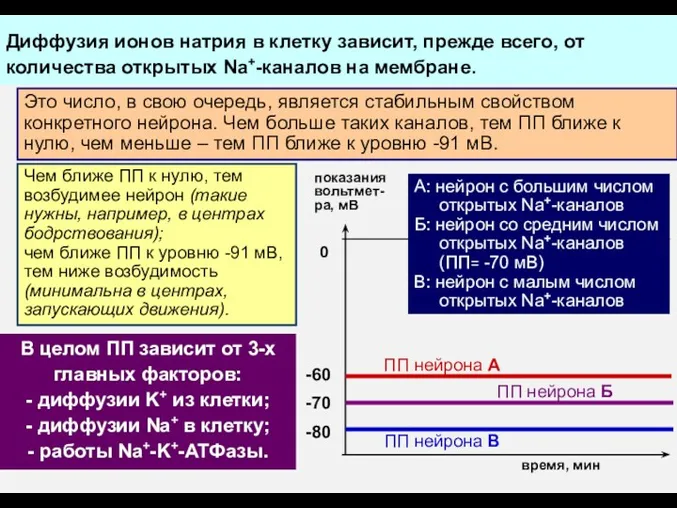
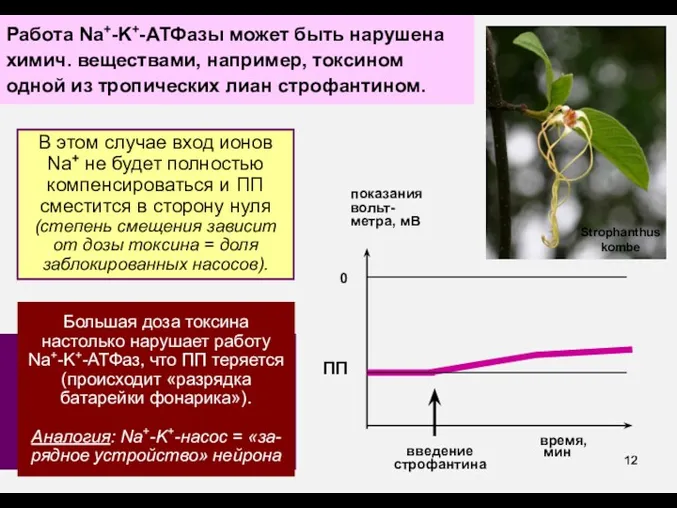
Работа Na+-K+-АТФазы может быть нарушена химич. веществами, например, токсином одной из
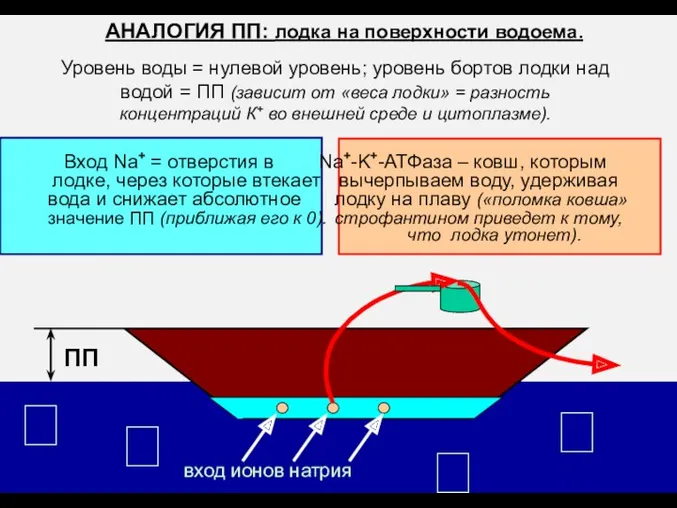
ПП
вход ионов натрия
АНАЛОГИЯ ПП: лодка на поверхности водоема.
Уровень воды =
ПП
вход ионов натрия
АНАЛОГИЯ ПП: лодка на поверхности водоема.
Уровень воды =
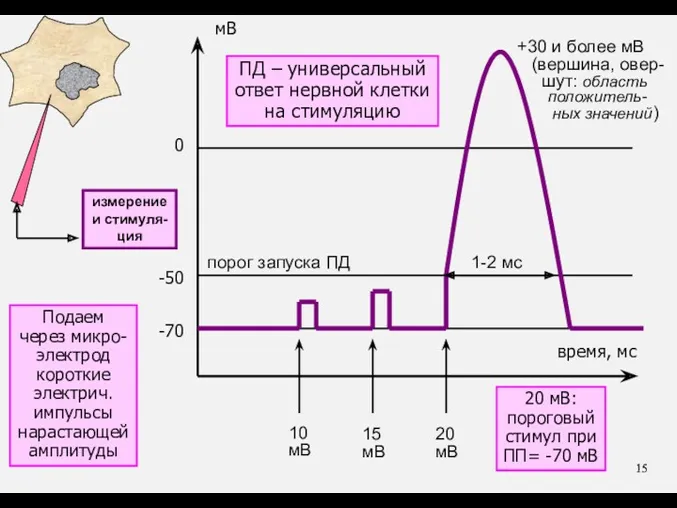
ПД – универсальный
ответ нервной клетки
на стимуляцию
20 мВ:
пороговый
стимул при
ПП= -70
ПД – универсальный
ответ нервной клетки
на стимуляцию
20 мВ:
пороговый
стимул при
ПП= -70
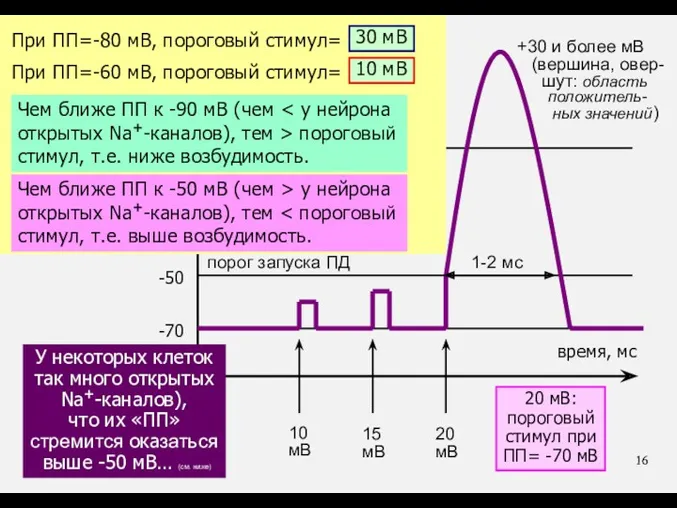
При ПП=-80 мВ, пороговый стимул= …?
30 мВ
При ПП=-60 мВ, пороговый стимул=
При ПП=-80 мВ, пороговый стимул= …?
30 мВ
При ПП=-60 мВ, пороговый стимул=
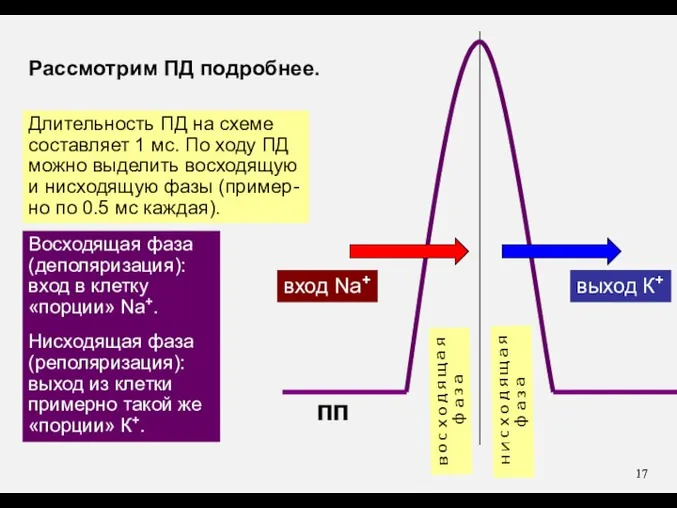
Рассмотрим ПД подробнее.
Длительность ПД на схеме
составляет 1 мс. По ходу
Рассмотрим ПД подробнее.
Длительность ПД на схеме
составляет 1 мс. По ходу
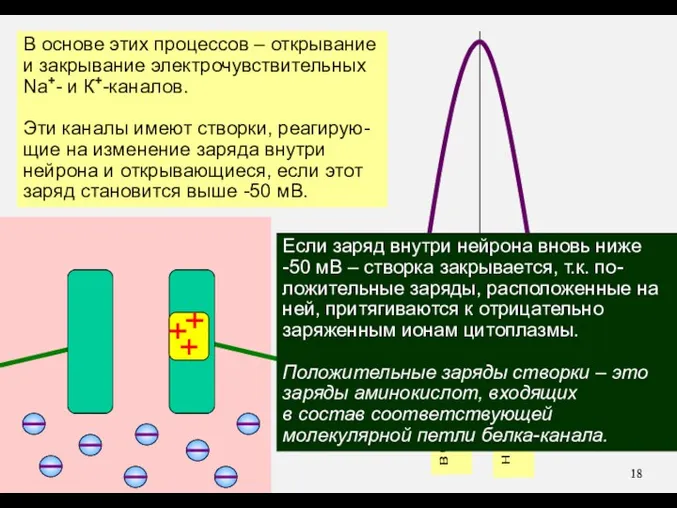
В основе этих процессов – открывание
и закрывание электрочувствительных
Na+- и К+-каналов.
Эти каналы
В основе этих процессов – открывание
и закрывание электрочувствительных
Na+- и К+-каналов.
Эти каналы

В основе этих процессов – открывание
и закрывание электрочувствительных
Na+- и К+-каналов.
Эти каналы
В основе этих процессов – открывание
и закрывание электрочувствительных
Na+- и К+-каналов.
Эти каналы
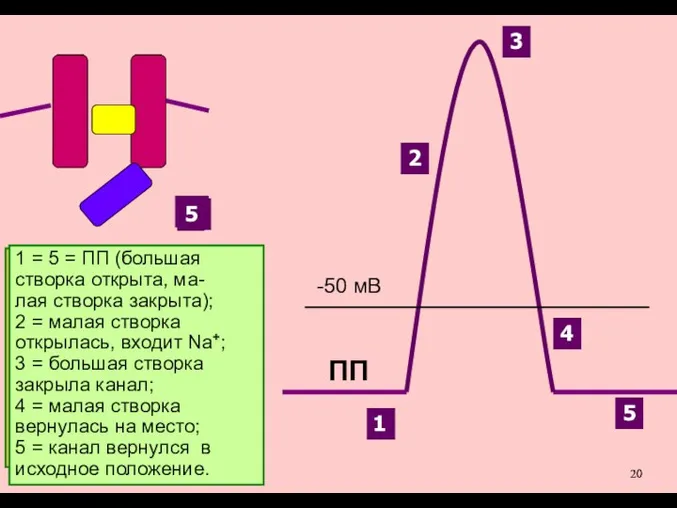
Для закрытия Na+-кана-лов на пике ПД служит дополнительная створка (внутриклеточная, «большая»,
Для закрытия Na+-кана-лов на пике ПД служит дополнительная створка (внутриклеточная, «большая»,
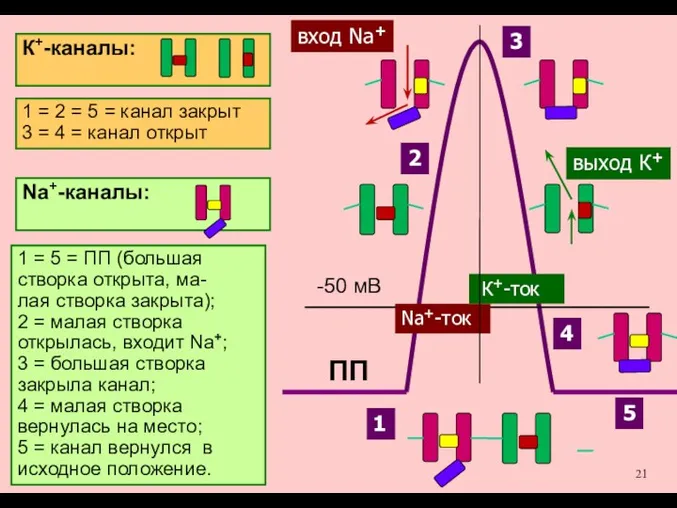
Именно разная скорость открытия
Na+-каналов и К+-каналов позволяет
возникнуть сначала восходящей, а
затем
Именно разная скорость открытия
Na+-каналов и К+-каналов позволяет
возникнуть сначала восходящей, а
затем

Мы познакомились с общими принципами генерации ПД.
Следующие три вопроса:
[1].
Мы познакомились с общими принципами генерации ПД.
Следующие три вопроса:
[1].
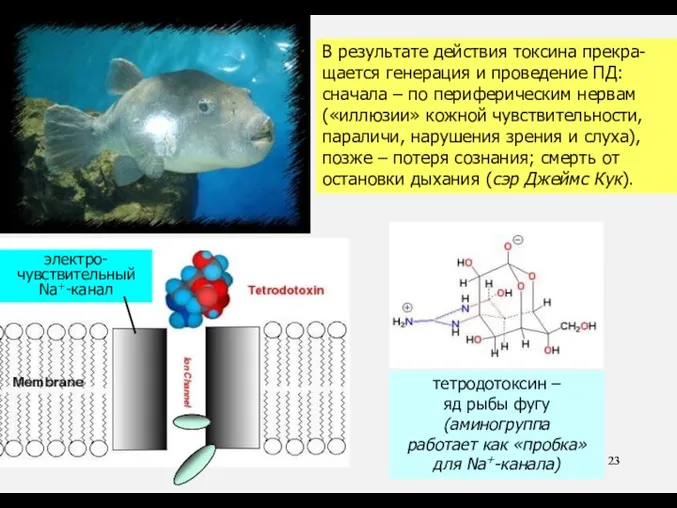
тетродотоксин –
яд рыбы фугу
(аминогруппа
работает как «пробка»
для Na+-канала)
В результате действия токсина прекра-
тетродотоксин –
яд рыбы фугу
(аминогруппа
работает как «пробка»
для Na+-канала)
В результате действия токсина прекра-

В результате действия токсина прекра- щается генерация и проведение ПД: сначала
В результате действия токсина прекра- щается генерация и проведение ПД: сначала
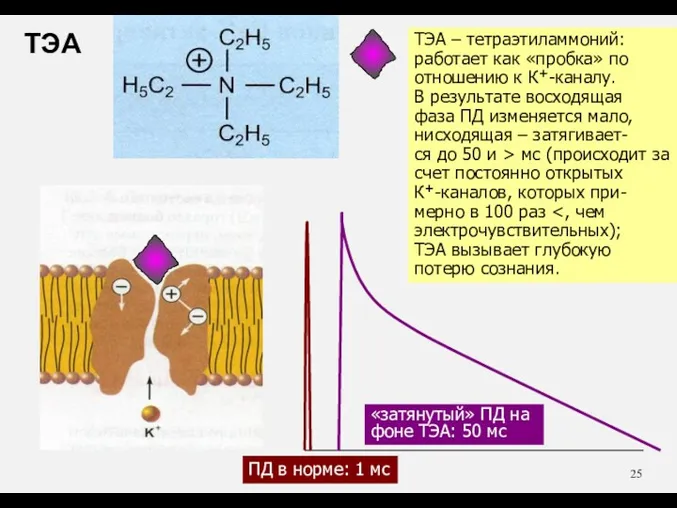
ТЭА – тетраэтиламмоний:
работает как «пробка» по
отношению к К+-каналу.
В результате восходящая
фаза ПД
ТЭА – тетраэтиламмоний:
работает как «пробка» по
отношению к К+-каналу.
В результате восходящая
фаза ПД
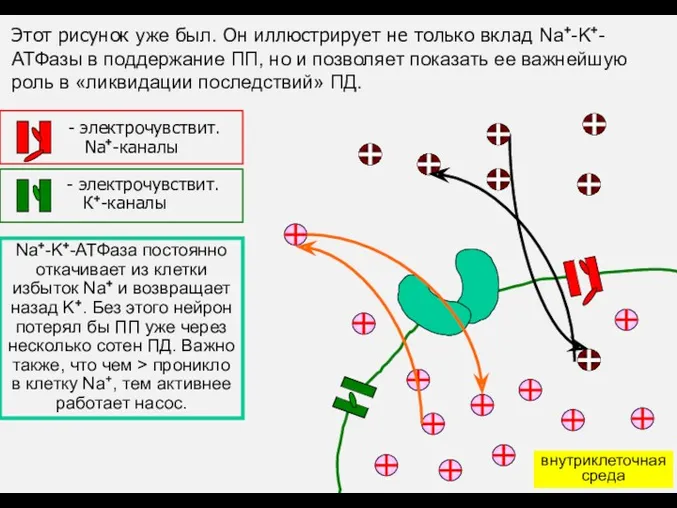
Этот рисунок уже был. Он иллюстрирует не только вклад Na+-K+-АТФазы в
Этот рисунок уже был. Он иллюстрирует не только вклад Na+-K+-АТФазы в
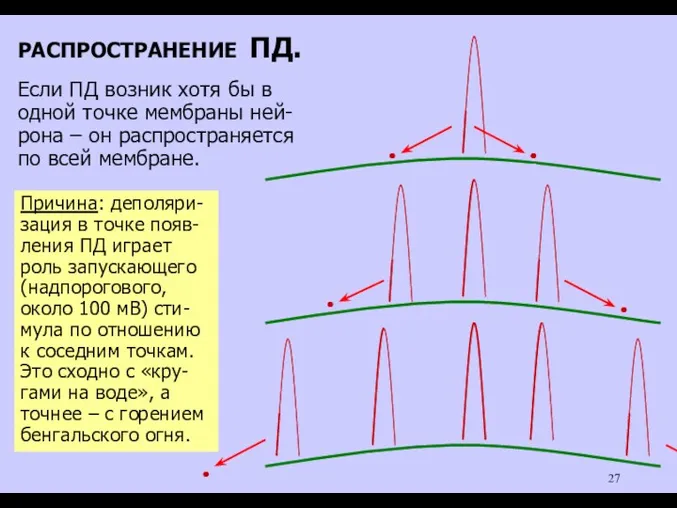
РАСПРОСТРАНЕНИЕ ПД.
Если ПД возник хотя бы в
одной точке мембраны ней-
рона
РАСПРОСТРАНЕНИЕ ПД.
Если ПД возник хотя бы в
одной точке мембраны ней-
рона
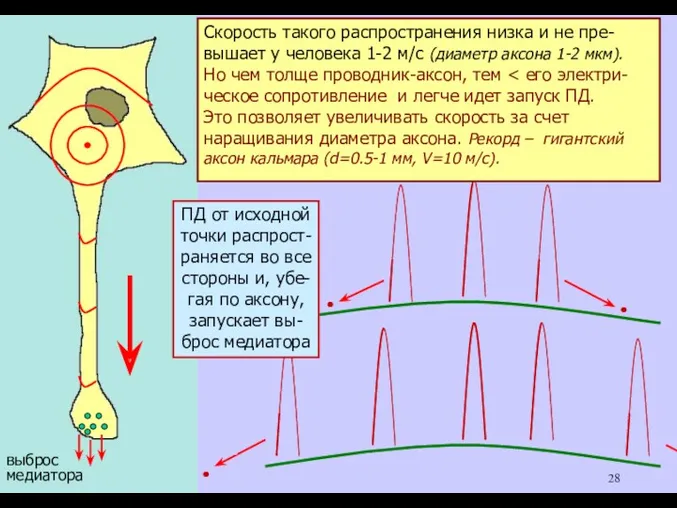
РАСПРОСТРАНЕНИЕ ПД.
Если ПД возник хотя бы в
одной точке мембраны ней-
рона
РАСПРОСТРАНЕНИЕ ПД.
Если ПД возник хотя бы в
одной точке мембраны ней-
рона
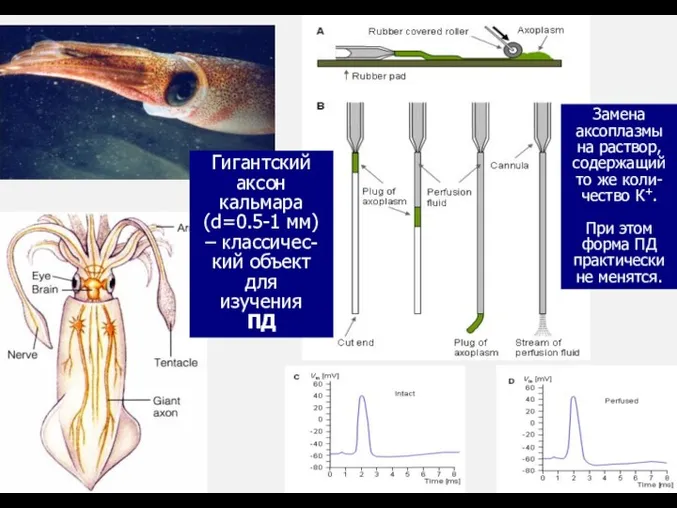
Гигантский
аксон
кальмара
(d=0.5-1 мм) – классичес-кий объект для
изучения
ПД
Замена аксоплазмы на
Гигантский
аксон
кальмара
(d=0.5-1 мм) – классичес-кий объект для
изучения
ПД
Замена аксоплазмы на
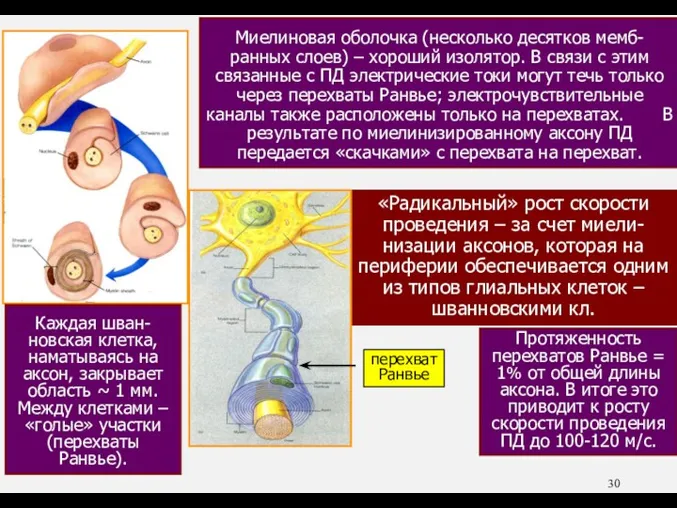
«Радикальный» рост скорости проведения – за счет миели-низации аксонов, которая на
«Радикальный» рост скорости проведения – за счет миели-низации аксонов, которая на
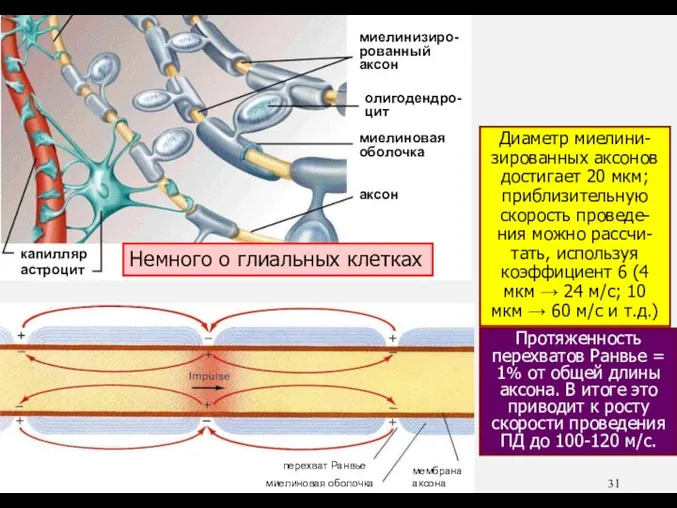
Миелиновая оболочка (несколько десятков мемб-ранных слоев) – хороший изолятор. В связи
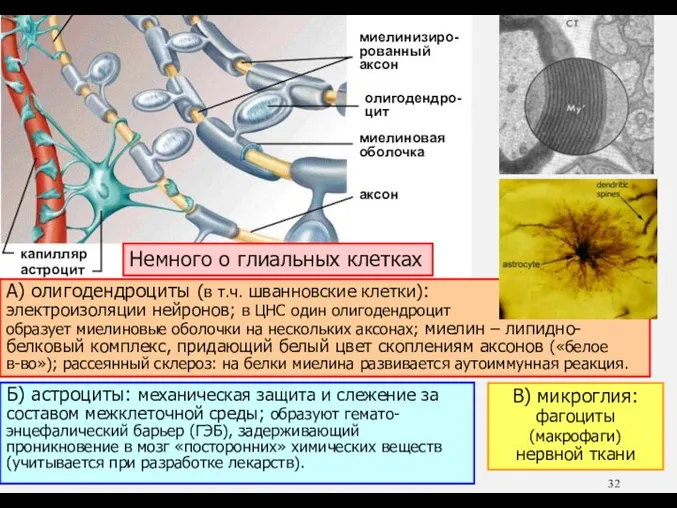
А) олигодендроциты (в т.ч. шванновские клетки):
электроизоляции нейронов; в ЦНС один
А) олигодендроциты (в т.ч. шванновские клетки):
электроизоляции нейронов; в ЦНС один

Что еще?
* нейроны-пейсмекеры
* местные анестетики
* батрахотоксин
* электрические рыбы
Что еще?
* нейроны-пейсмекеры
* местные анестетики
* батрахотоксин
* электрические рыбы
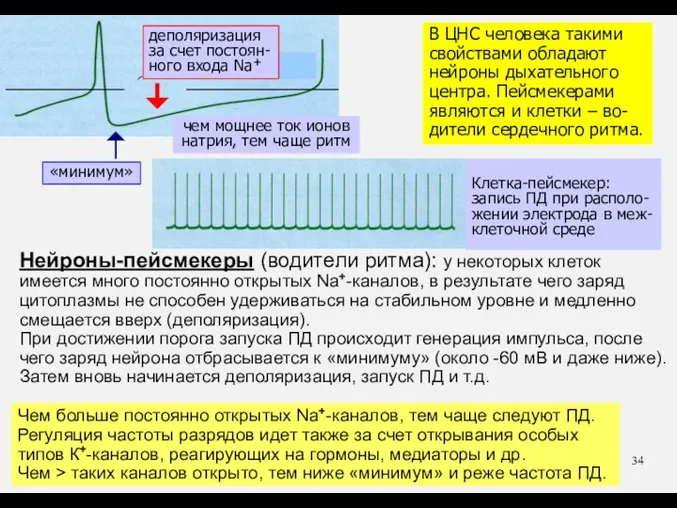
Чем больше постоянно открытых Na+-каналов, тем чаще следуют ПД. Регуляция частоты
Чем больше постоянно открытых Na+-каналов, тем чаще следуют ПД. Регуляция частоты
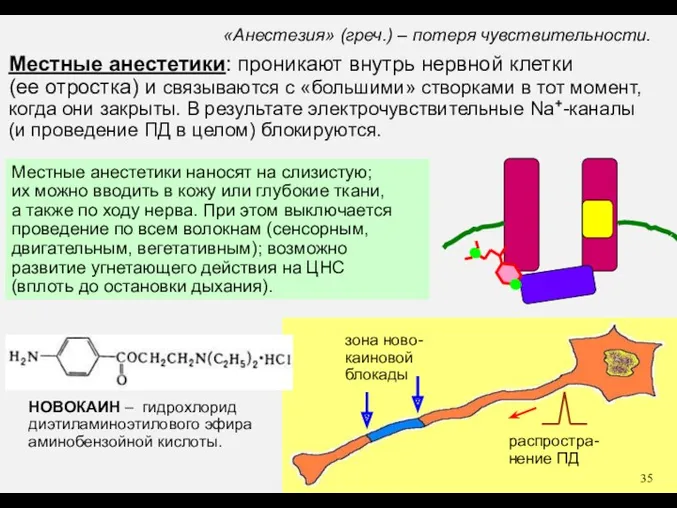
НОВОКАИН – гидрохлорид
диэтиламиноэтилового эфира
аминобензойной кислоты.
«Анестезия» (греч.) – потеря чувствительности.
Местные
НОВОКАИН – гидрохлорид
диэтиламиноэтилового эфира
аминобензойной кислоты.
«Анестезия» (греч.) – потеря чувствительности.
Местные
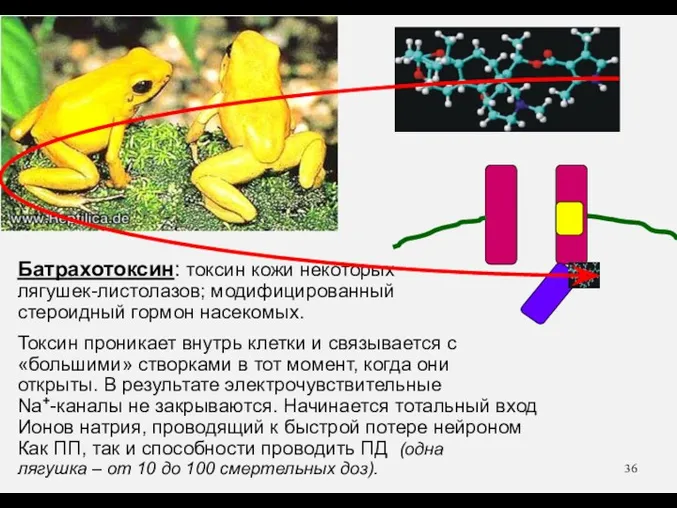
Батрахотоксин: токсин кожи некоторых
лягушек-листолазов; модифицированный
стероидный гормон насекомых.
Токсин проникает внутрь
Батрахотоксин: токсин кожи некоторых
лягушек-листолазов; модифицированный
стероидный гормон насекомых.
Токсин проникает внутрь
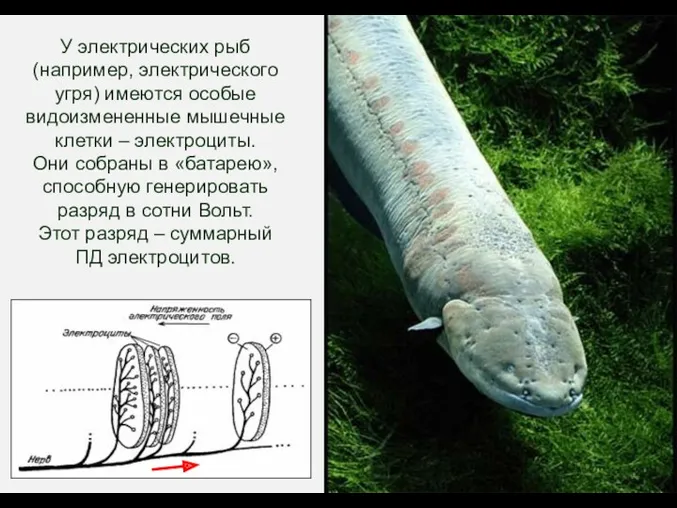
У электрических рыб (например, электрического угря) имеются особые видоизмененные мышечные клетки
У электрических рыб (например, электрического угря) имеются особые видоизмененные мышечные клетки

Генерация электрического поля (нильский слоник) + электрорецепция у рыб и млекопитающих
Генерация электрического поля (нильский слоник) + электрорецепция у рыб и млекопитающих
 Тип Членистоногие (Arthropoda). Класс Скрыточелюстные насекомые (Insecta - Entognatha)
Тип Членистоногие (Arthropoda). Класс Скрыточелюстные насекомые (Insecta - Entognatha) Эволюционное учение (урок обобщающего повторения для 10 класса)
Эволюционное учение (урок обобщающего повторения для 10 класса) Цветковые растения
Цветковые растения История развития биологических знаний. Предмет, задачи и методы биологии. Основные концепции современной биологии
История развития биологических знаний. Предмет, задачи и методы биологии. Основные концепции современной биологии Гідрологія. Показники якості води
Гідрологія. Показники якості води Сердечно-сосудистая система. Сердце. Кровеносные сосуды. Лимфатическая система
Сердечно-сосудистая система. Сердце. Кровеносные сосуды. Лимфатическая система Презентация к уроку биологии Царство: грибы 6 класс Часть 2 Диск
Презентация к уроку биологии Царство: грибы 6 класс Часть 2 Диск Цитоскелет
Цитоскелет Физиология ЦНС. Промежуточный мозг и ретикулярная формация
Физиология ЦНС. Промежуточный мозг и ретикулярная формация Урок-презентация по теме :Бабочки.
Урок-презентация по теме :Бабочки. Происхождение человека (антропогенез)
Происхождение человека (антропогенез) Розовый слон
Розовый слон Гепард
Гепард Презентация по биологии 6 класс Мхи
Презентация по биологии 6 класс Мхи Генетика человека
Генетика человека Живые организмы весной
Живые организмы весной Видоизменения побегов
Видоизменения побегов Cellular Respiration
Cellular Respiration 20230330_genetika_obnov_wecompress.com_
20230330_genetika_obnov_wecompress.com_ Применение проектной технологии на уроках биологии и во внеурочной деятельности
Применение проектной технологии на уроках биологии и во внеурочной деятельности Зоология позвоночных. Надкласс четвероногие. (Лекция 7)
Зоология позвоночных. Надкласс четвероногие. (Лекция 7) Гортензия метельчатая Bombshell
Гортензия метельчатая Bombshell Катаболизм аминокислот
Катаболизм аминокислот Разнообразие рептилий
Разнообразие рептилий Digestion
Digestion Перелетные птицы
Перелетные птицы Популяция. Свойства популяций
Популяция. Свойства популяций Красная книга Крыма
Красная книга Крыма