- Фазовые переходы в мембранах. Лиотропный мезоморфизм липидов

Содержание
- 2. Фазовые переходы липидов в мембранах В водной среде липидные структуры ведут себя как анизотропные жидкости (жидкие
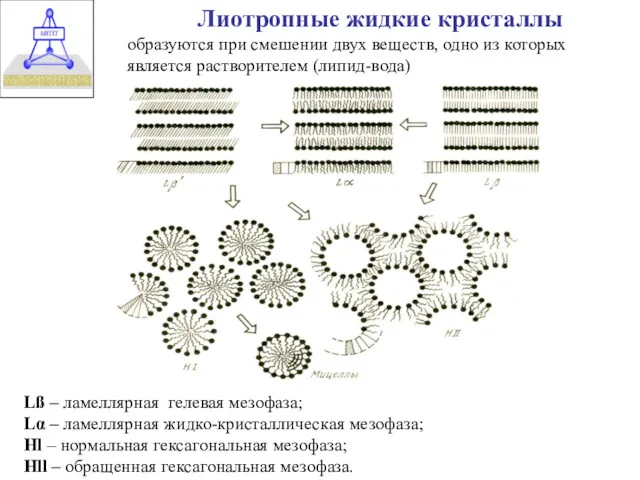
- 3. Лиотропные жидкие кристаллы образуются при смешении двух веществ, одно из которых является растворителем (липид-вода) Lß –
- 4. Гидратация липидов липид + Вода [мало ] гидратированный липид + Вода модификация полярной «головы» липида: увеличение
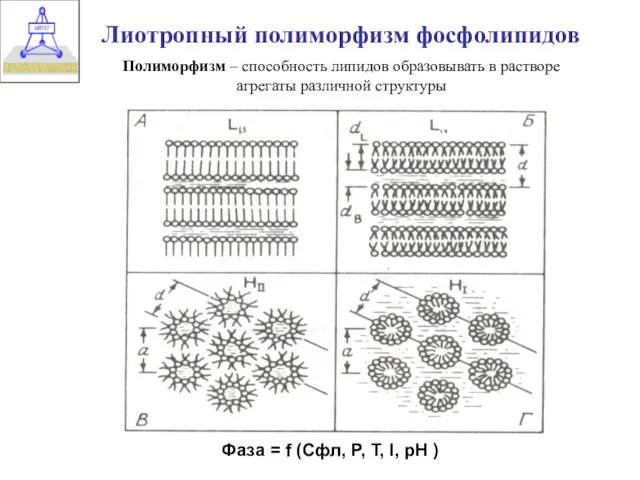
- 5. Лиотропный полиморфизм фосфолипидов Полиморфизм – способность липидов образовывать в растворе агрегаты различной структуры Фаза = f
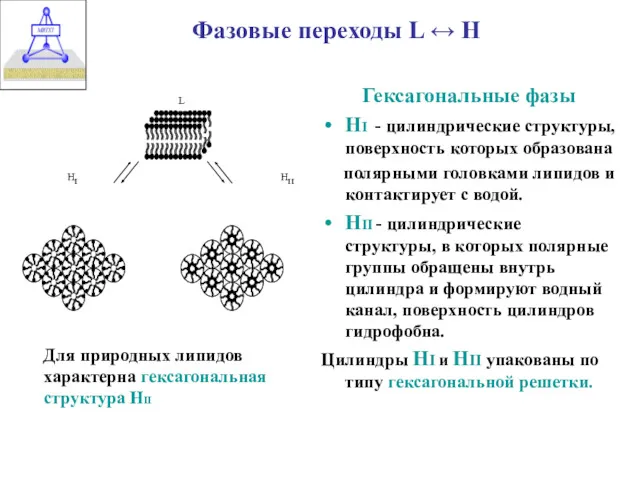
- 6. Фазовые переходы L ↔ H Гексагональные фазы НI - цилиндрические структуры, поверхность которых образована полярными головками
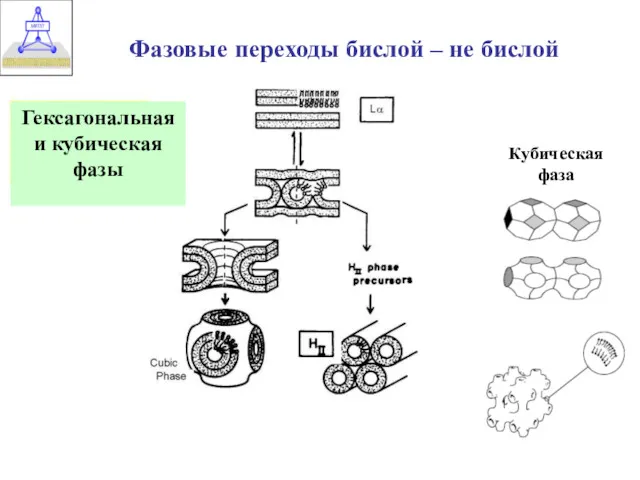
- 7. Фазовые переходы бислой – не бислой Гексагональная и кубическая фазы Кубическая фаза
- 8. Липидные агрегаты в воде Бислой Мицеллярная фаза Обращенные мицеллы Дискообразные мицеллы, бицеллы Мицеллы Ламеллярная фаза Гексагональная
- 9. Взаимосвязь лиотропного и термотропного мезоморфизма липидов в мембране Температура фазового перехода гель-жидкий кристалл Тф.п. зависит от
- 10. Фазовые диаграммы – однокомпонентных водно-липидных систем А – моногалактозилдиглицерид, Б - дигалактозилзиглицерид Дипальмитоилфосфатидилхолин в воде
- 11. Критический параметр упаковки липидов Структура полиморфной фазы (мезофазы) определяется динамической формой молекулы липида. Геометрические размеры молекул
- 12. Критический параметр упаковки липидов Полная поверхность мицеллы - M·Sо=4πR*2 Полный объем мицеллы - M·v=4/3πR*3 → Радиус
- 13. Полиморфные фазы, форма молекул и КПУ >1 ½ -1
- 14. Полиморфные фазы и форма молекул
- 15. Полиморфные фазы и динамическая форма молекул липидов
- 16. Методы изучения липидного полиморфизма - ЯМР – спектроскопия - Электронная микроскопия - Дифракционные методы
- 17. Метод Р ЯМР ФЛ – основные структурные элементы бислоя биомембран. ФЛ содержат, по крайней мере, один
- 18. Метод ЯМР на ядрах фосфора (31Р – ЯМР) 31Р-ЯМР – используют для изучения полиморфизма липидов с
- 19. Химический сдвиг сигналов зависит от ориентации фосфатной группы относительно линий напряженности магнитного поля. Электронная плотность вокруг
- 20. Метод Р ЯМР Химический сдвиг сигналов зависит от ориентации фосфатной группы относительно линий напряженности магнитного поля.
- 21. Метод Р ЯМР 31
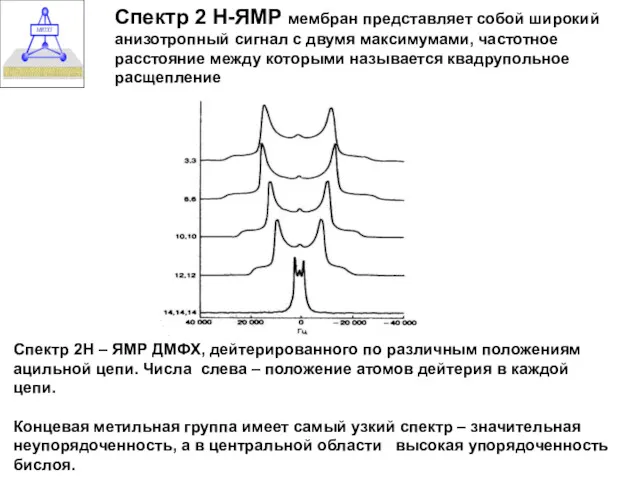
- 22. Метод Н ЯМР Спектр 2 Н-ЯМР мембран представляет собой широкий анизотропный сигнал с двумя максимумами, частотное
- 23. Спектр 2 Н-ЯМР мембран представляет собой широкий анизотропный сигнал с двумя максимумами, частотное расстояние между которыми
- 24. Структуры, которые образуются при диспергировании фосфолипидов в воде и соответствующие им спектры 31Р-ЯМР и 2Н-ЯМР
- 25. Электронная микроскопия Характеристика частиц по размеру и форме Методы: Просвечивающая микроскопия тонких срезов Замораживания-скалывания Негативное контрастирование
- 26. Просвечивающая электронная микроскопия тонких срезов 1950-е г.г. - Выявлена характерная трехслойная структура мембран, состоящая из 2-х
- 27. Метод замораживания-скалывания Усилия при скалывании замороженной клетки приводят к образованию среза, проходящего через образец. Оказалось, что
- 28. Метод «замораживания-скалывания» и «замораживания-травления» 1. Замораживание образца в жидком азоте (T=-196ºC, V=20ºC/мс) 2. Скалывание ножом-микротомом в
- 29. Электронные микрофотографии замороженных сколов Метод замораживания-скалывания
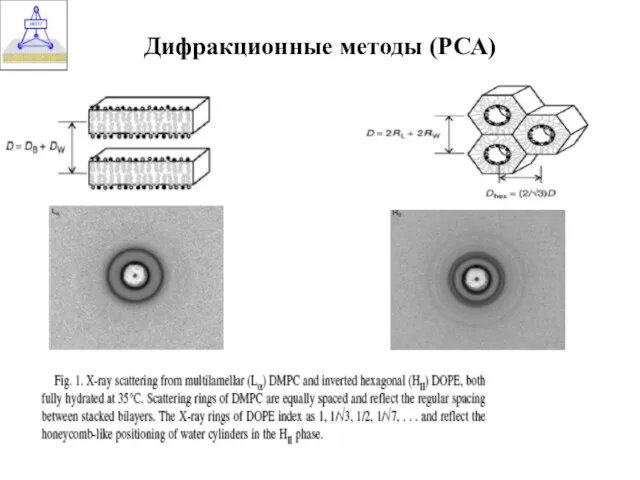
- 30. Дифракция рентгеновских лучей (РСА) Высокая упорядоченность кристаллических образцов – необходимое условие получения данных о структуре с
- 31. Дифракционные методы (РСА)
- 32. Электронная микроскопия «замораживание-скалывание» и 31Р –ЯМР спектроскопия Спектры 31Р-ЯМР водных дисперсий яичного фосфатидилэтаноламина (я-ФЭ) при различных
- 33. Спектры 31Р-ЯМР водных дисперсий яичного фосфатидилэтаноламина при различных температурах : а - 25º С - L
- 34. Полиморфизм отрицательно заряженных фосфолипидов Структурные перестройки водных дисперсий кардиолипина (КЛ) при добавлении ионов Са2+ 2 Na+
- 35. Полиморфизм смесей фосфолипидов ФЭ ФЭ/ФХ 85:15 ФЭ/ФХ 65:35 ФЭ/ФХ 50:50 Спектры 31Р-ЯМР водных дисперсий смесей соевого
- 36. Влияние белков на полиморфизм фосфолипидов Белки оказывают сильное влияние на полиморфизм ФЛ: - Гидрофобный полипептид грамицидин
- 37. Биологическое значение полиморфизма липидов Бислой – структурная основа биологических мембран НЕбислойные структуры в мембранах В настоящее
- 39. Скачать презентацию
Фазовые переходы липидов в мембранах
В водной среде липидные структуры ведут себя как анизотропные
Фазовые переходы липидов в мембранах
В водной среде липидные структуры ведут себя как анизотропные

Лиотропные жидкие кристаллы
образуются при смешении двух веществ, одно из которых является растворителем (липид-вода)
Лиотропные жидкие кристаллы
образуются при смешении двух веществ, одно из которых является растворителем (липид-вода)
Гидратация липидов
липид
+ Вода [мало ]
гидратированный липид
+ Вода
модификация полярной «головы»
липида: увеличение объема;
ослабление
Гидратация липидов
липид
+ Вода [мало ]
гидратированный липид
+ Вода
модификация полярной «головы»
липида: увеличение объема;
ослабление
![Гидратация липидов липид + Вода [мало ] гидратированный липид + Вода модификация полярной](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/146151/slide-3.jpg)
Лиотропный полиморфизм фосфолипидов
Полиморфизм – способность липидов образовывать в растворе агрегаты различной структуры
Фаза =
Лиотропный полиморфизм фосфолипидов
Полиморфизм – способность липидов образовывать в растворе агрегаты различной структуры
Фаза =
Фазовые переходы L ↔ H
Гексагональные фазы
НI - цилиндрические структуры, поверхность которых образована
Фазовые переходы L ↔ H
Гексагональные фазы
НI - цилиндрические структуры, поверхность которых образована
Фазовые переходы бислой – не бислой
Гексагональная
и кубическая
фазы
Кубическая
фаза
Фазовые переходы бислой – не бислой
Гексагональная
и кубическая
фазы
Кубическая
фаза
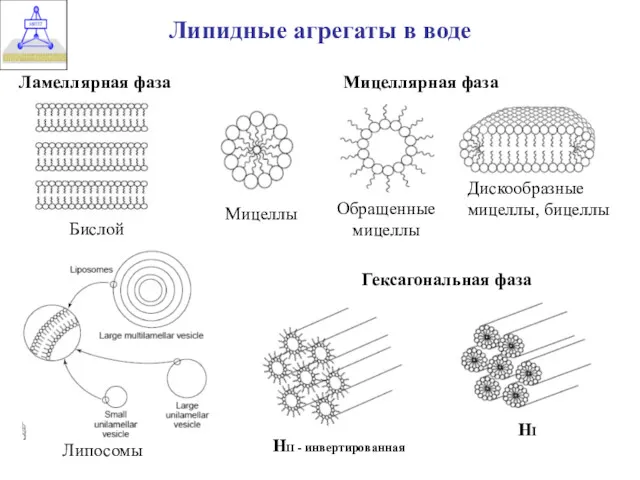
Липидные агрегаты в воде
Бислой
Мицеллярная фаза
Обращенные
мицеллы
Дискообразные
мицеллы, бицеллы
Мицеллы
Ламеллярная фаза
Гексагональная фаза
HI
HII - инвертированная
Липосомы
Липидные агрегаты в воде
Бислой
Мицеллярная фаза
Обращенные
мицеллы
Дискообразные
мицеллы, бицеллы
Мицеллы
Ламеллярная фаза
Гексагональная фаза
HI
HII - инвертированная
Липосомы
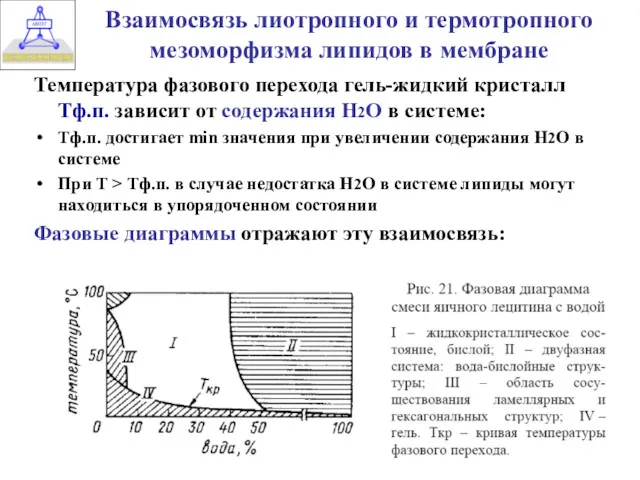
Взаимосвязь лиотропного и термотропного мезоморфизма липидов в мембране
Температура фазового перехода гель-жидкий кристалл Тф.п.
Взаимосвязь лиотропного и термотропного мезоморфизма липидов в мембране
Температура фазового перехода гель-жидкий кристалл Тф.п.
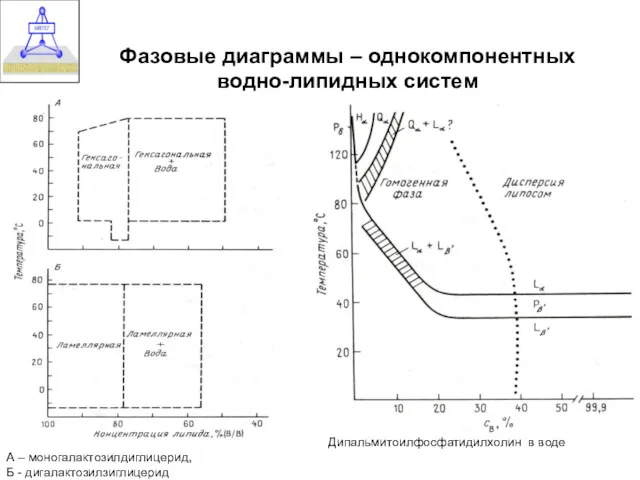
Фазовые диаграммы – однокомпонентных водно-липидных систем
А – моногалактозилдиглицерид,
Б - дигалактозилзиглицерид
Дипальмитоилфосфатидилхолин в воде
Фазовые диаграммы – однокомпонентных водно-липидных систем
А – моногалактозилдиглицерид,
Б - дигалактозилзиглицерид
Дипальмитоилфосфатидилхолин в воде
Критический параметр упаковки липидов
Структура полиморфной фазы (мезофазы) определяется динамической формой молекулы липида.
Геометрические
Критический параметр упаковки липидов
Структура полиморфной фазы (мезофазы) определяется динамической формой молекулы липида.
Геометрические

Критический параметр упаковки липидов
Полная поверхность мицеллы - M·Sо=4πR*2
Полный объем мицеллы - M·v=4/3πR*3 →
Радиус
Критический параметр упаковки липидов
Полная поверхность мицеллы - M·Sо=4πR*2
Полный объем мицеллы - M·v=4/3πR*3 →
Радиус

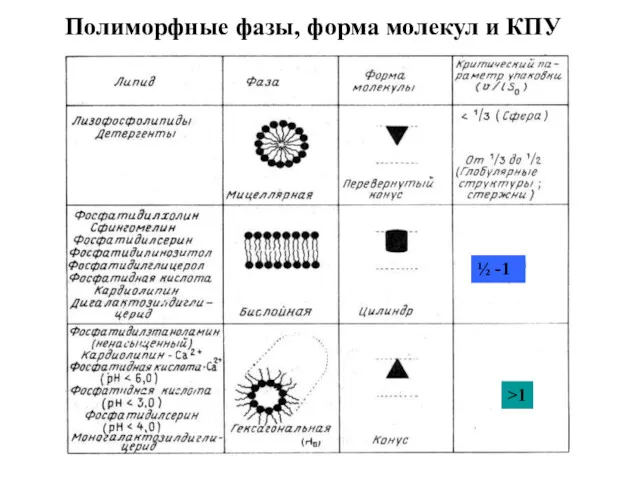
Полиморфные фазы, форма молекул и КПУ
>1
½ -1
Полиморфные фазы, форма молекул и КПУ
>1
½ -1
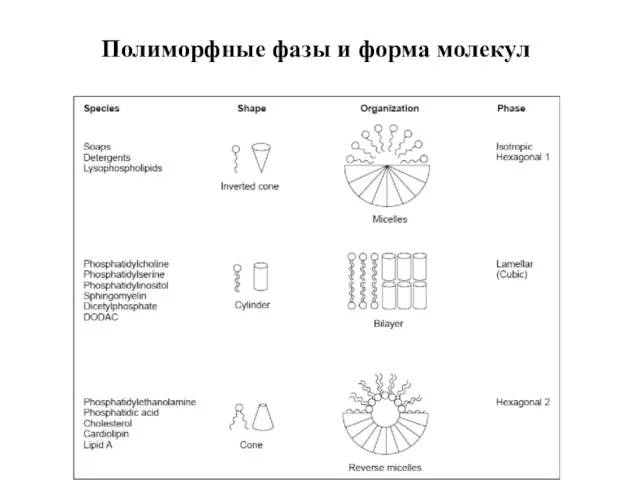
Полиморфные фазы и форма молекул
Полиморфные фазы и форма молекул
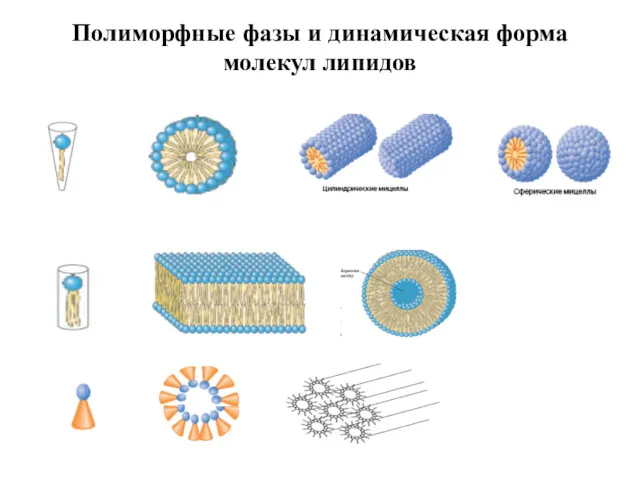
Полиморфные фазы и динамическая форма молекул липидов
Полиморфные фазы и динамическая форма молекул липидов
Методы изучения липидного полиморфизма
- ЯМР – спектроскопия
- Электронная микроскопия
- Дифракционные методы
Методы изучения липидного полиморфизма
- ЯМР – спектроскопия
- Электронная микроскопия
- Дифракционные методы

Метод Р ЯМР
ФЛ – основные структурные элементы бислоя биомембран.
ФЛ содержат, по крайней мере,
Метод Р ЯМР
ФЛ – основные структурные элементы бислоя биомембран.
ФЛ содержат, по крайней мере,

Метод ЯМР на ядрах фосфора (31Р – ЯМР)
31Р-ЯМР – используют для изучения полиморфизма
Метод ЯМР на ядрах фосфора (31Р – ЯМР)
31Р-ЯМР – используют для изучения полиморфизма

Химический сдвиг сигналов зависит от ориентации фосфатной группы относительно линий напряженности магнитного поля.
Химический сдвиг сигналов зависит от ориентации фосфатной группы относительно линий напряженности магнитного поля.
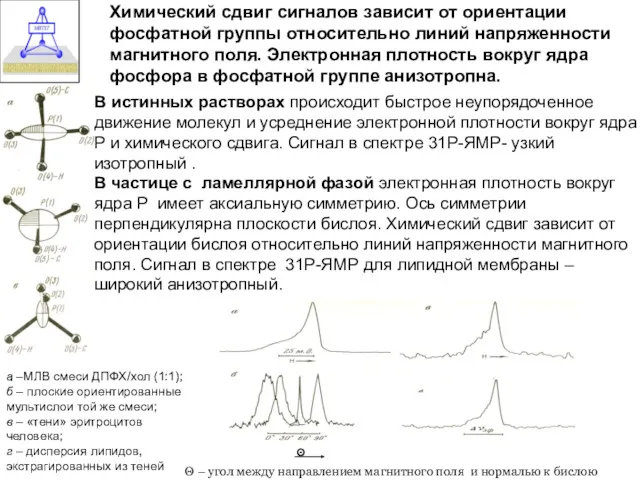
Метод Р ЯМР
Химический сдвиг сигналов зависит от ориентации фосфатной группы относительно линий напряженности
Метод Р ЯМР
Химический сдвиг сигналов зависит от ориентации фосфатной группы относительно линий напряженности

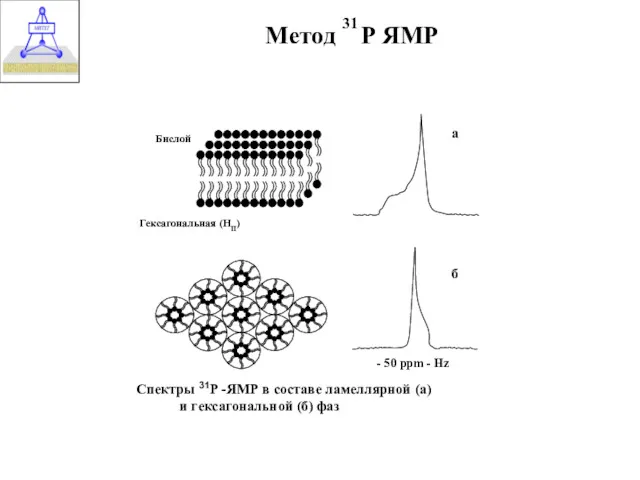
Метод Р ЯМР
31
Метод Р ЯМР
31
Метод Н ЯМР
Спектр 2 Н-ЯМР мембран представляет собой широкий анизотропный сигнал с двумя
Метод Н ЯМР
Спектр 2 Н-ЯМР мембран представляет собой широкий анизотропный сигнал с двумя

Спектр 2 Н-ЯМР мембран представляет собой широкий анизотропный сигнал с двумя максимумами, частотное
Спектр 2 Н-ЯМР мембран представляет собой широкий анизотропный сигнал с двумя максимумами, частотное
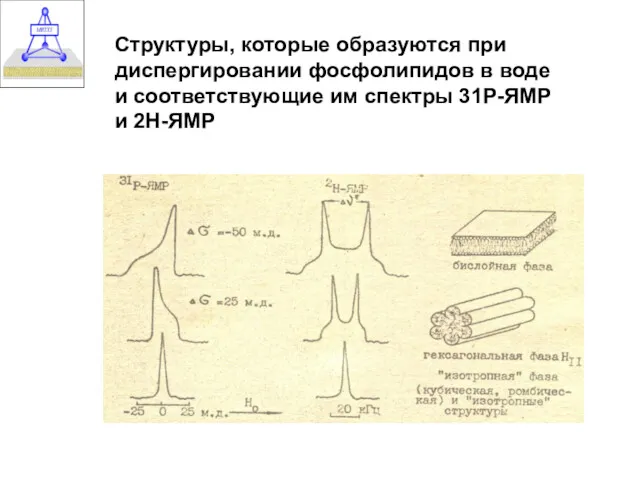
Структуры, которые образуются при диспергировании фосфолипидов в воде и соответствующие им спектры 31Р-ЯМР
Структуры, которые образуются при диспергировании фосфолипидов в воде и соответствующие им спектры 31Р-ЯМР
Электронная микроскопия
Характеристика частиц по размеру и форме
Методы:
Просвечивающая микроскопия тонких срезов
Замораживания-скалывания
Негативное контрастирование
Криоэлектронная микроскопия
Электронная микроскопия
Характеристика частиц по размеру и форме
Методы:
Просвечивающая микроскопия тонких срезов
Замораживания-скалывания
Негативное контрастирование
Криоэлектронная микроскопия

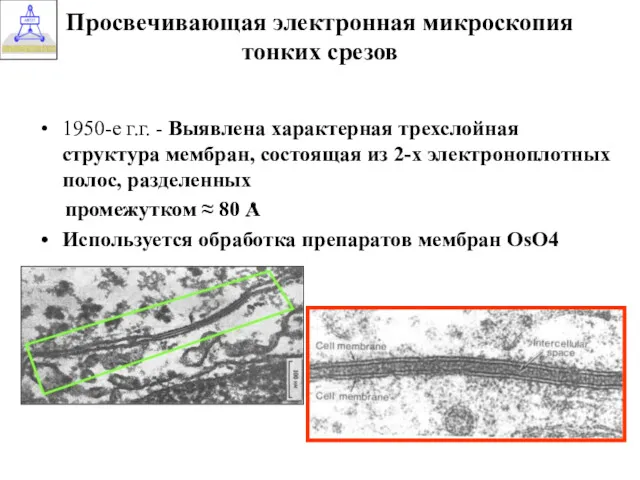
Просвечивающая электронная микроскопия тонких срезов
1950-е г.г. - Выявлена характерная трехслойная структура мембран, состоящая
Просвечивающая электронная микроскопия тонких срезов
1950-е г.г. - Выявлена характерная трехслойная структура мембран, состоящая
Метод замораживания-скалывания
Усилия при скалывании замороженной клетки приводят к образованию среза, проходящего через
Метод замораживания-скалывания
Усилия при скалывании замороженной клетки приводят к образованию среза, проходящего через

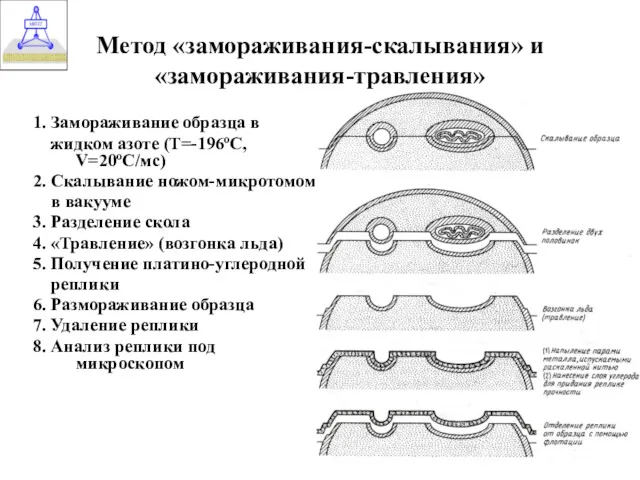
Метод «замораживания-скалывания» и «замораживания-травления»
1. Замораживание образца в
жидком азоте (T=-196ºC, V=20ºC/мс)
2.
Метод «замораживания-скалывания» и «замораживания-травления»
1. Замораживание образца в
жидком азоте (T=-196ºC, V=20ºC/мс)
2.
Электронные микрофотографии замороженных сколов
Метод замораживания-скалывания
Электронные микрофотографии замороженных сколов
Метод замораживания-скалывания

Дифракция рентгеновских лучей (РСА)
Высокая упорядоченность кристаллических образцов – необходимое условие получения
Дифракция рентгеновских лучей (РСА)
Высокая упорядоченность кристаллических образцов – необходимое условие получения

Дифракционные методы (РСА)
Дифракционные методы (РСА)
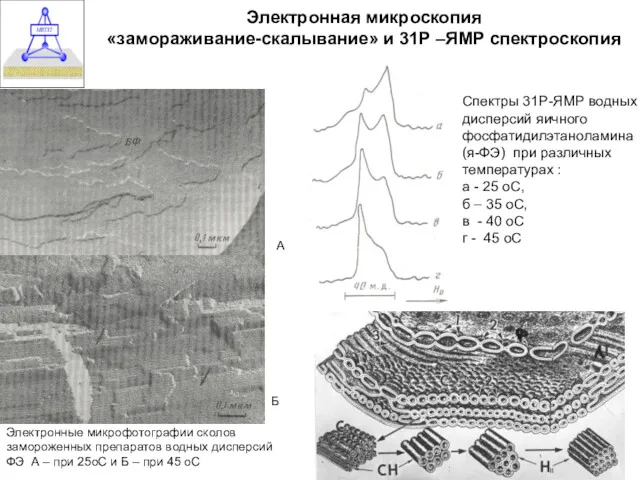
Электронная микроскопия
«замораживание-скалывание» и 31Р –ЯМР спектроскопия
Спектры 31Р-ЯМР водных дисперсий яичного фосфатидилэтаноламина (я-ФЭ)
Электронная микроскопия
«замораживание-скалывание» и 31Р –ЯМР спектроскопия
Спектры 31Р-ЯМР водных дисперсий яичного фосфатидилэтаноламина (я-ФЭ)
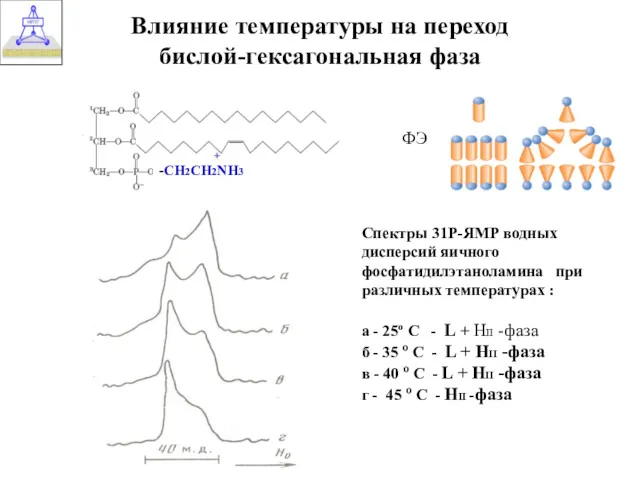
Спектры 31Р-ЯМР водных дисперсий яичного фосфатидилэтаноламина при различных температурах :
а - 25º
Спектры 31Р-ЯМР водных дисперсий яичного фосфатидилэтаноламина при различных температурах :
а - 25º
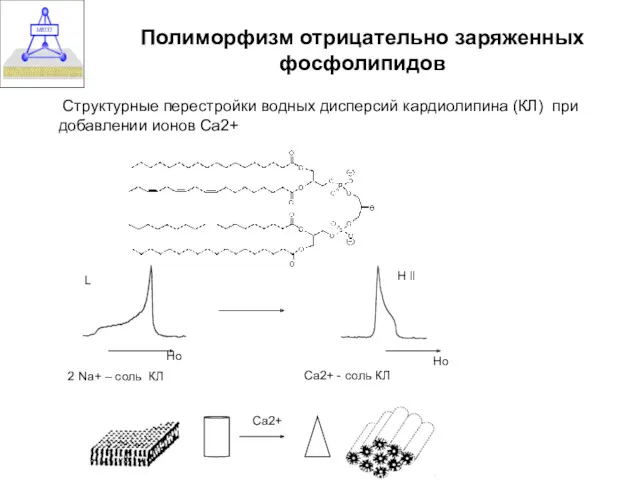
Полиморфизм отрицательно заряженных фосфолипидов
Структурные перестройки водных дисперсий кардиолипина (КЛ) при добавлении ионов
Полиморфизм отрицательно заряженных фосфолипидов
Структурные перестройки водных дисперсий кардиолипина (КЛ) при добавлении ионов
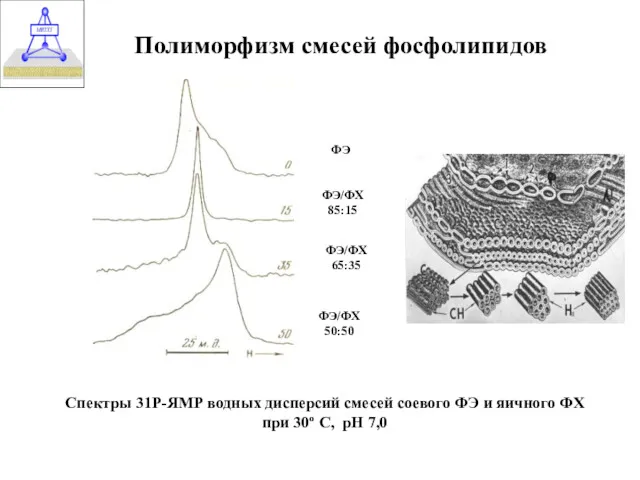
Полиморфизм смесей фосфолипидов
ФЭ
ФЭ/ФХ
85:15
ФЭ/ФХ
65:35
ФЭ/ФХ
50:50
Спектры 31Р-ЯМР водных дисперсий смесей соевого ФЭ и яичного ФХ
Полиморфизм смесей фосфолипидов
ФЭ
ФЭ/ФХ
85:15
ФЭ/ФХ
65:35
ФЭ/ФХ
50:50
Спектры 31Р-ЯМР водных дисперсий смесей соевого ФЭ и яичного ФХ
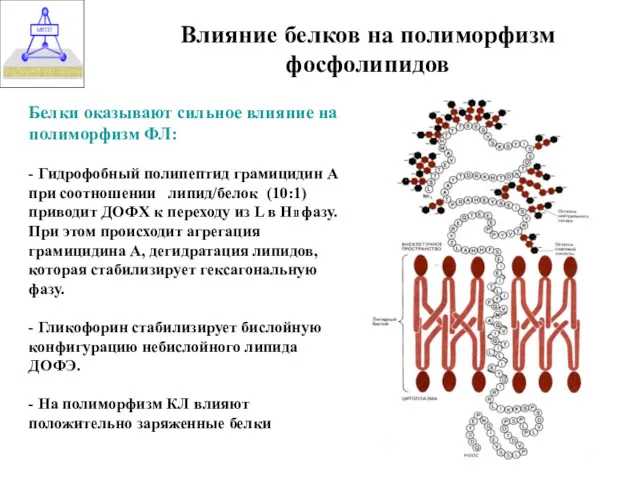
Влияние белков на полиморфизм фосфолипидов
Белки оказывают сильное влияние на полиморфизм ФЛ:
- Гидрофобный полипептид
Влияние белков на полиморфизм фосфолипидов
Белки оказывают сильное влияние на полиморфизм ФЛ:
- Гидрофобный полипептид
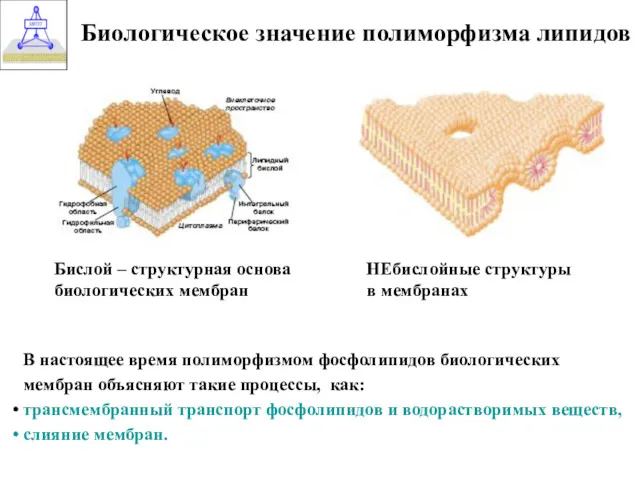
Биологическое значение полиморфизма липидов
Бислой – структурная основа
биологических мембран
НЕбислойные структуры
в мембранах
В
Биологическое значение полиморфизма липидов
Бислой – структурная основа
биологических мембран
НЕбислойные структуры
в мембранах
В
 Понимание биологии. (Часть 2)
Понимание биологии. (Часть 2) Биология - наука о живом мире. Общие закономерности жизни
Биология - наука о живом мире. Общие закономерности жизни Модификационная изменчивость
Модификационная изменчивость Критерии вида (биологические задачи)
Критерии вида (биологические задачи) Презентация Здоровье – ключ к успеху
Презентация Здоровье – ключ к успеху Як змінюється навесні життя риб?
Як змінюється навесні життя риб? Асқорыту жүйесі
Асқорыту жүйесі Разнообразие насекомых
Разнообразие насекомых Терморегуляция
Терморегуляция История развития генетики. Основные понятия
История развития генетики. Основные понятия Перешеек ромбовидного мозга. Средний и промежуточный мозг
Перешеек ромбовидного мозга. Средний и промежуточный мозг Непрямой онтогенез
Непрямой онтогенез Inorganic substances. Water. Mineral salts
Inorganic substances. Water. Mineral salts Тварини-рекордсмени
Тварини-рекордсмени Презентация Отряд Перепончатокрылые
Презентация Отряд Перепончатокрылые Развитие пищеварительной системы, оболочки пищеварительной трубки. Слизистая оболочка полости рта
Развитие пищеварительной системы, оболочки пищеварительной трубки. Слизистая оболочка полости рта Газообмен. Диффузия свободных молекул газа (O2 и СО2)
Газообмен. Диффузия свободных молекул газа (O2 и СО2) Антигени. Класифікація антигенів
Антигени. Класифікація антигенів Человек как уникальный вид живой природы
Человек как уникальный вид живой природы Биологическое действие радиации
Биологическое действие радиации Хвойные растения
Хвойные растения Яблони. Классификация сортов
Яблони. Классификация сортов Презентация к уроку Тип Плоские черви
Презентация к уроку Тип Плоские черви Влияние хозяйственной деятельности на окружающую среду
Влияние хозяйственной деятельности на окружающую среду ГМО. Еда. Питание
ГМО. Еда. Питание Наука биология
Наука биология Сердечнососудистая система
Сердечнососудистая система Растения Дальневосточного региона
Растения Дальневосточного региона