- Понимание биологии. (Часть 2)

Содержание
- 2. Я - не любитель слова «концепция», я – любитель слова «ясность» Однако речь пойдёт о концепциях,
- 3. Надеюсь, что всё будет понятно.
- 4. «Мало кто знает сегодня, что в начале ХХ века микробиология могла пойти другим путём. В этом
- 5. «Главные результаты микробиологии являются «полным позорищем», неудачной наукоподобной попыткой познать основополагающие принципы организации живого» Stanier R.Y.,
- 6. Об одном умалчиваемом тупике «И вот в середине ХХ века микробиология зашла в тупик, — одной
- 7. Объяснение с единой точки зрения: Целостный взгляд на прокариотно-вирусное единство. Почему существование бактерий и архей без
- 8. Но главное в другом… Эта презентация о том, как мы сильно недооцениваем роль процессов, которые как
- 9. Данное обобщение позволяет предсказать всеобщий характер распространения бактериофагов в бактериальных сообществах Предсказание: без бактериофагов большинство прокариотических
- 10. Для контраста вернёмся ненадолго в мир эвкариот, где картина кажется более ясной.
- 11. Вспомним одно фундаментальное обобщение Копирование генетической информации обязательно происходит с теми или иными ошибками. Например, человеческий
- 12. Даже слабовредные мутации при клональном размножении будут неизбежно накапливаться. Вступает в действие так называемый Храповик Меллера.
- 13. Правило Мёллера (Muller 1950) Смертность потомков в половых популяциях будет меньше, чем в бесполых. Для удаления
- 14. Рекомбинация порождает разнообразие: искусство немножко отличаться Делает некоторых потомков исключительными, некоторых – безнадёжными, некоторых – очень
- 15. У эвкариот половой процесс и его аналоги являются правилом для размножения. Многочисленные модели, объясняющие роль полового
- 16. В случае прокариот процессы рекомбинации на первый взгляд кажутся менее регулярными и более хаотичными. НА САМОМ
- 17. Постепенно возникает новое более цельное понимание того, как в мире прокариот поддерживается генетическая стабильность видов.
- 18. У прокариот. У прокариот рекомбинация выглядит более случайной. При рекомбинации часть генетического материала, как правило, теряется.
- 19. Однако… Понимание того, как прокариоты осуществляют рекомбинацию возникло не сразу. Поначалу большое внимание уделялось такому хорошо
- 20. Пойдём от частного к общему. Как конъюгация происходит у кишечной палочки?
- 21. Конъюгация у бактерий Перенос генетического материала от одной бактериальной клетки (донора) к другой (реципиенту) при их
- 22. Ледерберг и Татум доказали, что при конъюгации происходит рекомбинация генетической информации Дж. Ледерберг Э. Татум
- 23. В основе их доказательства лежало использование ауксотрофных сред Культуры, растущие на наиболее бедной питательной среде из
- 24. Met- Bio- Thr- Leu- Thi- Смешанная культура Клетки высевали на чашки с минимальной средой. В результате
- 25. Пили бывают разные… Помимо жгутиков, бактериальная клетка может нести тонкие отростки – пили. Обычно они нужны,
- 26. Пили бывают разные…
- 27. Донорные, мужские клетки, содержащие F-плазмиду, имеют от 1 до 3 пилей. Пили имеют осевой канал. Они
- 29. Обычно конъюгация рассматривается на одном частном примере у E. coli, хотя её механизмы очень разнообразны.
- 30. После установления контакта пили сокращаются и клетка входит в тесный контакт. В сайт плазмиды оri (точка
- 31. В донорной и реципиентной клетках синтезируется комплементарная нить. В реципиентной клетке линейная ДНК замыкается в кольцо.
- 32. Но здесь же просто перенос плазмиды, а где же рекомбинация хромосомного материала?
- 33. F-плазмида может интегрировать в хромосому. При ее переносе реципиентную клетку также будет происходить перенос хромосомной ДНК.
- 34. Фундаментальный факт Если в бактериальную клетку проникнет фрагмент ДНК, гомологичный участку хромосомы, то с высокой частотой
- 35. Почему при мейозе у эвкариота возможен только один кроссинговер, а при конъюгации у прокариот обязательно хотя
- 36. Одиночный кроссинговер привел бы к образованию линейной молекулы и гибели клетки
- 37. Очень важное замечание F-фактор ведет себя как геномный паразит, а конъюгация и f-пили несомненно являются всего
- 38. Насколько распространена конъюгация у бактерий? Обнаружены разные механизмы конъюгации, однако лишь у небольшой части бактерий. Встречается
- 39. Наряду с конъюгативными плазмидами в рекомбинации могут участвовать конъюгативные транспозоны В отличие от плазмид конъюгативные транспозоны
- 40. Системы конъюгативного переноса у грамположительных бактерий более разнообразны. Как правило, клетки не образуют пилей, а просто
- 41. Разнообразие конъюгативного переноса
- 42. Насколько распространена конъюгация? Даже у хорошо изученных видов бактерий в последние годы обнаруживают разные виды конъюгаций.
- 43. Первый намёк на понимание возник в связи с открытием явления трасформации у бактерий. Эта история началась
- 44. Ф. Гриффитс и О. Эвери инъецировали мышам культуру живого пневмококка R-штамма вместе с S-штаммом, убитым высокой
- 45. Живые S-клетки (контроль) Живые R-клетки (контроль) Убитые Нагреванием S-клетки Смесь убитых Нагреванием S-клеток и живых R-клеток
- 46. Трансформация - направленный перенос и встраивание в генетический аппарат клетки небольшого фрагмента чужеродной ДНК, происходящая участия
- 47. В 90-е годы прошлого века стало ясно, что трансформация важна и для того, чтобы организмы в
- 48. Однако… Как увидим ниже, трансформация лежит в основе и другого фундаментального явления, связанного с избавлением от
- 49. Как перенос генов спасает популяцию
- 50. Инфекция вирусами – это прежде всего критически необходимое для прокариот приобретение эволюции.
- 51. Трансдукция - процесс переноса бактериальной ДНК из одной клетки в другую бактериофагом. Попадая в другую бактериальную
- 53. Таким образом, фаги оказываются полезными для популяций бактерий. Фаги, убивая часть бактерий, переносят их ДНК, обеспечивая
- 54. А теперь внимание!!! Принципиально новая идея №1! Задача на логическое воображение: могут ли эти два процесса
- 55. ТРАНСФОРМАЦИЯ ТРАНСДУКЦИЯ
- 56. Ответ – ДА! Лизис фагом прокариотической клетки приводит к образованию свободной бактериальной ДНК, которая может трансформировать.
- 57. ТРАНСФОРМАЦИЯ ТРАНСДУКЦИЯ
- 58. Попробуем кое в чём посомневаться. «Наука — не место для догм. Учёный имеет право и обязан
- 59. Какой процесс более значим? Казалось бы, фаги, благодаря специфичности инфицирования должны быть более важны для осуществления
- 60. Система рестрикции-модификации обеспечивается двумя типами ферментов. Первый тип – «ножницы», которые режут ДНК по определённым последовательностями
- 61. • Рестриктаза расщепляет чужеродную ДНК по определённому сайту узнавания. • Собственная ДНК клетки не расщепляется рестриктазой,
- 63. Кстати: Даже порезанная ДНК из фага может участвовать в рекомбинации.
- 64. Почему рестриктазы так называются? Система рестрикции-модификации предотвращает фаговую инфекции не на 100%, а только уменьшает её
- 65. Нетривиальная идея №2! В научной и учебной литературе чаще всего пишут, что системы рестрикции-модификации нужны для
- 66. Система рестрикции модификации помогает осуществлять преимущественно рекомбинацию только между очень родственными штаммами! Фаги с определённой частотой
- 68. В настоящее время общепризнанным является тот фундаментальный факт, что бактериофаги и археофаги играют ключевую роль в
- 69. Это утверждение может быть дополнено не менее фундаментальным обобщением: массовый лизис бактерий – критически необходимый элемент
- 70. Бактерии (в прямоугольнике) и вирусные частицы (овал) из морской воды.
- 71. Таким образом Фаги оказываются исключительно полезными не только благодаря тому, что они сами (быть может таксоноспецифично)
- 72. Бактериофаги и археофаги – это не гибель прокариотических популяций, а критически необходимый элемент их выживания
- 73. Бактериально-архейное-вирусное единство
- 75. Программируемый лизис клетки – альтернатива фаговому лизису.
- 83. У Bacillus subtilis есть простая генетическая система, приводящая к лизису половины популяции. http://elementy.ru/news/430139
- 84. Программируемый лизис бактериальных клеток необходим для рекомбинации
- 85. P.S. Ещё один недооцененный возможно распространённейший механизм спасения прокариот от Храповика Мёллера «Бактерии гаплоидны» - такая
- 86. Полиплоидия – возможность частично избавиться от храповика Меллера. Нередко сестринские хромосомы в прокариотах рекомбинируют, давая возможность
- 87. Почему Deinococcus radiodurans способен выдерживать чудовищные дозы радиации? Дейнококк является тетраплоидом — в его клетках присутствует
- 91. Скачать презентацию

Я - не любитель слова «концепция», я – любитель слова «ясность»
Однако
Я - не любитель слова «концепция», я – любитель слова «ясность»
Однако

Надеюсь, что всё будет понятно.
Надеюсь, что всё будет понятно.

«Мало кто знает сегодня, что в начале ХХ века микробиология могла пойти другим
«Мало кто знает сегодня, что в начале ХХ века микробиология могла пойти другим
«Главные результаты микробиологии являются «полным позорищем», неудачной наукоподобной попыткой познать основополагающие
«Главные результаты микробиологии являются «полным позорищем», неудачной наукоподобной попыткой познать основополагающие

Об одном умалчиваемом тупике
«И вот в середине ХХ века микробиология зашла в тупик, — одной только
Об одном умалчиваемом тупике
«И вот в середине ХХ века микробиология зашла в тупик, — одной только

Объяснение с единой точки зрения:
Целостный взгляд на прокариотно-вирусное единство.
Почему существование бактерий
Объяснение с единой точки зрения:
Целостный взгляд на прокариотно-вирусное единство.
Почему существование бактерий

Но главное в другом…
Эта презентация о том, как мы сильно недооцениваем
Но главное в другом…
Эта презентация о том, как мы сильно недооцениваем

Данное обобщение позволяет предсказать всеобщий характер распространения бактериофагов в бактериальных сообществах
Предсказание:
Данное обобщение позволяет предсказать всеобщий характер распространения бактериофагов в бактериальных сообществах
Предсказание:

Для контраста вернёмся ненадолго в мир эвкариот, где картина кажется более
Для контраста вернёмся ненадолго в мир эвкариот, где картина кажется более

Вспомним одно фундаментальное обобщение
Копирование генетической информации обязательно происходит с теми или
Вспомним одно фундаментальное обобщение
Копирование генетической информации обязательно происходит с теми или

Даже слабовредные мутации при клональном размножении будут неизбежно накапливаться.
Вступает в
Даже слабовредные мутации при клональном размножении будут неизбежно накапливаться.
Вступает в

Правило Мёллера (Muller 1950)
Смертность потомков в половых популяциях будет меньше, чем
Правило Мёллера (Muller 1950)
Смертность потомков в половых популяциях будет меньше, чем

Рекомбинация порождает разнообразие: искусство немножко отличаться
Делает некоторых потомков исключительными, некоторых –
Рекомбинация порождает разнообразие: искусство немножко отличаться
Делает некоторых потомков исключительными, некоторых –

У эвкариот половой процесс и его аналоги являются правилом для размножения.
Многочисленные
У эвкариот половой процесс и его аналоги являются правилом для размножения.
Многочисленные

В случае прокариот процессы рекомбинации на первый взгляд кажутся менее регулярными
В случае прокариот процессы рекомбинации на первый взгляд кажутся менее регулярными

Постепенно возникает новое более цельное понимание того, как в мире прокариот
Постепенно возникает новое более цельное понимание того, как в мире прокариот

У прокариот.
У прокариот рекомбинация выглядит более случайной.
При рекомбинации часть генетического
У прокариот.
У прокариот рекомбинация выглядит более случайной.
При рекомбинации часть генетического

Однако…
Понимание того, как прокариоты осуществляют рекомбинацию возникло не сразу.
Поначалу большое
Однако…
Понимание того, как прокариоты осуществляют рекомбинацию возникло не сразу.
Поначалу большое

Пойдём от частного к общему.
Как конъюгация происходит у кишечной палочки?
Пойдём от частного к общему.
Как конъюгация происходит у кишечной палочки?

Конъюгация у бактерий
Перенос генетического материала от одной бактериальной клетки (донора) к
Конъюгация у бактерий
Перенос генетического материала от одной бактериальной клетки (донора) к

Ледерберг и Татум доказали, что при конъюгации происходит рекомбинация генетической информации
Дж.
Ледерберг и Татум доказали, что при конъюгации происходит рекомбинация генетической информации
Дж.

В основе их доказательства лежало использование ауксотрофных сред
Культуры, растущие на наиболее
В основе их доказательства лежало использование ауксотрофных сред
Культуры, растущие на наиболее
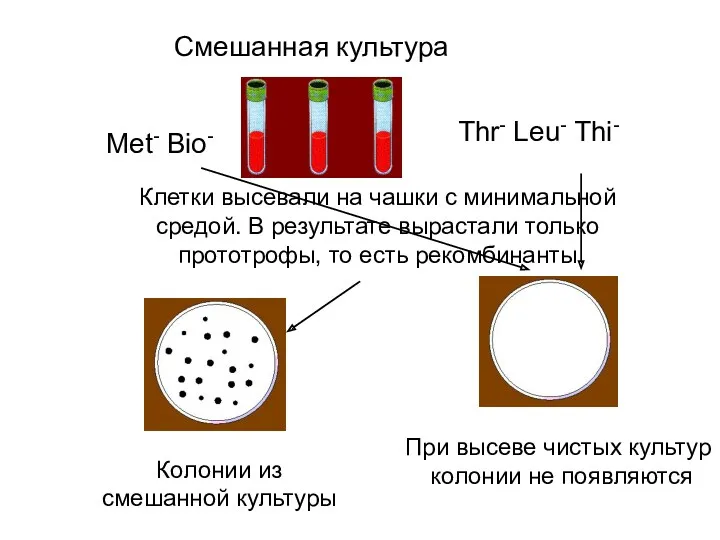
Met- Bio-
Thr- Leu- Thi-
Смешанная культура
Клетки высевали на чашки с минимальной средой.
Met- Bio-
Thr- Leu- Thi-
Смешанная культура
Клетки высевали на чашки с минимальной средой.

Пили бывают разные…
Помимо жгутиков, бактериальная клетка может нести тонкие отростки –
Пили бывают разные…
Помимо жгутиков, бактериальная клетка может нести тонкие отростки –
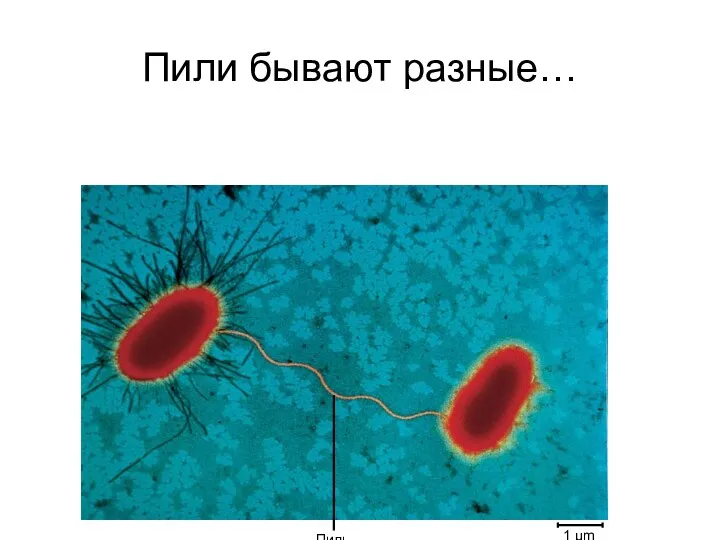
Пили бывают разные…
Пили бывают разные…

Донорные, мужские клетки, содержащие F-плазмиду, имеют от 1 до 3 пилей.
Пили
Донорные, мужские клетки, содержащие F-плазмиду, имеют от 1 до 3 пилей.
Пили
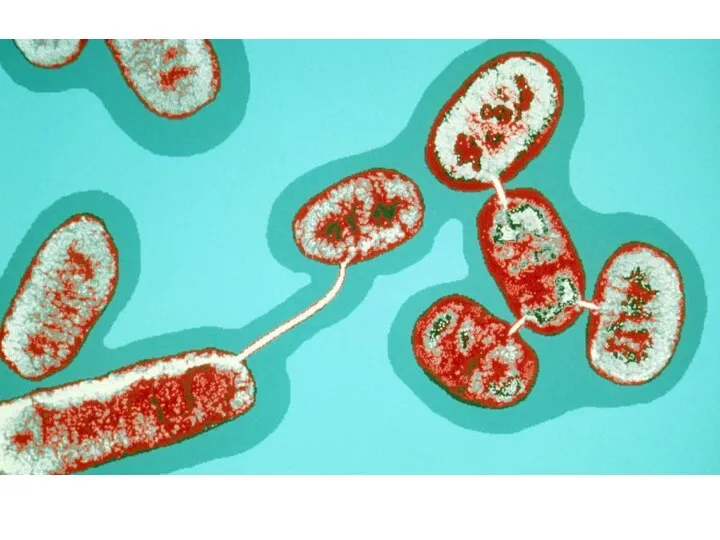

Обычно конъюгация рассматривается на одном частном примере у E. coli, хотя
Обычно конъюгация рассматривается на одном частном примере у E. coli, хотя
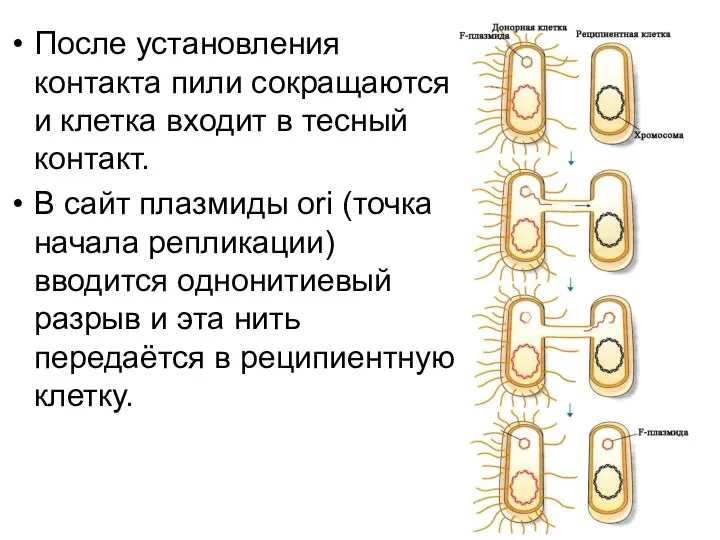
После установления контакта пили сокращаются и клетка входит в тесный контакт.
В
После установления контакта пили сокращаются и клетка входит в тесный контакт.
В
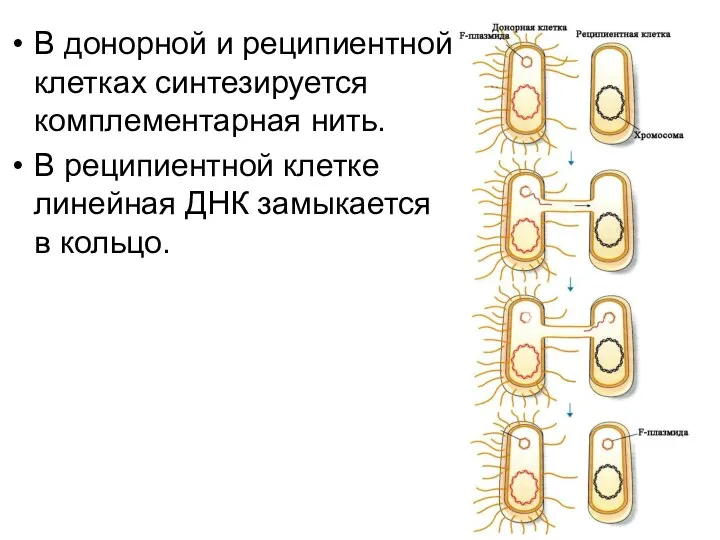
В донорной и реципиентной клетках синтезируется комплементарная нить.
В реципиентной клетке
В донорной и реципиентной клетках синтезируется комплементарная нить.
В реципиентной клетке

Но здесь же просто перенос плазмиды, а где же рекомбинация хромосомного
Но здесь же просто перенос плазмиды, а где же рекомбинация хромосомного
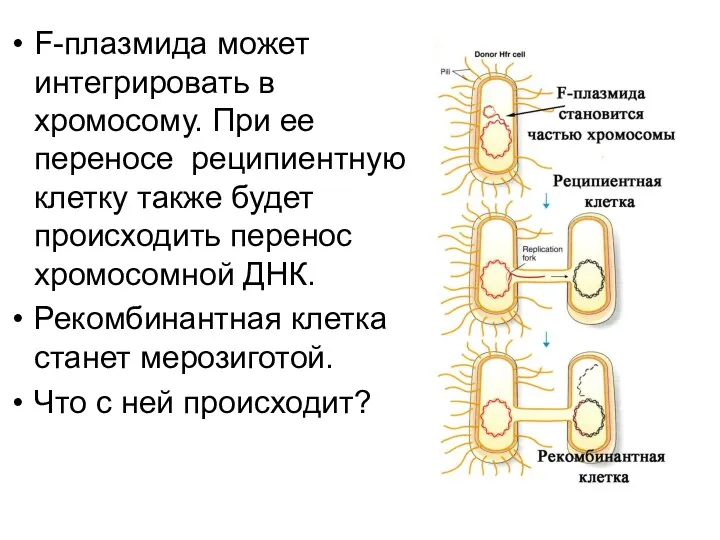
F-плазмида может интегрировать в хромосому. При ее переносе реципиентную клетку также
F-плазмида может интегрировать в хромосому. При ее переносе реципиентную клетку также

Фундаментальный факт
Если в бактериальную клетку проникнет фрагмент ДНК, гомологичный участку хромосомы,
Фундаментальный факт
Если в бактериальную клетку проникнет фрагмент ДНК, гомологичный участку хромосомы,
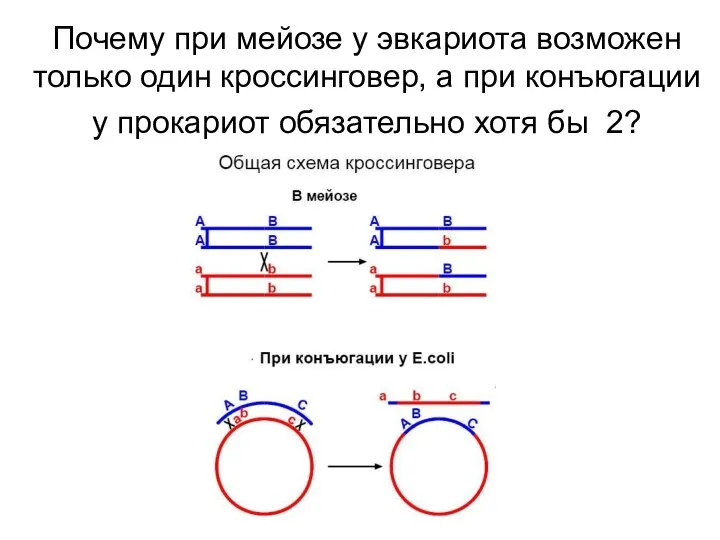
Почему при мейозе у эвкариота возможен только один кроссинговер, а при
Почему при мейозе у эвкариота возможен только один кроссинговер, а при
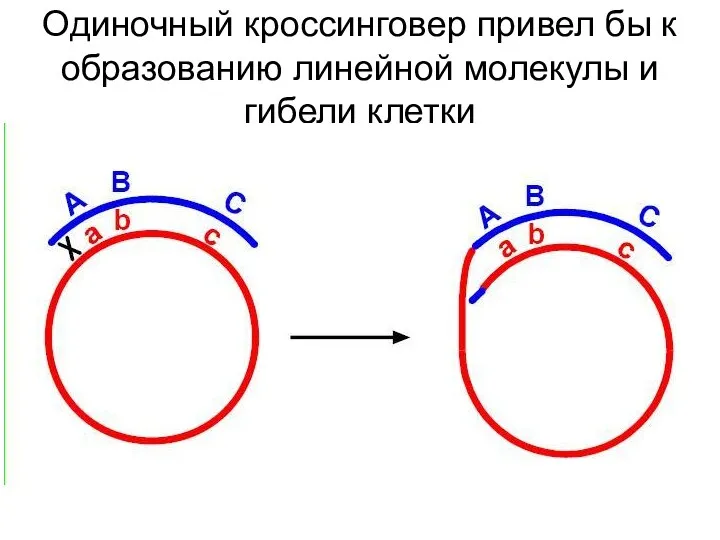
Одиночный кроссинговер привел бы к образованию линейной молекулы и гибели клетки
Одиночный кроссинговер привел бы к образованию линейной молекулы и гибели клетки

Очень важное замечание
F-фактор ведет себя как геномный паразит, а конъюгация и
Очень важное замечание
F-фактор ведет себя как геномный паразит, а конъюгация и

Насколько распространена конъюгация у бактерий?
Обнаружены разные механизмы конъюгации, однако лишь у
Насколько распространена конъюгация у бактерий?
Обнаружены разные механизмы конъюгации, однако лишь у

Наряду с конъюгативными плазмидами в рекомбинации могут участвовать конъюгативные транспозоны
В отличие
Наряду с конъюгативными плазмидами в рекомбинации могут участвовать конъюгативные транспозоны
В отличие

Системы конъюгативного переноса у грамположительных бактерий более разнообразны.
Как правило, клетки не
Системы конъюгативного переноса у грамположительных бактерий более разнообразны.
Как правило, клетки не
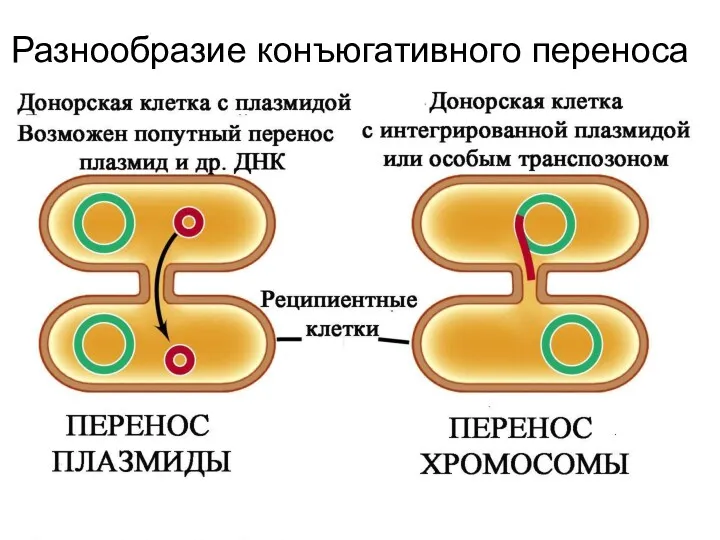
Разнообразие конъюгативного переноса
Разнообразие конъюгативного переноса

Насколько распространена конъюгация?
Даже у хорошо изученных видов бактерий в последние годы
Насколько распространена конъюгация?
Даже у хорошо изученных видов бактерий в последние годы

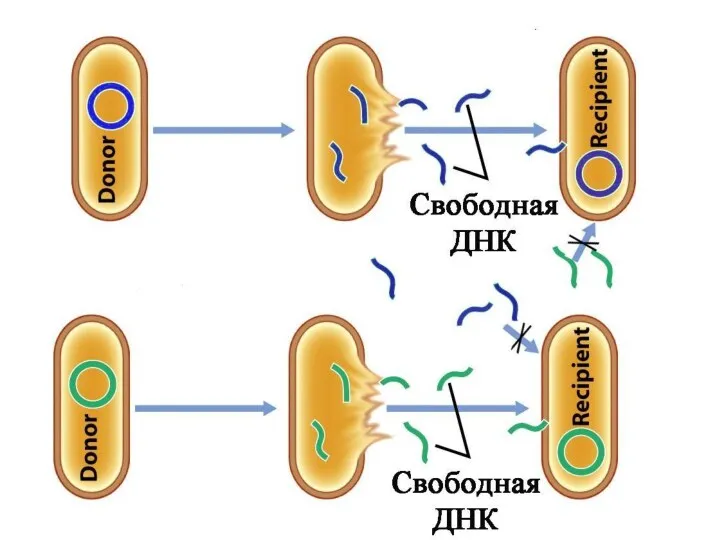
Первый намёк на понимание возник в связи с открытием явления трасформации
Первый намёк на понимание возник в связи с открытием явления трасформации

Ф. Гриффитс и О. Эвери инъецировали мышам культуру живого пневмококка R-штамма
Ф. Гриффитс и О. Эвери инъецировали мышам культуру живого пневмококка R-штамма
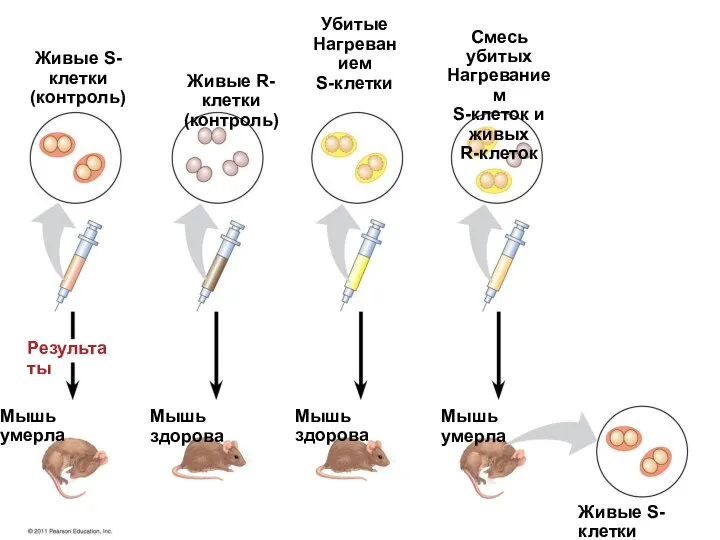
Живые S-клетки
(контроль)
Живые R-клетки
(контроль)
Убитые
Нагреванием
S-клетки
Смесь убитых
Нагреванием
S-клеток и живых
R-клеток
Мышь умерла
Мышь умерла
Мышь здорова
Мышь
Живые S-клетки
(контроль)
Живые R-клетки
(контроль)
Убитые
Нагреванием
S-клетки
Смесь убитых
Нагреванием
S-клеток и живых
R-клеток
Мышь умерла
Мышь умерла
Мышь здорова
Мышь

Трансформация - направленный перенос и встраивание в генетический аппарат клетки небольшого
Трансформация - направленный перенос и встраивание в генетический аппарат клетки небольшого

В 90-е годы прошлого века стало ясно, что трансформация важна и
В 90-е годы прошлого века стало ясно, что трансформация важна и

Однако…
Как увидим ниже, трансформация лежит в основе и другого фундаментального явления,
Однако…
Как увидим ниже, трансформация лежит в основе и другого фундаментального явления,
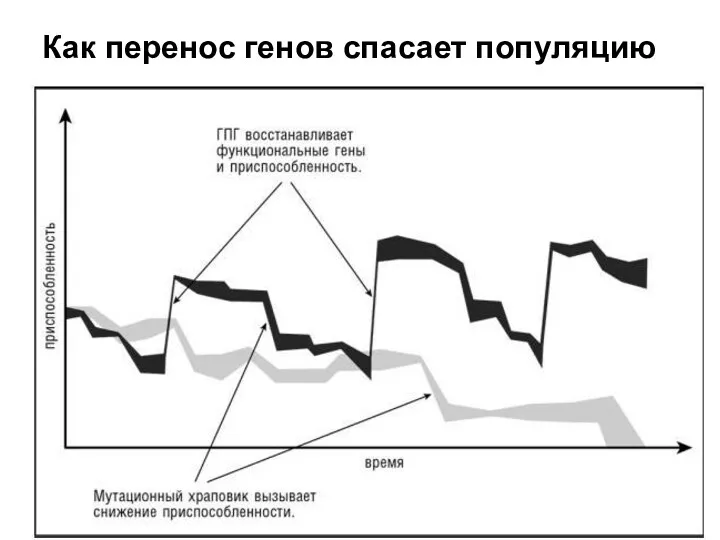
Как перенос генов спасает популяцию
Как перенос генов спасает популяцию

Инфекция вирусами – это прежде всего критически необходимое для прокариот приобретение
Инфекция вирусами – это прежде всего критически необходимое для прокариот приобретение

Трансдукция - процесс переноса бактериальной ДНК из одной клетки в другую
Трансдукция - процесс переноса бактериальной ДНК из одной клетки в другую
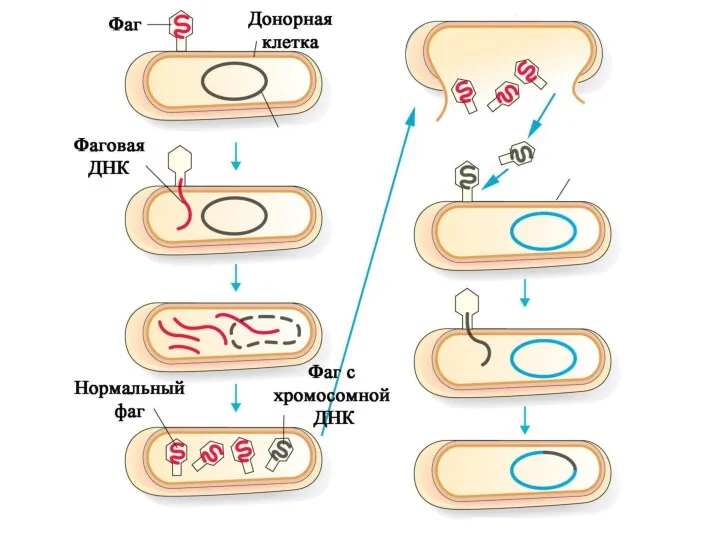

Таким образом, фаги оказываются полезными для популяций бактерий.
Фаги, убивая часть бактерий,
Таким образом, фаги оказываются полезными для популяций бактерий.
Фаги, убивая часть бактерий,

А теперь внимание!!!
Принципиально новая идея №1!
Задача на логическое воображение: могут ли
А теперь внимание!!!
Принципиально новая идея №1!
Задача на логическое воображение: могут ли
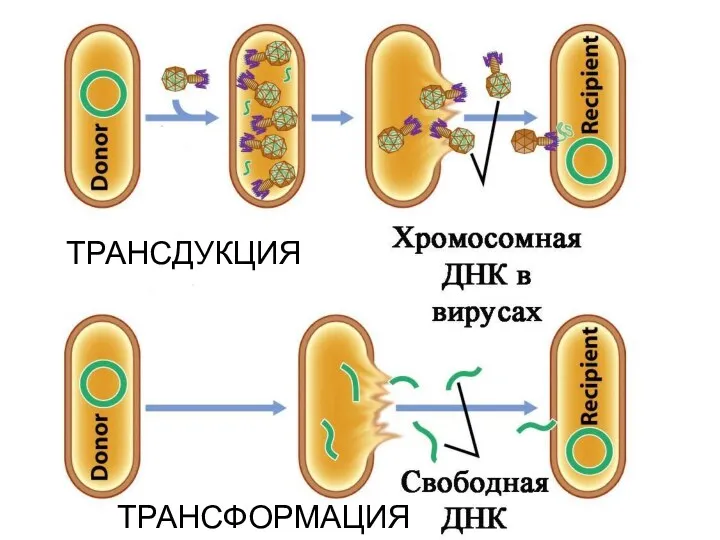
ТРАНСФОРМАЦИЯ
ТРАНСДУКЦИЯ
ТРАНСФОРМАЦИЯ
ТРАНСДУКЦИЯ

Ответ – ДА!
Лизис фагом прокариотической клетки приводит к образованию свободной бактериальной
Ответ – ДА!
Лизис фагом прокариотической клетки приводит к образованию свободной бактериальной
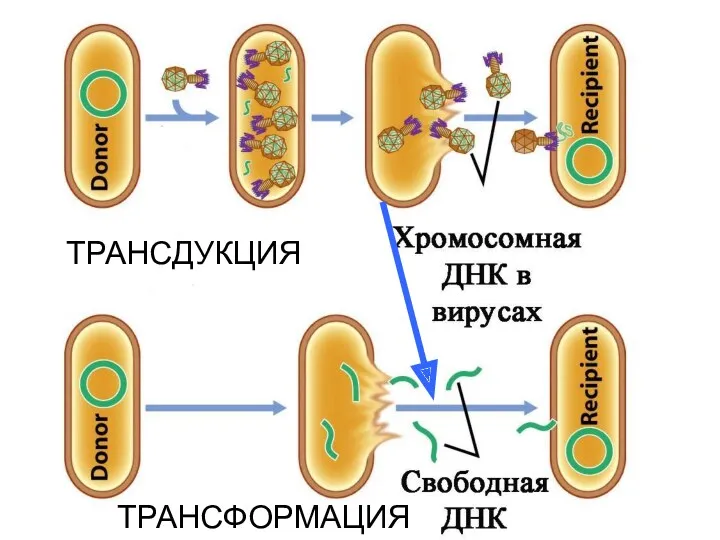
ТРАНСФОРМАЦИЯ
ТРАНСДУКЦИЯ
ТРАНСФОРМАЦИЯ
ТРАНСДУКЦИЯ

Попробуем кое в чём посомневаться.
«Наука — не место для догм. Учёный имеет право
Попробуем кое в чём посомневаться.
«Наука — не место для догм. Учёный имеет право

Какой процесс более значим?
Казалось бы, фаги, благодаря специфичности инфицирования должны быть
Какой процесс более значим?
Казалось бы, фаги, благодаря специфичности инфицирования должны быть

Система рестрикции-модификации обеспечивается двумя типами ферментов.
Первый тип – «ножницы», которые
Система рестрикции-модификации обеспечивается двумя типами ферментов.
Первый тип – «ножницы», которые
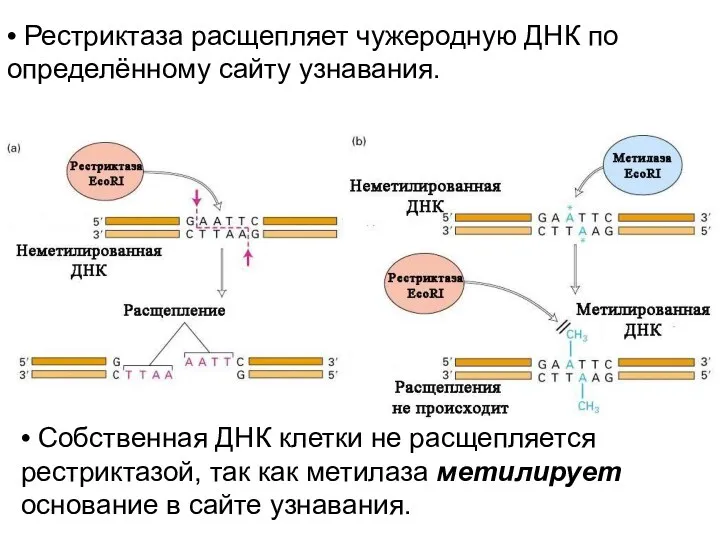
• Рестриктаза расщепляет чужеродную ДНК по определённому сайту узнавания.
• Собственная ДНК
• Рестриктаза расщепляет чужеродную ДНК по определённому сайту узнавания.
• Собственная ДНК
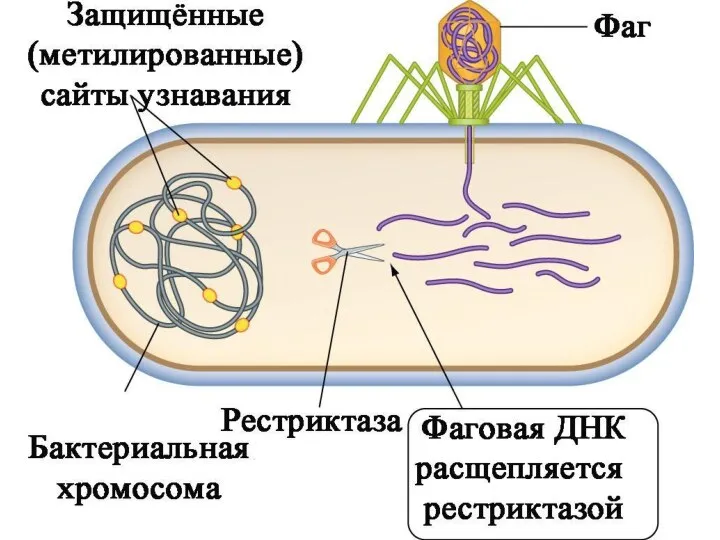

Кстати:
Даже порезанная ДНК из фага может участвовать в рекомбинации.
Кстати:
Даже порезанная ДНК из фага может участвовать в рекомбинации.

Почему рестриктазы так называются?
Система рестрикции-модификации предотвращает фаговую инфекции не на 100%,
Почему рестриктазы так называются?
Система рестрикции-модификации предотвращает фаговую инфекции не на 100%,

Нетривиальная идея №2!
В научной и учебной литературе чаще всего пишут, что
Нетривиальная идея №2!
В научной и учебной литературе чаще всего пишут, что

Система рестрикции модификации помогает осуществлять преимущественно рекомбинацию только между очень родственными
Система рестрикции модификации помогает осуществлять преимущественно рекомбинацию только между очень родственными

В настоящее время общепризнанным является тот фундаментальный факт, что бактериофаги и
В настоящее время общепризнанным является тот фундаментальный факт, что бактериофаги и
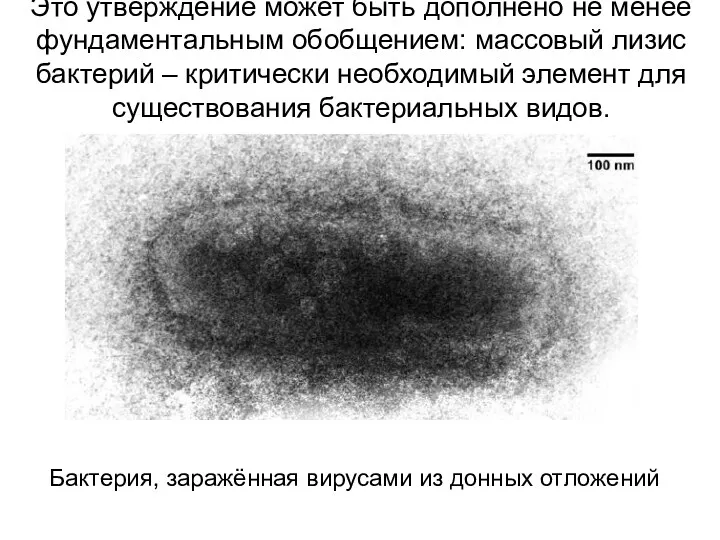
Это утверждение может быть дополнено не менее фундаментальным обобщением: массовый лизис
Это утверждение может быть дополнено не менее фундаментальным обобщением: массовый лизис

Бактерии (в прямоугольнике) и вирусные частицы (овал) из морской воды.
Бактерии (в прямоугольнике) и вирусные частицы (овал) из морской воды.

Таким образом
Фаги оказываются исключительно полезными не только благодаря тому, что они
Таким образом
Фаги оказываются исключительно полезными не только благодаря тому, что они
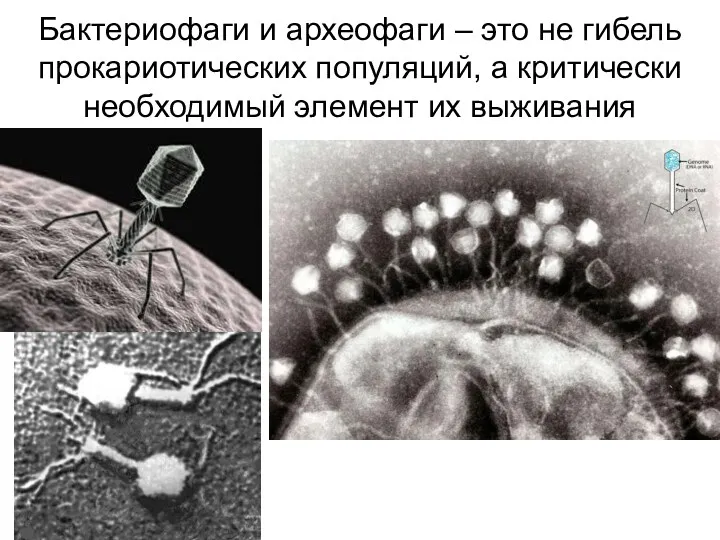
Бактериофаги и археофаги – это не гибель прокариотических популяций, а критически
Бактериофаги и археофаги – это не гибель прокариотических популяций, а критически
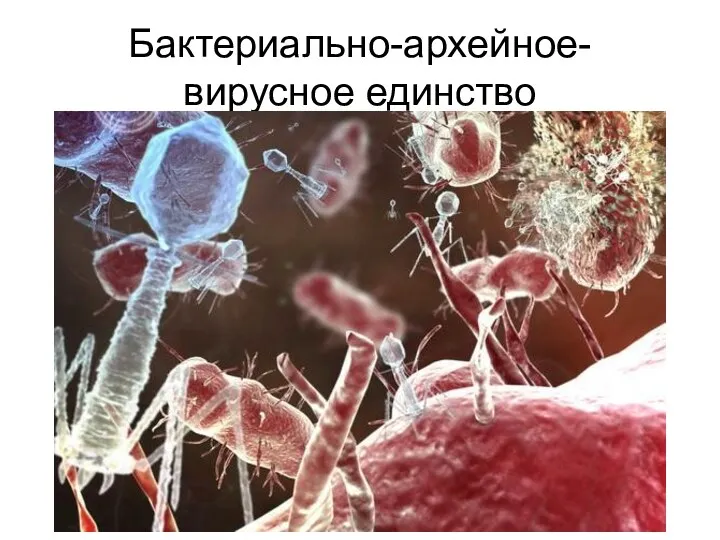
Бактериально-архейное-вирусное единство
Бактериально-архейное-вирусное единство
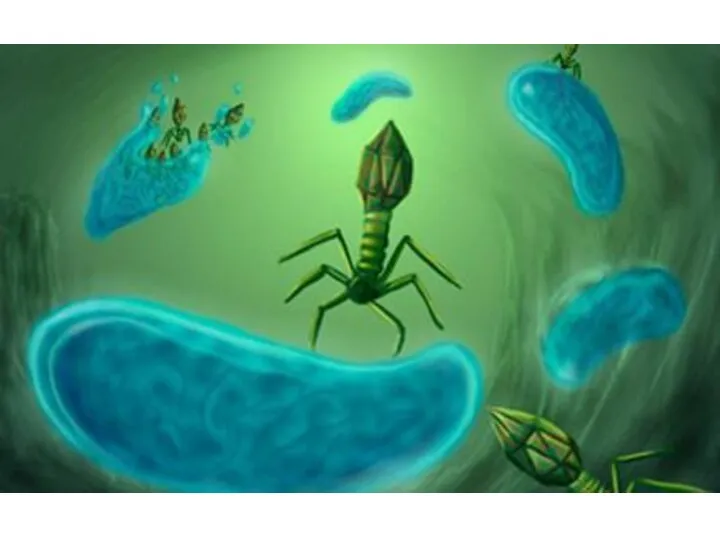

Программируемый лизис клетки – альтернатива фаговому лизису.
Программируемый лизис клетки – альтернатива фаговому лизису.
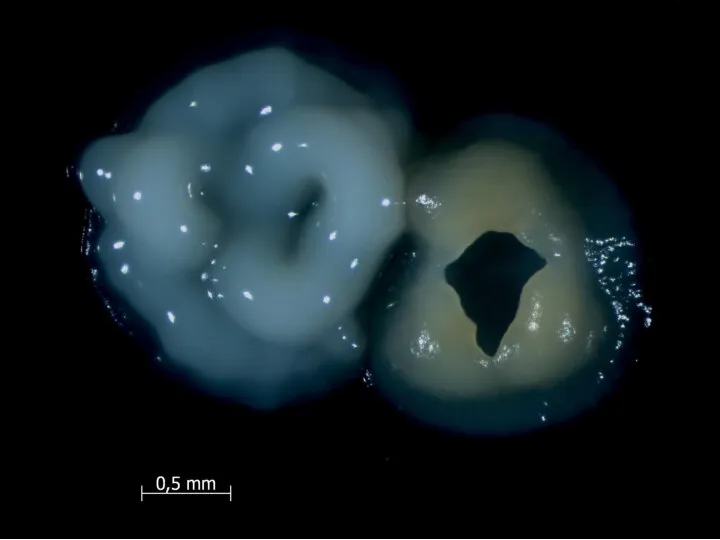
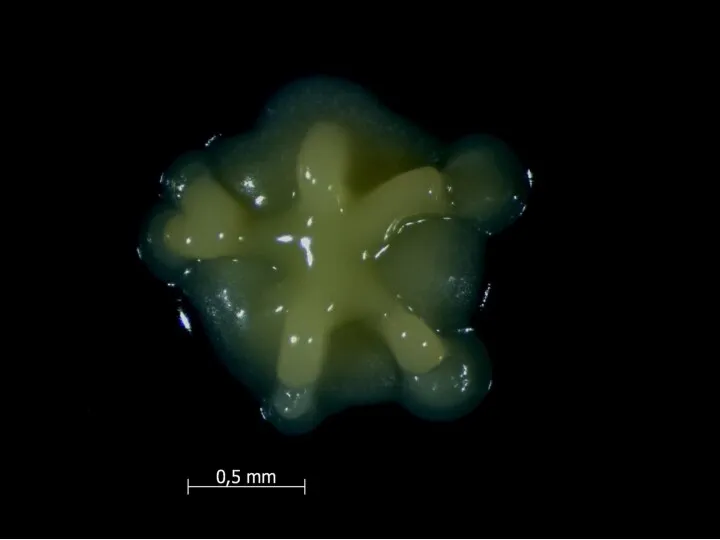


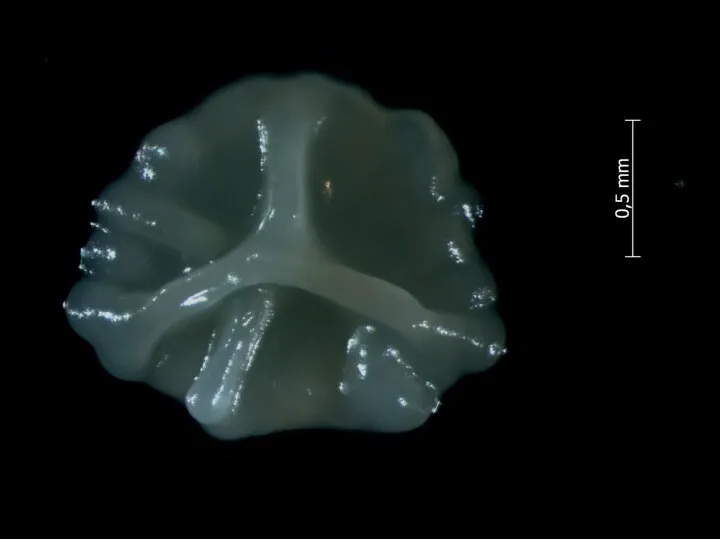

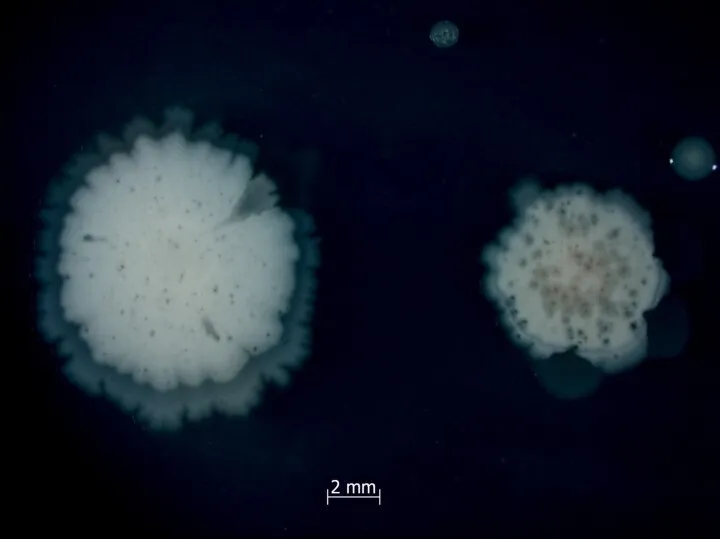
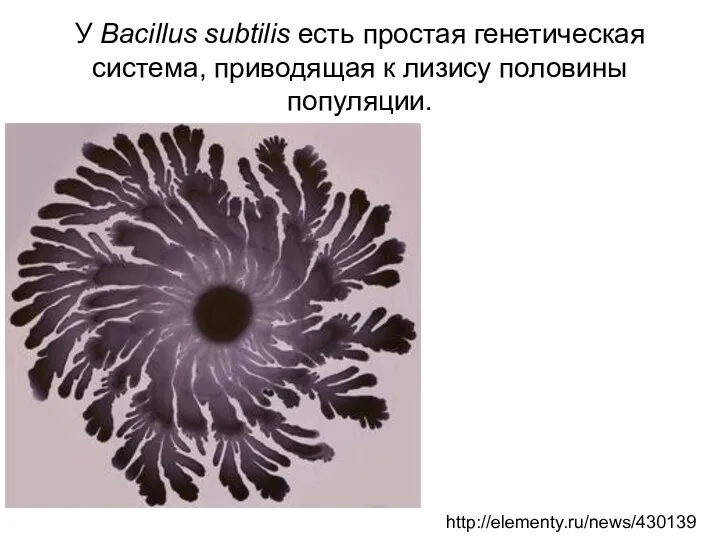
У Bacillus subtilis есть простая генетическая система, приводящая к лизису половины
У Bacillus subtilis есть простая генетическая система, приводящая к лизису половины
Программируемый лизис бактериальных клеток необходим для рекомбинации
Программируемый лизис бактериальных клеток необходим для рекомбинации

P.S. Ещё один недооцененный возможно распространённейший механизм спасения прокариот от Храповика
P.S. Ещё один недооцененный возможно распространённейший механизм спасения прокариот от Храповика

Полиплоидия – возможность частично избавиться от храповика Меллера.
Нередко сестринские хромосомы в
Полиплоидия – возможность частично избавиться от храповика Меллера.
Нередко сестринские хромосомы в

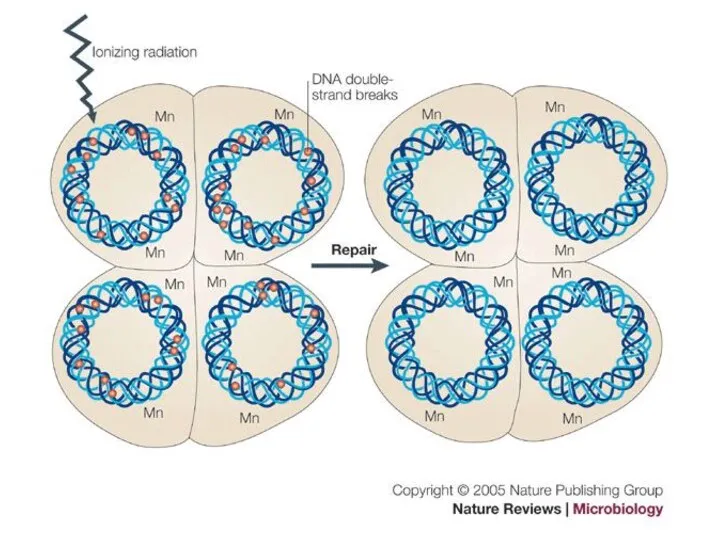
Почему Deinococcus radiodurans способен выдерживать чудовищные дозы радиации?
Дейнококк является тетраплоидом —
Почему Deinococcus radiodurans способен выдерживать чудовищные дозы радиации?
Дейнококк является тетраплоидом —

 Развитие жизни в архее и протерозое
Развитие жизни в архее и протерозое Наземно-воздушная среда обитания организмов
Наземно-воздушная среда обитания организмов Царство Животные
Царство Животные Многоствольные формы
Многоствольные формы Методологическая роль лабораторных и практических работ по биологии в рамках ФГОС.
Методологическая роль лабораторных и практических работ по биологии в рамках ФГОС. Происхождение человека. Теории происхождения человека
Происхождение человека. Теории происхождения человека Опорно-двигательный аппарат. Позвоночник и грудная клетка
Опорно-двигательный аппарат. Позвоночник и грудная клетка Среда обитания организмов. Тухватуллина Азалия
Среда обитания организмов. Тухватуллина Азалия Культивирование красных водорослей
Культивирование красных водорослей Содержание и разведение организмов в ботанических садах
Содержание и разведение организмов в ботанических садах Артерии большого круга кровообращения
Артерии большого круга кровообращения Виявлення пристосувань до способу життя птахів. Практична робота 3
Виявлення пристосувань до способу життя птахів. Практична робота 3 Функциональная анатомия кровеносной системы
Функциональная анатомия кровеносной системы Организм и окружающая средa
Организм и окружающая средa Реакции окисления и восстановления биоорганических соединений. (Лекция 5)
Реакции окисления и восстановления биоорганических соединений. (Лекция 5) Основні сенсорні системи організму людини
Основні сенсорні системи організму людини История органического мира земли
История органического мира земли Понятие о вирусах. (10 класс)
Понятие о вирусах. (10 класс) Неконкурентные взаимодействия между видами
Неконкурентные взаимодействия между видами Жизнь в морях и океанах
Жизнь в морях и океанах Особенности питания и пищеварения у животных
Особенности питания и пищеварения у животных Биоритмы человека
Биоритмы человека Формы естественного отбора
Формы естественного отбора The Digestive System in Humans
The Digestive System in Humans Морфологически дифференцированные клетки прокариот (МДК)
Морфологически дифференцированные клетки прокариот (МДК) Морфология и физиология вирусов
Морфология и физиология вирусов Клетка, как целостная живая система
Клетка, как целостная живая система Риби. Особливості будови та спосіб життя
Риби. Особливості будови та спосіб життя