- Клетка, как целостная живая система

Содержание
- 2. Клеточные мембрана - динамические и эластические молекулярные структуры, состоящие из белков и липидов. Также в них
- 4. Химический состав клеточной мембраны Классификация липидов
- 5. Собственно мембрана (плазмалемма) Липидный слой плазмалеммы состоит из четырех классов соединений: 1) фосфолипидов, 2) нейтральных липидов
- 6. В каждой эукариотической клетке присутствует более 1000 разновидностей липидов — что обеспечивается возможностью комбинации различных полярных
- 7. Каждая мембрана клетки замкнута, т.е. имеет внутреннюю и внешнюю поверхности, различающиеся по липидному и белковому составам
- 9. Латеральная гетерогенность обеспечивает существование липидов в мембране в двух фазах: Твердая фаза (гель) — So (от
- 10. Жидкие фазы: Упорядоченная жидкая фаза — Lo (от “liquid ordered”). Она отличается достаточно высокой упорядоченностью липидных
- 11. Твердая фаза (S) образует мембранные рафты — это маленькие (10–200 нм), гетерогенные участки плотно-упакованного липида являются
- 12. Рафты участвуют в: «почковании» вирусных частиц. Многие вирусы, покидая зараженную ранее клетку, облачаются в липидную оболочку
- 14. Ассоциация с рафтами может быть решающим фактором, определяющим активность интегральных мембранных белков, в том числе ионных
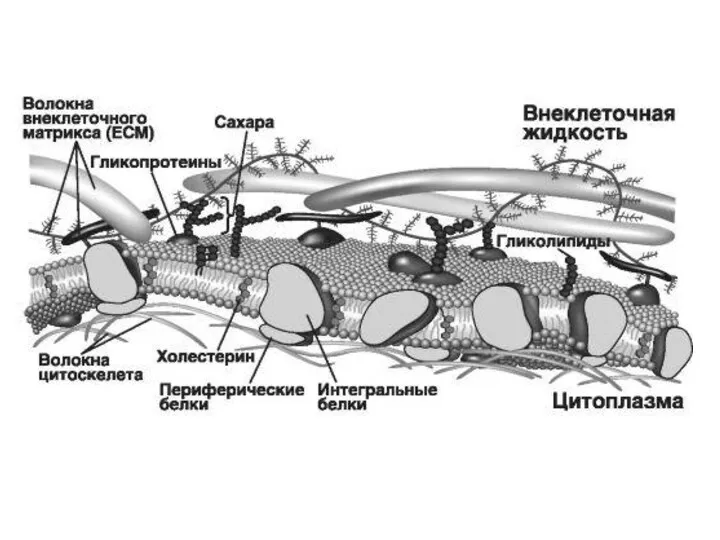
- 15. Гликокаликс Гликокаликс содержит полисахариды, рецепторные белки, ферменты. Гликокаликс представляет собой связанный с двойным слоем липидов гликопротеиновый
- 16. Благодаря способности образовывать связи со многими белками интегрины участвуют в процессах распознавания, адгезии, миграции клеток на
- 17. Гликокаликс
- 18. Гликокаликс
- 19. Подмембранный слой Представлен элементами цитоскелета, образующими сеть на внутренней поверхности плазмалеммы и прикрепленными к ней при
- 21. Подмембранный слой
- 22. Разновидности клеточных мембран Мембрана эритроцитов состоит из белков (50%), липидов (40%) и углеводов (10%). Липиды включают
- 23. Разновидности клеточных мембран Миелиновые мембраны, окружающие аксоны нейронов, многослойны, в них присутствует большое количество липидов (около
- 24. Белки составляют до 20% сухой массы миелина. Они бывают двух видов: белки, расположенные на поверхности, и
- 25. Разновидности клеточных мембран Мембраны митохондрий. Митохондрии имеют две бислойных мембраны. Гладкая внешняя мембрана полностью окружает всю
- 26. Разновидности клеточных мембран Внутренняя (цитоплазматическая) мембрана бактерий по структуре сходна с внутренними мембранами хлоропластов и митохондрий.
- 27. Структурные компоненты клетки Постоянные Непостоянные Выполняют специфические жизненно-важные функции Органоиды Могут появляться или исчезать в процессе
- 28. Классификация органелл
- 29. Клеточное ядро основной компонент эукариотической клетки, содержащий её генетический материал. Ядерная пора Тубулиновые микрофиламенты Участок эндоплазматической
- 30. Шероховатая эндоплазматическая сеть Ядро Ядерная оболочка Ядерная пора Рибосомы Гладкая эндоплазматическая сеть Аппарат Гольджи Гладкая эндоплазматическая
- 31. Ядро эукариотической клетки Функция ядра заключается: в хранении и реализации генетической информации (для синтеза белков) и
- 32. Кариолемма Кариолемма состоит из двух биологических мембран (наружной и внутренней) толщиной 6 - 7,5 нм, которые
- 33. Кариолемма Перинуклеарное пространство (люмен) переходит в полости ЭПС. Со стороны цитоплазмы наружная ядерная мембрана окружена сетью
- 34. Внутренняя мембрана кариолеммы ядерная ламина (80-300 нм) – структура, образованная белками-ламинами, к которой прикреплены нити хроматина.
- 35. Функции ламины Поддерживает форму ядра. Участвует в формировании порового комплекса. Отвечает за упорядоченное расположение хроматина в
- 36. Кариолемма В отличие от других органелл, кариолемма содержит поровые комплексы, состоящие из пор и белковых компонентов.
- 37. Области перехода внутренней мембраны кариолеммы в наружную. В поре расположен комплекс ядерной поры: Два параллельных белковых
- 38. Комплекс ядерной поры (поровый комплекс) Электронная микрофотография ядерных пор 80 нм наружная ядерная мембрана внутренняя ядерная
- 39. Комплекс ядерной поры
- 40. Цитоскелет Это сложная динамичная (лабильная), трехмерная цитоплазматическая сеть немембранных волокнистых и трубчатых структур различного типа. Элементы
- 41. Цитоскелет
- 42. Компоненты цитоскелета 1 Микротрубочки 2 Промежуточные филаменты 3 Микрофиламенты 1 2 3
- 44. Микротрубочки Микротрубочки – самые крупные элементы цитоскелета. Построены из димеров глобулярного белка тубулина (13 глобул по
- 45. Лабильность микротрубочек «–» конец связан с белками ЦОМТ. «+» конец свободный, участвует в процессах полимеризации /
- 46. Микротрубочки Работа микротрубочек регулируется специальными белками (динеин, кинезин, МАР-белки, и др.). МАР-белки (Microtubule-associated protein, MAP) регулируют
- 47. Динеин и кинезин обеспечивает скольжение микротрубочек относительно друг друга и перемещение мембранных органелл вдоль по поверхности
- 48. Центр организации микротрубочек ЦОМТ, связанный с клеточным центром. Микротрубочки могут образовываться в цитозоле и вне связи
- 49. Комплексы микротрубочек В клетке микротрубочки могут формировать комплексы, выполняющие специфические функции. К комплексам, построенным из микротрубочек
- 50. Ресничка На вертикальном разрезе ресничка состоит из корешка, базального тельца (в цитоплазме) и собственно реснички (над
- 51. Ресничка Строение ресничек и жгутиков одинаково – это цилиндрические выросты цитоплазмы, основу которых составляет каркас из
- 52. Неподвижные Все типы клеток (кроме клеток крови, мышечных клеток) в G0-периоде формируют первичные реснички. Рост аксонемы
- 53. Клеточный центр (диплосома) в 1888 г. Теодор Бовери Универсальный немембранный органоид всех эукариотических клеток, органелла клеточного
- 54. Клеточный центр образован двумя перпендикулярно расположенными центриолями и центросферой. Центриоли имеют цилиндрическую форму. Их поперечный диаметр
- 55. Клеточный центр в интерфазной клетке организует ее компартменты, участвует в циклозе (двидении цитоплазмы). При митотическом и
- 56. Клеточный центр Дочерняя центриоль. Триплеты микротрубочек Материнская центриоль Центросфера Молекулы динеина.
- 57. Центросфера (сома) Сателлиты/гало Микротрубочки Фокусы схождения микротрубочек Сателлиты – ЦОМТ (Центры Организации МикроТрубочек), состоящие из белковой
- 58. Центросомный цикл М-фаза: две диплосомы на полюсах клетки, от них отходят нити ахроматинового веретена деления. Материнская
- 59. Базальное тельце (кинетосома) Состоит из 9 триплетов микротрубочек, имеет центральную белковую фибриллу и спицы идущие к
- 60. Функции микротрубочек Поддержание формы клетки Обеспечение внутриклеточного транспорта Формирование органелл Обеспечение подвижности клетки Формирования веретена деления
- 61. Промежуточные филаменты Прочные устойчивые стабильные и самые долгоживущие компоненты цитоскелета. Представляют собой неполярные нити (фибриллы) толщиной
- 62. Строение промежуточных филаментов Димер Тетрамер Протофиламент Волокно 1 2 3 4
- 63. Классы промежуточных филаментов Тонофиламенты (кератины); Десминовые филаменты (десмин); Виментиновые филаменты (виментин); Нейрофиламенты (NF-L, NF-M, NF-H); Глиальные
- 64. Функции промежуточных филаментов Поддержание формы клетки; Распределение органелл в цитоплазме; Формирование рогового вещества; Формирование остова отростков
- 65. Микрофиламенты Тонкие белковые полярные нити диаметром 5-7 нм, расположенные в цитоплазме поодиночке, пучками или в виде
- 66. Белки микрофиламентов Актин – глобулярный мономерный белок (G-актин), способный к полимеризации (F-актин). Молекула актина имеет вид
- 67. Микрофиламенты
- 68. Актин-связывающие белки ингибирующие полимеризацию актина (профиллин, ДНКаза I); стабилизирующие белки (тропомиозины); кэпактины (гельзолин/вилин, фрагмин/северин) сшивающие актиновые
- 69. Микроворсинки Выросты цитоплазмы, окруженные плазмолеммой, каркас которых образован пучком микрофиламентов. У основания микроворсинки пучок микрофиламентов переходит
- 70. Функции микрофиламентов Обеспечение сокращения клеток Обеспечение движения клеток. Обеспечение мембранных функций. Перемещение в цитозоле органелл, макромолекул.
- 71. Микротрабекулы Наименее изученная часть цитоскелета. Выявляются только высоковольтной (мегавольтной) трансмиссионной электронной микроскопией в виде сети нитей
- 72. Микротрабекулы Система тонких белковых нитей, пересекающих цитоплазму в различных направлениях. С микротрабекулярной системой связаны микротрубочки и
- 73. ВЕЗИКУЛЯРНАЯ СИСТЕМА КЛЕТКИ 1. Гранулярная эндоплазматическая сеть. 2. Агранулярная эндоплазматическая сеть. 3. Аппарат Гольджи. 4. Лизосомы
- 74. Эндоплазматическая сеть Эндоплазматическая сеть представлена трубочками, цистернами, мембранами, пузырьками. Большинство веществ синтезируется на наружной поверхности мембран.
- 75. Эндоплазматическая сеть Кариолемма Шероховатая ЭПС Цистерны
- 76. Является местом конденсации и накопления продуктов секреции, вырабатываемых в других участках клетки, в основном, в эндоплазматической
- 78. ЛИЗОСОМЫ Ферментный состав внутреннего содержимого 0,2-0,5 мкм насос АТФ АДФ Кислые гидролазы: нуклеазы протеазы гликозидазы липазы
- 79. Лизосомальный цикл
- 80. Пероксисомы – основной центр утилизации кислорода в клетках, с последующим образованием перекиси водорода (H2O2). Образуются из
- 82. Скачать презентацию

Клеточные мембрана - динамические и эластические молекулярные структуры, состоящие из белков и
Клеточные мембрана - динамические и эластические молекулярные структуры, состоящие из белков и

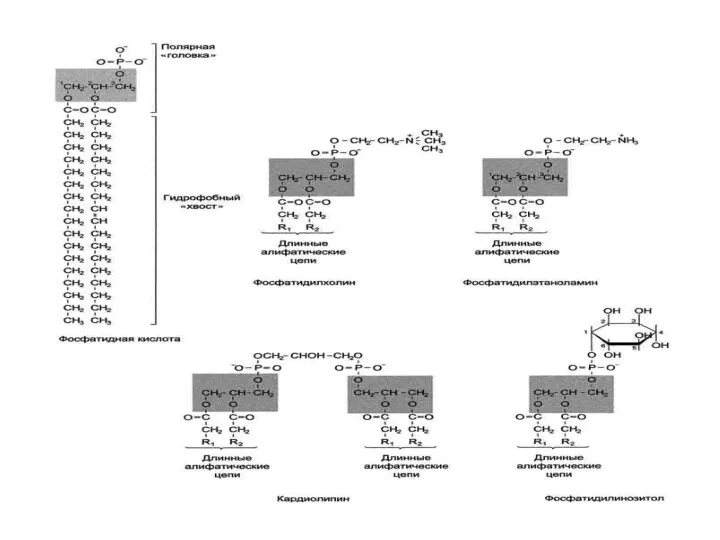
Химический состав клеточной мембраны
Классификация липидов
Химический состав клеточной мембраны
Классификация липидов
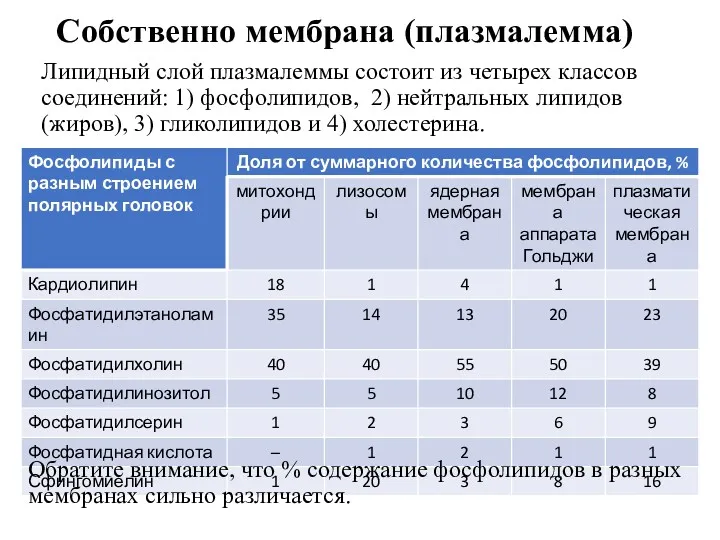
Собственно мембрана (плазмалемма)
Липидный слой плазмалеммы состоит из четырех классов соединений: 1)
Собственно мембрана (плазмалемма)
Липидный слой плазмалеммы состоит из четырех классов соединений: 1)

В каждой эукариотической клетке присутствует более 1000 разновидностей липидов — что обеспечивается возможностью
В каждой эукариотической клетке присутствует более 1000 разновидностей липидов — что обеспечивается возможностью

Каждая мембрана клетки замкнута, т.е. имеет внутреннюю и внешнюю поверхности, различающиеся
Каждая мембрана клетки замкнута, т.е. имеет внутреннюю и внешнюю поверхности, различающиеся

Латеральная гетерогенность обеспечивает существование липидов в мембране в двух фазах:
Твердая фаза
Латеральная гетерогенность обеспечивает существование липидов в мембране в двух фазах:
Твердая фаза

Жидкие фазы:
Упорядоченная жидкая фаза — Lo (от “liquid ordered”). Она отличается достаточно высокой упорядоченностью
Жидкие фазы: Упорядоченная жидкая фаза — Lo (от “liquid ordered”). Она отличается достаточно высокой упорядоченностью

Твердая фаза (S) образует мембранные рафты — это маленькие (10–200 нм), гетерогенные участки
Твердая фаза (S) образует мембранные рафты — это маленькие (10–200 нм), гетерогенные участки

Рафты участвуют в:
«почковании» вирусных частиц. Многие вирусы, покидая зараженную ранее клетку,
Рафты участвуют в:
«почковании» вирусных частиц. Многие вирусы, покидая зараженную ранее клетку,
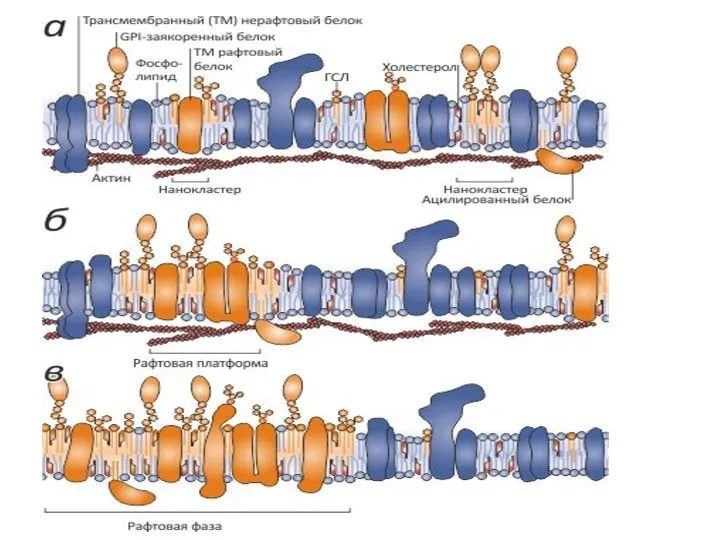

Ассоциация с рафтами может быть решающим фактором, определяющим активность интегральных мембранных
Ассоциация с рафтами может быть решающим фактором, определяющим активность интегральных мембранных

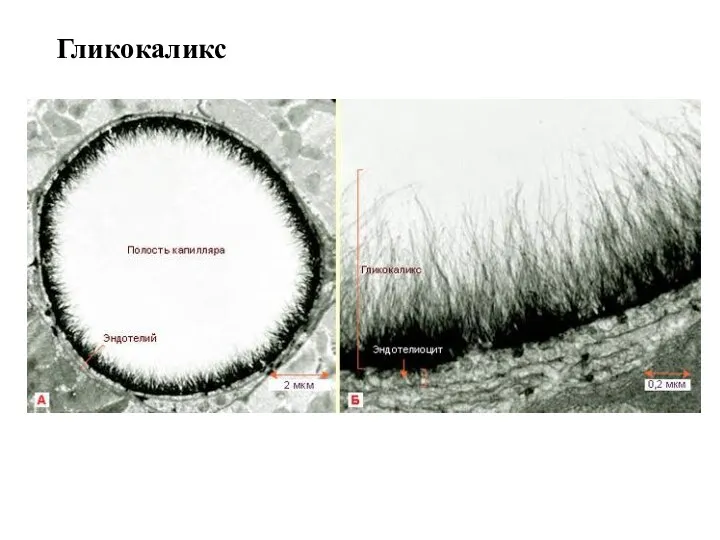
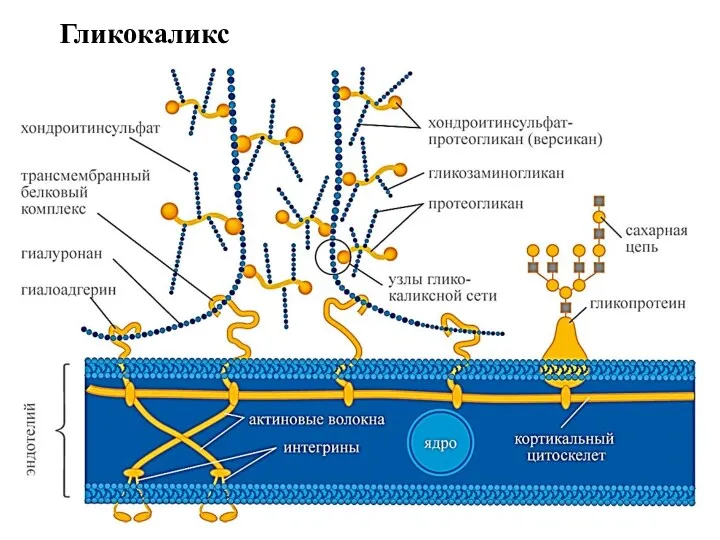
Гликокаликс
Гликокаликс содержит полисахариды, рецепторные белки, ферменты. Гликокаликс представляет собой связанный с
Гликокаликс
Гликокаликс содержит полисахариды, рецепторные белки, ферменты. Гликокаликс представляет собой связанный с

Благодаря способности образовывать связи со многими белками интегрины участвуют в процессах
Благодаря способности образовывать связи со многими белками интегрины участвуют в процессах
Гликокаликс
Гликокаликс
Гликокаликс
Гликокаликс

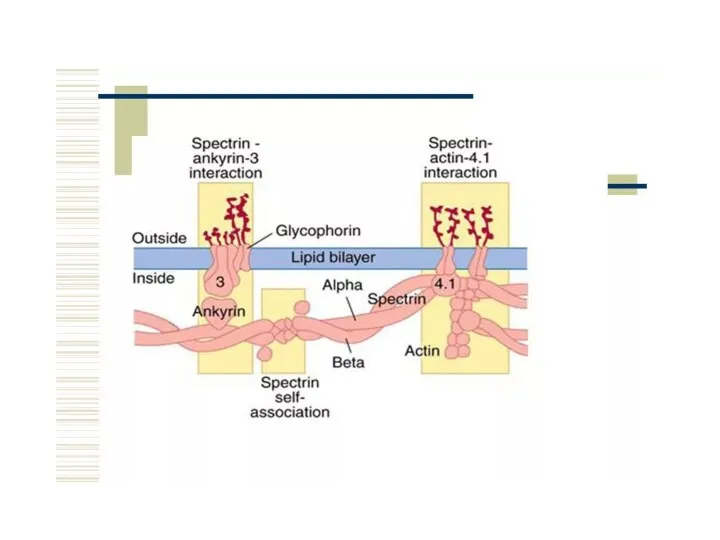
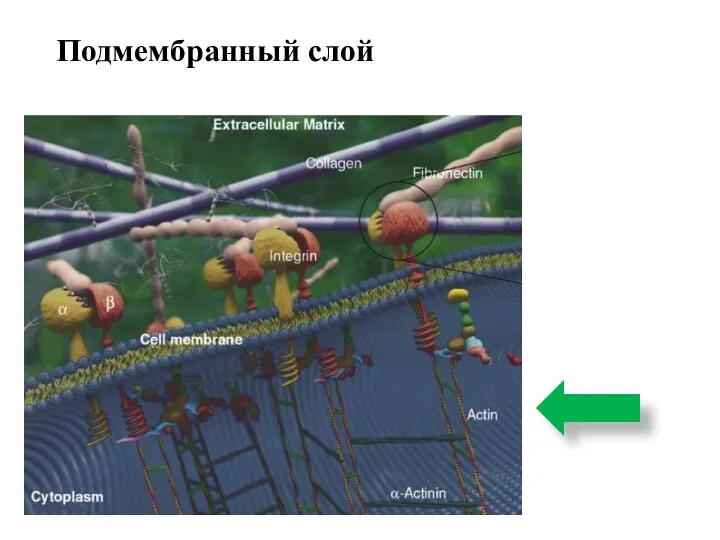
Подмембранный слой
Представлен элементами цитоскелета, образующими сеть на внутренней поверхности плазмалеммы
Подмембранный слой
Представлен элементами цитоскелета, образующими сеть на внутренней поверхности плазмалеммы
Подмембранный слой
Подмембранный слой

Разновидности клеточных мембран
Мембрана эритроцитов состоит из белков (50%), липидов (40%) и
Разновидности клеточных мембран
Мембрана эритроцитов состоит из белков (50%), липидов (40%) и

Разновидности клеточных мембран
Миелиновые мембраны, окружающие аксоны нейронов, многослойны, в них присутствует
Разновидности клеточных мембран
Миелиновые мембраны, окружающие аксоны нейронов, многослойны, в них присутствует

Белки составляют до 20% сухой массы миелина. Они бывают двух видов: белки,
Белки составляют до 20% сухой массы миелина. Они бывают двух видов: белки,

Разновидности клеточных мембран
Мембраны митохондрий. Митохондрии имеют две бислойных мембраны. Гладкая внешняя мембрана
Разновидности клеточных мембран
Мембраны митохондрий. Митохондрии имеют две бислойных мембраны. Гладкая внешняя мембрана

Разновидности клеточных мембран
Внутренняя (цитоплазматическая) мембрана бактерий по структуре сходна с внутренними мембранами
Разновидности клеточных мембран
Внутренняя (цитоплазматическая) мембрана бактерий по структуре сходна с внутренними мембранами
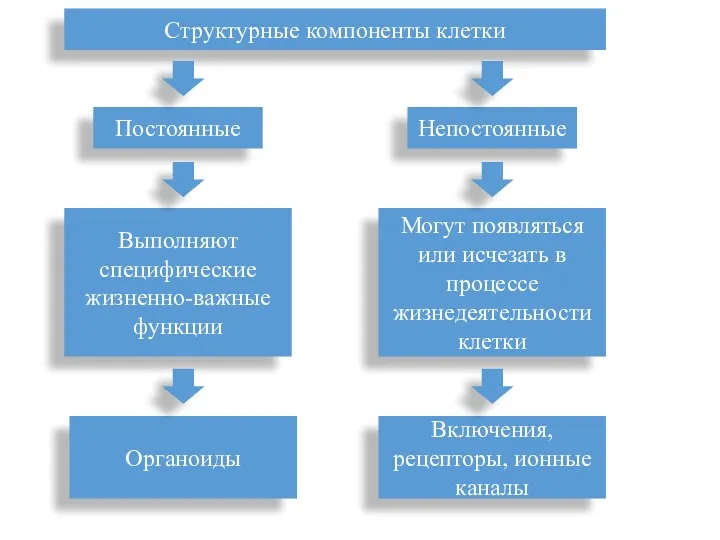
Структурные компоненты клетки
Постоянные
Непостоянные
Выполняют специфические жизненно-важные функции
Органоиды
Могут появляться или исчезать в процессе
Структурные компоненты клетки
Постоянные
Непостоянные
Выполняют специфические жизненно-важные функции
Органоиды
Могут появляться или исчезать в процессе

Классификация органелл
Классификация органелл
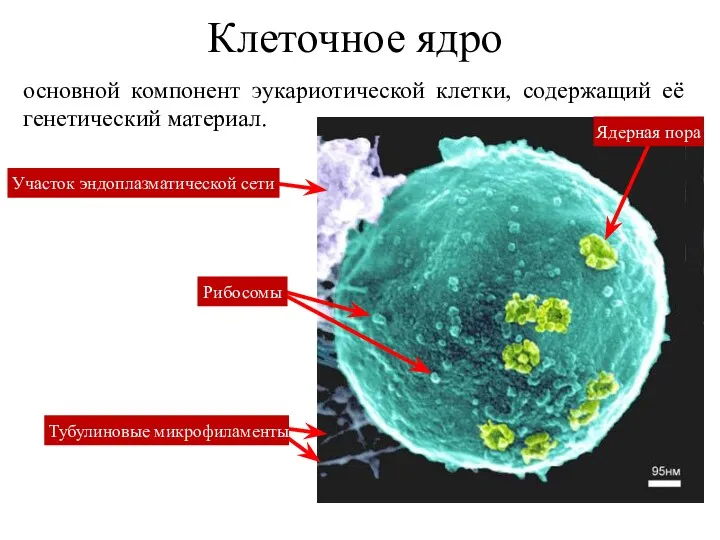
Клеточное ядро
основной компонент эукариотической клетки, содержащий её генетический материал.
Ядерная пора
Тубулиновые микрофиламенты
Участок
Клеточное ядро
основной компонент эукариотической клетки, содержащий её генетический материал.
Ядерная пора
Тубулиновые микрофиламенты
Участок
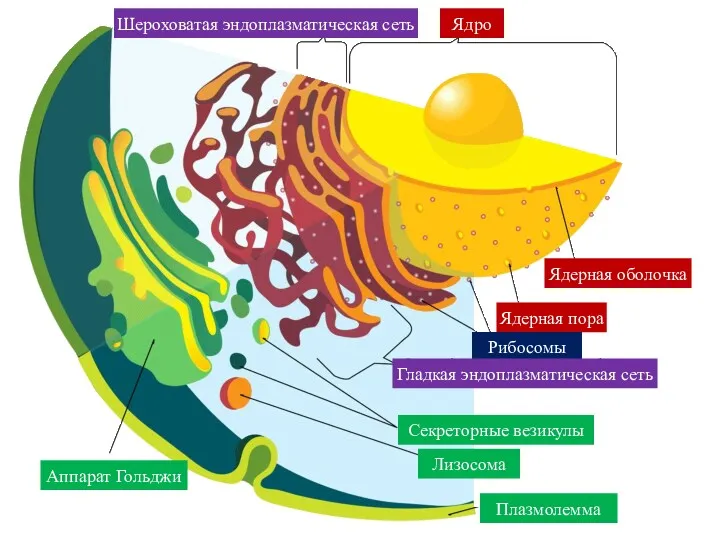
Шероховатая эндоплазматическая сеть
Ядро
Ядерная оболочка
Ядерная пора
Рибосомы
Гладкая эндоплазматическая сеть
Аппарат Гольджи
Гладкая эндоплазматическая сеть
Секреторные везикулы
Лизосома
Плазмолемма
Шероховатая эндоплазматическая сеть
Ядро
Ядерная оболочка
Ядерная пора
Рибосомы
Гладкая эндоплазматическая сеть
Аппарат Гольджи
Гладкая эндоплазматическая сеть
Секреторные везикулы
Лизосома
Плазмолемма

Ядро эукариотической клетки
Функция ядра заключается: в хранении и реализации генетической информации
Ядро эукариотической клетки
Функция ядра заключается: в хранении и реализации генетической информации
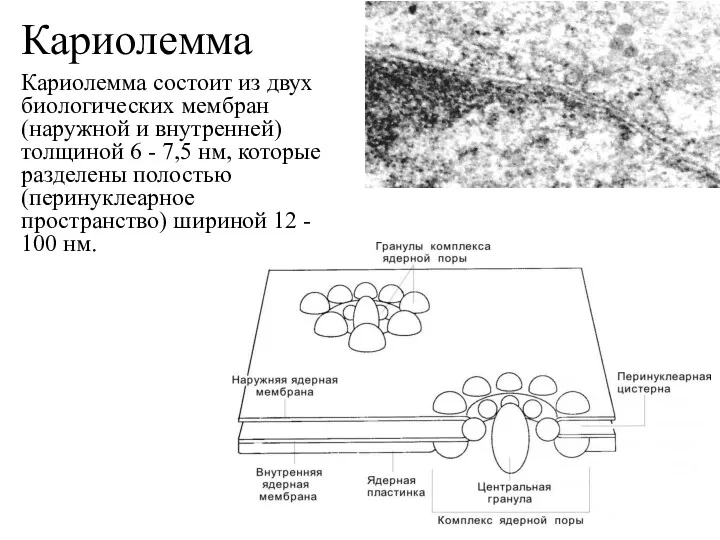
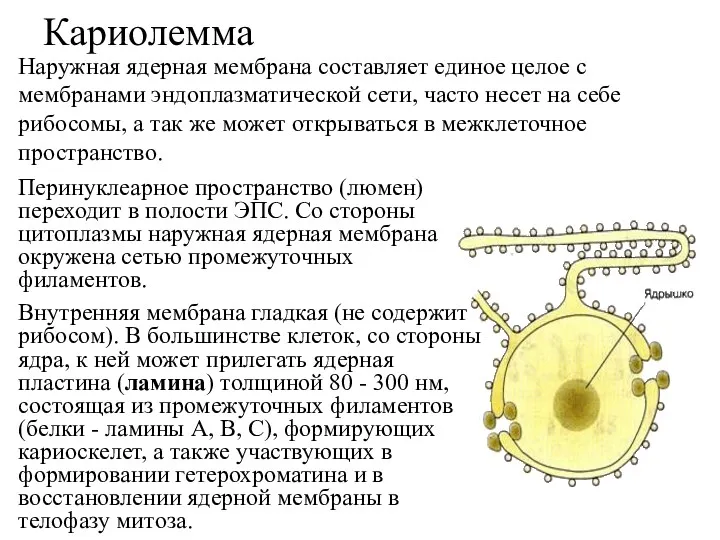
Кариолемма
Кариолемма состоит из двух биологических мембран (наружной и внутренней) толщиной 6
Кариолемма
Кариолемма состоит из двух биологических мембран (наружной и внутренней) толщиной 6
Кариолемма
Перинуклеарное пространство (люмен) переходит в полости ЭПС. Со стороны цитоплазмы наружная
Кариолемма
Перинуклеарное пространство (люмен) переходит в полости ЭПС. Со стороны цитоплазмы наружная
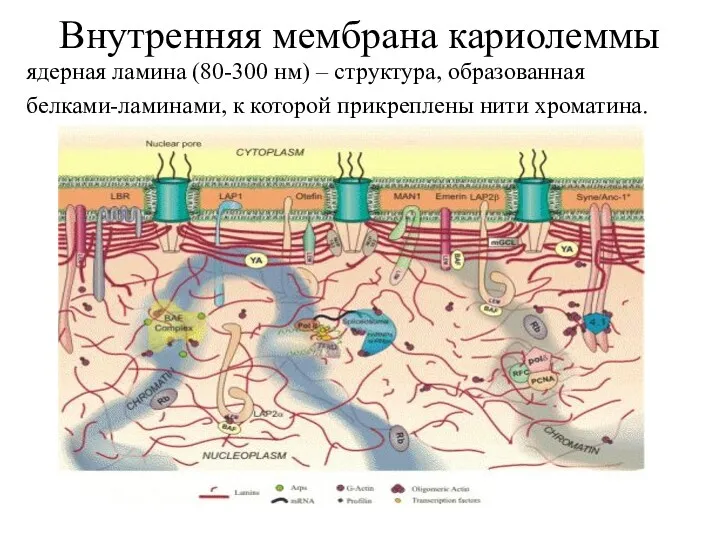
Внутренняя мембрана кариолеммы
ядерная ламина (80-300 нм) – структура, образованная
белками-ламинами, к
Внутренняя мембрана кариолеммы
ядерная ламина (80-300 нм) – структура, образованная
белками-ламинами, к

Функции ламины
Поддерживает форму ядра.
Участвует в формировании порового комплекса.
Отвечает за упорядоченное расположение
Функции ламины
Поддерживает форму ядра.
Участвует в формировании порового комплекса.
Отвечает за упорядоченное расположение

Кариолемма
В отличие от других органелл, кариолемма содержит поровые комплексы, состоящие из
Кариолемма
В отличие от других органелл, кариолемма содержит поровые комплексы, состоящие из
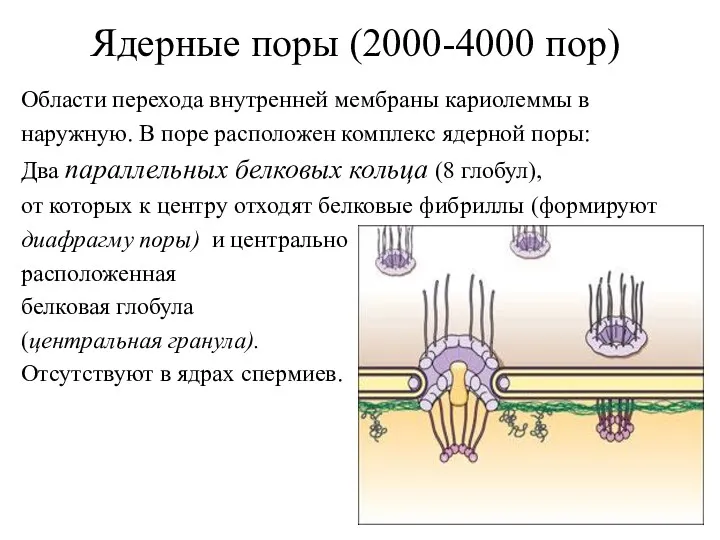
Области перехода внутренней мембраны кариолеммы в
наружную. В поре расположен комплекс ядерной
Области перехода внутренней мембраны кариолеммы в
наружную. В поре расположен комплекс ядерной
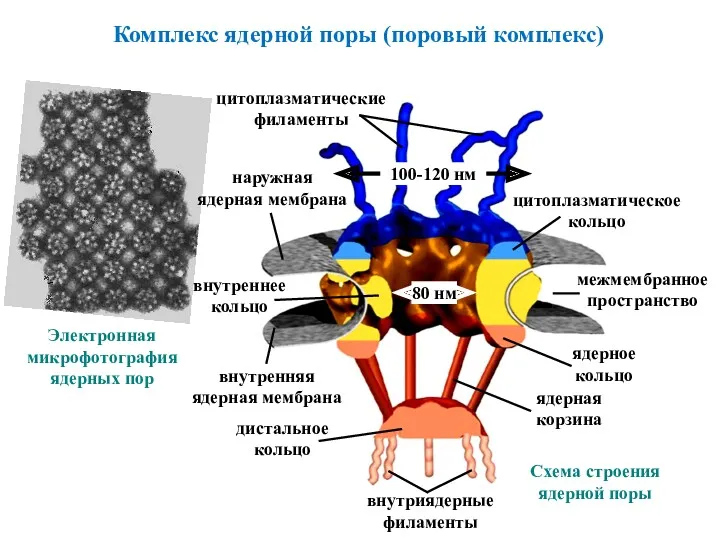
Комплекс ядерной поры (поровый комплекс)
Электронная
микрофотография
ядерных пор
80 нм
наружная
ядерная мембрана
внутренняя
ядерная мембрана
цитоплазматические
филаменты
внутриядерные
филаменты
дистальное
кольцо
ядерная
корзина
ядерное
кольцо
цитоплазматическое
кольцо
межмембранное
пространство
100-120 нм
Схема строения
ядерной
Комплекс ядерной поры (поровый комплекс)
Электронная
микрофотография
ядерных пор
80 нм
наружная
ядерная мембрана
внутренняя
ядерная мембрана
цитоплазматические
филаменты
внутриядерные
филаменты
дистальное
кольцо
ядерная
корзина
ядерное
кольцо
цитоплазматическое
кольцо
межмембранное
пространство
100-120 нм
Схема строения
ядерной
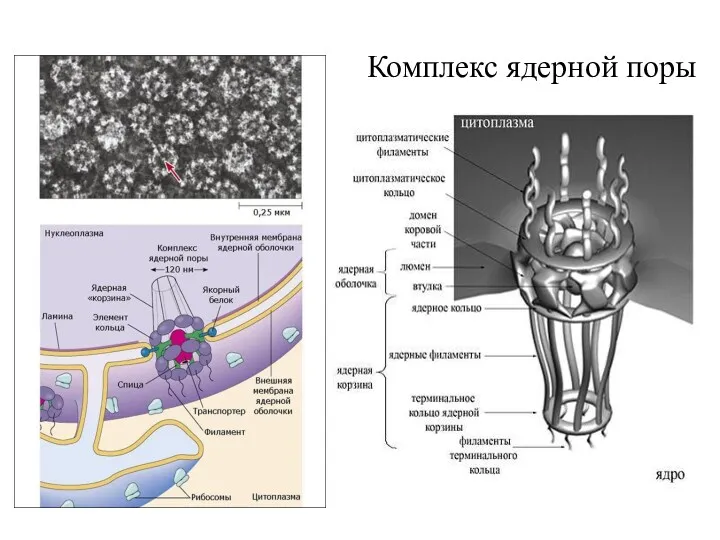
Комплекс ядерной поры
Комплекс ядерной поры

Цитоскелет
Это сложная динамичная (лабильная), трехмерная цитоплазматическая сеть немембранных волокнистых и
Цитоскелет
Это сложная динамичная (лабильная), трехмерная цитоплазматическая сеть немембранных волокнистых и
Цитоскелет
Цитоскелет
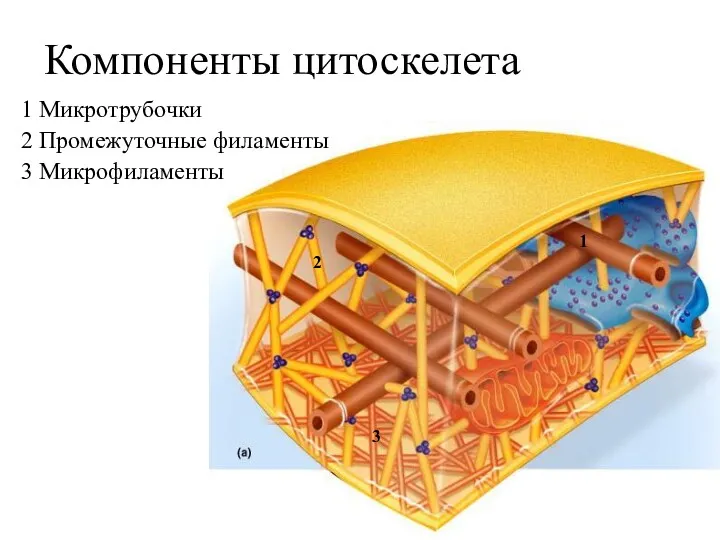
Компоненты цитоскелета
1 Микротрубочки
2 Промежуточные филаменты
3 Микрофиламенты
1
2
3
Компоненты цитоскелета
1 Микротрубочки
2 Промежуточные филаменты
3 Микрофиламенты
1
2
3
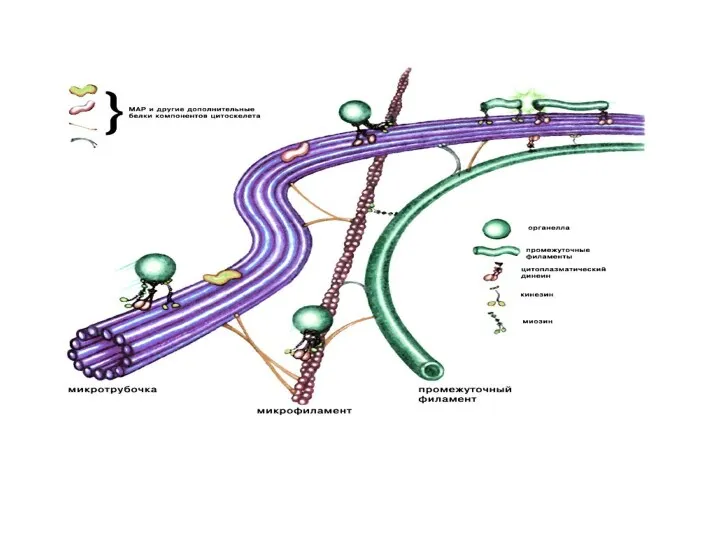
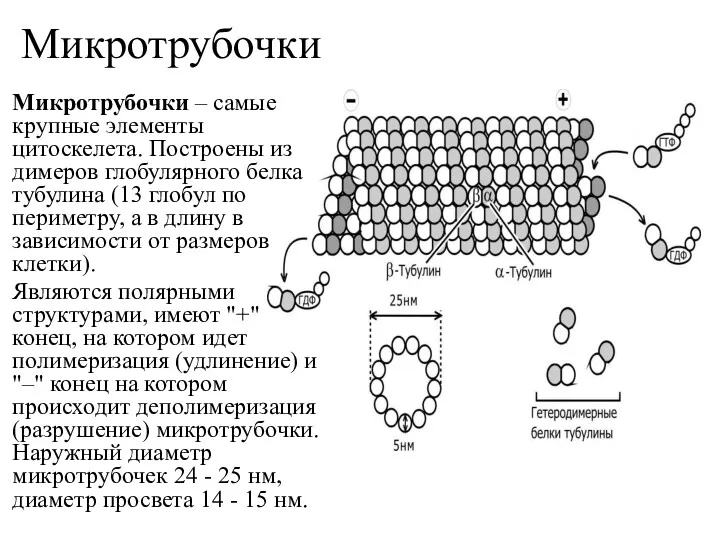
Микротрубочки
Микротрубочки – самые крупные элементы цитоскелета. Построены из димеров глобулярного белка
Микротрубочки
Микротрубочки – самые крупные элементы цитоскелета. Построены из димеров глобулярного белка

Лабильность микротрубочек
«–» конец связан с белками ЦОМТ.
«+» конец свободный, участвует в
Лабильность микротрубочек
«–» конец связан с белками ЦОМТ.
«+» конец свободный, участвует в

Микротрубочки
Работа микротрубочек регулируется специальными белками (динеин, кинезин, МАР-белки, и др.).
МАР-белки
Микротрубочки
Работа микротрубочек регулируется специальными белками (динеин, кинезин, МАР-белки, и др.).
МАР-белки
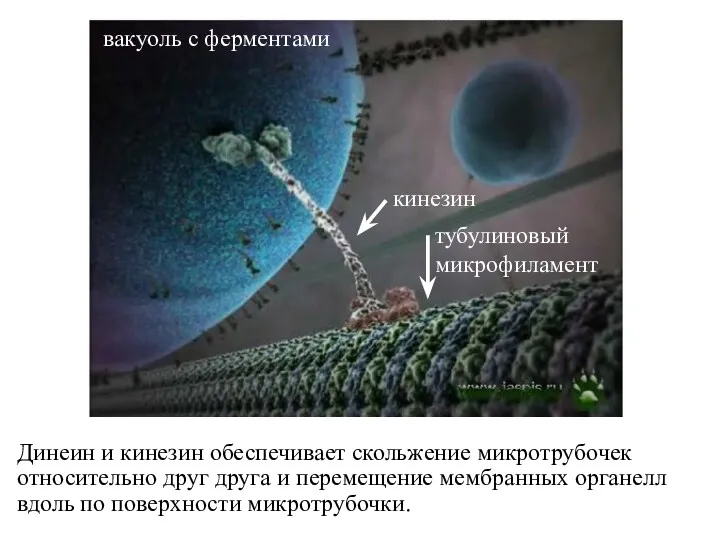
Динеин и кинезин обеспечивает скольжение микротрубочек относительно друг друга и перемещение
Динеин и кинезин обеспечивает скольжение микротрубочек относительно друг друга и перемещение

Центр организации микротрубочек
ЦОМТ, связанный с клеточным центром.
Микротрубочки могут образовываться в цитозоле
Центр организации микротрубочек
ЦОМТ, связанный с клеточным центром.
Микротрубочки могут образовываться в цитозоле

Комплексы микротрубочек
В клетке микротрубочки могут формировать комплексы, выполняющие специфические функции.
Комплексы микротрубочек
В клетке микротрубочки могут формировать комплексы, выполняющие специфические функции.
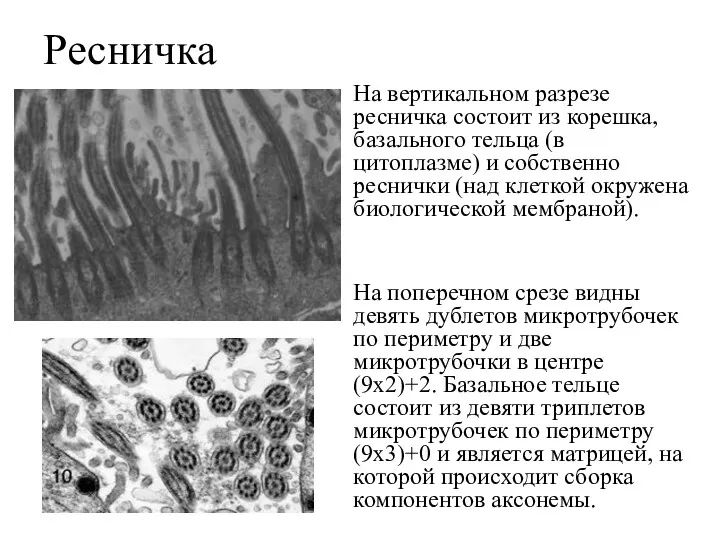
Ресничка
На вертикальном разрезе ресничка состоит из корешка, базального тельца (в цитоплазме)
Ресничка
На вертикальном разрезе ресничка состоит из корешка, базального тельца (в цитоплазме)
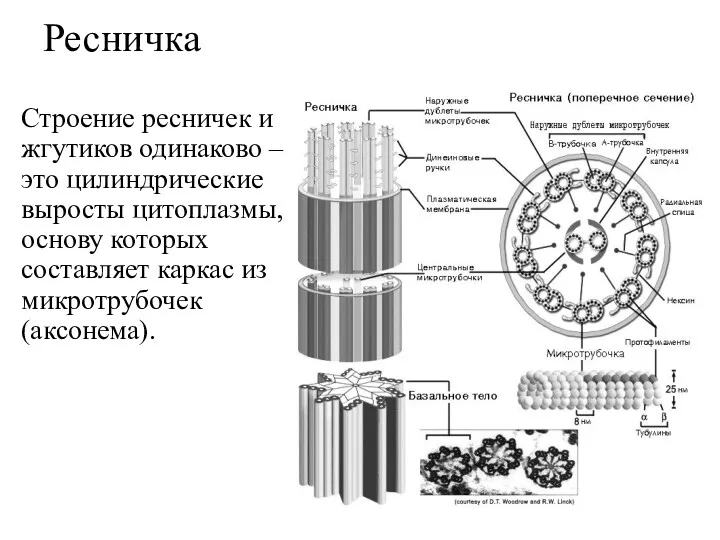
Ресничка
Строение ресничек и жгутиков одинаково – это цилиндрические выросты цитоплазмы, основу
Ресничка
Строение ресничек и жгутиков одинаково – это цилиндрические выросты цитоплазмы, основу
Неподвижные
Все типы клеток (кроме клеток крови, мышечных клеток) в G0-периоде формируют
Неподвижные
Все типы клеток (кроме клеток крови, мышечных клеток) в G0-периоде формируют

Клеточный центр (диплосома)
в 1888 г. Теодор Бовери
Универсальный немембранный органоид
Клеточный центр (диплосома)
в 1888 г. Теодор Бовери
Универсальный немембранный органоид

Клеточный центр образован двумя перпендикулярно расположенными центриолями и центросферой. Центриоли имеют
Клеточный центр образован двумя перпендикулярно расположенными центриолями и центросферой. Центриоли имеют

Клеточный центр в интерфазной клетке организует ее компартменты, участвует в циклозе
Клеточный центр в интерфазной клетке организует ее компартменты, участвует в циклозе
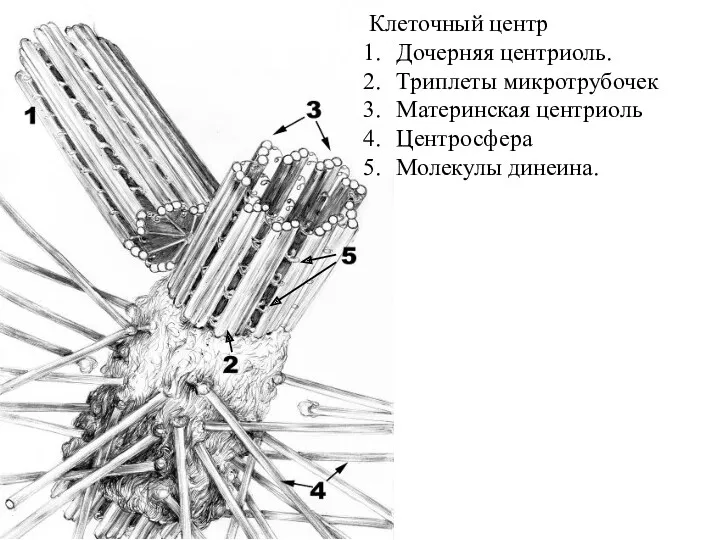
Клеточный центр
Дочерняя центриоль.
Триплеты микротрубочек
Материнская центриоль
Центросфера
Молекулы динеина.
Клеточный центр
Дочерняя центриоль.
Триплеты микротрубочек
Материнская центриоль
Центросфера
Молекулы динеина.
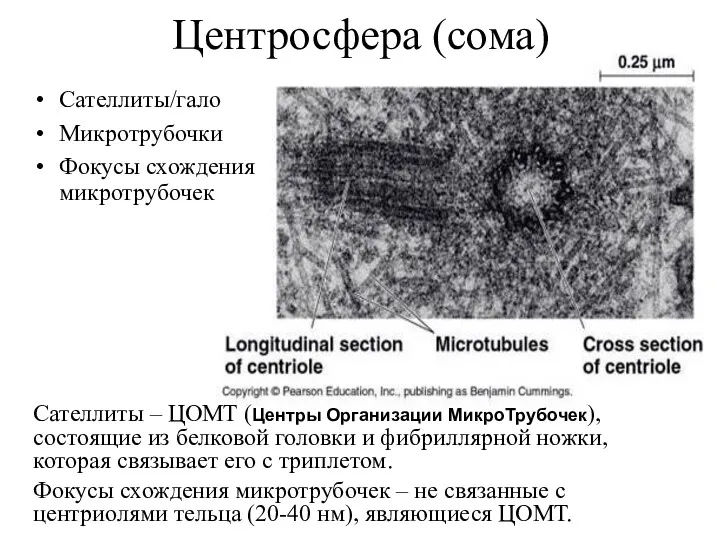
Центросфера (сома)
Сателлиты/гало
Микротрубочки
Фокусы схождения микротрубочек
Сателлиты – ЦОМТ (Центры Организации МикроТрубочек), состоящие из белковой головки
Центросфера (сома)
Сателлиты/гало
Микротрубочки
Фокусы схождения микротрубочек
Сателлиты – ЦОМТ (Центры Организации МикроТрубочек), состоящие из белковой головки

Центросомный цикл
М-фаза: две диплосомы на полюсах клетки, от них отходят нити
Центросомный цикл
М-фаза: две диплосомы на полюсах клетки, от них отходят нити

Базальное тельце (кинетосома)
Состоит из 9 триплетов микротрубочек, имеет центральную
белковую фибриллу
Базальное тельце (кинетосома)
Состоит из 9 триплетов микротрубочек, имеет центральную
белковую фибриллу

Функции микротрубочек
Поддержание формы клетки
Обеспечение внутриклеточного транспорта
Формирование органелл
Обеспечение подвижности клетки
Формирования веретена
Функции микротрубочек
Поддержание формы клетки
Обеспечение внутриклеточного транспорта
Формирование органелл
Обеспечение подвижности клетки
Формирования веретена

Промежуточные филаменты
Прочные устойчивые стабильные и самые долгоживущие компоненты цитоскелета.
Представляют собой
Промежуточные филаменты
Прочные устойчивые стабильные и самые долгоживущие компоненты цитоскелета.
Представляют собой
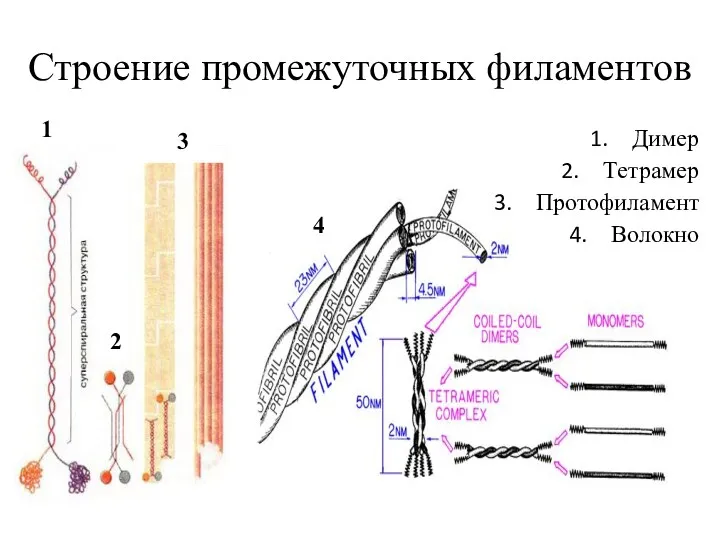
Строение промежуточных филаментов
Димер
Тетрамер
Протофиламент
Волокно
1
2
3
4
Строение промежуточных филаментов
Димер
Тетрамер
Протофиламент
Волокно
1
2
3
4

Классы промежуточных филаментов
Тонофиламенты (кератины);
Десминовые филаменты (десмин);
Виментиновые филаменты (виментин);
Нейрофиламенты (NF-L, NF-M, NF-H);
Глиальные
Классы промежуточных филаментов
Тонофиламенты (кератины);
Десминовые филаменты (десмин);
Виментиновые филаменты (виментин);
Нейрофиламенты (NF-L, NF-M, NF-H);
Глиальные

Функции промежуточных филаментов
Поддержание формы клетки;
Распределение органелл в цитоплазме;
Формирование рогового вещества;
Формирование остова
Функции промежуточных филаментов
Поддержание формы клетки;
Распределение органелл в цитоплазме;
Формирование рогового вещества;
Формирование остова

Микрофиламенты
Тонкие белковые полярные нити диаметром 5-7 нм, расположенные в цитоплазме поодиночке,
Микрофиламенты
Тонкие белковые полярные нити диаметром 5-7 нм, расположенные в цитоплазме поодиночке,
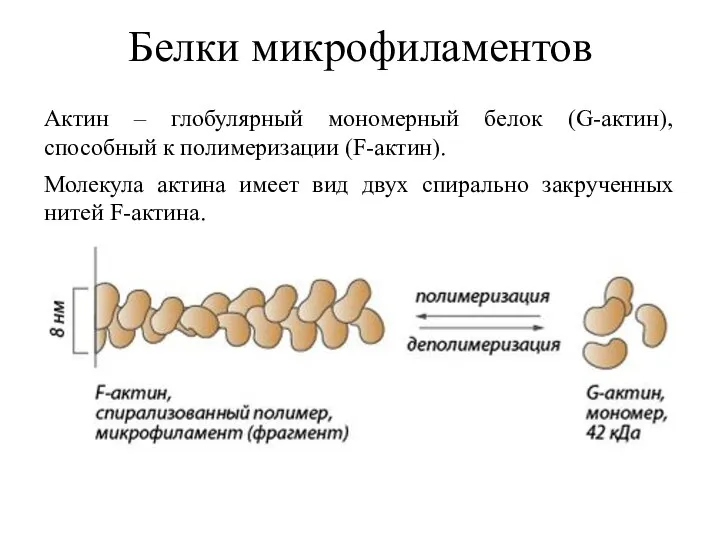
Белки микрофиламентов
Актин – глобулярный мономерный белок (G-актин), способный к полимеризации (F-актин).
Молекула
Белки микрофиламентов
Актин – глобулярный мономерный белок (G-актин), способный к полимеризации (F-актин).
Молекула
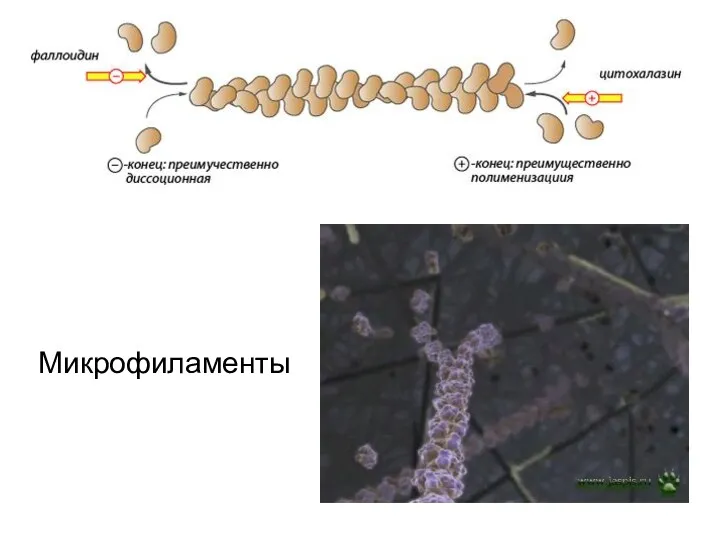
Микрофиламенты
Микрофиламенты

Актин-связывающие белки
ингибирующие полимеризацию актина (профиллин, ДНКаза I);
стабилизирующие белки (тропомиозины);
кэпактины (гельзолин/вилин, фрагмин/северин)
сшивающие
Актин-связывающие белки
ингибирующие полимеризацию актина (профиллин, ДНКаза I);
стабилизирующие белки (тропомиозины);
кэпактины (гельзолин/вилин, фрагмин/северин)
сшивающие
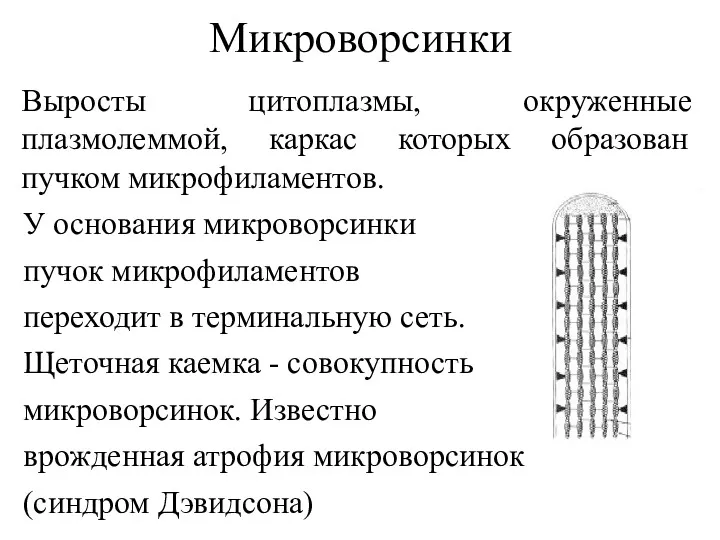
Микроворсинки
Выросты цитоплазмы, окруженные плазмолеммой, каркас которых образован пучком микрофиламентов.
У основания микроворсинки
пучок
Микроворсинки
Выросты цитоплазмы, окруженные плазмолеммой, каркас которых образован пучком микрофиламентов.
У основания микроворсинки
пучок

Функции микрофиламентов
Обеспечение сокращения клеток
Обеспечение движения клеток.
Обеспечение мембранных функций.
Перемещение в цитозоле органелл,
Функции микрофиламентов
Обеспечение сокращения клеток
Обеспечение движения клеток.
Обеспечение мембранных функций.
Перемещение в цитозоле органелл,

Микротрабекулы
Наименее изученная часть цитоскелета.
Выявляются только высоковольтной (мегавольтной) трансмиссионной электронной микроскопией
Микротрабекулы
Наименее изученная часть цитоскелета.
Выявляются только высоковольтной (мегавольтной) трансмиссионной электронной микроскопией
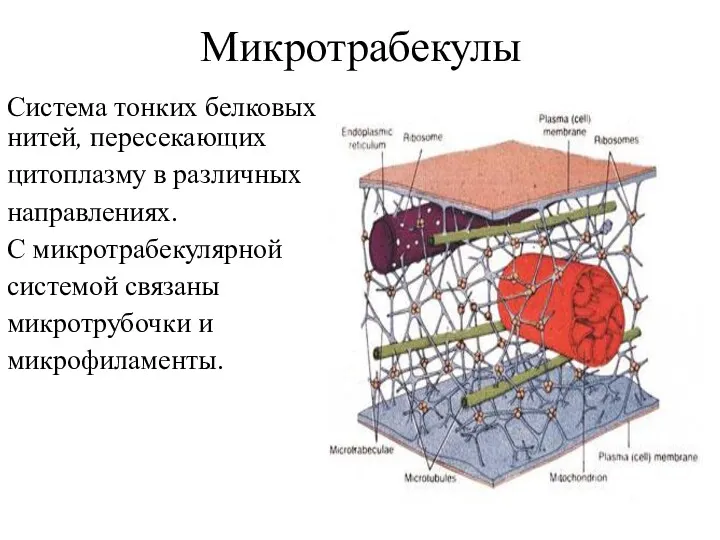
Микротрабекулы
Система тонких белковых нитей, пересекающих
цитоплазму в различных
направлениях.
С микротрабекулярной
Микротрабекулы
Система тонких белковых нитей, пересекающих
цитоплазму в различных
направлениях.
С микротрабекулярной

ВЕЗИКУЛЯРНАЯ СИСТЕМА КЛЕТКИ
1. Гранулярная эндоплазматическая сеть.
2. Агранулярная эндоплазматическая сеть.
3. Аппарат Гольджи.
4.
ВЕЗИКУЛЯРНАЯ СИСТЕМА КЛЕТКИ
1. Гранулярная эндоплазматическая сеть.
2. Агранулярная эндоплазматическая сеть.
3. Аппарат Гольджи.
4.

Эндоплазматическая сеть
Эндоплазматическая сеть представлена трубочками, цистернами, мембранами, пузырьками. Большинство веществ синтезируется
Эндоплазматическая сеть
Эндоплазматическая сеть представлена трубочками, цистернами, мембранами, пузырьками. Большинство веществ синтезируется
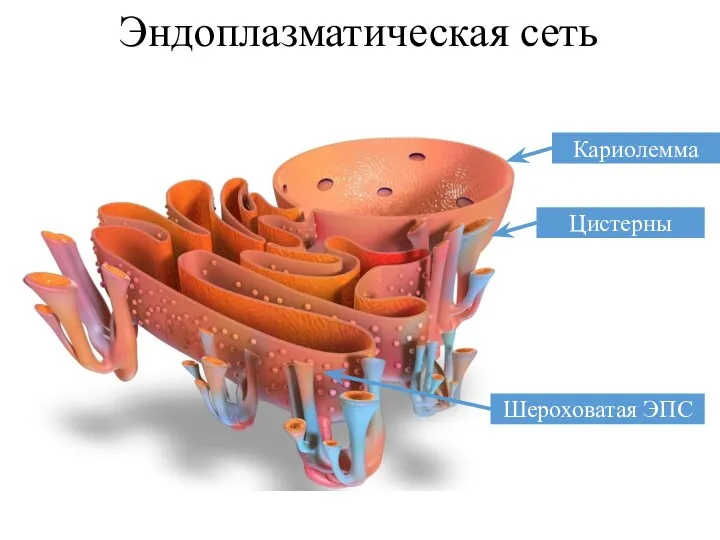
Эндоплазматическая сеть
Кариолемма
Шероховатая ЭПС
Цистерны
Эндоплазматическая сеть
Кариолемма
Шероховатая ЭПС
Цистерны

Является местом конденсации и накопления продуктов секреции, вырабатываемых в других участках
Является местом конденсации и накопления продуктов секреции, вырабатываемых в других участках
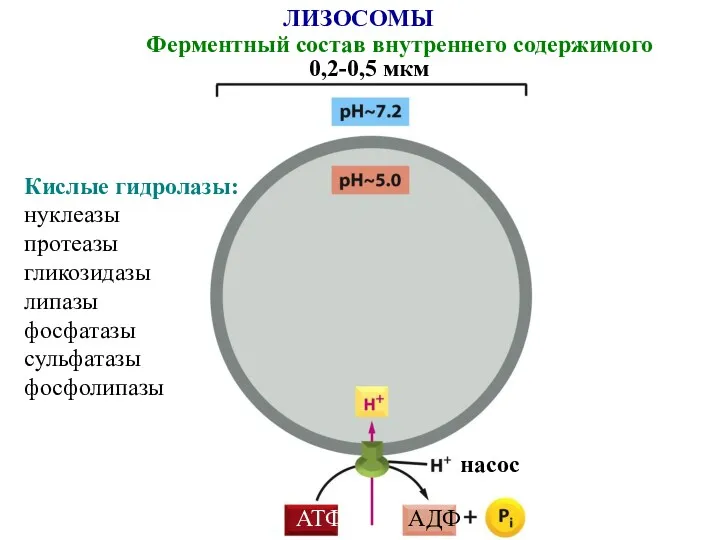
ЛИЗОСОМЫ
Ферментный состав внутреннего содержимого
0,2-0,5 мкм
насос
АТФ
АДФ
Кислые гидролазы:
нуклеазы
протеазы
гликозидазы
липазы
фосфатазы
сульфатазы
фосфолипазы
ЛИЗОСОМЫ
Ферментный состав внутреннего содержимого
0,2-0,5 мкм
насос
АТФ
АДФ
Кислые гидролазы:
нуклеазы
протеазы
гликозидазы
липазы
фосфатазы
сульфатазы
фосфолипазы
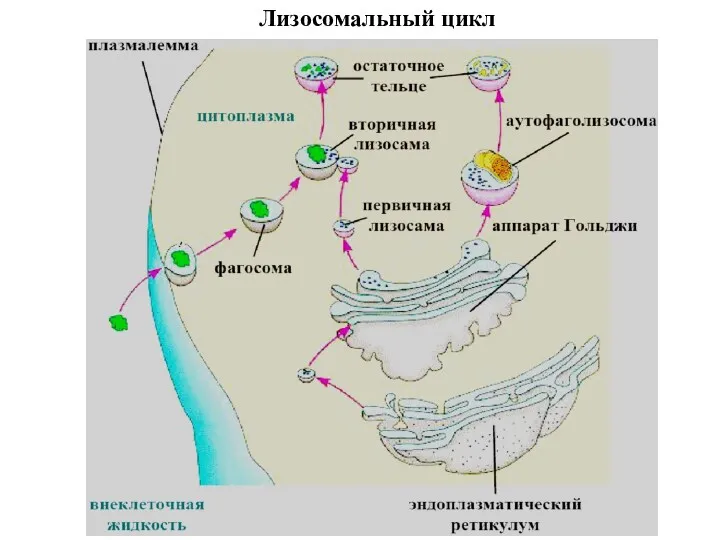
Лизосомальный цикл
Лизосомальный цикл

Пероксисомы – основной центр утилизации кислорода в клетках, с последующим образованием
Пероксисомы – основной центр утилизации кислорода в клетках, с последующим образованием
 Тонкая кишка
Тонкая кишка Полость рта
Полость рта Физиология спорта. Физиологические основы классификации физических упражнений
Физиология спорта. Физиологические основы классификации физических упражнений Посттрансляционные модификации белков
Посттрансляционные модификации белков Зоология беспозвоночных
Зоология беспозвоночных Свойства воды. Интерактивный кроссворд
Свойства воды. Интерактивный кроссворд Нервная система
Нервная система Опорно-двигательный аппарат
Опорно-двигательный аппарат Окружающая среда
Окружающая среда Микрофлора пищевых продуктов и кулинарной продукции
Микрофлора пищевых продуктов и кулинарной продукции Пагін і його будова
Пагін і його будова Микробиология ойын
Микробиология ойын Division Ranunculаles and Polygonales
Division Ranunculаles and Polygonales Пищеварительная система
Пищеварительная система Периоды становления микробиологии. Систематика и номенклатура микробов
Периоды становления микробиологии. Систематика и номенклатура микробов Механизмы половой дифференцировки
Механизмы половой дифференцировки Фотосинтез. Хемосинтез
Фотосинтез. Хемосинтез Спланхнология. Учение о внутренностях
Спланхнология. Учение о внутренностях Школьная клумба
Школьная клумба обмен веществ и энергии в клетке.
обмен веществ и энергии в клетке. Метаболизм клетки. Клеточное дыхание. Фотосинтез, хемосинтез
Метаболизм клетки. Клеточное дыхание. Фотосинтез, хемосинтез Презентация урока 6 класс ФГОС.
Презентация урока 6 класс ФГОС. Функциональная анатомия сердечно-сосудистой системы
Функциональная анатомия сердечно-сосудистой системы Оригинальная технология эффективных микроорганизмов в России: микробиологическое удобрение Восток ЭМ-1
Оригинальная технология эффективных микроорганизмов в России: микробиологическое удобрение Восток ЭМ-1 Гортань как орган фонации
Гортань как орган фонации Все про кошек
Все про кошек Генетика пола. Наследование,сцепленное с полом
Генетика пола. Наследование,сцепленное с полом Речные рыбы
Речные рыбы