- Ферменты

Содержание
- 2. Ферменты (энзимы) специфические и высокоэффективные катализаторы химических реакций, протекающих в живой клетке Энзимология один из главных
- 3. Обмен веществ был бы невозможен без участия биологических катализаторов – ферментов Биокаталитически ускоряются самые различные превращения
- 4. Сходство и отличия ферментов и традиционных катализаторов
- 5. не создают реакции, а лишь ускоряют достижение равновесия ускоряют реакции за счет снижения энергии активации Катализаторы
- 6. Н2О2 → Н2О + ½ О2 Еа = 75,24 кДж/моль (18 ккал/моль) Мелкодисперсная Pt ускоряет реакцию
- 7. Бо́льшая эффективность Отличия ферментов и традиционных катализаторов
- 8. Ферменты проявляют высокую каталитическую активность в очень мягких условиях температура (?) давление (?) рН (?) Гидролиз
- 9. Высокая специфичность действия пределы ее у разных ферментов различны Отличия ферментов и традиционных катализаторов
- 10. Виды специфичности ферментов Относительная (групповая) Наибольшее значение – тип химической связи в молекуле субстрата Пепсин расщепляет
- 11. Виды специфичности ферментов Абсолютная Фермент катализирует превращение одного-единственного субстрата Аргиназа ускоряет гидролиз аргинина Уреаза – мочевины
- 12. Виды специфичности ферментов стереохимическая обусловлена существованием оптически активных субстратов оксидазы L- и D-аминокислот аспартатдекарбоксилаза (отщепление СО2
- 13. Аконитатгидратаза
- 14. Отличия, связанные с белковой природой ферментов (термолабильность, зависимость активности от рН, наличие активаторов и ингибиторов и
- 15. Строение ферментов
- 16. Природа ферментативного катализа Фермент (энзим) – катализатор, ускоряющий только одну химическую реакцию Фермент – белок Белки
- 17. Простые (однокомпонентные) Сложные (двухкомпонентные)
- 18. Однокомпонентные ферменты (ферменты-протеины) Каталитический центр Субстратный центр Субстратный центр может совпадать (или перекрываться) с каталитическим Каталитический
- 19. Трипсин Каталитический центр Субстратный центр Активный центр
- 20. Двухкомпонентные ферменты (ферменты-протеиды) Простетическая группа Кофермент 1. Белковая часть – апофермент
- 21. Природа коферментов (О. Г. Варбург, Р. Кун, П. Каррер) Витамины (Е, К, Q, В1, В2, В6,
- 22. построены из нескольких субъединиц – протомеров каталаза – 6 протомеров Максимальная каталитическая активность Наряду с каталитически
- 23. лактатдегидрогеназа НННН, НННМ, ННММ, НМММ, ММММ H – от англ. heart – сердце M – от
- 25. отличаются по степени активности по некоторым физическим свойствам по локализации в органах и тканях и т.
- 26. ферменты, способные одновременно ускорять несколько химических реакций и осуществлять сложные превращения субстрата пируватдегидрогеназный комплекс Мультиэнзимы
- 27. Модель пируватдегидрогеназного комплекса 12 димерных молекул пируватдекарбоксилазы 6 димерных молекул дигидролипоилдегидрогеназы 24 молекулы липоат-ацетилтрансферазы
- 28. мультиэнзимный комплекс, обслуживающий единый многоступенчатый процесс биохимических превращений метаболоны гликолиза, цикла Кребса и т. д. Метаболон
- 29. состоят из одной полипептидной цепи, содержащей несколько доменов, каждый из которых характеризуется своей каталитической активностью Полифункциональные
- 30. Ферменты и ферментные комплексы
- 31. Физиологический смысл мультиэнзимных комплексов и полифункциональных ферментов слаженность во времени и пространстве действия ферментов или доменов
- 32. Кинетика ферментативных реакций (самостоятельно + лабораторные занятия)
- 33. Номенклатура и классификация ферментов
- 34. Номенклатура ферментов
- 35. 1. Тривиальная номенклатура пепсин – от греч. пепсис – пищеварение трипсин – от греч. трипсис –
- 36. 2. Рациональная номенклатура Название субстрата + окончание –аза (1883 г., П. Э. Дюкло) амилаза – от
- 37. 3. По названию простетической группы (кофермента) геминфермент (простетическая группа – гем) пиридоксальфермент (кофермент – пиридоксаль, витамин
- 38. 4. По характеру субстрата и типу катализируемой реакции Сукцинатдегидрогеназа НООС–СН2–СН2–СООН → НООС–СН=СН–СООН + 2Н янтарная кислота
- 39. 5. Международная (московская) номенклатура V Международный биохимический конгресс, 10–16 августа 1961 г., Москва Химическое название субстрата
- 40. пиридоксальфермент, ускоряющий реакцию переаминирования между L-ала и α-кетоглутаровой кислотой L-аланин: 2-оксоглутаратаминотрансфераза
- 41. Названия ферментов выигрывают в точности, НО в ряде случаев гораздо сложнее тривиальных уреаза – карбамид-амидогидролаза трегалаза
- 42. Классификация ферментов
- 43. Список ферментов
- 44. Международной комиссией был составлен детальный список ферментов, в котором на основании классификации каждому ферменту присвоен шифр,
- 45. 1 число Класс Оксидоредуктазы Трансферазы Гидролазы Лиазы Изомеразы Лигазы (синтетазы)
- 46. 2 число Подкласс у оксидоредуктаз показывает природу той группы в молекуле донора, которая подвергается окислению (1
- 47. 3 число Подподкласс у оксидоредуктаз – тип участвующего в реакции акцептора (1 – НАД или НАДФ;
- 48. 4 число Порядковый номер Уреаза 3.5.1.5 3 – класс гидролаз 5 – подкласс С–N-гидролаз 1 –
- 49. Оксидоредуктазы
- 50. I. Оксидоредуктазы ускоряют ОВР Окисление – процесс отнятия атомов Н или электронов от субстрата, а восстановление
- 51. Особенности оксидоредуктаз Способность образовывать цепи, в которых осуществляется многоступенчатый перенос атомов Н или электронов от первичного
- 52. Особенности оксидоредуктаз Аэробные дегидрогеназы (оксидазы) переносят атомы Н или электроны непосредственно на атомы О Анаэробные дегидрогеназы
- 53. Особенности оксидоредуктаз Первичные дегидрогеназы катализируют реакции отнятия атомов Н непосредственно от окисляемого вещества (первичного субстрата) Вторичные
- 54. Особенности оксидоредуктаз Двухкомпонентные ферменты с ограниченным набором коферментов, но способные ускорять большое число самых разнообразных ОВР,
- 55. Особенности оксидоредуктаз Пиридинпротеины Аэробные дегидрогеназы Пример – алкогольдегидрогеназа Коферменты никотинамидадениндинуклеотид (NAD+, НАД+) никотинамидадениндинуклеотидфосфат (NADP+, НАДФ+)
- 57. Особенности оксидоредуктаз Флавопротеины (ФП) Партнеры восстановленных форм пиридинпротеинов в окислительно-восстановительной цепи В большинстве случаев – вторичные
- 59. FAD
- 60. Особенности оксидоредуктаз Другие коферменты оксидоредуктаз хиноны (убихинон – витамин Q) железопорфирины (цитохромы)
- 61. Особенности оксидоредуктаз Ускоряют протекание реакций, связанных с высвобождением энергии (цитохромная система)
- 62. Трансферазы
- 63. II. Трансферазы ускоряют реакции переноса функциональных групп и молекулярных остатков от одного соединения к другому
- 64. Подклассы Фосфотрансферазы (киназы) переносят фосфатные группы с участием АТФ Гексокиназа (АТФ: D-глюкоза-6-фосфотрансфераза)
- 65. глюкоза АТФ Глюкозо-6-фосфат (Г-6-Ф) АДФ
- 66. Подклассы Аминотрансферазы (трансаминазы) ускоряют реакции переаминирования аминокислот с кетокислотами Кофермент – пиридоксальфосфат
- 67. Подклассы Гликозилтрансферазы ускоряют реакции переноса остатков сахаров на различные акцепторы фосфорилазы катализируют реакции переноса гликозильных групп
- 68. сахароза глюкозо-1-фосфат (Г-1-Ф) фруктоза
- 69. Подклассы Ацилтрансферазы ускоряют перенос ацильных групп на аминокислоты, амины, спирты и др. Кофермент – коэнзим А
- 70. ацетил-КоА холин ацетилхолин
- 71. Подклассы ферменты, ускоряющие перенос одноуглеродных фрагментов Метилтрансферазы Оксиметилтрансферазы Формилтрансферазы и др. нуклеотидных остатков Нуклеотидилтрансферазы
- 72. Гидролазы
- 73. III. Гидролазы ускоряют реакции гидролиза органических соединений Делятся на подклассы по характеру субстрата
- 74. Подклассы Эстеразы ускоряют гидролиз сложных эфиров Липаза ускоряет гидролиз α-сложноэфирных связей в молекулах триглицеридов (жиров) Фосфатазы
- 75. Подклассы Гликозидазы ускоряют гидролиз гликозидов Мальтаза Сахараза Амилаза
- 76. Подклассы Пептидазы (пептид-гидролазы) ускоряют гидролиз пептидных связей в белках и пептидах протеиназы (пептидил-пептидогидролазы) ускоряют гидролиз внутренних
- 77. Подклассы Амидазы ускоряют гидролиз амидов кислот уреаза (гидролиз мочевины до NH3 и СО2) аспарагиназа и глутаминаза
- 78. Лиазы
- 79. IV. Лиазы ускоряют реакции негидролитического распада органических соединений При этом замыкаются двойные связи и выделяются такие
- 80. Подклассы С–С-лиазы декарбоксилазы кетокислот (кофермент – витамин В1) декарбокислазы аминокислот (кофермент – витамин В6) альдегид-лиазы альдолаза
- 81. фруктозо-1,6-дифосфат (Ф-1,6-дФ) диоксиацетонфосфат (ДОАФ) 3-фосфоглицериновый альдегид (3-ФГА)
- 82. Подклассы С–О-лиазы (гидратазы) ускоряют гидратацию и дегидратацию органических соединений Фумаратгидратаза НООС–СН(ОН)–СН2–СООН ⇄ НООС–СН=СН–СООН + Н2О Яблочная
- 83. Подклассы С–N-лиазы аспартат-аммиак-лиаза ускоряет прямое дезаминирование асп НООС–СН(NH2)–СН2–СООН ⇄ НООС–СН=СН–СООNH4 Аспарагиновая кислота (асп) Фумарат аммония
- 84. Изомеразы
- 85. V. Изомеразы ускоряют геометрические или структурные изменения в пределах одной молекулы
- 86. Подклассы Рацемазы и эпимеразы Мутаротаза α-D-глюкопираноза ⇄ β-D-глюкопираноза Цис-транс-изомеразы Ретинол-изомераза
- 87. Подклассы Внутримолекулярные трансферазы (мутазы) Фосфоглицерат-фосфомутаза 3-фосфоглицериновая кислота (3-ФГК) 2-фосфоглицериновая кислота (2-ФГК)
- 88. Подклассы Внутримолекулярные оксидоредуктазы триозофосфатизомераза Диоксиацетонфосфат (ДОАФ) 3-фосфоглицериновый альдегид (3-ФГА)
- 89. Лигазы
- 90. VI. Лигазы Ускоряют присоединение друг к другу двух молекул Процесс сопряжен с разрывом макроэргической связи АТФ;
- 92. Скачать презентацию

Ферменты (энзимы)
специфические и высокоэффективные катализаторы химических реакций, протекающих в живой клетке
Энзимология
один
Ферменты (энзимы)
специфические и высокоэффективные катализаторы химических реакций, протекающих в живой клетке
Энзимология
один

Обмен веществ был бы невозможен без участия биологических катализаторов – ферментов
Биокаталитически
Обмен веществ был бы невозможен без участия биологических катализаторов – ферментов
Биокаталитически

Сходство и отличия ферментов и традиционных катализаторов
Сходство и отличия ферментов и традиционных катализаторов

не создают реакции, а лишь ускоряют достижение равновесия
ускоряют реакции за счет
не создают реакции, а лишь ускоряют достижение равновесия
ускоряют реакции за счет

Н2О2 → Н2О + ½ О2
Еа = 75,24 кДж/моль (18 ккал/моль)
Мелкодисперсная
Н2О2 → Н2О + ½ О2
Еа = 75,24 кДж/моль (18 ккал/моль)
Мелкодисперсная

Бо́льшая эффективность
Отличия ферментов и традиционных катализаторов
Бо́льшая эффективность
Отличия ферментов и традиционных катализаторов

Ферменты проявляют высокую каталитическую активность в очень мягких условиях
температура (?)
давление (?)
Ферменты проявляют высокую каталитическую активность в очень мягких условиях
температура (?)
давление (?)

Высокая специфичность действия
пределы ее у разных ферментов различны
Отличия ферментов и традиционных
Высокая специфичность действия
пределы ее у разных ферментов различны
Отличия ферментов и традиционных

Виды специфичности ферментов
Относительная (групповая)
Наибольшее значение – тип химической связи в молекуле
Виды специфичности ферментов
Относительная (групповая)
Наибольшее значение – тип химической связи в молекуле

Виды специфичности ферментов
Абсолютная
Фермент катализирует превращение одного-единственного субстрата
Аргиназа ускоряет гидролиз аргинина
Уреаза
Виды специфичности ферментов
Абсолютная
Фермент катализирует превращение одного-единственного субстрата
Аргиназа ускоряет гидролиз аргинина
Уреаза

Виды специфичности ферментов
стереохимическая
обусловлена существованием
оптически активных субстратов
оксидазы L- и D-аминокислот
аспартатдекарбоксилаза (отщепление
Виды специфичности ферментов
стереохимическая
обусловлена существованием
оптически активных субстратов
оксидазы L- и D-аминокислот
аспартатдекарбоксилаза (отщепление
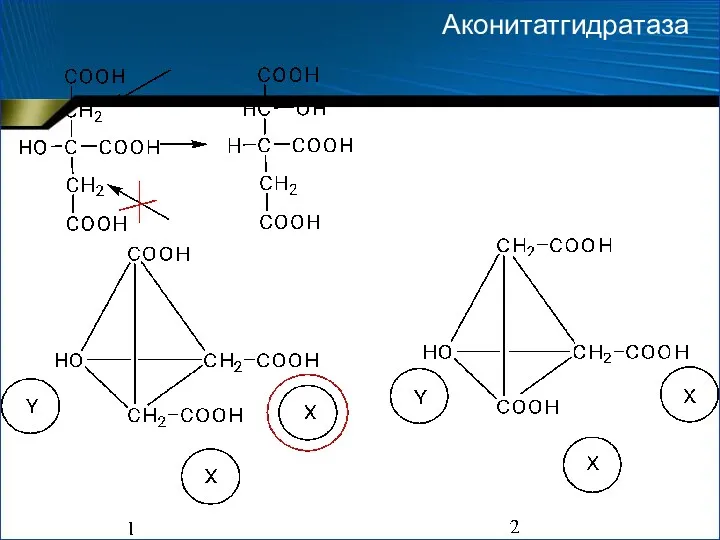
Аконитатгидратаза
Аконитатгидратаза

Отличия, связанные с белковой природой ферментов (термолабильность, зависимость активности от рН,
Отличия, связанные с белковой природой ферментов (термолабильность, зависимость активности от рН,

Строение ферментов
Строение ферментов
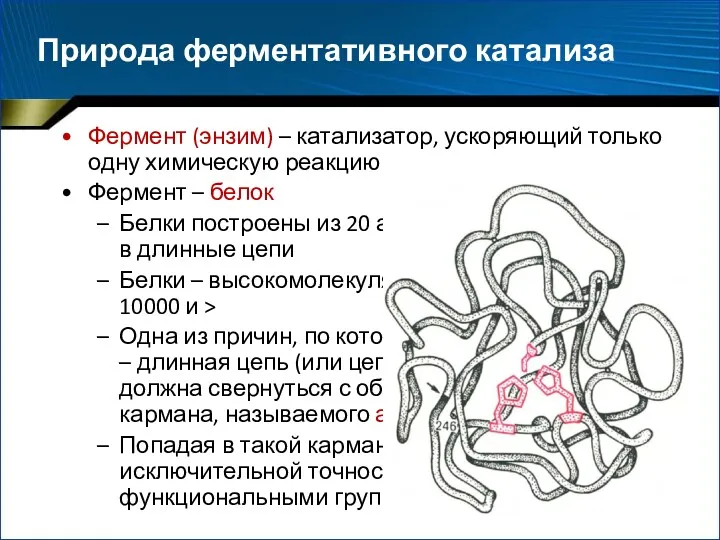
Природа ферментативного катализа
Фермент (энзим) – катализатор, ускоряющий только одну химическую
Природа ферментативного катализа
Фермент (энзим) – катализатор, ускоряющий только одну химическую
Простые
(однокомпонентные)
Сложные
(двухкомпонентные)
Простые
(однокомпонентные)
Сложные
(двухкомпонентные)

Однокомпонентные ферменты (ферменты-протеины)
Каталитический центр
Субстратный центр
Субстратный центр может совпадать (или перекрываться)
Однокомпонентные ферменты (ферменты-протеины)
Каталитический центр
Субстратный центр
Субстратный центр может совпадать (или перекрываться)
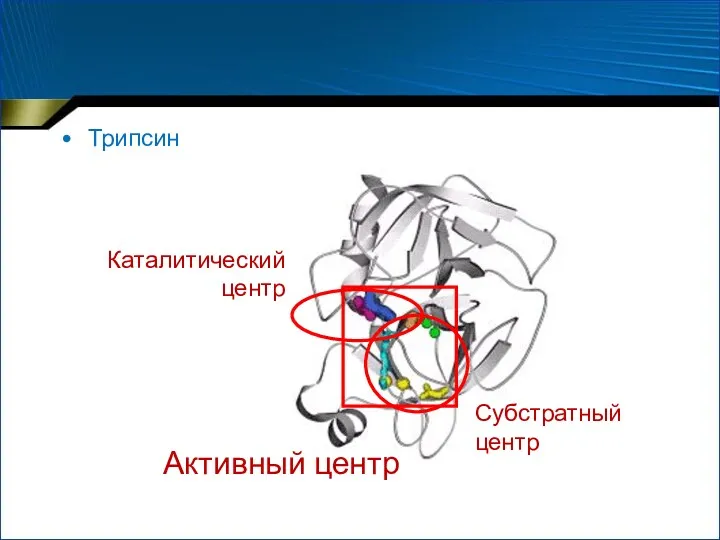
Трипсин
Каталитический центр
Субстратный центр
Активный центр
Трипсин
Каталитический центр
Субстратный центр
Активный центр
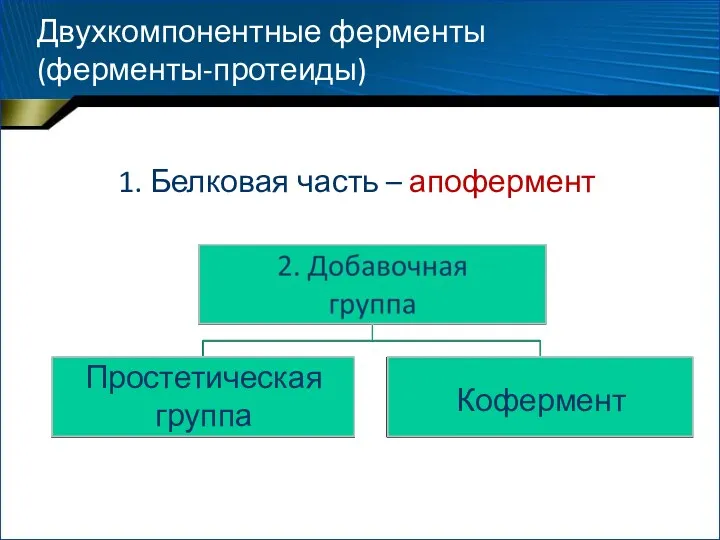
Двухкомпонентные ферменты (ферменты-протеиды)
Простетическая группа
Кофермент
1. Белковая часть – апофермент
Двухкомпонентные ферменты (ферменты-протеиды)
Простетическая группа
Кофермент
1. Белковая часть – апофермент

Природа коферментов
(О. Г. Варбург, Р. Кун, П. Каррер)
Витамины (Е, К,
Природа коферментов
(О. Г. Варбург, Р. Кун, П. Каррер)
Витамины (Е, К,
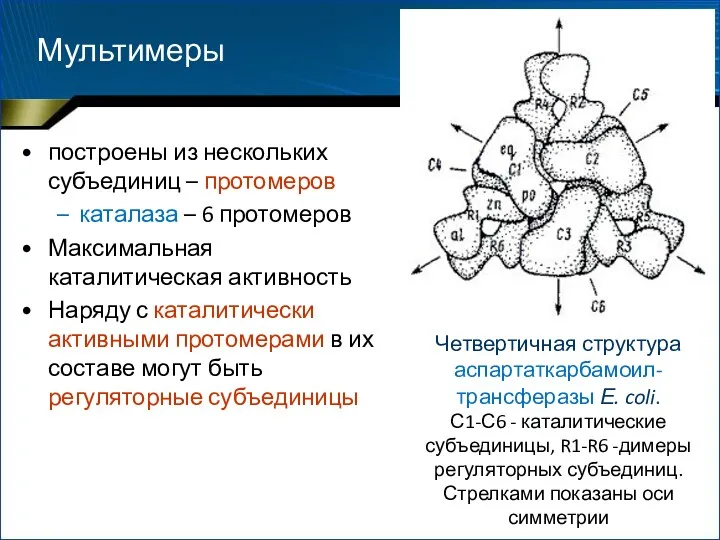
построены из нескольких субъединиц – протомеров
каталаза – 6 протомеров
Максимальная каталитическая активность
Наряду
построены из нескольких субъединиц – протомеров
каталаза – 6 протомеров
Максимальная каталитическая активность
Наряду

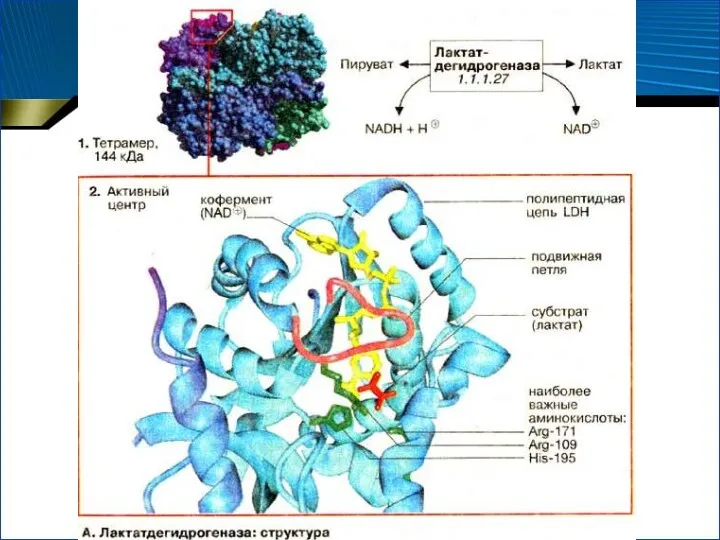
лактатдегидрогеназа
НННН, НННМ, ННММ, НМММ, ММММ
H – от англ. heart –
лактатдегидрогеназа
НННН, НННМ, ННММ, НМММ, ММММ
H – от англ. heart –

отличаются
по степени активности
по некоторым физическим свойствам
по локализации в органах и
отличаются
по степени активности
по некоторым физическим свойствам
по локализации в органах и

ферменты, способные одновременно ускорять несколько химических реакций и осуществлять сложные превращения
ферменты, способные одновременно ускорять несколько химических реакций и осуществлять сложные превращения
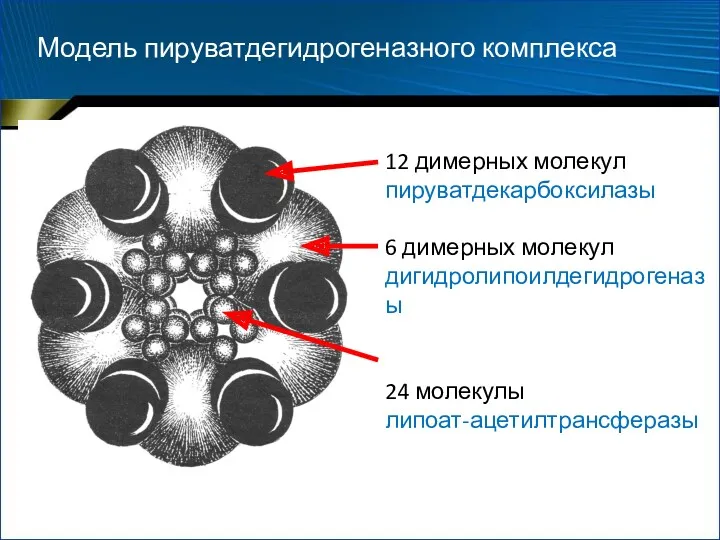
Модель пируватдегидрогеназного комплекса
12 димерных молекул пируватдекарбоксилазы
6 димерных молекул дигидролипоилдегидрогеназы
24 молекулы
липоат-ацетилтрансферазы
Модель пируватдегидрогеназного комплекса
12 димерных молекул пируватдекарбоксилазы
6 димерных молекул дигидролипоилдегидрогеназы
24 молекулы
липоат-ацетилтрансферазы
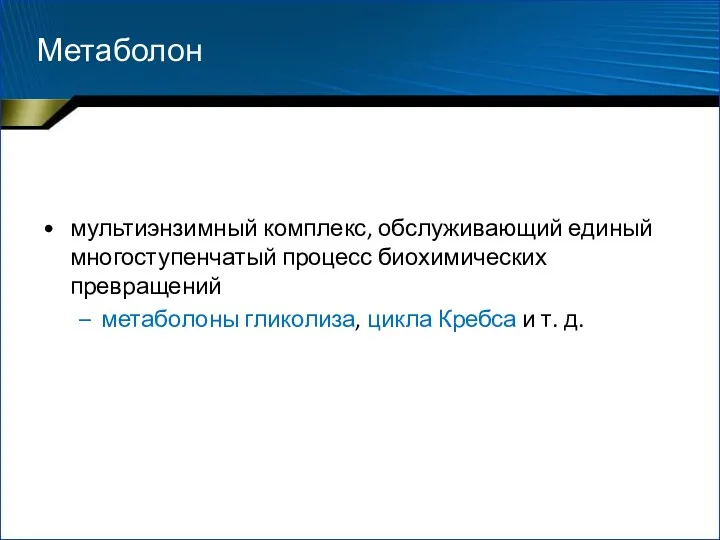
мультиэнзимный комплекс, обслуживающий единый многоступенчатый процесс биохимических превращений
метаболоны гликолиза, цикла Кребса
мультиэнзимный комплекс, обслуживающий единый многоступенчатый процесс биохимических превращений
метаболоны гликолиза, цикла Кребса

состоят из одной полипептидной цепи, содержащей несколько доменов, каждый из которых
состоят из одной полипептидной цепи, содержащей несколько доменов, каждый из которых
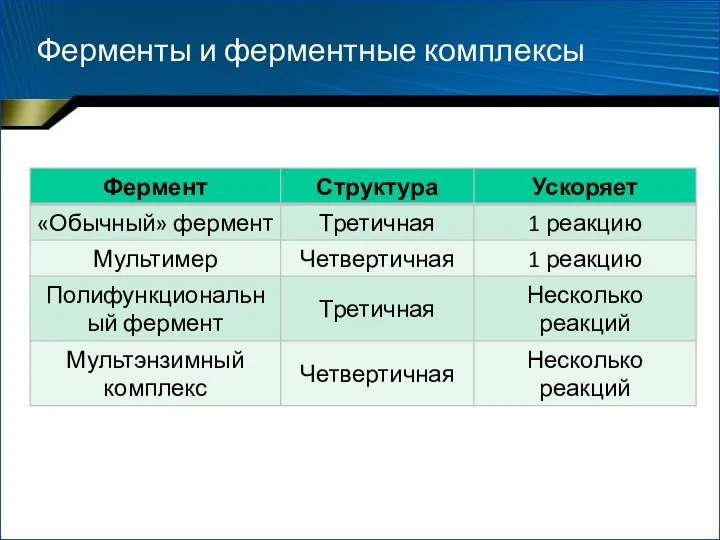
Ферменты и ферментные комплексы
Ферменты и ферментные комплексы

Физиологический смысл мультиэнзимных комплексов и полифункциональных ферментов
слаженность во времени и пространстве
Физиологический смысл мультиэнзимных комплексов и полифункциональных ферментов
слаженность во времени и пространстве

Кинетика ферментативных реакций (самостоятельно + лабораторные занятия)
Кинетика ферментативных реакций (самостоятельно + лабораторные занятия)

Номенклатура и классификация ферментов
Номенклатура и классификация ферментов

Номенклатура ферментов
Номенклатура ферментов

1. Тривиальная номенклатура
пепсин – от греч. пепсис – пищеварение
трипсин – от
1. Тривиальная номенклатура
пепсин – от греч. пепсис – пищеварение
трипсин – от

2. Рациональная номенклатура
Название субстрата + окончание –аза (1883 г., П. Э.
2. Рациональная номенклатура
Название субстрата + окончание –аза (1883 г., П. Э.

3. По названию простетической группы (кофермента)
геминфермент (простетическая группа – гем)
пиридоксальфермент (кофермент
3. По названию простетической группы (кофермента)
геминфермент (простетическая группа – гем)
пиридоксальфермент (кофермент

4. По характеру субстрата и типу катализируемой реакции
Сукцинатдегидрогеназа
НООС–СН2–СН2–СООН → НООС–СН=СН–СООН +
4. По характеру субстрата и типу катализируемой реакции
Сукцинатдегидрогеназа
НООС–СН2–СН2–СООН → НООС–СН=СН–СООН +

5. Международная (московская) номенклатура
V Международный биохимический конгресс, 10–16 августа 1961
5. Международная (московская) номенклатура
V Международный биохимический конгресс, 10–16 августа 1961

пиридоксальфермент, ускоряющий реакцию переаминирования между L-ала и α-кетоглутаровой кислотой
L-аланин: 2-оксоглутаратаминотрансфераза
пиридоксальфермент, ускоряющий реакцию переаминирования между L-ала и α-кетоглутаровой кислотой
L-аланин: 2-оксоглутаратаминотрансфераза

Названия ферментов выигрывают в точности, НО в ряде случаев гораздо сложнее
Названия ферментов выигрывают в точности, НО в ряде случаев гораздо сложнее

Классификация ферментов
Классификация ферментов

Список ферментов
Список ферментов

Международной комиссией был составлен детальный список ферментов, в котором на основании
Международной комиссией был составлен детальный список ферментов, в котором на основании

1 число
Класс
Оксидоредуктазы
Трансферазы
Гидролазы
Лиазы
Изомеразы
Лигазы (синтетазы)
1 число
Класс
Оксидоредуктазы
Трансферазы
Гидролазы
Лиазы
Изомеразы
Лигазы (синтетазы)

2 число
Подкласс
у оксидоредуктаз показывает природу той группы в молекуле донора, которая
2 число
Подкласс
у оксидоредуктаз показывает природу той группы в молекуле донора, которая
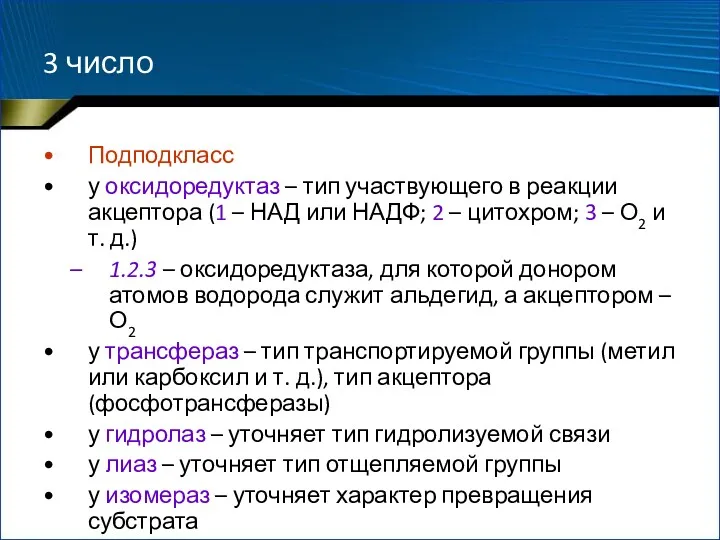
3 число
Подподкласс
у оксидоредуктаз – тип участвующего в реакции акцептора (1 –
3 число
Подподкласс
у оксидоредуктаз – тип участвующего в реакции акцептора (1 –

4 число
Порядковый номер
Уреаза 3.5.1.5
3 – класс гидролаз
5 – подкласс С–N-гидролаз
1 –
4 число
Порядковый номер
Уреаза 3.5.1.5
3 – класс гидролаз
5 – подкласс С–N-гидролаз
1 –

Оксидоредуктазы
Оксидоредуктазы

I. Оксидоредуктазы
ускоряют ОВР
Окисление – процесс отнятия атомов Н или электронов
I. Оксидоредуктазы
ускоряют ОВР
Окисление – процесс отнятия атомов Н или электронов

Особенности оксидоредуктаз
Способность образовывать цепи, в которых осуществляется многоступенчатый перенос атомов
Особенности оксидоредуктаз
Способность образовывать цепи, в которых осуществляется многоступенчатый перенос атомов
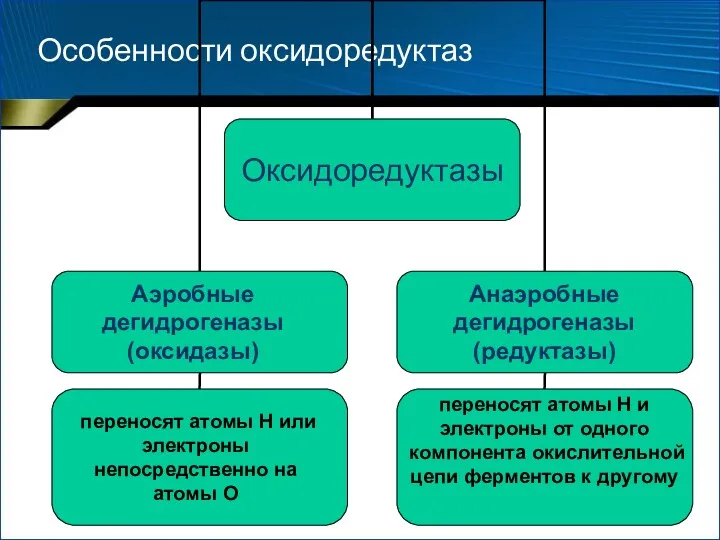
Особенности оксидоредуктаз
Аэробные дегидрогеназы (оксидазы)
переносят атомы Н или электроны непосредственно на
Особенности оксидоредуктаз
Аэробные дегидрогеназы (оксидазы)
переносят атомы Н или электроны непосредственно на
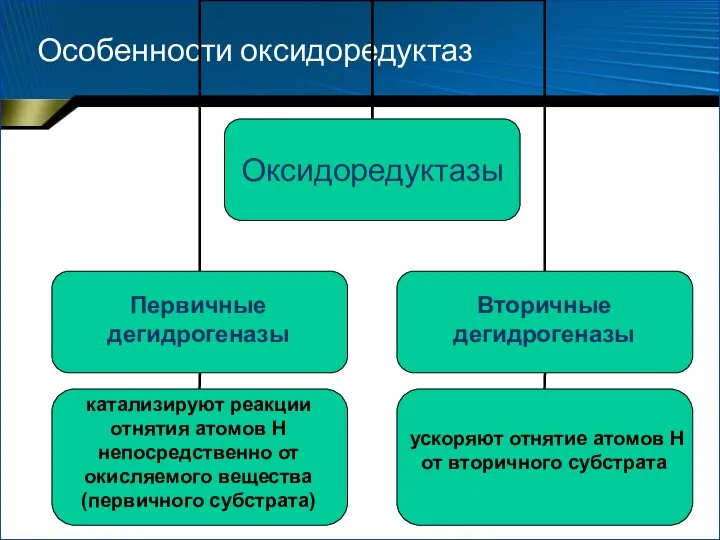
Особенности оксидоредуктаз
Первичные дегидрогеназы
катализируют реакции отнятия атомов Н непосредственно от окисляемого
Особенности оксидоредуктаз
Первичные дегидрогеназы
катализируют реакции отнятия атомов Н непосредственно от окисляемого

Особенности оксидоредуктаз
Двухкомпонентные ферменты с ограниченным набором коферментов, но способные ускорять
Особенности оксидоредуктаз
Двухкомпонентные ферменты с ограниченным набором коферментов, но способные ускорять

Особенности оксидоредуктаз
Пиридинпротеины
Аэробные дегидрогеназы
Пример – алкогольдегидрогеназа
Коферменты
никотинамидадениндинуклеотид (NAD+, НАД+)
никотинамидадениндинуклеотидфосфат (NADP+, НАДФ+)
Особенности оксидоредуктаз
Пиридинпротеины
Аэробные дегидрогеназы
Пример – алкогольдегидрогеназа
Коферменты
никотинамидадениндинуклеотид (NAD+, НАД+)
никотинамидадениндинуклеотидфосфат (NADP+, НАДФ+)
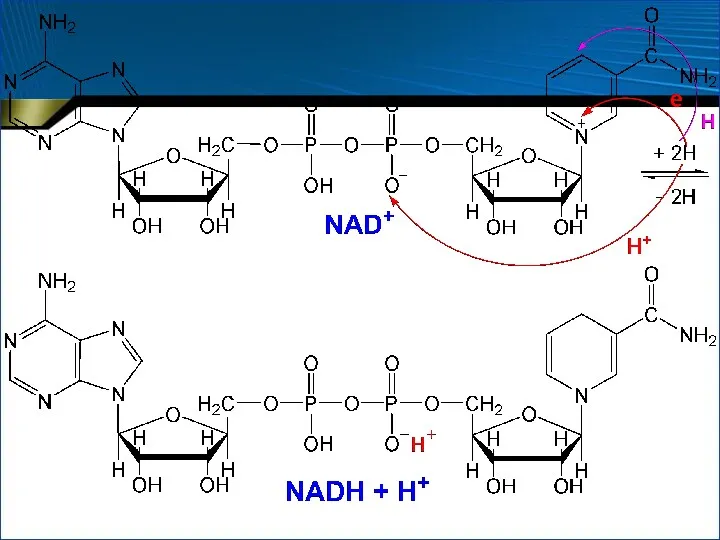

Особенности оксидоредуктаз
Флавопротеины (ФП)
Партнеры восстановленных форм пиридинпротеинов в окислительно-восстановительной цепи
В большинстве
Особенности оксидоредуктаз
Флавопротеины (ФП)
Партнеры восстановленных форм пиридинпротеинов в окислительно-восстановительной цепи
В большинстве
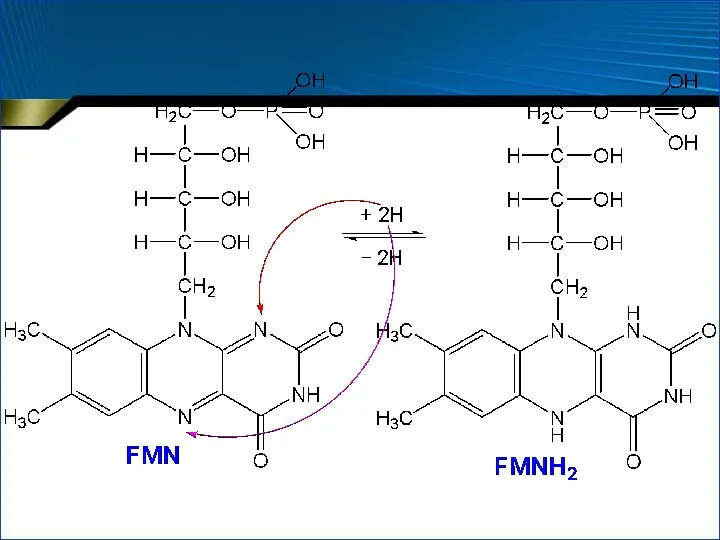
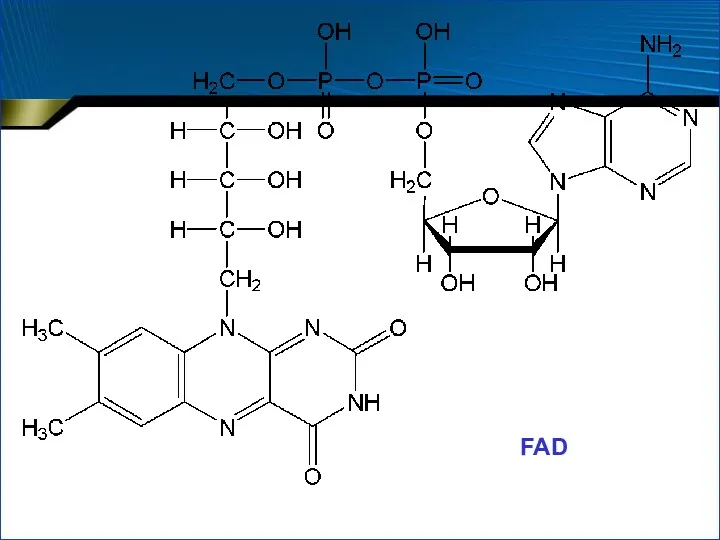
FAD
FAD

Особенности оксидоредуктаз
Другие коферменты оксидоредуктаз
хиноны (убихинон – витамин Q)
железопорфирины
Особенности оксидоредуктаз
Другие коферменты оксидоредуктаз
хиноны (убихинон – витамин Q)
железопорфирины

Особенности оксидоредуктаз
Ускоряют протекание реакций, связанных с высвобождением энергии (цитохромная система)
Особенности оксидоредуктаз
Ускоряют протекание реакций, связанных с высвобождением энергии (цитохромная система)

Трансферазы
Трансферазы

II. Трансферазы
ускоряют реакции переноса функциональных групп и молекулярных остатков от
II. Трансферазы
ускоряют реакции переноса функциональных групп и молекулярных остатков от

Подклассы
Фосфотрансферазы (киназы) переносят фосфатные группы с участием АТФ
Гексокиназа (АТФ: D-глюкоза-6-фосфотрансфераза)
Подклассы
Фосфотрансферазы (киназы) переносят фосфатные группы с участием АТФ
Гексокиназа (АТФ: D-глюкоза-6-фосфотрансфераза)
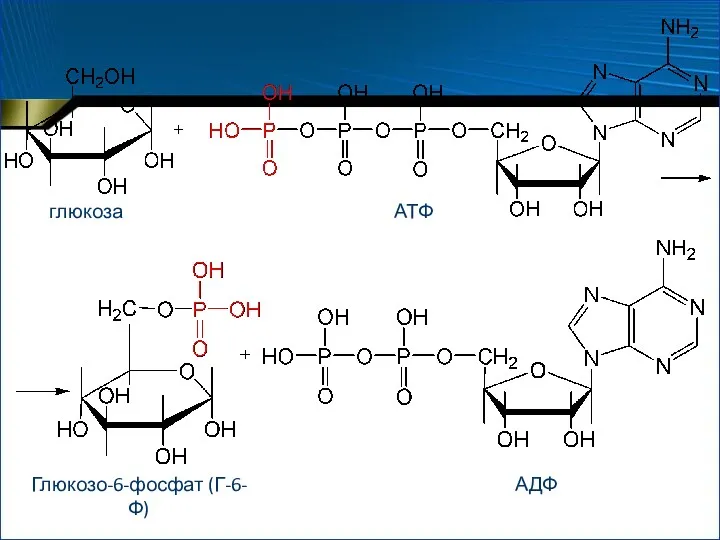
глюкоза
АТФ
Глюкозо-6-фосфат (Г-6-Ф)
АДФ
глюкоза
АТФ
Глюкозо-6-фосфат (Г-6-Ф)
АДФ

Подклассы
Аминотрансферазы (трансаминазы) ускоряют реакции переаминирования аминокислот с кетокислотами
Кофермент – пиридоксальфосфат
Подклассы
Аминотрансферазы (трансаминазы) ускоряют реакции переаминирования аминокислот с кетокислотами
Кофермент – пиридоксальфосфат

Подклассы
Гликозилтрансферазы ускоряют реакции переноса остатков сахаров на различные акцепторы
фосфорилазы катализируют
Подклассы
Гликозилтрансферазы ускоряют реакции переноса остатков сахаров на различные акцепторы
фосфорилазы катализируют
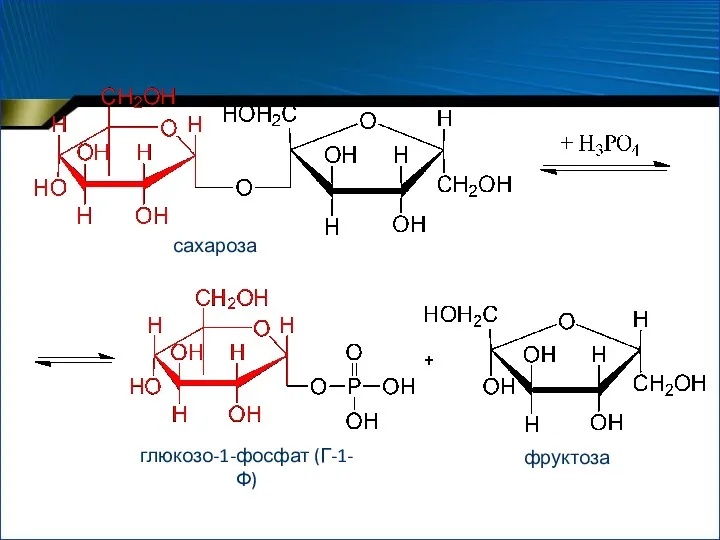
сахароза
глюкозо-1-фосфат (Г-1-Ф)
фруктоза
сахароза
глюкозо-1-фосфат (Г-1-Ф)
фруктоза

Подклассы
Ацилтрансферазы ускоряют перенос ацильных групп на аминокислоты, амины, спирты и
Подклассы
Ацилтрансферазы ускоряют перенос ацильных групп на аминокислоты, амины, спирты и
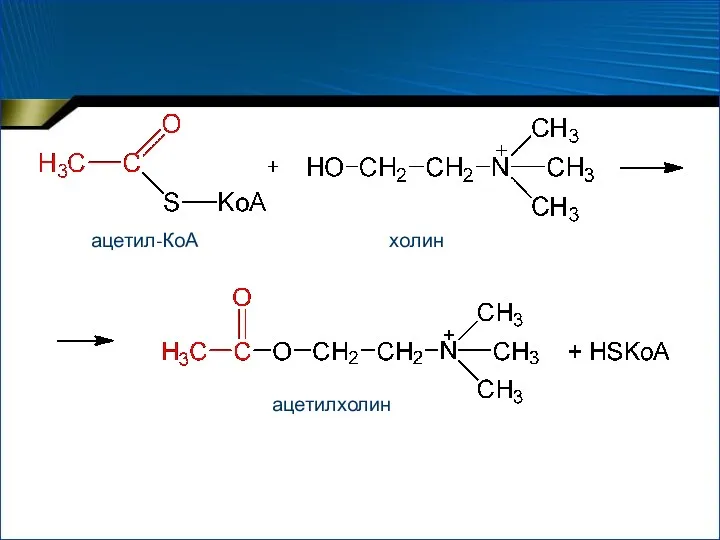
ацетил-КоА
холин
ацетилхолин
ацетил-КоА
холин
ацетилхолин

Подклассы
ферменты, ускоряющие перенос
одноуглеродных фрагментов
Метилтрансферазы
Оксиметилтрансферазы
Формилтрансферазы и др.
нуклеотидных остатков
Нуклеотидилтрансферазы
Подклассы
ферменты, ускоряющие перенос
одноуглеродных фрагментов
Метилтрансферазы
Оксиметилтрансферазы
Формилтрансферазы и др.
нуклеотидных остатков
Нуклеотидилтрансферазы

Гидролазы
Гидролазы

III. Гидролазы
ускоряют реакции гидролиза органических соединений
Делятся на подклассы по характеру
III. Гидролазы
ускоряют реакции гидролиза органических соединений
Делятся на подклассы по характеру

Подклассы
Эстеразы ускоряют гидролиз сложных эфиров
Липаза ускоряет гидролиз α-сложноэфирных связей
Подклассы
Эстеразы ускоряют гидролиз сложных эфиров
Липаза ускоряет гидролиз α-сложноэфирных связей

Подклассы
Гликозидазы ускоряют гидролиз гликозидов
Мальтаза
Сахараза
Амилаза
Подклассы
Гликозидазы ускоряют гидролиз гликозидов
Мальтаза
Сахараза
Амилаза

Подклассы
Пептидазы (пептид-гидролазы) ускоряют гидролиз пептидных связей в белках и пептидах
Подклассы
Пептидазы (пептид-гидролазы) ускоряют гидролиз пептидных связей в белках и пептидах

Подклассы
Амидазы ускоряют гидролиз амидов кислот
уреаза (гидролиз мочевины до NH3
Подклассы
Амидазы ускоряют гидролиз амидов кислот
уреаза (гидролиз мочевины до NH3

Лиазы
Лиазы

IV. Лиазы
ускоряют реакции негидролитического распада органических соединений
При этом замыкаются двойные
IV. Лиазы
ускоряют реакции негидролитического распада органических соединений
При этом замыкаются двойные

Подклассы
С–С-лиазы
декарбоксилазы кетокислот (кофермент – витамин В1)
декарбокислазы аминокислот (кофермент –
Подклассы
С–С-лиазы
декарбоксилазы кетокислот (кофермент – витамин В1)
декарбокислазы аминокислот (кофермент –
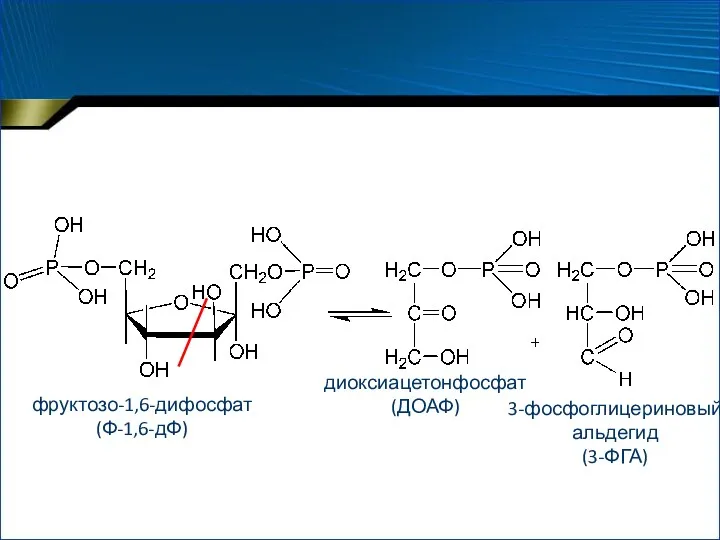
фруктозо-1,6-дифосфат
(Ф-1,6-дФ)
диоксиацетонфосфат
(ДОАФ)
3-фосфоглицериновый альдегид
(3-ФГА)
фруктозо-1,6-дифосфат
(Ф-1,6-дФ)
диоксиацетонфосфат
(ДОАФ)
3-фосфоглицериновый альдегид
(3-ФГА)

Подклассы
С–О-лиазы (гидратазы) ускоряют гидратацию и дегидратацию органических соединений
Фумаратгидратаза
НООС–СН(ОН)–СН2–СООН ⇄
Подклассы
С–О-лиазы (гидратазы) ускоряют гидратацию и дегидратацию органических соединений
Фумаратгидратаза
НООС–СН(ОН)–СН2–СООН ⇄

Подклассы
С–N-лиазы
аспартат-аммиак-лиаза ускоряет прямое дезаминирование асп
НООС–СН(NH2)–СН2–СООН ⇄ НООС–СН=СН–СООNH4
Аспарагиновая кислота
Подклассы
С–N-лиазы
аспартат-аммиак-лиаза ускоряет прямое дезаминирование асп
НООС–СН(NH2)–СН2–СООН ⇄ НООС–СН=СН–СООNH4
Аспарагиновая кислота

Изомеразы
Изомеразы

V. Изомеразы
ускоряют геометрические или структурные изменения в пределах одной молекулы
V. Изомеразы
ускоряют геометрические или структурные изменения в пределах одной молекулы

Подклассы
Рацемазы и эпимеразы
Мутаротаза
α-D-глюкопираноза ⇄ β-D-глюкопираноза
Цис-транс-изомеразы
Ретинол-изомераза
Подклассы
Рацемазы и эпимеразы
Мутаротаза
α-D-глюкопираноза ⇄ β-D-глюкопираноза
Цис-транс-изомеразы
Ретинол-изомераза
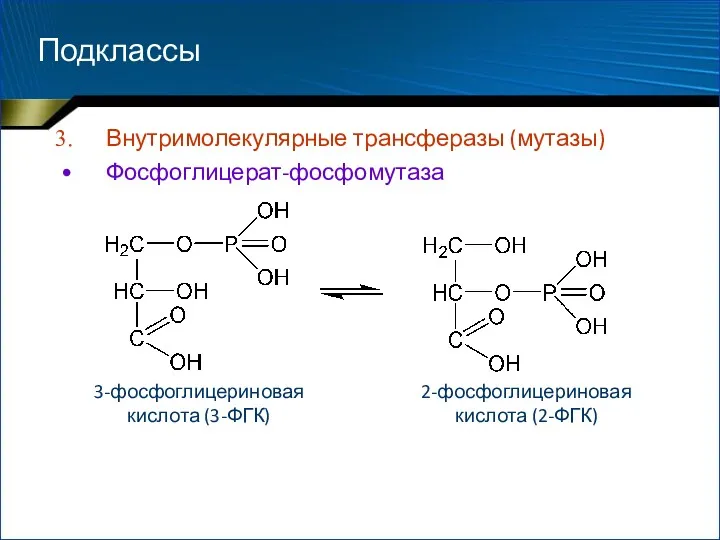
Подклассы
Внутримолекулярные трансферазы (мутазы)
Фосфоглицерат-фосфомутаза
3-фосфоглицериновая кислота (3-ФГК)
2-фосфоглицериновая кислота (2-ФГК)
Подклассы
Внутримолекулярные трансферазы (мутазы)
Фосфоглицерат-фосфомутаза
3-фосфоглицериновая кислота (3-ФГК)
2-фосфоглицериновая кислота (2-ФГК)
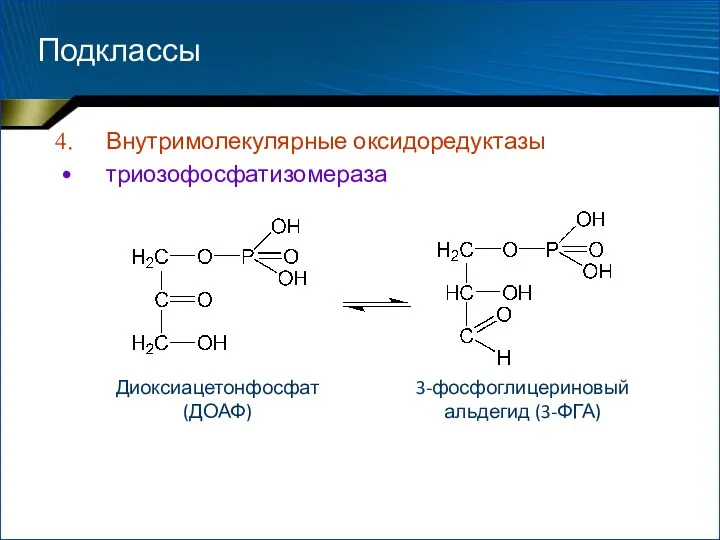
Подклассы
Внутримолекулярные оксидоредуктазы
триозофосфатизомераза
Диоксиацетонфосфат (ДОАФ)
3-фосфоглицериновый альдегид (3-ФГА)
Подклассы
Внутримолекулярные оксидоредуктазы
триозофосфатизомераза
Диоксиацетонфосфат (ДОАФ)
3-фосфоглицериновый альдегид (3-ФГА)

Лигазы
Лигазы

VI. Лигазы
Ускоряют присоединение друг к другу двух молекул
Процесс сопряжен с
VI. Лигазы
Ускоряют присоединение друг к другу двух молекул
Процесс сопряжен с
 Морфология и ультраструктура бактерий
Морфология и ультраструктура бактерий A review of studies on quantum biology
A review of studies on quantum biology Генная инженерия
Генная инженерия Знакомый и незнакомый картофель
Знакомый и незнакомый картофель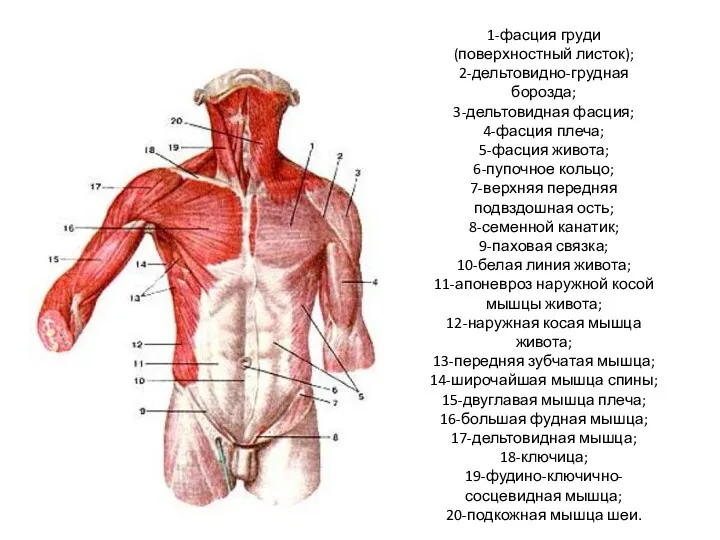 Мышцы туловища
Мышцы туловища Метаболизм белков, пептидов, аминокислот. Лекция 19-21
Метаболизм белков, пептидов, аминокислот. Лекция 19-21 Мастер-класс Я есть то, что я ем
Мастер-класс Я есть то, что я ем Физиология микроорганизмов. Бактерии
Физиология микроорганизмов. Бактерии Естественный отбор и борьба за существование
Естественный отбор и борьба за существование Презентация Печень, строение и функции
Презентация Печень, строение и функции Журавлеподібні
Журавлеподібні Продукты питания, ядовитые для собак и кошек
Продукты питания, ядовитые для собак и кошек Respiration Module
Respiration Module Закономерности развития живой природы
Закономерности развития живой природы Онтогенез речи
Онтогенез речи Постройка и реальность (моделирование форм подводного мира)
Постройка и реальность (моделирование форм подводного мира) Конспект урока и презентация Внешнее строение листа
Конспект урока и презентация Внешнее строение листа Обмен липидов. (Часть 2)
Обмен липидов. (Часть 2) Мутационная изменчивость
Мутационная изменчивость Физиология дыхательной системы. Внешнее дыхание
Физиология дыхательной системы. Внешнее дыхание Як тварини визначають напрямок руху
Як тварини визначають напрямок руху Биотехнология. Достижения и перспективы развития
Биотехнология. Достижения и перспективы развития Деление царств на группы
Деление царств на группы История развития генетики. Основные генетические понятия
История развития генетики. Основные генетические понятия Синтез белков в клетке
Синтез белков в клетке Биохимия липидов. Классификация, биологические функции. Переваривание и всасывание. Обмен липопротеидов
Биохимия липидов. Классификация, биологические функции. Переваривание и всасывание. Обмен липопротеидов Наука генетика. Генетика пола и наследование, сцепленное с полом
Наука генетика. Генетика пола и наследование, сцепленное с полом Водоросли. Общая характеристика. Классификация
Водоросли. Общая характеристика. Классификация