- Физиология микроорганизмов

Содержание
- 2. Энергия в бактериальной клетке накапливается в форме молекул АТФ. У хемоорганотрофных бактерий реакции, связанные с получением
- 3. При использовании в качестве источника углерода и энергии глюкозы или других гексоз начальные этапы окисления глюкозы
- 4. Пути расщепления глюкозы. Расщепление глюкозы до пировиноградной кислоты, одному из важнейших промежуточных продуктов обмена веществ, у
- 5. Пути расщепления глюкозы 1) через образование фруктозо-1,6-дифосфата (ФДФ-путем, или гликолитическим распадом, или, по имени изучавших его
- 6. Глюкоза в бактериальной клетке сначала фосфорилируется при участии АТФ и фермента гексокиназы до метаболически активной формы
- 7. ФДФ-путь. Г-6-Ф изомеризуется до фруктозо-6-фосфата, который под действием фосфофруктокиназы превращается во фруктозо-1,6-дифосфат, который в дальнейшем через
- 8. ПФ-путь. В этом случае глюкозо-6-фосфат через реакции дегидрирования и декарбоксилирования превращается в рибулезо-5-фосфат (Ри-5-Ф), который находится
- 9. Образовавшиеся пентозофосфаты превращаются в результате транскетолазных и трансальдолазных реакций во фруктозо-6-фосфат, замыкая реакции в цикл, и
- 10. КДФГ-путь (путь Этнера—Дудорова) Этот путь расщепления глюкозы специфичен только для бактерий. Встречается у бактерий, потерявших фермент
- 11. Процесс начинается с дегидрирования глюкозо-6-фосфата до 6-фосфоглюконовой кислоты. От нее под действием дегидрогеназы отщепляется вода и
- 12. На каждую молекулу глюкозы образуется 1 молекула АТФ, 1 молекула восстановленного НАД и 1 молекула восстановленного
- 13. Окислительный метаболизм у бактерий (дыхание)
- 14. Окислительный метаболизм Бактерии, обладающие окислительным метаболизмом, энергию получают путем дыхания. Дыхание— процесс получения энергии в реакциях
- 15. В зависимости от акцепторов протонов и электронов среди бактерий различают аэробы, факультативные анаэробы и облигатные анаэробы.
- 16. Облигатные аэробы (бруцеллы, легионеллы, псевдомонады, микобактерии, возбудитель сибирской язвы) растут и размножаются только в присутствии кислорода.
- 17. Облигатные анаэробы (бифидобактерии, лактобактерии, клостридии) не используют кислород для получения энергии. Тип метаболизма у них бродильный.
- 18. Факультативные анаэробы (пневмококки, энтерококки, энтеробактерии, коринебактерии, франциселлы) способны расти и размножаться как в присутствии кислорода, так
- 19. В окислительных процессах, протекающих в атмосфере кислорода образуются токсические продукты: перекись водорода Н2О2 и закисный радикал
- 20. У бактерий, обладающих окислительным метаболизмом, акцептором электронов (или водорода (Н+)) является молекулярный кислород. В этом случае
- 21. Цикл трикарбоновых кислот (цикл Кребса)
- 22. Цикл трикарбоновых кислот выполняет функции как поставщика предшественников для биосинтетических процессов, так и атомов водорода, который
- 23. Типичная цепь выглядит следующим образом: ЦТК → НАД(Н2)→флавопротеид→хинон → →цитохромы: в→с→а→О2
- 24. Среди бактериальных цитохромов различают цитохромы в, с, а и а3. Конечным этапом переноса электронов (протонов) по
- 25. Образование АТФ вдыхательной цепи связывают с хемоосмотическим процессом. Особая ориентация переносчиков в ЦПМ приводит к тому,
- 26. У некоторых бактерий цитохромы отсутствуют, и при контакте с кислородом происходит непосредственный перенос водорода на кислород
- 27. Помимо углеводов прокариоты способны использовать другие органические соединения, в частности белки, в качестве источника энергии, окисляя
- 29. Скачать презентацию

Энергия в бактериальной клетке накапливается в форме молекул АТФ. У хемоорганотрофных
Энергия в бактериальной клетке накапливается в форме молекул АТФ. У хемоорганотрофных

При использовании в качестве источника углерода и энергии глюкозы или других
При использовании в качестве источника углерода и энергии глюкозы или других
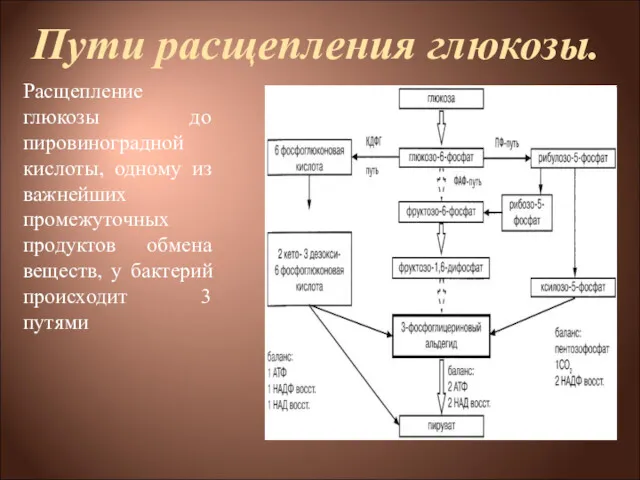
Пути расщепления глюкозы.
Расщепление глюкозы до пировиноградной кислоты, одному из важнейших
Пути расщепления глюкозы.
Расщепление глюкозы до пировиноградной кислоты, одному из важнейших

Пути расщепления глюкозы
1) через образование фруктозо-1,6-дифосфата (ФДФ-путем, или гликолитическим распадом, или,
Пути расщепления глюкозы
1) через образование фруктозо-1,6-дифосфата (ФДФ-путем, или гликолитическим распадом, или,

Глюкоза в бактериальной клетке сначала фосфорилируется при участии АТФ и фермента
Глюкоза в бактериальной клетке сначала фосфорилируется при участии АТФ и фермента

ФДФ-путь.
Г-6-Ф изомеризуется до фруктозо-6-фосфата, который под действием фосфофруктокиназы превращается во
ФДФ-путь.
Г-6-Ф изомеризуется до фруктозо-6-фосфата, который под действием фосфофруктокиназы превращается во

ПФ-путь.
В этом случае глюкозо-6-фосфат через реакции дегидрирования и декарбоксилирования превращается
ПФ-путь.
В этом случае глюкозо-6-фосфат через реакции дегидрирования и декарбоксилирования превращается

Образовавшиеся пентозофосфаты превращаются в результате транскетолазных и трансальдолазных реакций во фруктозо-6-фосфат,
Образовавшиеся пентозофосфаты превращаются в результате транскетолазных и трансальдолазных реакций во фруктозо-6-фосфат,

КДФГ-путь (путь Этнера—Дудорова)
Этот путь расщепления глюкозы специфичен только для бактерий.
КДФГ-путь (путь Этнера—Дудорова)
Этот путь расщепления глюкозы специфичен только для бактерий.

Процесс начинается с дегидрирования глюкозо-6-фосфата до 6-фосфоглюконовой кислоты. От нее под
Процесс начинается с дегидрирования глюкозо-6-фосфата до 6-фосфоглюконовой кислоты. От нее под

На каждую молекулу глюкозы образуется 1 молекула АТФ, 1 молекула восстановленного
На каждую молекулу глюкозы образуется 1 молекула АТФ, 1 молекула восстановленного
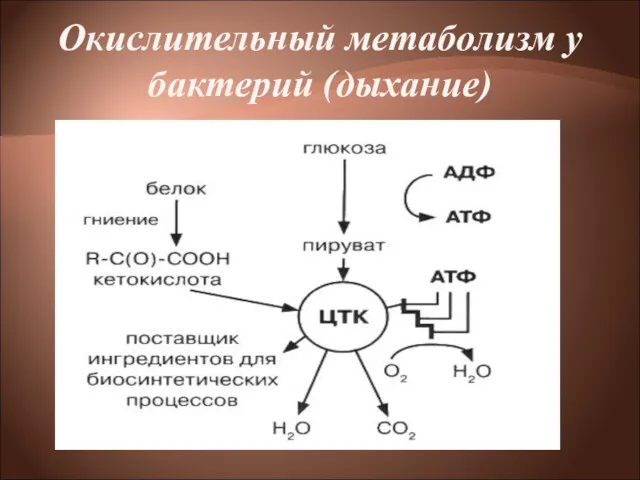
Окислительный метаболизм у бактерий (дыхание)
Окислительный метаболизм у бактерий (дыхание)

Окислительный метаболизм
Бактерии, обладающие окислительным метаболизмом, энергию получают путем дыхания.
Дыхание— процесс получения
Окислительный метаболизм
Бактерии, обладающие окислительным метаболизмом, энергию получают путем дыхания.
Дыхание— процесс получения

В зависимости от акцепторов протонов и электронов среди бактерий различают аэробы,
В зависимости от акцепторов протонов и электронов среди бактерий различают аэробы,

Облигатные аэробы (бруцеллы, легионеллы, псевдомонады, микобактерии, возбудитель сибирской язвы) растут и
Облигатные аэробы (бруцеллы, легионеллы, псевдомонады, микобактерии, возбудитель сибирской язвы) растут и

Облигатные анаэробы (бифидобактерии, лактобактерии, клостридии) не используют кислород для получения энергии.
Облигатные анаэробы (бифидобактерии, лактобактерии, клостридии) не используют кислород для получения энергии.

Факультативные анаэробы (пневмококки, энтерококки, энтеробактерии, коринебактерии, франциселлы) способны расти и размножаться
Факультативные анаэробы (пневмококки, энтерококки, энтеробактерии, коринебактерии, франциселлы) способны расти и размножаться

В окислительных процессах, протекающих в атмосфере кислорода образуются токсические продукты: перекись
В окислительных процессах, протекающих в атмосфере кислорода образуются токсические продукты: перекись

У бактерий, обладающих окислительным метаболизмом, акцептором электронов (или водорода (Н+)) является
У бактерий, обладающих окислительным метаболизмом, акцептором электронов (или водорода (Н+)) является
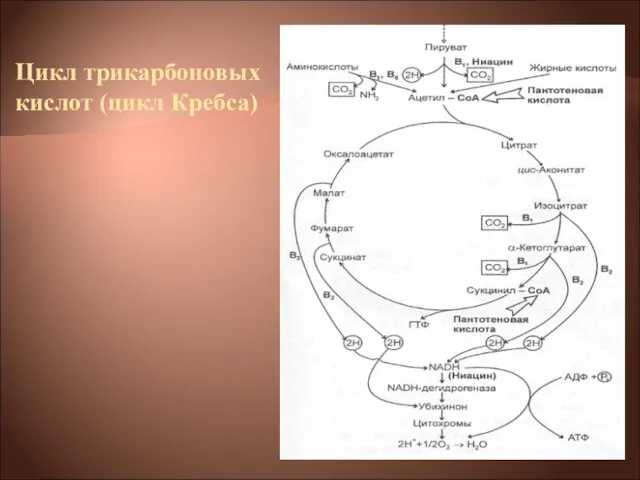
Цикл трикарбоновых кислот (цикл Кребса)
Цикл трикарбоновых кислот (цикл Кребса)

Цикл трикарбоновых кислот выполняет функции как поставщика предшественников для биосинтетических процессов,
Цикл трикарбоновых кислот выполняет функции как поставщика предшественников для биосинтетических процессов,

Типичная цепь выглядит следующим образом:
ЦТК → НАД(Н2)→флавопротеид→хинон → →цитохромы: в→с→а→О2
Типичная цепь выглядит следующим образом:
ЦТК → НАД(Н2)→флавопротеид→хинон → →цитохромы: в→с→а→О2

Среди бактериальных цитохромов различают цитохромы в, с, а и а3. Конечным
Среди бактериальных цитохромов различают цитохромы в, с, а и а3. Конечным

Образование АТФ вдыхательной цепи связывают с хемоосмотическим процессом. Особая ориентация переносчиков
Образование АТФ вдыхательной цепи связывают с хемоосмотическим процессом. Особая ориентация переносчиков

У некоторых бактерий цитохромы отсутствуют, и при контакте с кислородом происходит
У некоторых бактерий цитохромы отсутствуют, и при контакте с кислородом происходит

Помимо углеводов прокариоты способны использовать другие органические соединения, в частности белки,
Помимо углеводов прокариоты способны использовать другие органические соединения, в частности белки,
 Болезни и вредители цитрусовых
Болезни и вредители цитрусовых Рыба речная: Окунь
Рыба речная: Окунь Витамины – понятие о гиповитаминозах, гипервитаминозах, авитаминозах
Витамины – понятие о гиповитаминозах, гипервитаминозах, авитаминозах Мышечные цепи. Взаимосвязи миофасциальной сети
Мышечные цепи. Взаимосвязи миофасциальной сети Вода. Роль воды в жизни человека
Вода. Роль воды в жизни человека Строение. Функции. Значение кожи
Строение. Функции. Значение кожи Тип Хордовые класс Млекопитающие
Тип Хордовые класс Млекопитающие Профилактика употреблениия ПАВ через преподавание биологии
Профилактика употреблениия ПАВ через преподавание биологии Простейшие
Простейшие Наружные и внутренние мужские и женские половые органы
Наружные и внутренние мужские и женские половые органы Процессы жизнеобеспечения в организме человека. Эндокринные железы (железы внутренней секреции)
Процессы жизнеобеспечения в организме человека. Эндокринные железы (железы внутренней секреции) Применение проектной технологии на уроках биологии и во внеурочной деятельности
Применение проектной технологии на уроках биологии и во внеурочной деятельности Ядовитые змеи мира
Ядовитые змеи мира Пластичность клеток разных тканей
Пластичность клеток разных тканей الزواحف
الزواحف Домашние животные. Кошки
Домашние животные. Кошки Биоритмы. Характеристика хронологических типов человека
Биоритмы. Характеристика хронологических типов человека Рост и развитие волос
Рост и развитие волос Биотехнология түсініктері, даму тарихы, негізгі әдістері
Биотехнология түсініктері, даму тарихы, негізгі әдістері Суринам ұн жемірі
Суринам ұн жемірі Використання тваринами знарядь праці
Використання тваринами знарядь праці Гүл, оның құрылысы мен маңызы
Гүл, оның құрылысы мен маңызы Морские животные. Тип мягкотелые
Морские животные. Тип мягкотелые Экология и природопользование. Экосистемы
Экология и природопользование. Экосистемы Передвижение веществ в организме растения
Передвижение веществ в организме растения Транспорт веществ в организме. 6 класс
Транспорт веществ в организме. 6 класс Класс Пресмыкающиеся, или Рептилии
Класс Пресмыкающиеся, или Рептилии Методы иммуноанализа с применением различных меток
Методы иммуноанализа с применением различных меток