- Физиология микроорганизмов

Содержание
- 2. Энергия в бактериальной клетке накапливается в форме молекул АТФ. У хемоорганотрофных бактерий реакции, связанные с получением
- 3. При использовании в качестве источника углерода и энергии глюкозы или других гексоз начальные этапы окисления глюкозы
- 4. Расщепление глюкозы до пировиноградной кислоты, одному из важнейших промежуточных продуктов обмена веществ, у бактерий происходит 3
- 6. Пути расщепления глюкозы 1) через образование фруктозо-1,6-дифосфата (ФДФ-путем, или гликолитическим распадом, или, по имени изучавших его
- 7. Глюкоза в бактериальной клетке сначала фосфорилируется при участии АТФ и фермента гексокиназы до метаболически активной формы
- 8. ФДФ-путь. Г-6-Ф изомеризуется до фруктозо-6-фосфата, который под действием фосфофруктокиназы превращается во фруктозо-1,6-дифосфат, который в дальнейшем через
- 9. ПФ-путь. В этом случае глюкозо-6-фосфат через реакции дегидрирования и декарбоксилирования превращается в рибулезо-5-фосфат (Ри-5-Ф), который находится
- 10. Образовавшиеся пентозофосфаты превращаются в результате транскетолазных и трансальдолазных реакций во фруктозо-6-фосфат, замыкая реакции в цикл, и
- 11. КДФГ-путь (путь Этнера—Дудорова) Этот путь расщепления глюкозы специфичен только для бактерий. Встречается у бактерий, потерявших фермент
- 12. бактерии рода Pseudomonas
- 13. Процесс начинается с дегидрирования глюкозо-6-фосфата до 6-фосфоглюконовой кислоты. От нее под действием дегидрогеназы отщепляется вода и
- 14. На каждую молекулу глюкозы образуется: молекула АТФ, 1 молекула восстановленного НАД 1 молекула восстановленного НАДФ, которая
- 16. Окислительный метаболизм Бактерии, обладающие окислительным метаболизмом, энергию получают путем дыхания. Дыхание — процесс получения энергии в
- 17. В зависимости от акцепторов протонов и электронов среди бактерий различают аэробы, факультативные анаэробы и облигатные анаэробы.
- 18. Облигатные аэробы (бруцеллы, легионеллы, псевдомонады, микобактерии, возбудитель сибирской язвы) растут и размножаются только в присутствии кислорода.
- 19. Облигатные анаэробы (бифидобактерии, лактобактерии, клостридии) не используют кислород для получения энергии. Тип метаболизма у них бродильный.
- 20. Факультативные анаэробы (пневмококки, энтерококки, энтеробактерии, коринебактерии, франциселлы) способны расти и размножаться как в присутствии кислорода, так
- 21. В окислительных процессах, протекающих в атмосфере кислорода образуются токсические продукты: перекись водорода Н2О2 и закисный радикал
- 22. У бактерий, обладающих окислительным метаболизмом, акцептором электронов (или водорода (Н+) является молекулярный кислород. В этом случае
- 23. цикл кребса
- 24. Цикл трикарбоновых кислот (цикл Кребса)
- 25. Цикл трикарбоновых кислот выполняет функции как поставщика предшественников для биосинтетических процессов, так и атомов водорода, который
- 26. Типичная цепь выглядит следующим образом: ЦТК → НАД(Н2)→флавопротеид→хинон → →цитохромы: в→с→а→О2 Среди бактериальных цитохромов различают цитохромы
- 27. Среди бактериальных цитохромов различают цитохромы в, с, а и а3. Конечным этапом переноса электронов (протонов) по
- 28. Образование АТФ в дыхательной цепи связывают с хемоосмотическим процессом. Особая ориентация переносчиков в ЦПМ приводит к
- 29. У некоторых бактерий цитохромы отсутствуют, и при контакте с кислородом происходит непосредственный перенос водорода на кислород
- 31. Скачать презентацию

Энергия в бактериальной клетке накапливается в форме молекул АТФ.
У хемоорганотрофных
Энергия в бактериальной клетке накапливается в форме молекул АТФ.
У хемоорганотрофных

При использовании в качестве источника углерода и энергии глюкозы или других
При использовании в качестве источника углерода и энергии глюкозы или других

Расщепление глюкозы до пировиноградной кислоты, одному из важнейших промежуточных продуктов обмена
Расщепление глюкозы до пировиноградной кислоты, одному из важнейших промежуточных продуктов обмена
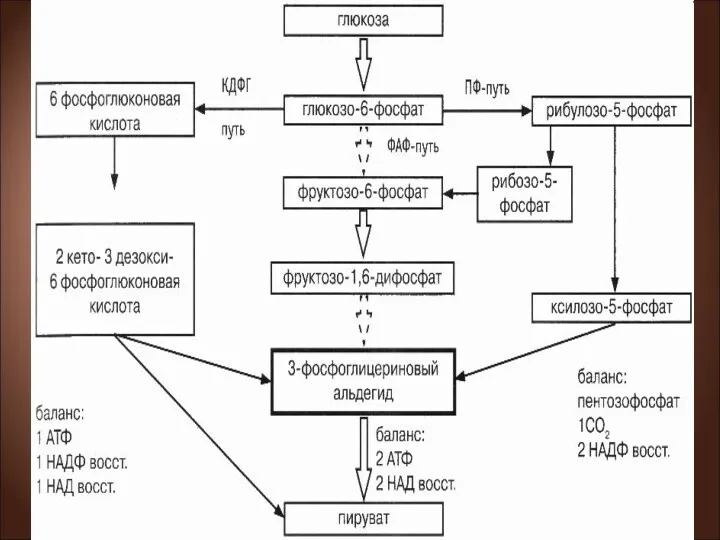

Пути расщепления глюкозы
1) через образование фруктозо-1,6-дифосфата (ФДФ-путем, или гликолитическим распадом, или,
Пути расщепления глюкозы
1) через образование фруктозо-1,6-дифосфата (ФДФ-путем, или гликолитическим распадом, или,

Глюкоза в бактериальной клетке сначала фосфорилируется при участии АТФ и фермента
Глюкоза в бактериальной клетке сначала фосфорилируется при участии АТФ и фермента

ФДФ-путь.
Г-6-Ф изомеризуется до фруктозо-6-фосфата, который под действием фосфофруктокиназы превращается во
ФДФ-путь.
Г-6-Ф изомеризуется до фруктозо-6-фосфата, который под действием фосфофруктокиназы превращается во

ПФ-путь.
В этом случае глюкозо-6-фосфат через реакции дегидрирования и декарбоксилирования превращается
ПФ-путь.
В этом случае глюкозо-6-фосфат через реакции дегидрирования и декарбоксилирования превращается

Образовавшиеся пентозофосфаты превращаются в результате транскетолазных и трансальдолазных реакций во фруктозо-6-фосфат,
Образовавшиеся пентозофосфаты превращаются в результате транскетолазных и трансальдолазных реакций во фруктозо-6-фосфат,

КДФГ-путь (путь Этнера—Дудорова)
Этот путь расщепления глюкозы специфичен только для бактерий.
КДФГ-путь (путь Этнера—Дудорова)
Этот путь расщепления глюкозы специфичен только для бактерий.
бактерии рода Pseudomonas
бактерии рода Pseudomonas

Процесс начинается с дегидрирования глюкозо-6-фосфата до 6-фосфоглюконовой кислоты. От нее под
Процесс начинается с дегидрирования глюкозо-6-фосфата до 6-фосфоглюконовой кислоты. От нее под

На каждую молекулу глюкозы образуется:
молекула АТФ,
1 молекула восстановленного НАД
На каждую молекулу глюкозы образуется:
молекула АТФ,
1 молекула восстановленного НАД
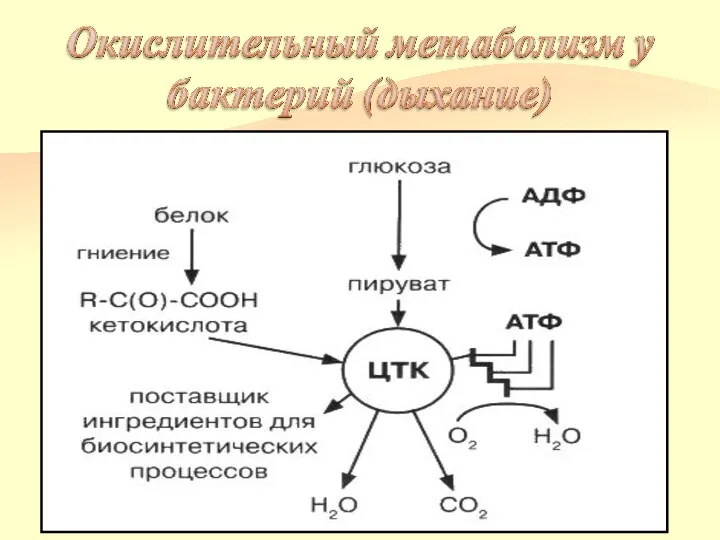

Окислительный метаболизм
Бактерии, обладающие окислительным метаболизмом, энергию получают путем дыхания.
Дыхание — процесс
Окислительный метаболизм
Бактерии, обладающие окислительным метаболизмом, энергию получают путем дыхания.
Дыхание — процесс

В зависимости от акцепторов протонов и электронов среди бактерий различают аэробы,
В зависимости от акцепторов протонов и электронов среди бактерий различают аэробы,

Облигатные аэробы (бруцеллы, легионеллы, псевдомонады, микобактерии, возбудитель сибирской язвы) растут и
Облигатные аэробы (бруцеллы, легионеллы, псевдомонады, микобактерии, возбудитель сибирской язвы) растут и

Облигатные анаэробы (бифидобактерии, лактобактерии, клостридии) не используют кислород для получения энергии.
Облигатные анаэробы (бифидобактерии, лактобактерии, клостридии) не используют кислород для получения энергии.

Факультативные анаэробы (пневмококки, энтерококки, энтеробактерии, коринебактерии, франциселлы) способны расти и размножаться
Факультативные анаэробы (пневмококки, энтерококки, энтеробактерии, коринебактерии, франциселлы) способны расти и размножаться

В окислительных процессах, протекающих в атмосфере кислорода образуются токсические продукты: перекись
В окислительных процессах, протекающих в атмосфере кислорода образуются токсические продукты: перекись

У бактерий, обладающих окислительным метаболизмом, акцептором электронов (или водорода (Н+) является
У бактерий, обладающих окислительным метаболизмом, акцептором электронов (или водорода (Н+) является
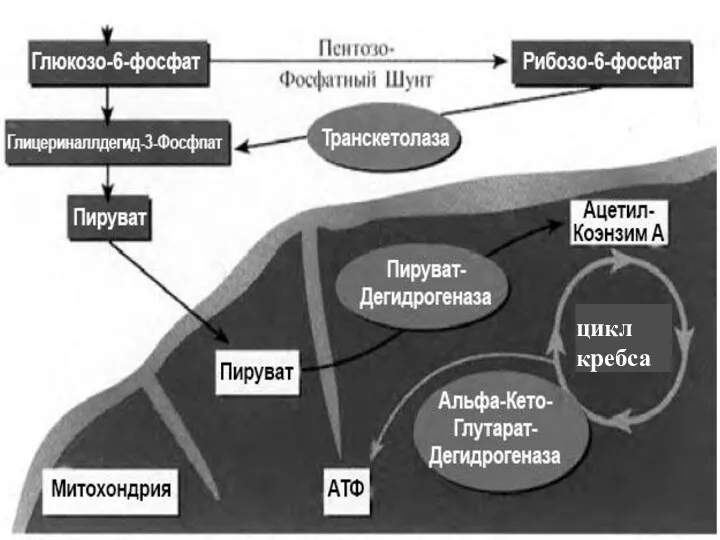
цикл
кребса
цикл
кребса
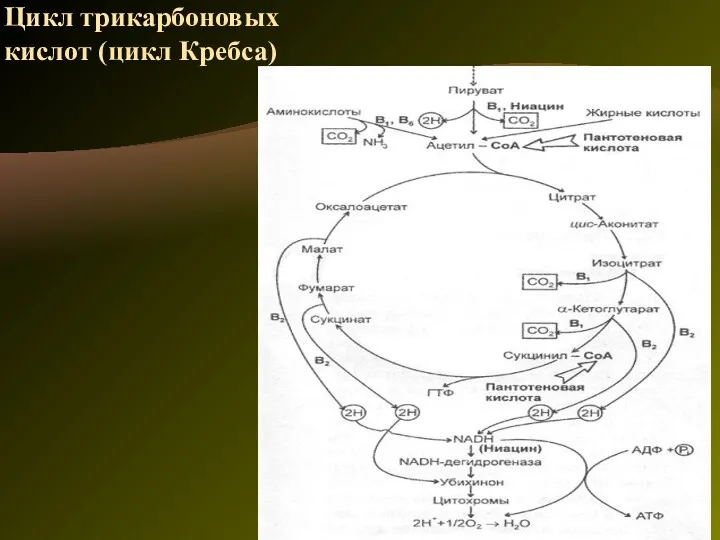
Цикл трикарбоновых кислот (цикл Кребса)
Цикл трикарбоновых кислот (цикл Кребса)
Цикл трикарбоновых кислот выполняет функции как поставщика предшественников для биосинтетических процессов,
Цикл трикарбоновых кислот выполняет функции как поставщика предшественников для биосинтетических процессов,

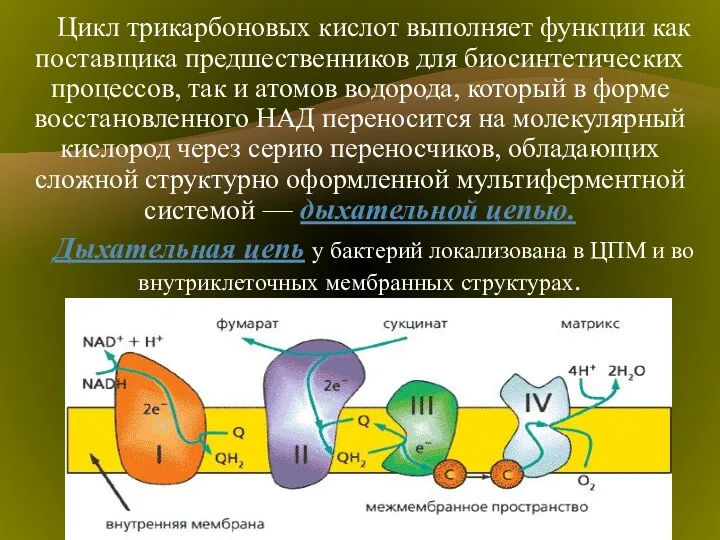
Типичная цепь выглядит следующим образом:
ЦТК → НАД(Н2)→флавопротеид→хинон → →цитохромы: в→с→а→О2
Среди
Типичная цепь выглядит следующим образом:
ЦТК → НАД(Н2)→флавопротеид→хинон → →цитохромы: в→с→а→О2
Среди

Среди бактериальных цитохромов различают цитохромы в, с, а и а3. Конечным
Среди бактериальных цитохромов различают цитохромы в, с, а и а3. Конечным
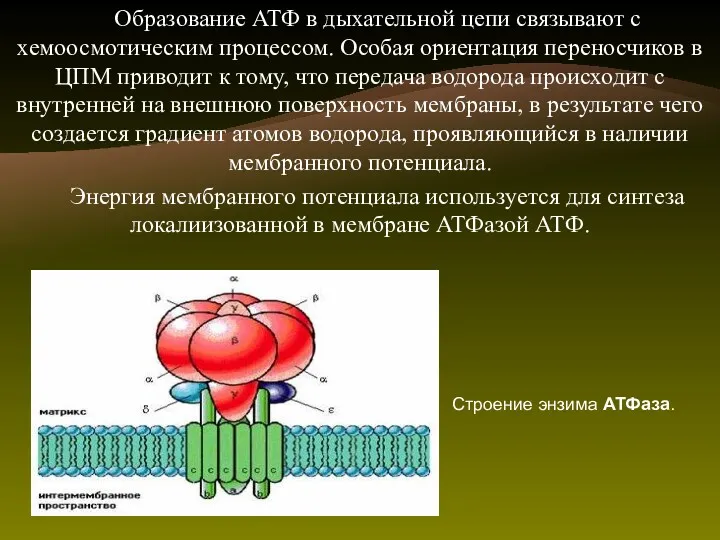
Образование АТФ в дыхательной цепи связывают с хемоосмотическим процессом. Особая ориентация
Образование АТФ в дыхательной цепи связывают с хемоосмотическим процессом. Особая ориентация

У некоторых бактерий цитохромы отсутствуют, и при контакте с кислородом происходит
У некоторых бактерий цитохромы отсутствуют, и при контакте с кислородом происходит
 Обмен веществ и превращение энергии в клетке
Обмен веществ и превращение энергии в клетке Определение чувствительности к антибиотикам. Культивирование анаэробов. Диагностика с помощью бактериофагов
Определение чувствительности к антибиотикам. Культивирование анаэробов. Диагностика с помощью бактериофагов Птицы Адамовского района
Птицы Адамовского района Нуклеиновые кислоты
Нуклеиновые кислоты Як вчаться пташенята
Як вчаться пташенята Презентация по теме Основные типы экологических взаимодействий
Презентация по теме Основные типы экологических взаимодействий Биологическое действие радиации. Радиационная безопасность
Биологическое действие радиации. Радиационная безопасность изменчивость
изменчивость Лишайники. Организм- сфинкс. 5 класс (1)
Лишайники. Организм- сфинкс. 5 класс (1) Строение и работа сердца. 8 класс
Строение и работа сердца. 8 класс Нервная система
Нервная система Обмін речовин та перетворення енергії в клітині. Електронний журнал
Обмін речовин та перетворення енергії в клітині. Електронний журнал Популяция. Функция популяции как системы
Популяция. Функция популяции как системы Основные вегетативные органы растений. Лекция 6
Основные вегетативные органы растений. Лекция 6 Дәнді дақылдардың вирустық аурулары
Дәнді дақылдардың вирустық аурулары Клонирование и очистка рекомбинантного белка
Клонирование и очистка рекомбинантного белка Коала. Образ жизни и питания
Коала. Образ жизни и питания Семейство Enterobacteriaceae
Семейство Enterobacteriaceae Интеллектуальная игра Аист над Амуром
Интеллектуальная игра Аист над Амуром Физиология, биохимия микроорганизмов, прокариотов-бактерий, эукариотов-простейших, грибов
Физиология, биохимия микроорганизмов, прокариотов-бактерий, эукариотов-простейших, грибов Генетичні основи селекції рослин
Генетичні основи селекції рослин Обрезка плодовых деревьев и кустарников
Обрезка плодовых деревьев и кустарников Средства борьбы с вредителями растений. Инсектициды и акарициды. (Лекция 6)
Средства борьбы с вредителями растений. Инсектициды и акарициды. (Лекция 6) Древнейшие люди
Древнейшие люди Катонқарағай ауданы
Катонқарағай ауданы Анатомо-физиологические особенности системы органов дыхания
Анатомо-физиологические особенности системы органов дыхания Окружающий мир. Мир растений. (3 класс)
Окружающий мир. Мир растений. (3 класс) Методика подготовки к ЕГЭ по биологии
Методика подготовки к ЕГЭ по биологии