- Физиология мышц

Содержание
- 2. Скелетные мышцы подразделяются на белые и красные: первые по функции являются быстро сокращающимися мышцами, а вторые
- 3. СЕРДЕЧНАЯ МЫШЦА подразделяется на сократительные клетки, которые образуют предсердия и желудочки (миокард), и клетки, которые не
- 5. Функции скелетных мышц Передвижение тела в пространстве Перемещение частей тела относительно друг друга Поддержание позы Передвижение
- 6. Функции гладких мышц Обеспечивают функции полых органов, стенки которых они образуют Обеспечивают сфинктерную функцию Поддерживают АД
- 7. СВОЙСТВА СКЕЛЕТНЫХ МЫШЦ 1. ВОЗБУДИМОСТЬ определяют по силе мышечного возбуждения, вызываемого через электроды, наложенные непосредственно на
- 8. 2.РАСТЯЖИМОСТЬ. Это свойство мышцы удлиняться под влиянием нагрузки. Степень растяжения мышцы зависит от величины груза. Растяжимость
- 9. 3. ЭЛАСТИЧНОСТЬ. Это свойство деформированного тела возвращаться к первоначальному своему состоянию после удаления силы, вызвавшей деформацию.
- 10. Строение мышечного волокна (I) и миофибриллы (II): А — анизотропные диски, I — изотропные диски, Н
- 11. Миофибриллы с саркоплазматическим ретикулумом (по Дж. Бендоллу, 1970): I — саркомеры, II — саркомеры с удаленным
- 12. СТРОЕНИЕ СКЕЛЕТНОЙ МЫШЦЫ Скелетная мышца представляет собой комплекс отдельных многоядерных мышечных клеток или волокон, которые состоят
- 13. Кроме того, в состав нитей актина входят белки — тропонин и тропомиозин, которые необходимы для развития
- 14. Поэтому каждый глобулярный G-актин представляет собой место связывания с миозином при сокращении мышцы. В состоянии покоя
- 15. СХЕМАТИЧЕСКОЕ ИЗОБРАЖЕНИЕ ПРОСТРАНСТВЕННОГО РАСПОЛОЖЕНИЯ АКТИНА, МИОЗИНА, ТРОПОНИНА, ТРОПОМИОЗИНА (РАССЛАБЛЕНИЕ)
- 16. ПОСЛЕ СВЯЗЫВАНИЯ ДВУХ ИОНОВ КАЛЬЦИЯ
- 17. Толстый филамент представляет собой биполярный полимер белка миозина, который способен взаимодействовать с актином и развивать в
- 18. В скелетных мышцах от миозинового стержня по его окружности отходят с равным интервалом в 14,3 нм
- 19. МЕХАНИЗМ ОДИНОЧНОГО МЫШЕЧНОГО СОКРАЩЕНИЯ 1.Процесс, при котором потенциалы действия, генерируемые на сарколемме в области нервно-мышечного синапса,
- 20. В состоянии покоя поперечный мостик расположен параллельно миозиновому стержию , а его головка — перпедикулярно относительно
- 21. 2.Для того чтобы вызвать сокращение, потенциал действия мышцы, вызваемый двигательным нервом, распространяется по сарколемме мышечной клетки
- 22. 3.Когда концентрация ионов Са в межфибриллярном пространстве, т. е. в саркоалазме между нитями актина и миозина,
- 23. После того, как ионы Са2 высвобождаются из саркоплазматического ретикулума и активируют сокращение мышцы, активновый филамент может
- 24. 4.Энергия гидролиза АТФ вызывает движение головки поперечного мостика, она наклоняется, что является основной силой поперечного мостика,
- 25. В результате головка мостика совершает движение на 40° к центру саркомера и перемещает при этом на
- 26. В результате одиночного мышечного сокращения : Актиновые и миозиновые нити не укорачиваются Взаимодействие актина и миозина
- 27. Расслабление скелетной мышцы Расслабление скелетной мышцы начинается только после того, как концентрация ионов Са в саркоплазме
- 28. ТИПЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ Сокращение мышцы в целом является результатом суммации одиночных мышечных сокращений. Сократительный ответ мышечного
- 30. Сократимость и возбудимость мышц разного вида: 1 — кривая сокращения, 2 — кривая возбудимости; а —
- 31. Тетаническим называется сокращение мышцы, при котором происходит суммация одиночных мышечных сокращений в результате ее раздражения частыми
- 32. Гладкий тетанус возникает в том случае, если нерв или мьшцу раздражать серией электрических импульсов с частотой,
- 33. РЕЖИМЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ
- 34. Если мышца развивает напряжение в условиях чрезмерной нагрузки, то ее длина не изменяется и такое сокращение
- 35. Во время перемещения тела в пространстве имеет место смешанный, или ауксотонический, или анизотонический тип сокращения, при
- 36. ЭНЕРГЕТИКА МЫШЕЧНОГО СОКРАЩЕНИЯ АТФ в мышце необходима для: Сокращения (образования мостиков) Расслабления (разрыва мостиков) Работы Са-насоса
- 37. МЕХАНИЗМЫ ВОССТАНОВЛЕНИЯ АТФ Креатинфосфокиназный. 3,6 моль АТФ/мин Гликолитический. 1,2 моль АТФ/мин Окислительный. - при окислении глюкозы
- 38. ПЕРЕДАЧА НА РАБОЧИЙ МЕХАНИЗМ МЫШЕЧНОГО ВОЛОКНА ЭНЕРГИИ, ОСВОБОЖДАЮЩЕЙСЯ ПРИ ОКИСЛИТЕЛЬНЫХ ПРОЦЕССАХ
- 39. ДВИГАТЕЛЬНАЯ ЕДИНИЦА
- 40. Единичное двигательное волокно мотонейрона и иннервируемые им мышечные волокна – ДВИГАТЕЛЬНАЯ ЕДИНИЦА (ДЕ). В большинстве скелетных
- 41. ОТЛИЧИЯ В НЕРВНЫХ КОМПОНЕНТАХ Возбудимость мотонейронов одной и той же мышцы находится в обратной зависимости от
- 42. ОТЛИЧИЯ МЫШЕЧНЫХ ВОЛОКОН ДЕ В быстрых мышечных волокнах : Больше ёмкость саркоплазматического ретикулума При поступлении ПД
- 43. Внутри этих волокон содержится большое количество миоглобина (придающего им красный цвет), являющегося депо кислорода в момент,
- 44. Физиологические показатели сокращения скелетной мышцы Сила Скорость развития силы и укорочения мышцы Мощность Работа
- 45. Скорость укорочения скелетной мышцы находится в обратной зависимости от величины нагрузки. Эта взаимосвязь описывается классическим уравнением
- 46. Работа мышц равна произведению силы, развиваемой мышцей, на путь перемещения груза, либо механическая работа мышц равна
- 47. Оптимальная скорость укорочения скелетной мышцы зависит от оптимальной исходной длины саркомеров перед началом сокращения, которая в
- 48. Мощность мышечной работы как физиологический параметр сокращения мышцы измеряется произведением величины силы мышцы на скорость укорочения
- 49. Утомление скелетной мышцы Утомлением называется временное снижение способности мышцы развивать усилие в результате предшествующей физической активности.
- 50. Унитарная гладкая мышца называется висцеральной, так как она окружает полые органы — желудок, кишечник, мочевой пузырь
- 51. ВЗАИМОДЕЙСТВИЕ ГЛАДКОМЫШЕЧНЫХ КЛЕТОК УНИТАРНОЙ МЫШЦЫ
- 52. Мультиунитарная гладкая мышца образована отдельными гладкомышечными клетками, каждая из которых иннервирована отдельным нервным волокном вегетативной нервной
- 54. Скачать презентацию

Скелетные мышцы подразделяются на белые и красные: первые по функции являются
Скелетные мышцы подразделяются на белые и красные: первые по функции являются

СЕРДЕЧНАЯ МЫШЦА подразделяется на сократительные клетки, которые образуют предсердия и желудочки
СЕРДЕЧНАЯ МЫШЦА подразделяется на сократительные клетки, которые образуют предсердия и желудочки


Функции скелетных мышц
Передвижение тела в пространстве
Перемещение частей тела относительно друг друга
Поддержание
Функции скелетных мышц
Передвижение тела в пространстве
Перемещение частей тела относительно друг друга
Поддержание

Функции гладких мышц
Обеспечивают функции полых органов, стенки которых они образуют
Обеспечивают сфинктерную
Функции гладких мышц
Обеспечивают функции полых органов, стенки которых они образуют
Обеспечивают сфинктерную

СВОЙСТВА СКЕЛЕТНЫХ МЫШЦ
1. ВОЗБУДИМОСТЬ определяют по силе мышечного возбуждения, вызываемого через
СВОЙСТВА СКЕЛЕТНЫХ МЫШЦ
1. ВОЗБУДИМОСТЬ определяют по силе мышечного возбуждения, вызываемого через

2.РАСТЯЖИМОСТЬ. Это свойство мышцы удлиняться под влиянием нагрузки. Степень растяжения
2.РАСТЯЖИМОСТЬ. Это свойство мышцы удлиняться под влиянием нагрузки. Степень растяжения

3. ЭЛАСТИЧНОСТЬ. Это свойство деформированного тела возвращаться к первоначальному своему состоянию
3. ЭЛАСТИЧНОСТЬ. Это свойство деформированного тела возвращаться к первоначальному своему состоянию
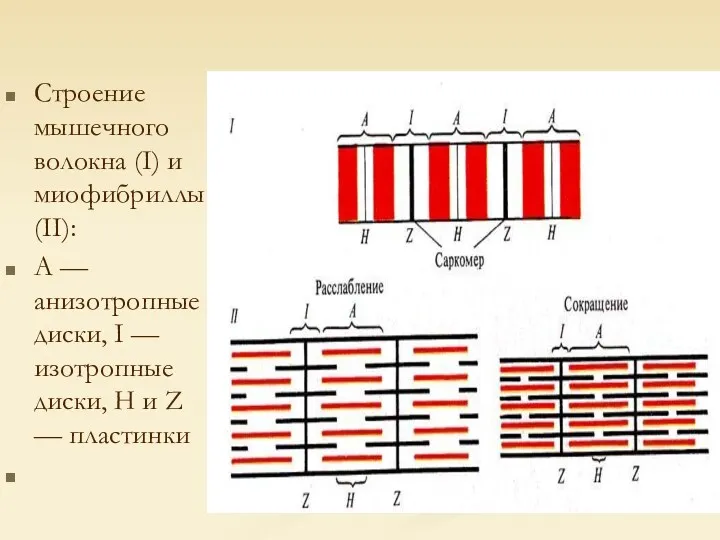
Строение мышечного волокна (I) и миофибриллы (II):
А — анизотропные диски, I
Строение мышечного волокна (I) и миофибриллы (II):
А — анизотропные диски, I
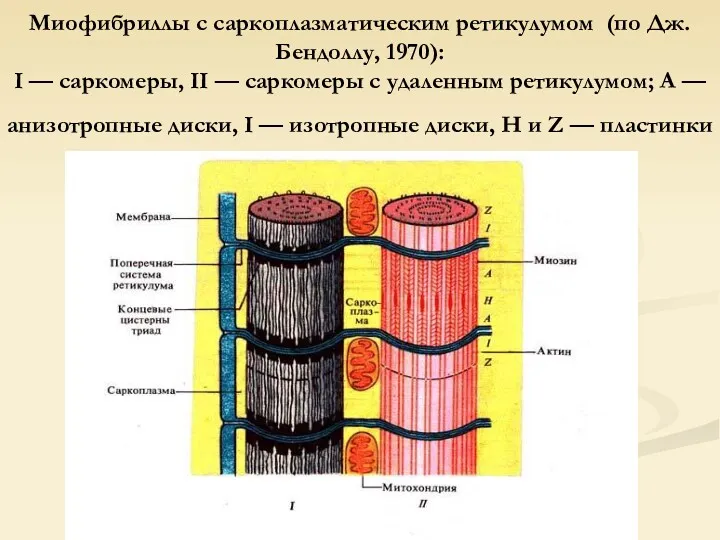
Миофибриллы с саркоплазматическим ретикулумом (по Дж. Бендоллу, 1970):
I — саркомеры, II
Миофибриллы с саркоплазматическим ретикулумом (по Дж. Бендоллу, 1970): I — саркомеры, II

СТРОЕНИЕ СКЕЛЕТНОЙ МЫШЦЫ
Скелетная мышца представляет собой комплекс отдельных многоядерных мышечных клеток
СТРОЕНИЕ СКЕЛЕТНОЙ МЫШЦЫ
Скелетная мышца представляет собой комплекс отдельных многоядерных мышечных клеток

Кроме того, в состав нитей актина входят белки — тропонин и
Кроме того, в состав нитей актина входят белки — тропонин и

Поэтому каждый глобулярный G-актин представляет собой место связывания с миозином при
Поэтому каждый глобулярный G-актин представляет собой место связывания с миозином при
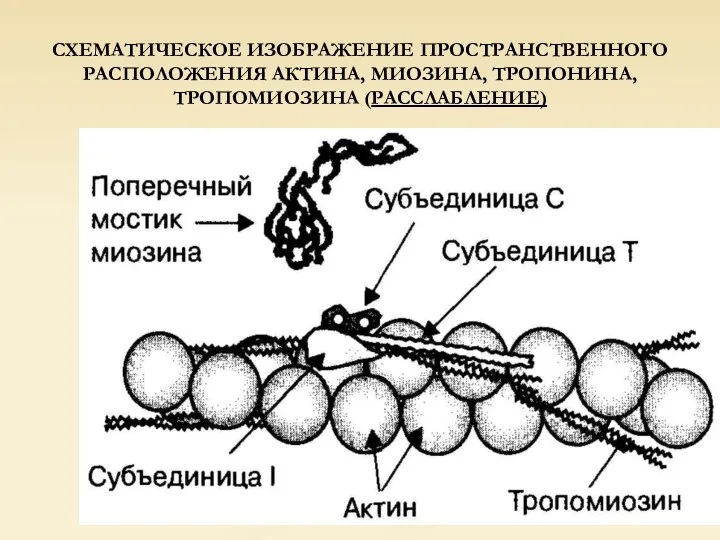
СХЕМАТИЧЕСКОЕ ИЗОБРАЖЕНИЕ ПРОСТРАНСТВЕННОГО РАСПОЛОЖЕНИЯ АКТИНА, МИОЗИНА, ТРОПОНИНА, ТРОПОМИОЗИНА (РАССЛАБЛЕНИЕ)
СХЕМАТИЧЕСКОЕ ИЗОБРАЖЕНИЕ ПРОСТРАНСТВЕННОГО РАСПОЛОЖЕНИЯ АКТИНА, МИОЗИНА, ТРОПОНИНА, ТРОПОМИОЗИНА (РАССЛАБЛЕНИЕ)
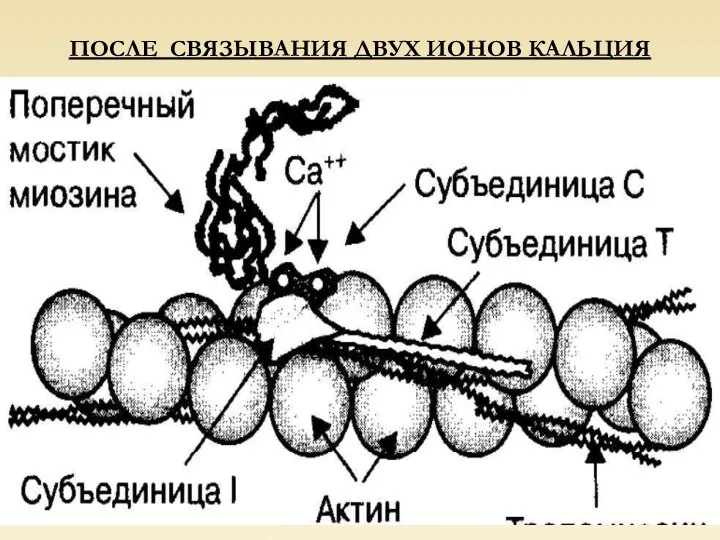
ПОСЛЕ СВЯЗЫВАНИЯ ДВУХ ИОНОВ КАЛЬЦИЯ
ПОСЛЕ СВЯЗЫВАНИЯ ДВУХ ИОНОВ КАЛЬЦИЯ

Толстый филамент представляет собой биполярный полимер белка миозина, который способен взаимодействовать
Толстый филамент представляет собой биполярный полимер белка миозина, который способен взаимодействовать

В скелетных мышцах от миозинового стержня по его окружности отходят с
В скелетных мышцах от миозинового стержня по его окружности отходят с

МЕХАНИЗМ ОДИНОЧНОГО МЫШЕЧНОГО СОКРАЩЕНИЯ
1.Процесс, при котором потенциалы действия, генерируемые на
МЕХАНИЗМ ОДИНОЧНОГО МЫШЕЧНОГО СОКРАЩЕНИЯ
1.Процесс, при котором потенциалы действия, генерируемые на
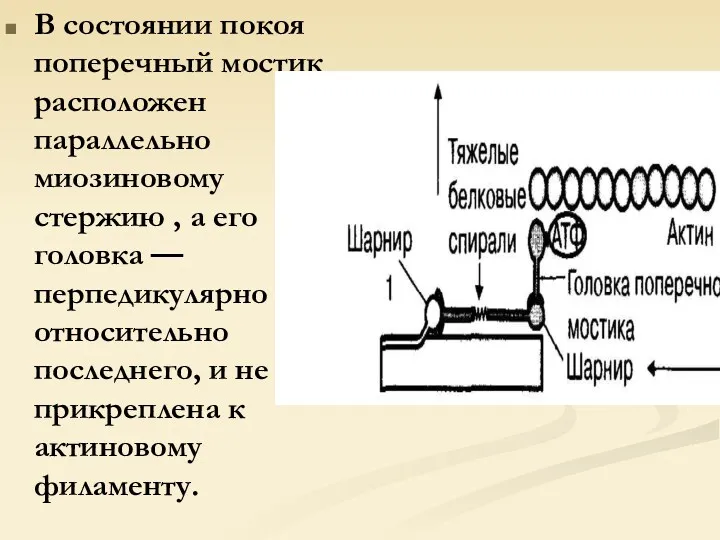
В состоянии покоя поперечный мостик расположен параллельно миозиновому стержию , а
В состоянии покоя поперечный мостик расположен параллельно миозиновому стержию , а

2.Для того чтобы вызвать сокращение, потенциал действия мышцы, вызваемый
2.Для того чтобы вызвать сокращение, потенциал действия мышцы, вызваемый

3.Когда концентрация ионов Са в межфибриллярном пространстве, т. е. в саркоалазме
3.Когда концентрация ионов Са в межфибриллярном пространстве, т. е. в саркоалазме
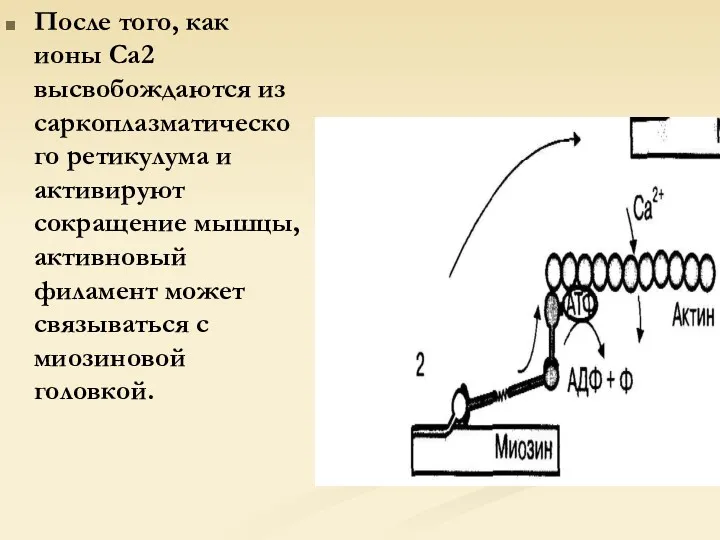
После того, как ионы Са2 высвобождаются из саркоплазматического ретикулума и активируют
После того, как ионы Са2 высвобождаются из саркоплазматического ретикулума и активируют
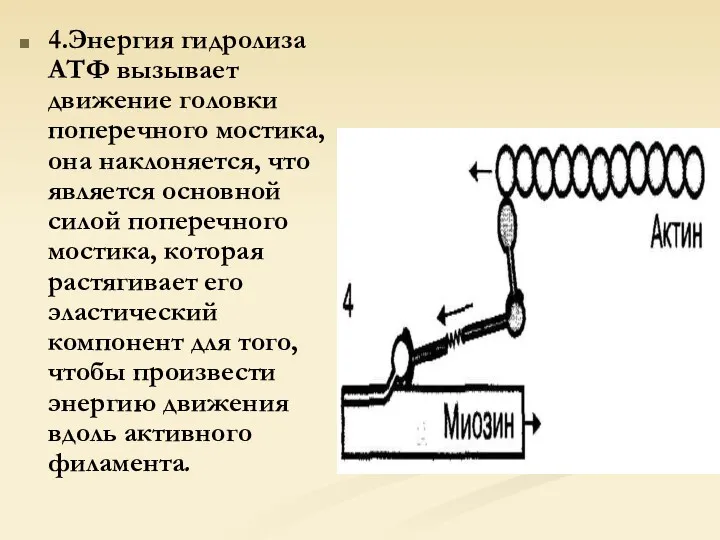
4.Энергия гидролиза АТФ вызывает движение головки поперечного мостика, она наклоняется, что
4.Энергия гидролиза АТФ вызывает движение головки поперечного мостика, она наклоняется, что

В результате головка мостика совершает движение на 40° к центру саркомера
В результате головка мостика совершает движение на 40° к центру саркомера

В результате одиночного мышечного сокращения :
Актиновые и миозиновые нити не укорачиваются
Взаимодействие
В результате одиночного мышечного сокращения :
Актиновые и миозиновые нити не укорачиваются
Взаимодействие

Расслабление скелетной мышцы
Расслабление скелетной мышцы начинается только после того,
Расслабление скелетной мышцы
Расслабление скелетной мышцы начинается только после того,

ТИПЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ
Сокращение мышцы в целом является результатом суммации одиночных
ТИПЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ
Сокращение мышцы в целом является результатом суммации одиночных
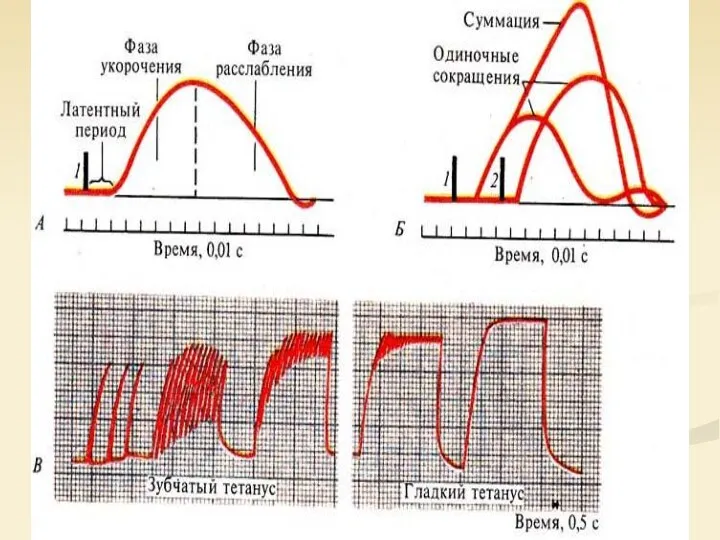
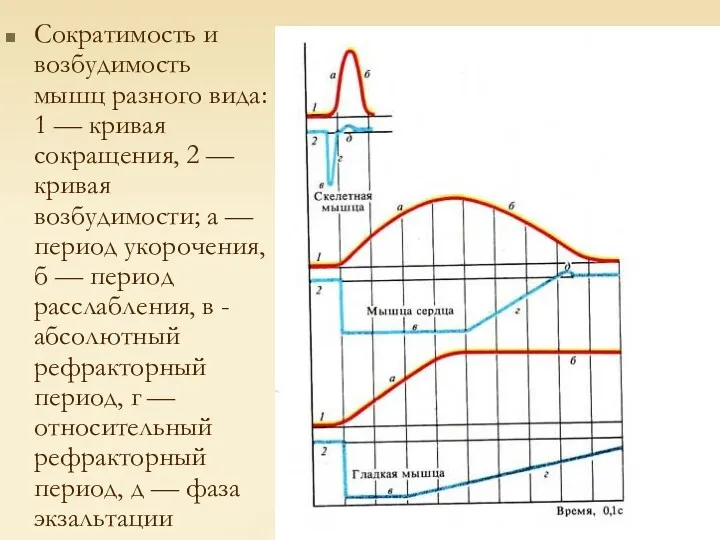
Сократимость и возбудимость мышц разного вида: 1 — кривая сокращения, 2
Сократимость и возбудимость мышц разного вида: 1 — кривая сокращения, 2

Тетаническим называется сокращение мышцы, при котором происходит суммация одиночных мышечных сокращений
Тетаническим называется сокращение мышцы, при котором происходит суммация одиночных мышечных сокращений

Гладкий тетанус возникает в том случае, если нерв или мьшцу раздражать
Гладкий тетанус возникает в том случае, если нерв или мьшцу раздражать
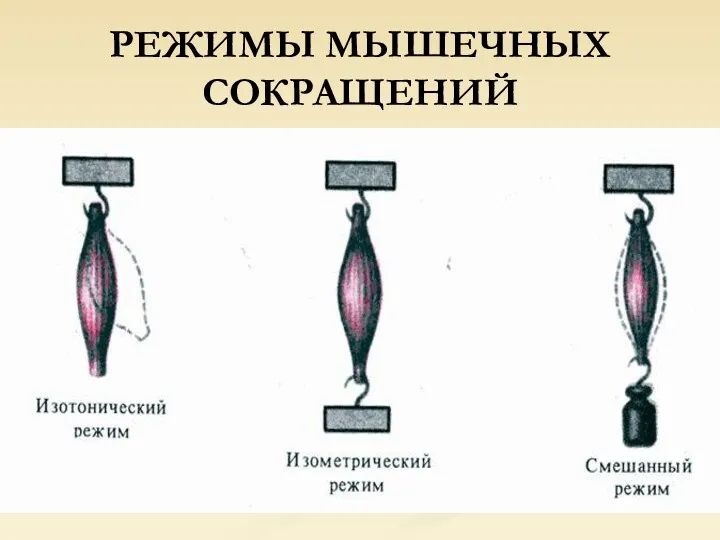
РЕЖИМЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ
РЕЖИМЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ

Если мышца развивает напряжение в условиях чрезмерной нагрузки, то ее длина
Если мышца развивает напряжение в условиях чрезмерной нагрузки, то ее длина

Во время перемещения тела в пространстве имеет место смешанный, или ауксотонический,
Во время перемещения тела в пространстве имеет место смешанный, или ауксотонический,

ЭНЕРГЕТИКА МЫШЕЧНОГО СОКРАЩЕНИЯ
АТФ в мышце необходима для:
Сокращения (образования мостиков)
Расслабления (разрыва мостиков)
Работы
ЭНЕРГЕТИКА МЫШЕЧНОГО СОКРАЩЕНИЯ
АТФ в мышце необходима для:
Сокращения (образования мостиков)
Расслабления (разрыва мостиков)
Работы

МЕХАНИЗМЫ ВОССТАНОВЛЕНИЯ АТФ
Креатинфосфокиназный. 3,6 моль АТФ/мин
Гликолитический. 1,2 моль АТФ/мин
Окислительный.
- при
МЕХАНИЗМЫ ВОССТАНОВЛЕНИЯ АТФ
Креатинфосфокиназный. 3,6 моль АТФ/мин
Гликолитический. 1,2 моль АТФ/мин
Окислительный.
- при
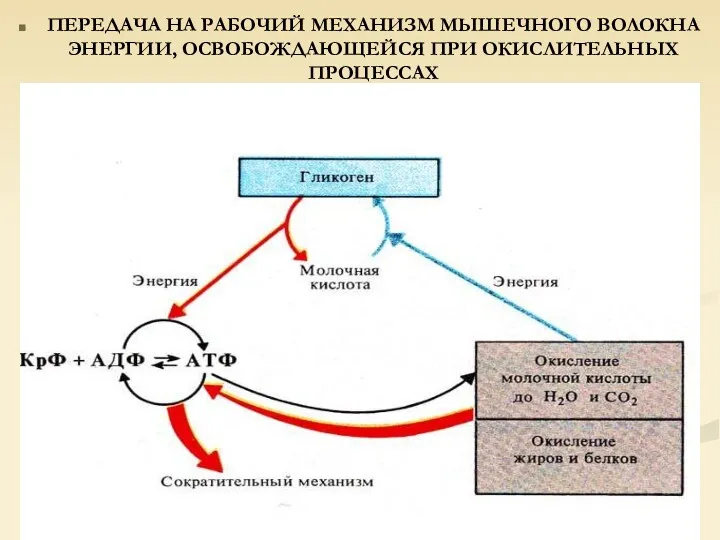
ПЕРЕДАЧА НА РАБОЧИЙ МЕХАНИЗМ МЫШЕЧНОГО ВОЛОКНА ЭНЕРГИИ, ОСВОБОЖДАЮЩЕЙСЯ ПРИ ОКИСЛИТЕЛЬНЫХ ПРОЦЕССАХ
ПЕРЕДАЧА НА РАБОЧИЙ МЕХАНИЗМ МЫШЕЧНОГО ВОЛОКНА ЭНЕРГИИ, ОСВОБОЖДАЮЩЕЙСЯ ПРИ ОКИСЛИТЕЛЬНЫХ ПРОЦЕССАХ
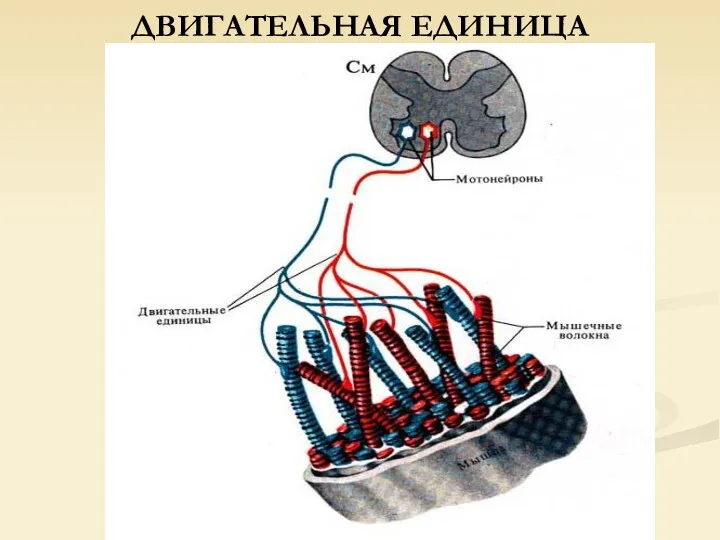
ДВИГАТЕЛЬНАЯ ЕДИНИЦА
ДВИГАТЕЛЬНАЯ ЕДИНИЦА

Единичное двигательное волокно мотонейрона и иннервируемые им мышечные волокна – ДВИГАТЕЛЬНАЯ
Единичное двигательное волокно мотонейрона и иннервируемые им мышечные волокна – ДВИГАТЕЛЬНАЯ

ОТЛИЧИЯ В НЕРВНЫХ КОМПОНЕНТАХ
Возбудимость мотонейронов одной и той же мышцы находится
ОТЛИЧИЯ В НЕРВНЫХ КОМПОНЕНТАХ
Возбудимость мотонейронов одной и той же мышцы находится

ОТЛИЧИЯ МЫШЕЧНЫХ ВОЛОКОН ДЕ
В быстрых мышечных волокнах :
Больше ёмкость саркоплазматического ретикулума
При
ОТЛИЧИЯ МЫШЕЧНЫХ ВОЛОКОН ДЕ
В быстрых мышечных волокнах :
Больше ёмкость саркоплазматического ретикулума
При

Внутри этих волокон содержится большое количество миоглобина (придающего им красный цвет),
Внутри этих волокон содержится большое количество миоглобина (придающего им красный цвет),

Физиологические показатели сокращения скелетной мышцы
Сила
Скорость развития силы и укорочения мышцы
Физиологические показатели сокращения скелетной мышцы
Сила
Скорость развития силы и укорочения мышцы

Скорость укорочения скелетной мышцы находится в обратной зависимости от величины нагрузки.
Скорость укорочения скелетной мышцы находится в обратной зависимости от величины нагрузки.

Работа мышц равна произведению силы, развиваемой мышцей, на путь перемещения груза,
Работа мышц равна произведению силы, развиваемой мышцей, на путь перемещения груза,

Оптимальная скорость укорочения скелетной мышцы зависит от оптимальной исходной длины саркомеров
Оптимальная скорость укорочения скелетной мышцы зависит от оптимальной исходной длины саркомеров

Мощность мышечной работы как физиологический параметр сокращения мышцы измеряется произведением величины
Мощность мышечной работы как физиологический параметр сокращения мышцы измеряется произведением величины

Утомление скелетной мышцы
Утомлением называется временное снижение способности мышцы развивать усилие
Утомление скелетной мышцы
Утомлением называется временное снижение способности мышцы развивать усилие

Унитарная гладкая мышца называется висцеральной, так как она окружает полые органы
Унитарная гладкая мышца называется висцеральной, так как она окружает полые органы
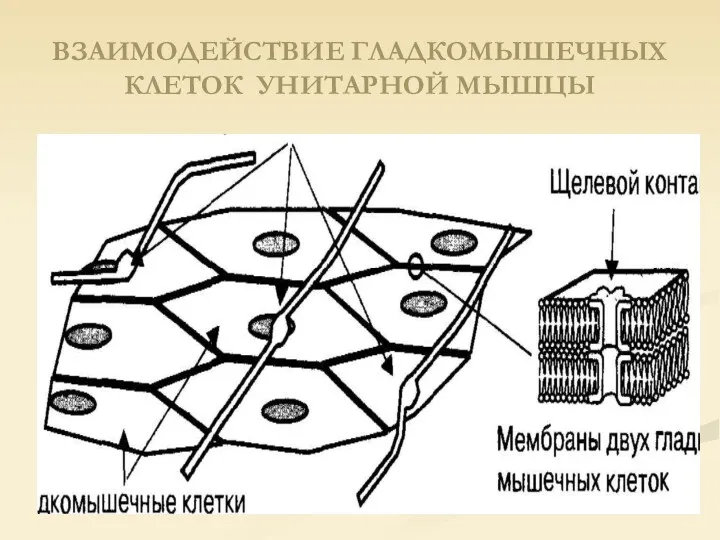
ВЗАИМОДЕЙСТВИЕ ГЛАДКОМЫШЕЧНЫХ КЛЕТОК УНИТАРНОЙ МЫШЦЫ
ВЗАИМОДЕЙСТВИЕ ГЛАДКОМЫШЕЧНЫХ КЛЕТОК УНИТАРНОЙ МЫШЦЫ

Мультиунитарная гладкая
мышца образована отдельными гладкомышечными клетками, каждая из которых иннервирована
Мультиунитарная гладкая мышца образована отдельными гладкомышечными клетками, каждая из которых иннервирована
 Ещё немного о белках
Ещё немного о белках Загадка человека. Наследственность. Урок 3
Загадка человека. Наследственность. Урок 3 Тип Членистоногие. Класс Паукообразные
Тип Членистоногие. Класс Паукообразные Кольчатые черви
Кольчатые черви Урок по теме Семейства класса двудольных растений
Урок по теме Семейства класса двудольных растений Гуморальная регуляция. Эндокринная система
Гуморальная регуляция. Эндокринная система Строение клеток живых организмов. Лабораторная работа
Строение клеток живых организмов. Лабораторная работа Мінез-құлық қалыптасуының жалпы принцептері. ЖЖІӘ туралы түсінік. Туа біткен және жүре пайда болған іс әрекеттерінің түрлері
Мінез-құлық қалыптасуының жалпы принцептері. ЖЖІӘ туралы түсінік. Туа біткен және жүре пайда болған іс әрекеттерінің түрлері Биоактивные пептиды
Биоактивные пептиды Нейромедиаторы. Критерии нейромедиатора
Нейромедиаторы. Критерии нейромедиатора Структурно-функциональные особенности биокатализа
Структурно-функциональные особенности биокатализа песчаная эфа
песчаная эфа Минеральное питание растений. Ассимиляция элементов
Минеральное питание растений. Ассимиляция элементов Цикличность поведения женских особей
Цикличность поведения женских особей Анатомия домашних животных
Анатомия домашних животных Понятие об организме. Основные части организма
Понятие об организме. Основные части организма Biological effects of nitric oxide and its role in cell signaling
Biological effects of nitric oxide and its role in cell signaling Формы размножения организмов. Бесполое размножение
Формы размножения организмов. Бесполое размножение Класс Пресмыкающиеся
Класс Пресмыкающиеся Многообразие насекомых
Многообразие насекомых Барыня тыква
Барыня тыква Филогенетический анализ и молекулярная эволюция. Лекция 5
Филогенетический анализ и молекулярная эволюция. Лекция 5 Черепно-мозговые нервы
Черепно-мозговые нервы Биофизика гормональных рецепторов
Биофизика гормональных рецепторов Подготовка к Всероссийской проверочной работе по биологии. 5 класс
Подготовка к Всероссийской проверочной работе по биологии. 5 класс Наука биология и условия жизни организмов
Наука биология и условия жизни организмов Нуклеїнові кислоти (ДНК та РНК)
Нуклеїнові кислоти (ДНК та РНК) Биомембраны. Пассивный и активный транспорт веществ
Биомембраны. Пассивный и активный транспорт веществ