- Физиология растений. Лекция 3

Содержание
- 2. Фотосинтетическое фосфорилирование (ФФ) ФФ – это синтез молекулы АТФ за счет энергии трансмембранного потенциала (ΔμН+) тилакоидной
- 3. Типы фотофосфорилирования Нециклическое фотофосфорилирование. 2АДФ + Фн + 2НАДФ+ + 2 Н2О 2АТФ + + О2
- 4. Механизм фотосинтетического фосфорилирования
- 5. Хемио-осмотическая гипотеза (Митчелл, Ягендорф)
- 6. Структура АТФ-синтазы
- 7. Биохимия фотосинтеза Сущность темновой фазы фотосинтеза заключается в трансформации макроэргической связи АТФ и НАДФН в стабильную
- 8. Цикл Кальвина В 1957 году Кальвин предложил биохимический механизм превращения СО2 в фосфоглицериновый альдегид (ФГА). 1
- 9. Методы Кальвина В лаборатории Кальвина, Басхэма и Бенсона впервые использовали методы: 1. Для фотосинтеза использовали *СО2
- 10. Цикл Кальвина Включение СО2 в цикл 1) Активирование первичного акцептора (рибулозомонофосфат) РМФ + АТФ → РБФ
- 11. 2). Карбоксилирование рибулозобисфосфата
- 12. Свойства рибулозобисфосфаткарбоксилазы РБФ-карбоксилаза (Rubisco) КФ 4.1.1.39 – самый распространенный белок биосферы: а) 50% от общего количества
- 13. Свойства Rubisco 2. Фермент двойного кодирования. Rubisco имеет молекулярную массу 550 кДа. Состоит из 8 больших
- 14. Свойства Rubisco Сборка белка происходит в хлоропластах при участии белка шаперона (Hsc60), который кодируется в ядре.
- 15. Свойства Rubisco Димеры больших субъединиц каталитически активны и в отсутствие малых субъединиц. Малые субъединицы играют регуляторную
- 16. Рибулозо-1,5-бисфосфаткарбоксилаза
- 17. II. Фаза восстановления Осуществляется с помощью двух реакций: А) активирование 3ФГК: 3ФГК + АТФ → 1,3ФГК
- 18. III. Регенерация первичного акцептора Общее уравнение образования ФГА 3 РМФ + 9 АТФ + 6 НАДФН
- 19. III.Регенерация первичного акцептора С3 – фосфоглицериновый альдегид (ФГА) С6 – фруктозо-1,6-БФ С4 – эритрозо-4-Ф С7 –
- 20. Фотодыхание Фотодыхание – это светоиндуцируемое выделение углекислого газа зелеными листьями растений. Фотодыхание обнаружено в 1965 году.
- 22. Причины фотодыхания 1. Наличие оксигеназной функции у рибулозобисфосфаткарбоксилазы. 2. Высокая концентрация кислорода в атмосфере, составляющая 21%.
- 23. Значение фотодыхания На фотодыхательный метаболизм у С3-растений тратится до 55% продуктов фотосинтеза. 1. Фотодыхание выступает как
- 24. Значение фотодыхания Фотодыхание – это «расплата» растений за созданную ими кислородную атмосферу. Потеря 40-50% продуктов фотосинтеза
- 25. Цикл Хэтча-Слэйка Карпилов И.С. в 1963 году обнаружил включение 14СО2 в яблочную кислоту в листьях кукурузы.
- 26. Анатомия листа у С4 - растений
- 27. Анатомия листа у С4 - растений Для С4-растений характерна кранц-анатомия листа. Внутреннее кольцо (обкладка) изолировано клетками
- 28. Биохимия цикла Хэтча-Слейка 1 – ФЕП-карбоксилаза, 2 – малатдегидрогеназа (НАД-зависимая), 3 – малатдегидрогеназа (декарбоксилир.), 4 -
- 29. Физиологическая роль цикла Хэтча-Слэйка Физиологическое значение цикла: 1) Увеличение концентрации СО2 в клетках обкладки. Используется новый
- 30. Физиологическая роль цикла Хэтча-Слэйка 3) ФЕП-карбоксилаза является уникальным ферментом, обладающим большим сродством к СО2, что имеет
- 31. Эволюция фотосинтеза Причины эволюции в последние 600 млн. лет обусловлены резким изменением факторов внешней среды. 1.
- 32. Метаболизм органических кислот по типу толстянковых Растения, осуществляющие метаболизм по типу толстянковых, являются суккулентами. Большинство принадлежат
- 33. САМ-метаболизм Данные растения адаптировались к дефициту воды на биохимическом уровне. Днем устьица у этих растений закрываются,
- 34. САМ-метаболизм
- 35. Сущность САМ-метаболизма 1. САМ-фотосинтез осуществляет разделение ассимиляции СО2 и цикла Кальвина не в пространстве, а во
- 36. Сравнительная характеристика основных показателей С3-, С4- и САМ-растений
- 37. Фотоокисление воды До 1931 года доминировала формальдегидная гипотеза происхождения кислорода. СО2 С + О2 С +
- 38. Происхождение кислорода Ван-Ниль в 1931 году предположил, что кислород образуется из воды. СО2 + Н218О СН2О
- 39. Механизм выделения кислорода Согласно Кутюрину (1968), молекула кислорода образуется из воды, в окислении которой участвует непосредственно
- 40. Механизм выделения кислорода Mn-содержащий ферментный комплекс
- 42. Скачать презентацию

Фотосинтетическое фосфорилирование (ФФ)
ФФ – это синтез молекулы АТФ за счет энергии
Фотосинтетическое фосфорилирование (ФФ)
ФФ – это синтез молекулы АТФ за счет энергии

Типы фотофосфорилирования
Нециклическое фотофосфорилирование.
2АДФ + Фн + 2НАДФ+ + 2 Н2О 2АТФ
Типы фотофосфорилирования
Нециклическое фотофосфорилирование.
2АДФ + Фн + 2НАДФ+ + 2 Н2О 2АТФ
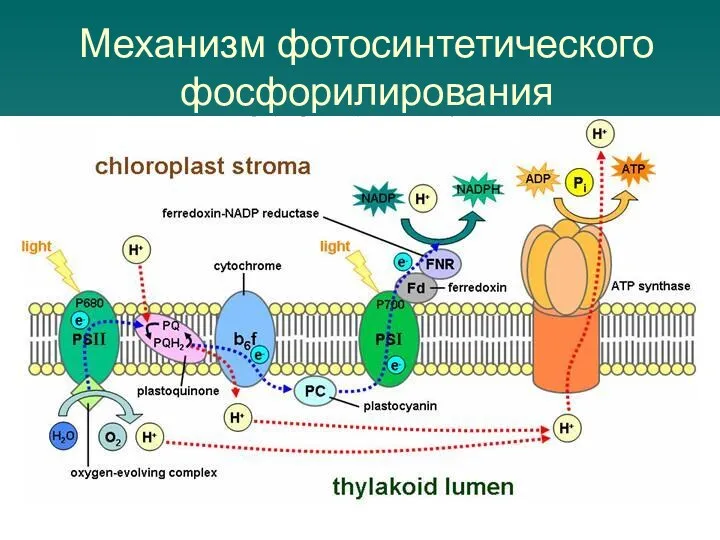
Механизм фотосинтетического фосфорилирования
Механизм фотосинтетического фосфорилирования
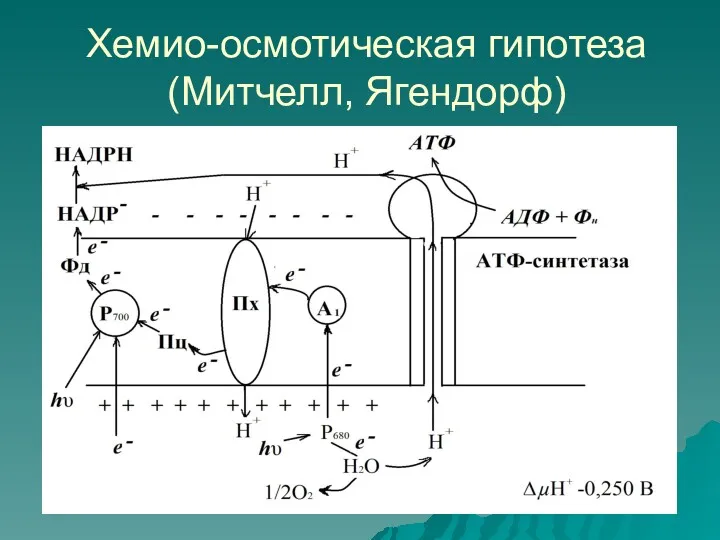
Хемио-осмотическая гипотеза (Митчелл, Ягендорф)
Хемио-осмотическая гипотеза (Митчелл, Ягендорф)
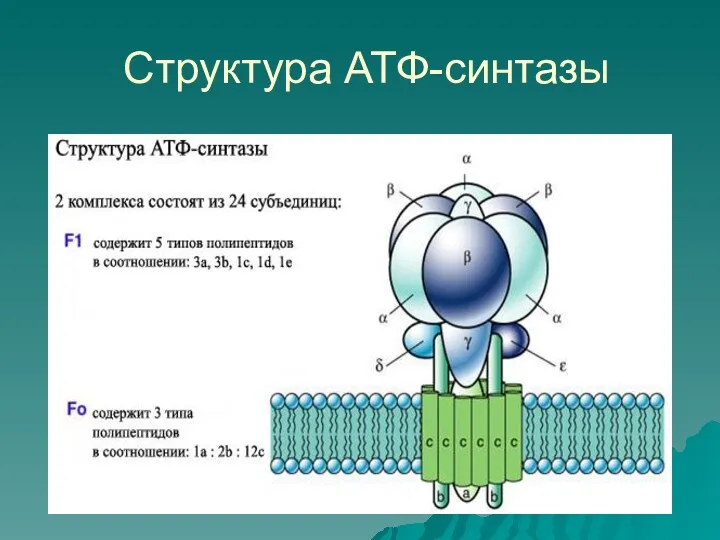
Структура АТФ-синтазы
Структура АТФ-синтазы

Биохимия фотосинтеза
Сущность темновой фазы фотосинтеза заключается в трансформации макроэргической связи АТФ
Биохимия фотосинтеза
Сущность темновой фазы фотосинтеза заключается в трансформации макроэргической связи АТФ

Цикл Кальвина
В 1957 году Кальвин предложил биохимический механизм превращения СО2 в
Цикл Кальвина
В 1957 году Кальвин предложил биохимический механизм превращения СО2 в

Методы Кальвина
В лаборатории Кальвина, Басхэма и Бенсона впервые использовали методы:
1.
Методы Кальвина
В лаборатории Кальвина, Басхэма и Бенсона впервые использовали методы:
1.

Цикл Кальвина
Включение СО2 в цикл
1) Активирование первичного акцептора
(рибулозомонофосфат)
РМФ + АТФ →
Цикл Кальвина
Включение СО2 в цикл
1) Активирование первичного акцептора
(рибулозомонофосфат)
РМФ + АТФ →
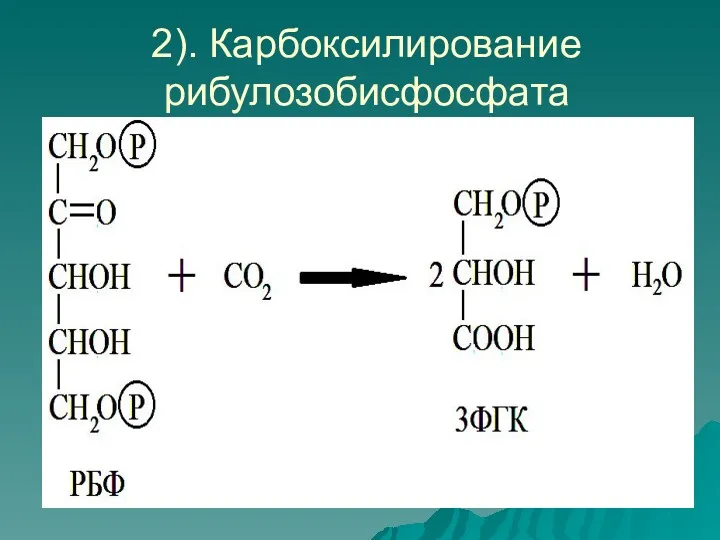
2). Карбоксилирование рибулозобисфосфата
2). Карбоксилирование рибулозобисфосфата

Свойства рибулозобисфосфаткарбоксилазы
РБФ-карбоксилаза (Rubisco)
КФ 4.1.1.39 – самый распространенный белок биосферы:
а)
Свойства рибулозобисфосфаткарбоксилазы
РБФ-карбоксилаза (Rubisco)
КФ 4.1.1.39 – самый распространенный белок биосферы:
а)

Свойства Rubisco
2. Фермент двойного кодирования.
Rubisco имеет молекулярную массу 550 кДа. Состоит
Свойства Rubisco
2. Фермент двойного кодирования.
Rubisco имеет молекулярную массу 550 кДа. Состоит

Свойства Rubisco
Сборка белка происходит в хлоропластах при участии белка шаперона (Hsc60),
Свойства Rubisco
Сборка белка происходит в хлоропластах при участии белка шаперона (Hsc60),

Свойства Rubisco
Димеры больших субъединиц каталитически активны и в отсутствие малых субъединиц.
Свойства Rubisco
Димеры больших субъединиц каталитически активны и в отсутствие малых субъединиц.
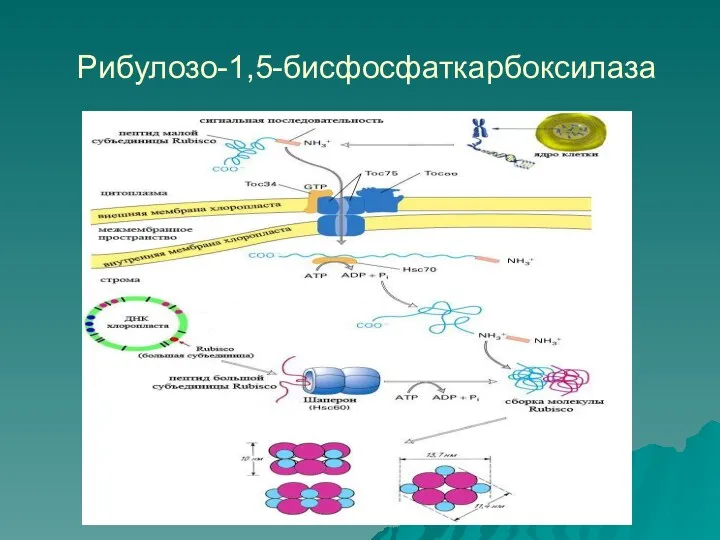
Рибулозо-1,5-бисфосфаткарбоксилаза
Рибулозо-1,5-бисфосфаткарбоксилаза

II. Фаза восстановления
Осуществляется с помощью двух реакций:
А) активирование 3ФГК:
3ФГК
II. Фаза восстановления
Осуществляется с помощью двух реакций:
А) активирование 3ФГК:
3ФГК
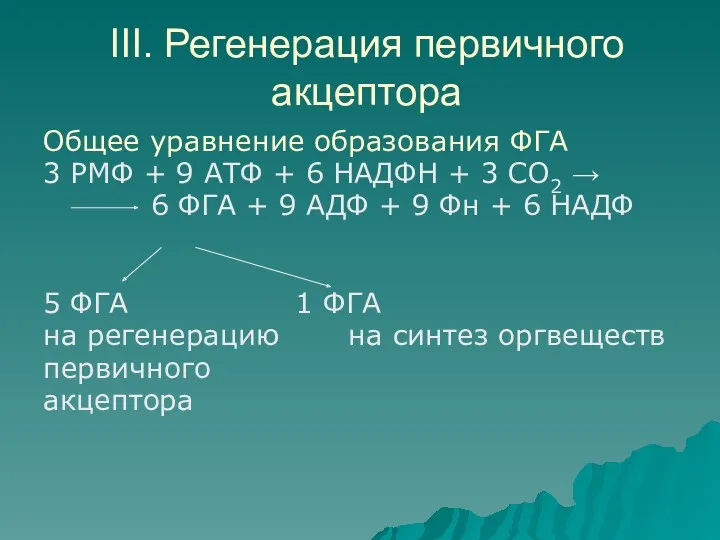
III. Регенерация первичного акцептора
Общее уравнение образования ФГА
3 РМФ + 9
III. Регенерация первичного акцептора
Общее уравнение образования ФГА
3 РМФ + 9
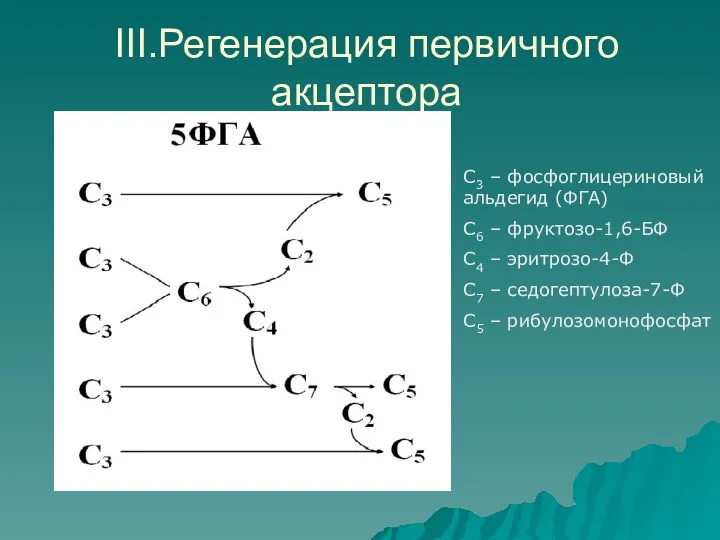
III.Регенерация первичного акцептора
С3 – фосфоглицериновый альдегид (ФГА)
С6 – фруктозо-1,6-БФ
С4 – эритрозо-4-Ф
С7
III.Регенерация первичного акцептора
С3 – фосфоглицериновый альдегид (ФГА)
С6 – фруктозо-1,6-БФ
С4 – эритрозо-4-Ф
С7

Фотодыхание
Фотодыхание – это светоиндуцируемое выделение углекислого газа зелеными листьями растений.
Фотодыхание
Фотодыхание
Фотодыхание – это светоиндуцируемое выделение углекислого газа зелеными листьями растений.
Фотодыхание
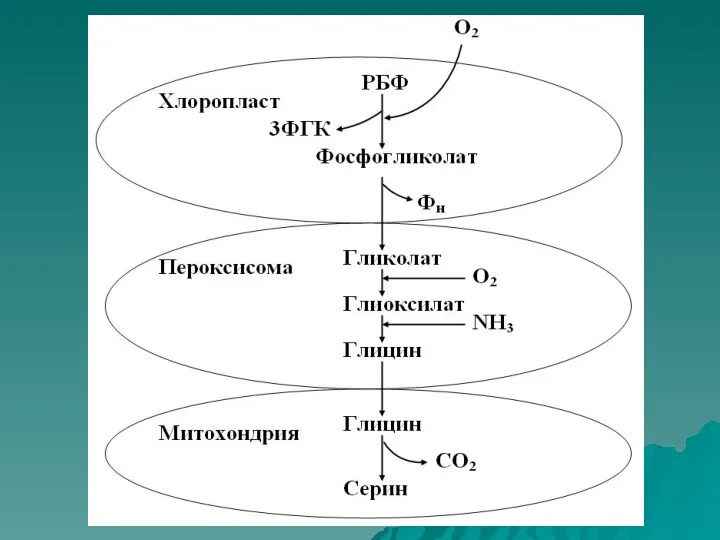

Причины фотодыхания
1. Наличие оксигеназной функции у рибулозобисфосфаткарбоксилазы.
2. Высокая концентрация кислорода в
Причины фотодыхания
1. Наличие оксигеназной функции у рибулозобисфосфаткарбоксилазы.
2. Высокая концентрация кислорода в

Значение фотодыхания
На фотодыхательный метаболизм у С3-растений тратится до 55% продуктов фотосинтеза.
1.
Значение фотодыхания
На фотодыхательный метаболизм у С3-растений тратится до 55% продуктов фотосинтеза.
1.

Значение фотодыхания
Фотодыхание – это «расплата» растений за созданную ими кислородную атмосферу.
Значение фотодыхания
Фотодыхание – это «расплата» растений за созданную ими кислородную атмосферу.

Цикл Хэтча-Слэйка
Карпилов И.С. в 1963 году обнаружил включение 14СО2 в яблочную
Цикл Хэтча-Слэйка
Карпилов И.С. в 1963 году обнаружил включение 14СО2 в яблочную
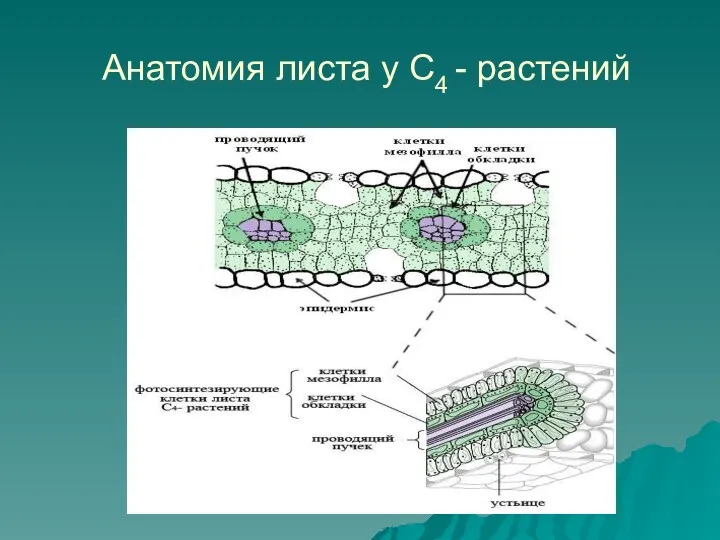
Анатомия листа у С4 - растений
Анатомия листа у С4 - растений

Анатомия листа у С4 - растений
Для С4-растений характерна кранц-анатомия листа.
Внутреннее
Анатомия листа у С4 - растений
Для С4-растений характерна кранц-анатомия листа.
Внутреннее
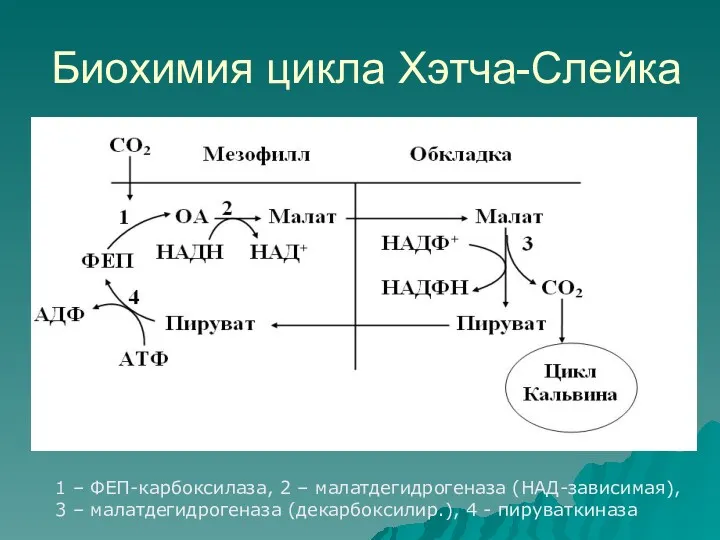
Биохимия цикла Хэтча-Слейка
1 – ФЕП-карбоксилаза, 2 – малатдегидрогеназа (НАД-зависимая), 3 –
Биохимия цикла Хэтча-Слейка
1 – ФЕП-карбоксилаза, 2 – малатдегидрогеназа (НАД-зависимая), 3 –

Физиологическая роль цикла Хэтча-Слэйка
Физиологическое значение цикла:
1) Увеличение концентрации СО2 в клетках
Физиологическая роль цикла Хэтча-Слэйка
Физиологическое значение цикла:
1) Увеличение концентрации СО2 в клетках

Физиологическая роль цикла Хэтча-Слэйка
3) ФЕП-карбоксилаза является уникальным ферментом, обладающим большим сродством
Физиологическая роль цикла Хэтча-Слэйка
3) ФЕП-карбоксилаза является уникальным ферментом, обладающим большим сродством

Эволюция фотосинтеза
Причины эволюции в последние 600 млн. лет обусловлены резким изменением
Эволюция фотосинтеза
Причины эволюции в последние 600 млн. лет обусловлены резким изменением

Метаболизм органических кислот по типу толстянковых
Растения, осуществляющие метаболизм по типу толстянковых,
Метаболизм органических кислот по типу толстянковых
Растения, осуществляющие метаболизм по типу толстянковых,

САМ-метаболизм
Данные растения адаптировались к дефициту воды на биохимическом уровне. Днем устьица
САМ-метаболизм
Данные растения адаптировались к дефициту воды на биохимическом уровне. Днем устьица
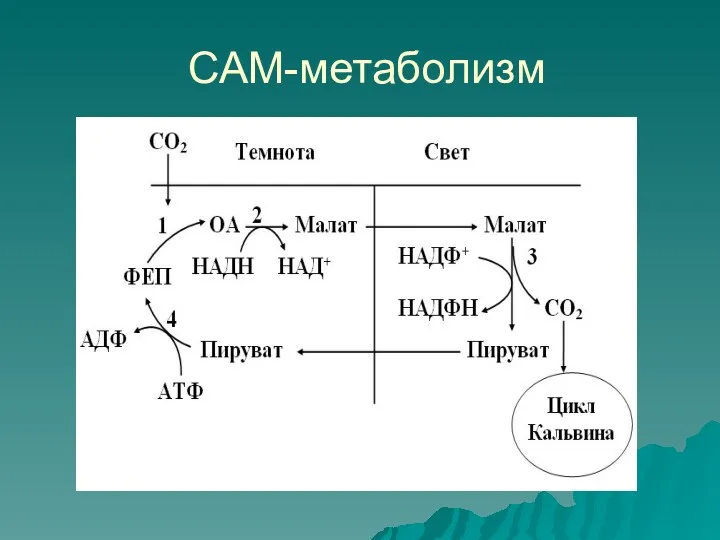
САМ-метаболизм
САМ-метаболизм

Сущность САМ-метаболизма
1. САМ-фотосинтез осуществляет разделение ассимиляции СО2 и цикла Кальвина не
Сущность САМ-метаболизма
1. САМ-фотосинтез осуществляет разделение ассимиляции СО2 и цикла Кальвина не
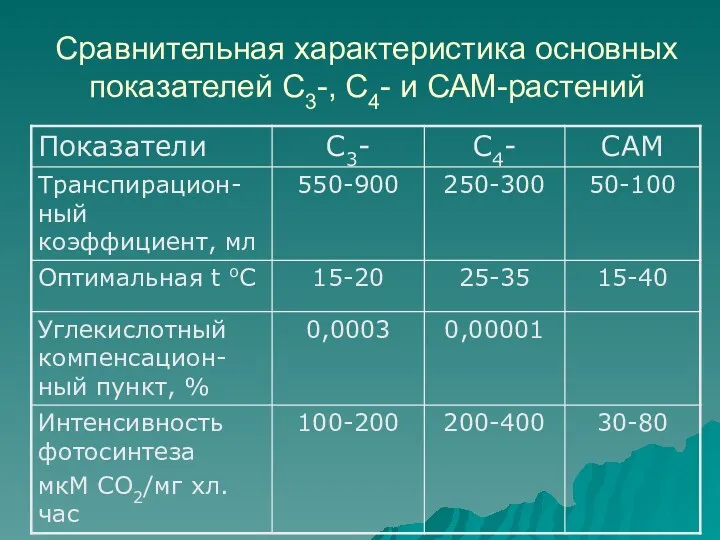
Сравнительная характеристика основных показателей С3-, С4- и САМ-растений
Сравнительная характеристика основных показателей С3-, С4- и САМ-растений

Фотоокисление воды
До 1931 года доминировала формальдегидная гипотеза происхождения кислорода.
СО2 С +
Фотоокисление воды
До 1931 года доминировала формальдегидная гипотеза происхождения кислорода.
СО2 С +

Происхождение кислорода
Ван-Ниль в 1931 году предположил, что кислород образуется из воды.
СО2
Происхождение кислорода
Ван-Ниль в 1931 году предположил, что кислород образуется из воды.
СО2

Механизм выделения кислорода
Согласно Кутюрину (1968), молекула кислорода образуется из воды, в
Механизм выделения кислорода
Согласно Кутюрину (1968), молекула кислорода образуется из воды, в

Механизм выделения кислорода
Mn-содержащий ферментный комплекс
Механизм выделения кислорода
Mn-содержащий ферментный комплекс
 Нормальная анатомия легких, плевры и средостения
Нормальная анатомия легких, плевры и средостения Черенкование комнатных растений
Черенкование комнатных растений Такие разные жуки. Коллекции. Занятие № 2. Консультация. Планирование деятельности
Такие разные жуки. Коллекции. Занятие № 2. Консультация. Планирование деятельности Введение в системную биологию
Введение в системную биологию Разнообразие животных
Разнообразие животных Болезни органов дыхания и их предупреждение. Гигиена дыхания
Болезни органов дыхания и их предупреждение. Гигиена дыхания Развитие низших хордовых. Амфибии
Развитие низших хордовых. Амфибии Специфика энергетических систем растительной клетки
Специфика энергетических систем растительной клетки Летучие мыши
Летучие мыши Анатомо-физиологические особенности сердечно сосудистой системы
Анатомо-физиологические особенности сердечно сосудистой системы Презентация у уроку Строение и функции пищеварительной системы человека 8 класс
Презентация у уроку Строение и функции пищеварительной системы человека 8 класс Вопросы экзамена по нервной системе
Вопросы экзамена по нервной системе Устройство речевого аппарата
Устройство речевого аппарата Жизнь и научная деятельность Ж.Б.Ламарка (1744-1829)
Жизнь и научная деятельность Ж.Б.Ламарка (1744-1829) Мышцы шеи
Мышцы шеи Половое размножение у покрытосеменных растений
Половое размножение у покрытосеменных растений Матричные биосинтезы
Матричные биосинтезы Мендель Грегор Иоганн
Мендель Грегор Иоганн Растения-рекордсмены
Растения-рекордсмены Выделительная система человека
Выделительная система человека Онтогенез. Лекция 14
Онтогенез. Лекция 14 Биологиялық тыңайтқыштар
Биологиялық тыңайтқыштар Все о грибах
Все о грибах Биологический метод исследования. Определение патогенности и вирулентности микроорганизмов
Биологический метод исследования. Определение патогенности и вирулентности микроорганизмов Научно-исследовательский проект Удивительные свойства сахара
Научно-исследовательский проект Удивительные свойства сахара Процеси гниття. Хімізм процесу гниття
Процеси гниття. Хімізм процесу гниття Ұлпаның және ағзаның электростимуляциясы
Ұлпаның және ағзаның электростимуляциясы Лабораторная диагностика стафилококковой инфекции
Лабораторная диагностика стафилококковой инфекции