- Физиология возбудимых клеток. Мембранный потенциал

Содержание
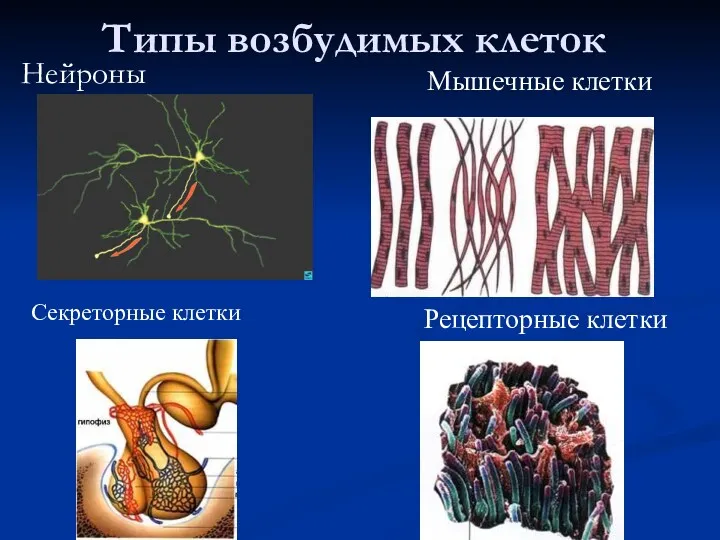
- 2. Типы возбудимых клеток Нейроны Мышечные клетки Секреторные клетки Рецепторные клетки
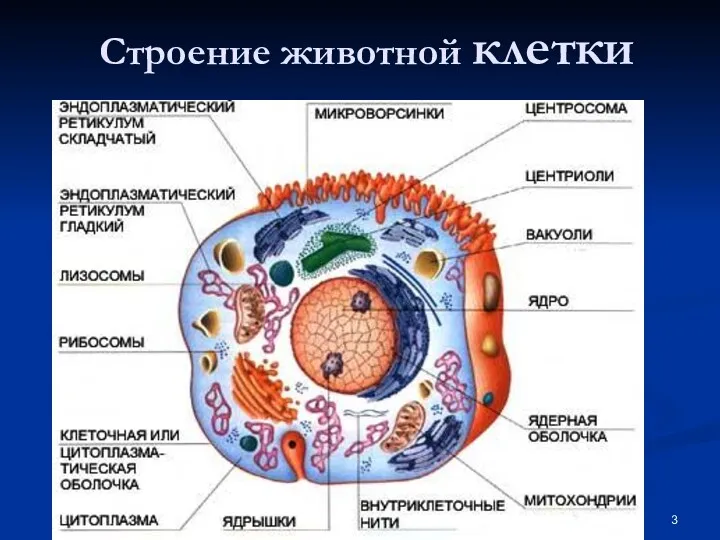
- 3. Строение животной клетки
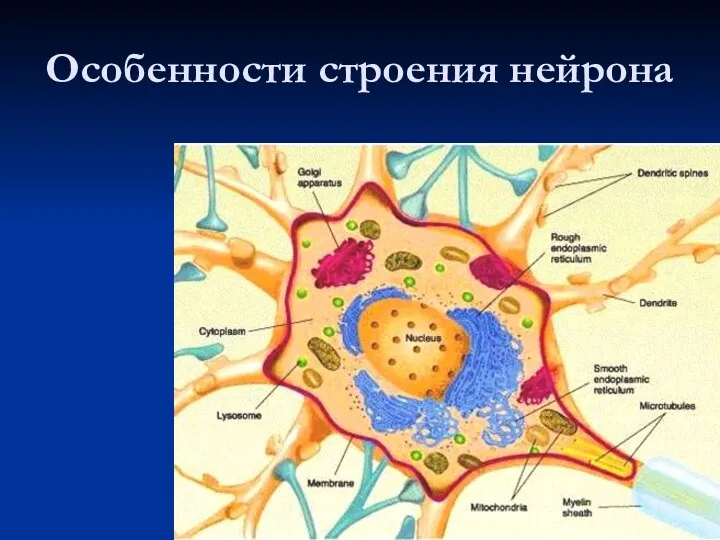
- 4. Особенности строения нейрона
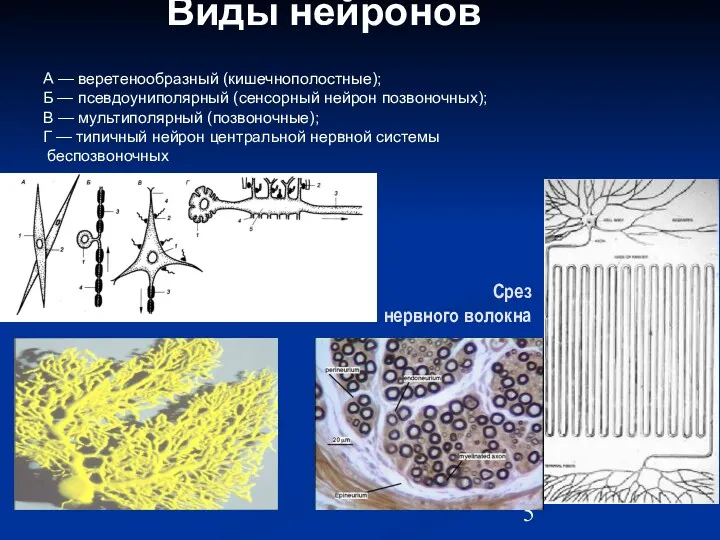
- 5. Виды нейронов А — веретенообразный (кишечнополостные); Б — псевдоуниполярный (сенсорный нейрон позвоночных); В — мультиполярный (позвоночные);
- 6. Формирование трансмембранного потенциала А. в чашке Петри KCl K+ Cl- Градиент концентрации Градиент заряда равновесие
- 7. Рассчет заряда на мембране Равновесный потенциал для какого-либо иона Х можно рассчитать из уравнения, полученного в
- 8. Б. мицелла – синтетический прообраз клетки К+ А- _ + К+
- 9. Мембрана живой клетки К+ Na+ Са++
- 10. Равновесные потенциалы(Е) Движущая сила (V- Е) Cl- -89 - 47 Cl-каналы
- 11. Мембрана живой клетки полупроницаема -61 К+ Na+ = 0,023 рК Са++ рСа++ = 0 Cl-
- 12. Проницаемость обеспечена ионные каналами мембраны 1-1000 каналов на квадратный микрометр мембраны Центральная водная пора Устья канала:
- 13. Создание градиента концентрации: 1. Na-K АТФ-аза 2. ионные обменники Транспорт 3 Na/2K за счет энергии 1
- 14. Изменения мембранного потенциала покоя 1. Деполяризация- уменьшение (ее скорость определяется постоянной времени (τm=RmCm)) 2. Гиперполяризация- увеличение
- 15. Внутриклеточная регистрация мембранного потенциала покоя Внутриклеточная микроэлектродная регистрация Величина МПП в возбудимых клетках – от -60
- 16. Потенциал действия Фаза деполяризации Фаза реполяризации Раздражающий импульс
- 17. Вызывается сверхпороговым раздражением Амплитуда не зависит от силы раздражения Распространяется по всей мембране не затухая Связан
- 18. Временной ход ионных токов во время потенциала действия
- 19. Фармакологическое разделение ионных токов ядами контроль Калиевый ток Натриевый ток Выводы Входящий ток переносится ионами натрия,
- 20. Фазы потенциала действия 1- порог (около 50 мв, ток Na>K) 2- деполяризация 0,5 мс (вход Na)
- 21. Исследование отдельного канала Возможность исследовать отдельный канал Возможность менять потенциал на мембране Возможность менять ионный состав
- 22. Нобелевская премия 1991 года в области физиологии и медицины Эрвин Нейер и Берт Сакманн «за открытия
- 23. Канал имеет воротный механизм 1- покой 2-деполяризация 3-рефрактерность Динамика открытия ворот 1 2 3 За один
- 24. Молекулярные механизмы активации и инактивации у большинства каналов общие
- 25. Работа Na+ канала
- 26. Белковая структура канала: 4 домена из 6 сегментов каждый Структура Cl- канала S4-воротный механизм, S5 и
- 27. Рефрактерность - снижение способности клетки отвечать на раздражение в результате временной инактивации натриевых каналов Абсолютная рефрактерность
- 28. Распространение потенциала действия по волокну Увеличение диаметра волокна повышает скорость проведения: Постоянная длины волокна (от 0,1
- 29. Миелинизированные волокна Эстафетный (до 40 м/с) и сальтаторный (до 120м/с) механизмы распространения возбуждения
- 30. Скорость проведения ПД по разным типам волокон
- 31. Виды регистрации ПД Внутриклеточная монополярная Внеклеточная биполярная
- 32. Использование флуоресцентных красителей
- 34. Скачать презентацию
Типы возбудимых клеток
Нейроны
Мышечные клетки
Секреторные клетки
Рецепторные клетки
Типы возбудимых клеток
Нейроны
Мышечные клетки
Секреторные клетки
Рецепторные клетки
Строение животной клетки
Строение животной клетки
Особенности строения нейрона
Особенности строения нейрона
Виды нейронов
А — веретенообразный (кишечнополостные);
Б — псевдоуниполярный (сенсорный нейрон позвоночных);
В
Виды нейронов
А — веретенообразный (кишечнополостные);
Б — псевдоуниполярный (сенсорный нейрон позвоночных);
В
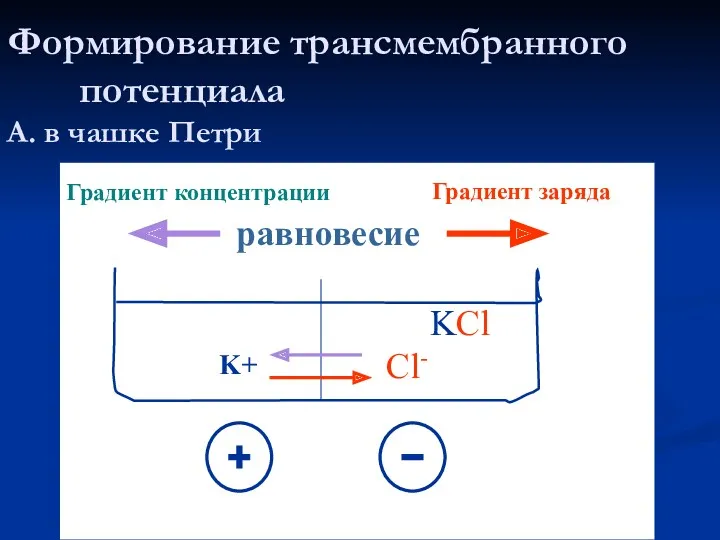
Формирование трансмембранного потенциала
А. в чашке Петри
KCl
K+
Cl-
Градиент концентрации
Градиент заряда
равновесие
Формирование трансмембранного потенциала
А. в чашке Петри
KCl
K+
Cl-
Градиент концентрации
Градиент заряда
равновесие

Рассчет заряда на мембране
Равновесный потенциал для какого-либо иона Х можно рассчитать
Рассчет заряда на мембране
Равновесный потенциал для какого-либо иона Х можно рассчитать
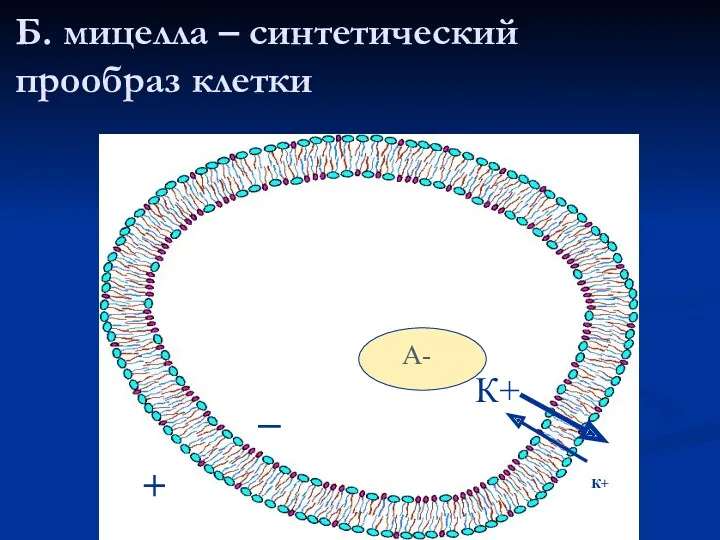
Б. мицелла – синтетический прообраз клетки
К+
А-
_
+
К+
Б. мицелла – синтетический прообраз клетки
К+
А-
_
+
К+
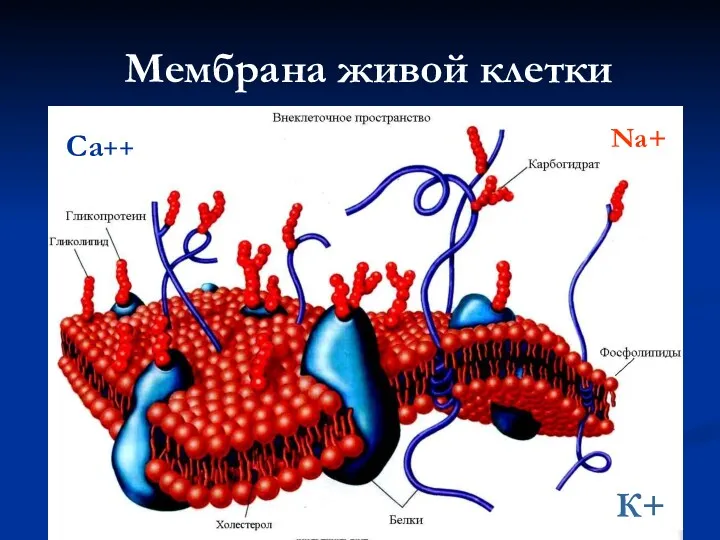
Мембрана живой клетки
К+
Na+
Са++
Мембрана живой клетки
К+
Na+
Са++

Равновесные потенциалы(Е) Движущая сила (V- Е)
Cl-
-89 - 47
Cl-каналы
Равновесные потенциалы(Е) Движущая сила (V- Е)
Cl-
-89 - 47
Cl-каналы
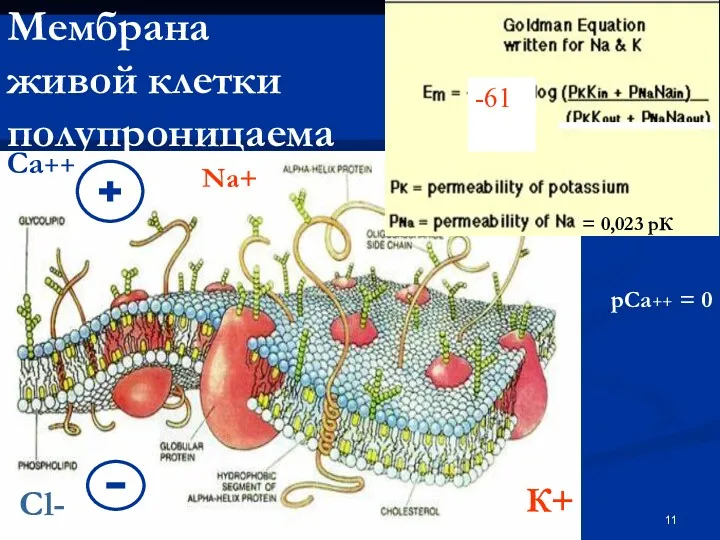
Мембрана
живой клетки полупроницаема
-61
К+
Na+
= 0,023 рК
Са++
рСа++ = 0
Cl-
Мембрана
живой клетки полупроницаема
-61
К+
Na+
= 0,023 рК
Са++
рСа++ = 0
Cl-

Проницаемость обеспечена ионные каналами мембраны
1-1000 каналов на квадратный микрометр мембраны
Центральная водная
Проницаемость обеспечена ионные каналами мембраны
1-1000 каналов на квадратный микрометр мембраны
Центральная водная
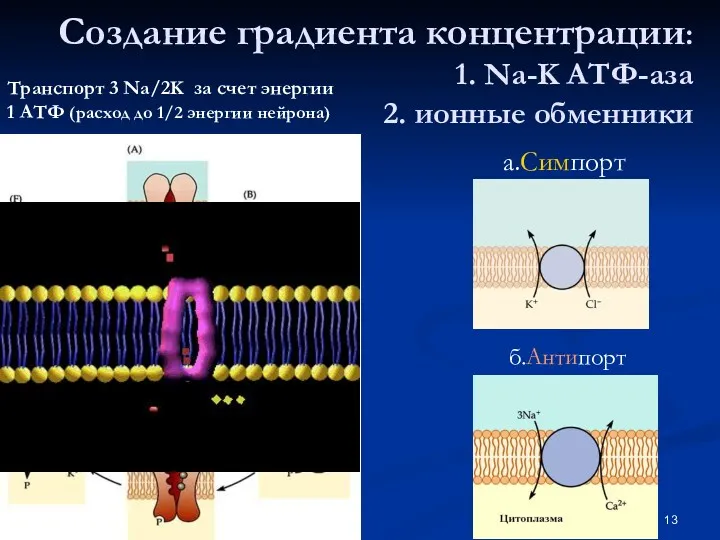
Создание градиента концентрации:
1. Na-K АТФ-аза
2. ионные обменники
Транспорт 3 Na/2K
Создание градиента концентрации:
1. Na-K АТФ-аза
2. ионные обменники
Транспорт 3 Na/2K
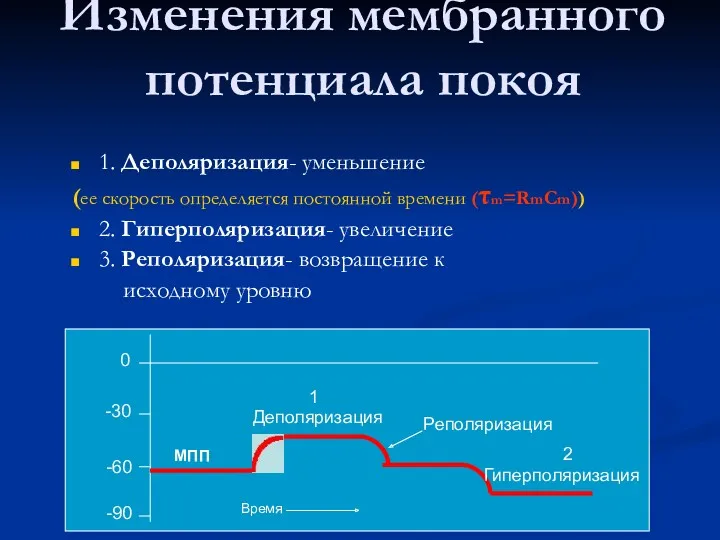
Изменения мембранного потенциала покоя
1. Деполяризация- уменьшение
(ее скорость определяется постоянной времени
Изменения мембранного потенциала покоя
1. Деполяризация- уменьшение
(ее скорость определяется постоянной времени
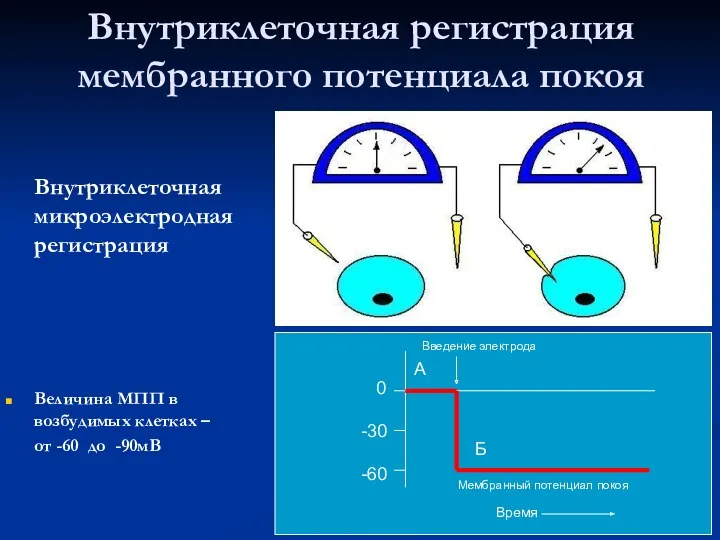
Внутриклеточная регистрация мембранного потенциала покоя
Внутриклеточная микроэлектродная регистрация
Величина МПП в возбудимых
Внутриклеточная регистрация мембранного потенциала покоя
Внутриклеточная микроэлектродная регистрация
Величина МПП в возбудимых
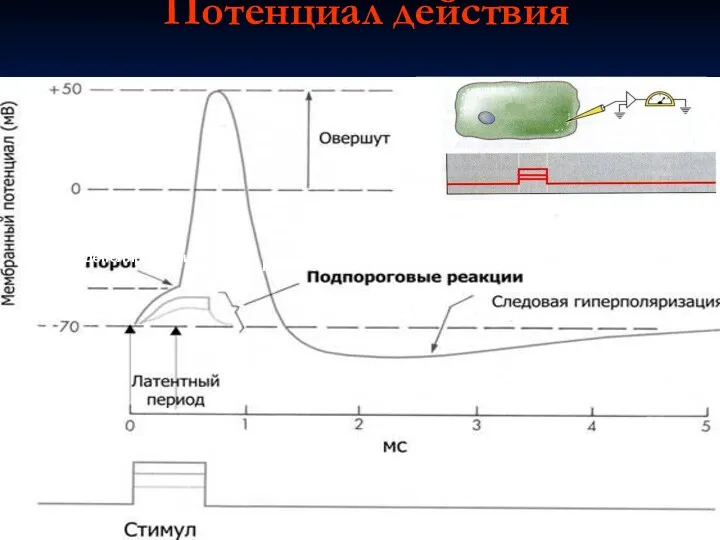
Потенциал действия
Фаза
деполяризации
Фаза
реполяризации
Раздражающий
импульс
Потенциал действия
Фаза
деполяризации
Фаза
реполяризации
Раздражающий
импульс

Вызывается сверхпороговым раздражением
Амплитуда не зависит от силы раздражения
Распространяется по всей мембране
Амплитуда не зависит от силы раздражения
Распространяется по всей мембране
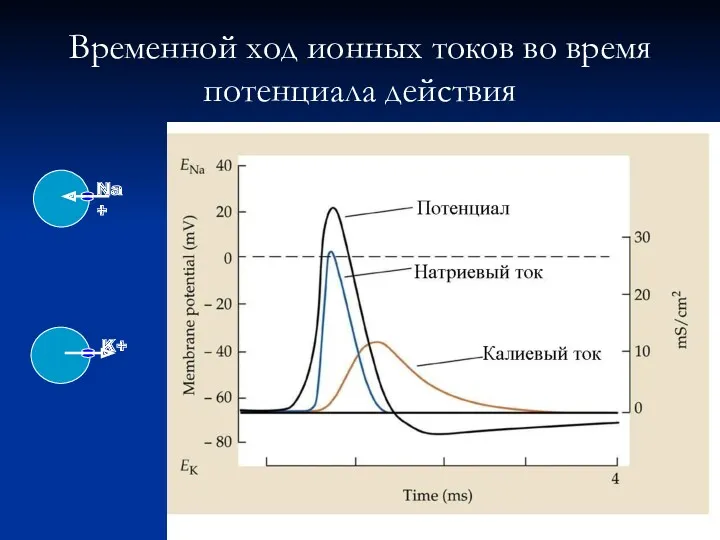
Временной ход ионных токов во время
потенциала действия
Временной ход ионных токов во время
потенциала действия
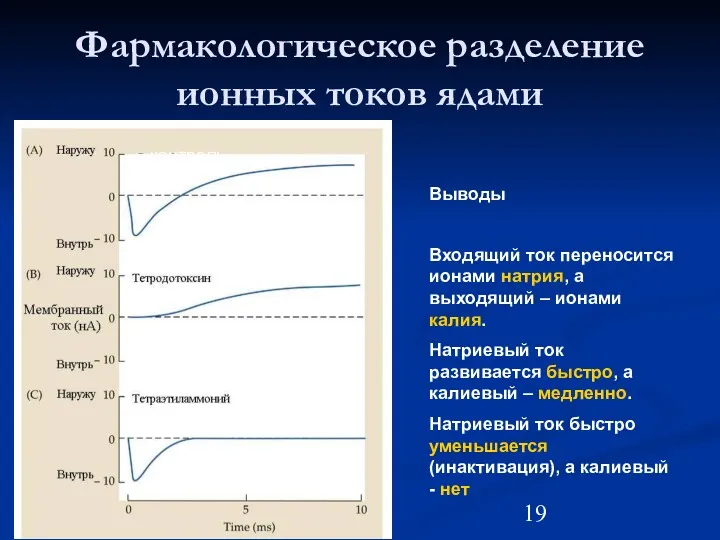
Фармакологическое разделение ионных токов ядами
контроль
Калиевый ток
Натриевый ток
Выводы
Входящий ток переносится ионами натрия,
Фармакологическое разделение ионных токов ядами
контроль
Калиевый ток
Натриевый ток
Выводы
Входящий ток переносится ионами натрия,
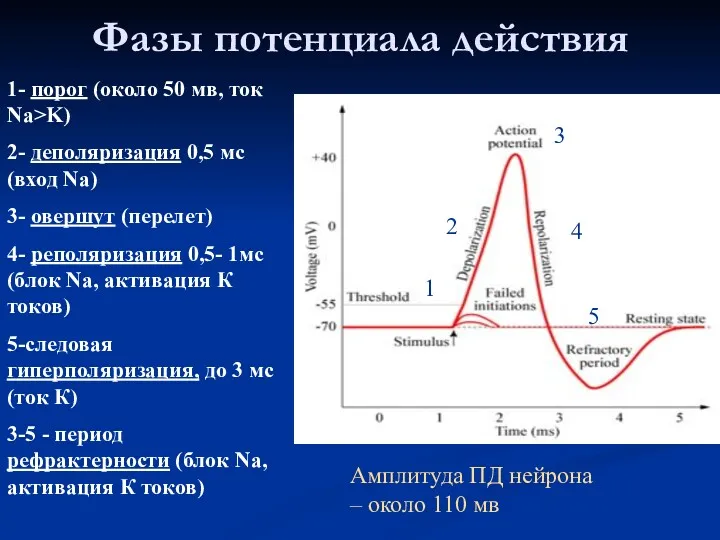
Фазы потенциала действия
1- порог (около 50 мв, ток Na>K)
2- деполяризация 0,5
Фазы потенциала действия
1- порог (около 50 мв, ток Na>K)
2- деполяризация 0,5
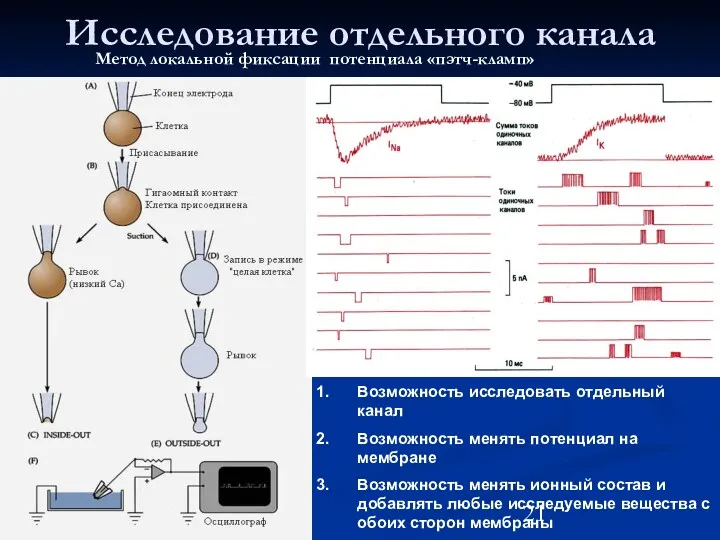
Исследование отдельного канала
Возможность исследовать отдельный канал
Возможность менять потенциал на мембране
Возможность менять
Исследование отдельного канала
Возможность исследовать отдельный канал
Возможность менять потенциал на мембране
Возможность менять

Нобелевская премия 1991 года в области физиологии и медицины
Эрвин Нейер и
Нобелевская премия 1991 года в области физиологии и медицины
Эрвин Нейер и
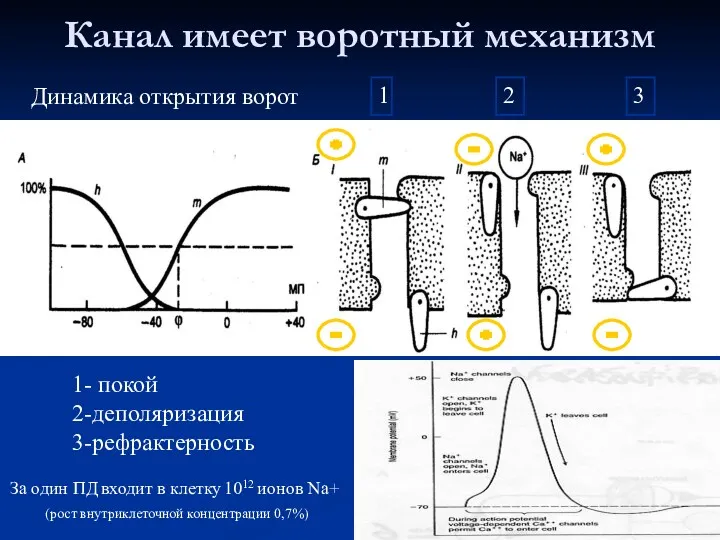
Канал имеет воротный механизм
1- покой
2-деполяризация
3-рефрактерность
Динамика открытия ворот
1
2
3
За один ПД входит в
Канал имеет воротный механизм
1- покой
2-деполяризация
3-рефрактерность
Динамика открытия ворот
1
2
3
За один ПД входит в
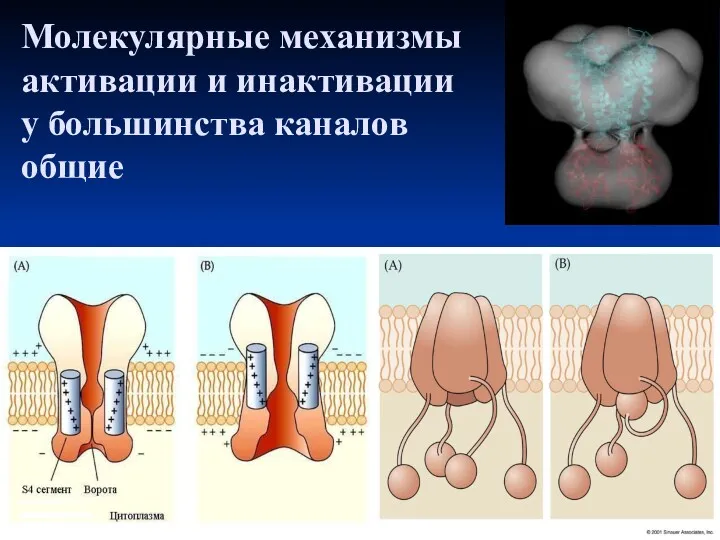
Молекулярные механизмы активации и инактивации
у большинства каналов
общие
Молекулярные механизмы активации и инактивации
у большинства каналов
общие

Работа Na+ канала
Работа Na+ канала
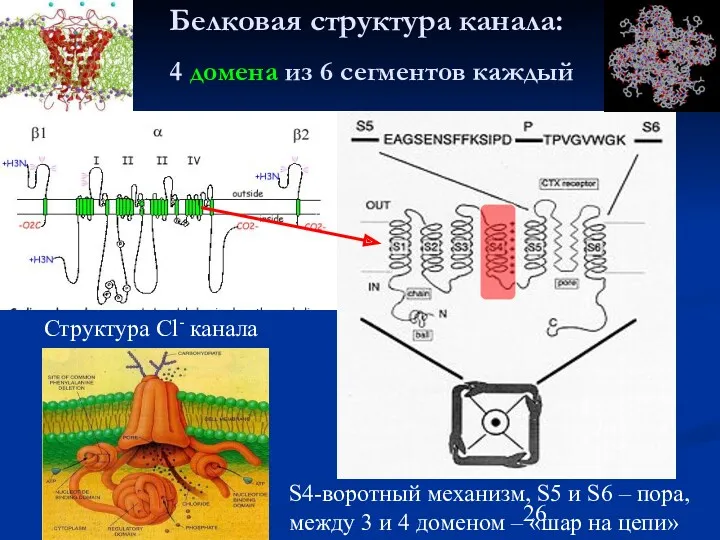
Белковая структура канала:
4 домена из 6 сегментов каждый
Структура Cl- канала
S4-воротный
Белковая структура канала:
4 домена из 6 сегментов каждый
Структура Cl- канала
S4-воротный
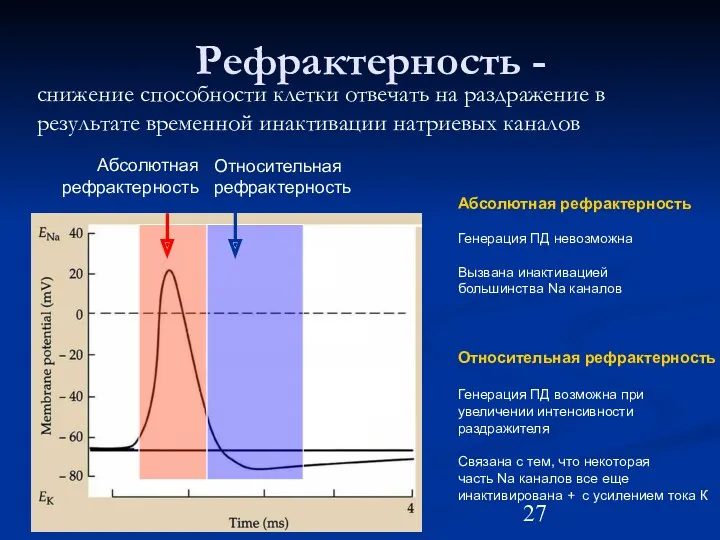
Рефрактерность -
снижение способности клетки отвечать на раздражение в результате временной
Рефрактерность -
снижение способности клетки отвечать на раздражение в результате временной
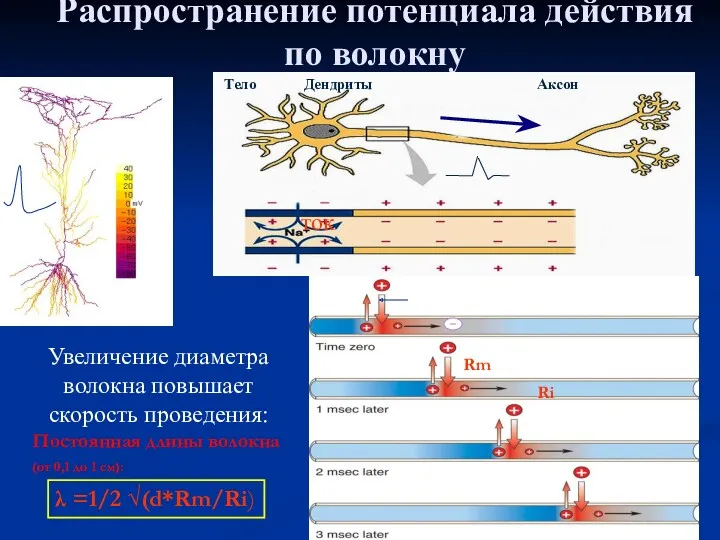
Распространение потенциала действия по волокну
Увеличение диаметра волокна повышает скорость проведения:
Постоянная длины
Распространение потенциала действия по волокну
Увеличение диаметра волокна повышает скорость проведения:
Постоянная длины
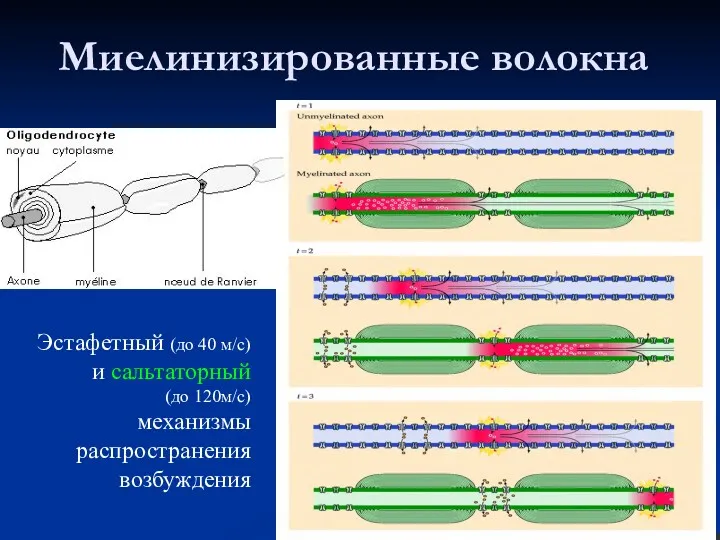
Миелинизированные волокна
Эстафетный (до 40 м/с) и сальтаторный
(до 120м/с)
механизмы распространения возбуждения
Миелинизированные волокна
Эстафетный (до 40 м/с) и сальтаторный
(до 120м/с)
механизмы распространения возбуждения
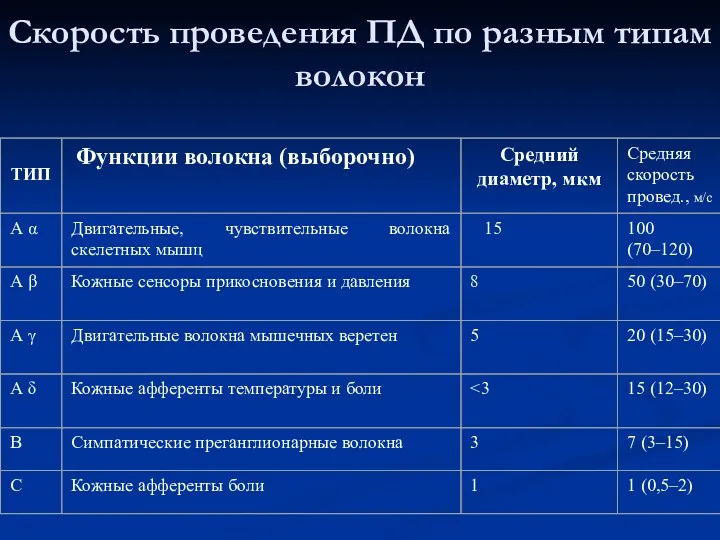
Скорость проведения ПД по разным типам волокон
Скорость проведения ПД по разным типам волокон
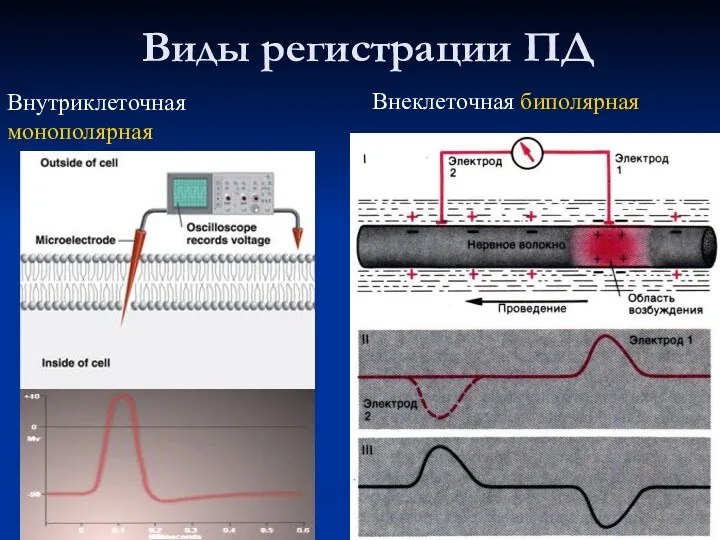
Виды регистрации ПД
Внутриклеточная монополярная
Внеклеточная биполярная
Виды регистрации ПД
Внутриклеточная монополярная
Внеклеточная биполярная
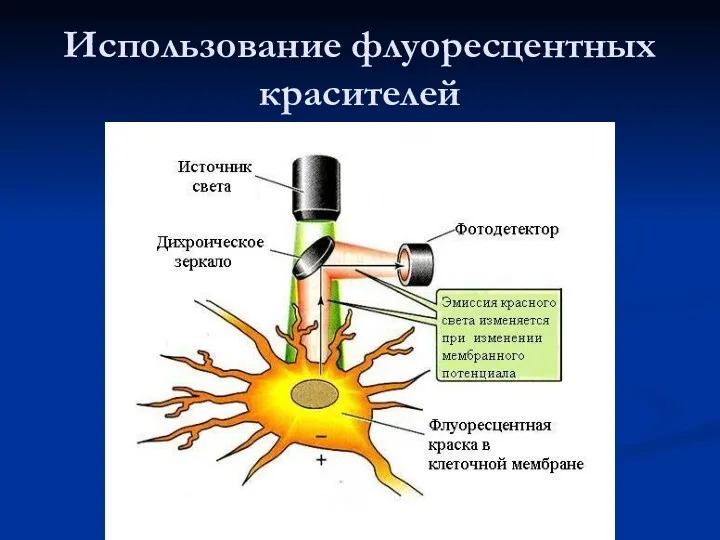
Использование флуоресцентных красителей
Использование флуоресцентных красителей
 Отряд Чешуйчатые. Ящерицы.
Отряд Чешуйчатые. Ящерицы. Introduction. Essential Cytology
Introduction. Essential Cytology Самые-самые Челябинской области
Самые-самые Челябинской области Огород здоровья
Огород здоровья Генетические аномалии человека
Генетические аномалии человека Биосинтез белка. Биосинтез углеводов
Биосинтез белка. Биосинтез углеводов Презентация к уроку биология 6 класс Устройство увеличительных приборов.
Презентация к уроку биология 6 класс Устройство увеличительных приборов. Методы и история развития биогеографии
Методы и история развития биогеографии Углеводы. Функции углеводов
Углеводы. Функции углеводов Будова клітинної мембрани. Хімічний склад. Транспортні властивості
Будова клітинної мембрани. Хімічний склад. Транспортні властивості Хоботные
Хоботные Интересные факты о кожном покрове и его производных
Интересные факты о кожном покрове и его производных Интерактивная игра-лабиринт Фотосинтез
Интерактивная игра-лабиринт Фотосинтез Физико-химические свойства крови
Физико-химические свойства крови Первая помощь при ушибах, вывихах, переломах
Первая помощь при ушибах, вывихах, переломах Популяционная генетика
Популяционная генетика Исследовательский практикум Цветение талой воды
Исследовательский практикум Цветение талой воды Повышение качества образования через нестандартные формы внеклассной работы
Повышение качества образования через нестандартные формы внеклассной работы 5a2a1377364c4c8bbec92a7e1c5f3f66
5a2a1377364c4c8bbec92a7e1c5f3f66 Загальна будова та функції нервової системи
Загальна будова та функції нервової системи Внутренняя среда организма. Кровь, тканевая жидкость и лимфа
Внутренняя среда организма. Кровь, тканевая жидкость и лимфа Сердечно – сосудистая система
Сердечно – сосудистая система Water as an Enviromental Factor
Water as an Enviromental Factor Sisteme parazitare
Sisteme parazitare Аюлар
Аюлар Презентация к уроку Вегетативное размножение растений 6 класс
Презентация к уроку Вегетативное размножение растений 6 класс Этапы развития жизни на Земле
Этапы развития жизни на Земле Лес и его голоса
Лес и его голоса