- Формирование осей зародыша дрозофилы и спецификация структур тела вдоль осей

Содержание
- 2. Гомейозисные (гомеотические) мутации впервые были описаны в 1894 Вильямом Бейтсоном (Willam Bateson) и характеризуются изменением места
- 3. Гомейозисная мутация гена Ultrabithorax у дрозофилы (подавление активности гена ultrabithorax) приводит к образованию второй пары крыльев
- 5. Эктопическая активность гомеотического гена Antennapedia (gain of function) в головном отделе (в норме активного во втором
- 6. Кластер гомеотических генов дрозофилы – Hom-C на 3-й хромосоме. Активность генов в зародыше и во взрослой
- 7. 8 гомеотических (гомейозисных) генов дрозофилы контролируют идентичность тела как у эмбриональной так и взрослой форм насекомого
- 8. Районы экспрессии гомеотических генов в бластодерме зародыша дрозофилы и несколькими часами позже в нервной системе (тёмные
- 9. Гомеотические ены Активности гомеотических генов инициируются продуктами экспрессии генов сегментации (gap и pair rule) и полярности
- 10. Гомеотические гены кодируют транскрипционные факторы -ТФ, содержащие ДНК- связывающий гомеодомен (60 а.о.), имеющий структуру - спираль-поворот-спираль-
- 11. Первоначальную экспрессию кластера гомеотических генов инициируют сегментационные гены: gap-гены и гены pair-rule. В дальнейшем за экспрессию
- 12. Имагинальные диски позиционированы в отношении сегментов личинки, что объясняет корреляцию сегментов личинки и взрослой мухи
- 14. Карта судьбы имагинальных дисков и гистобластов дрозофилы, отвечает за спецификацию структур, формирующихся из них.
- 15. Кластер гомеотических генов дрозофилы - Hom-C. Активность генов в зародыше и во взрослой особи. Последовательное расположение
- 16. Кластеры гомеотических генов высоко консервативны. Они сформированы в явном виде у Bilateria и присутствуют в организмах
- 17. Ортологичные гомеотические гены у всех животных (включая и саму дрозофилу) обозначаются сейчас как гены семейства Hox
- 18. У позвоночных выявляется не менее четырех кластеров паралогичных гомеотических генов. Число генов в Hox – кластерах
- 19. Гены гомеотических кластеров Hox-C мыши и HOX-C человека контролируют метамерию заднего отдела головного мозга и формирование
- 20. Гомеотические гены в кластере работают в гнездовом режиме. Кластеры гомеотических генов Hox-C мыши и HOX-C человека
- 21. Использование консерватизма гомеотических генов в молекулярном анализе филогенетических связей организмов. Поправка существующих представлений о филогенетических взаимоотношениях
- 22. Предполагаемая макроэволюционная роль гомейозисных мутаций - вклад биологии развития в эволюционную теорию. Однократная мутация одного или
- 23. Экспрессия гена Distall-less важна для образования конечностей у насекомых, в том числе и для образования ложных
- 24. Отсутствие экспрессии гомеотических генов Abd A и Ubx даёт возможность гену Distall-less экспрессироваться (маркировка красным цветом)
- 25. Формирование дорсо-вентральной оси у Drosophila. Дорсо-вентральная полярность зародыша формируется позднее передне-задней, хотя за неё также отвечают
- 26. Three general classes of genes specify the body plan: 1. Maternal effect genes (~33). 2. Segmentation
- 27. Начальный этап дорсолизации (проходит до оплодотворения) - миграция ядра ооцита из постериорного района ближе к трофоцитам
- 28. Градиент Gurken (белок, родственный эпидермальному ростовому фактору- EGF) в дорсальной части мембраны ооцита воспринимается фолликулярными клетками
- 29. Комплекс взаимодействий Gurken (ооцит)- Torpedo (фолликулярные клетки) приводит к поляризации клеток фолликулярного эпителия на дорсальные и
- 30. Сигнал из вентральных фолликулярных клеток в цитоплазму зародыша. События происходят в перивителлиновом пространстве уже после оплодотворения.
- 31. Гаструляция и дальнейшее развитие зародыша дрозофилы
- 32. Figure 21-52. Fate map of a Drosophila embryo at the cellular blastoderm stage. The embryo is
- 33. Начало гаструляции у Drosophila:16 наиболее вентральных клеток мигрируют внутрь зародыша и образуют в дальнейшем мезодерму (регистрируется
- 34. Схематическое представление гаструляции
- 35. Действие градиента Dorsal определяет дорсо-вентральную полярность в ходе гаструляции. Dorsal – транскрипционный фактор, по разному регулирует
- 36. Действие градиента Dorsal определяет дорсо-вентральную полярность в ходе гаструляции. Dorsal – транскрипционный фактор, по разному регулирует
- 37. Консерватизм сигнальной цепочки, действующей через Toll- рецепторы у дрозофилы и млекопитающих (эффект коопции). Сигнальная цепочка, действующая
- 38. Использование гомологичных компонентов сигнальной цепочки, включая ТФ: Dorsal/Dif /Relish, в иммунных реакциях у дрозофилы (антифунгальный-Relish и
- 40. Скачать презентацию

Гомейозисные (гомеотические) мутации впервые были описаны в 1894 Вильямом Бейтсоном (Willam
Гомейозисные (гомеотические) мутации впервые были описаны в 1894 Вильямом Бейтсоном (Willam
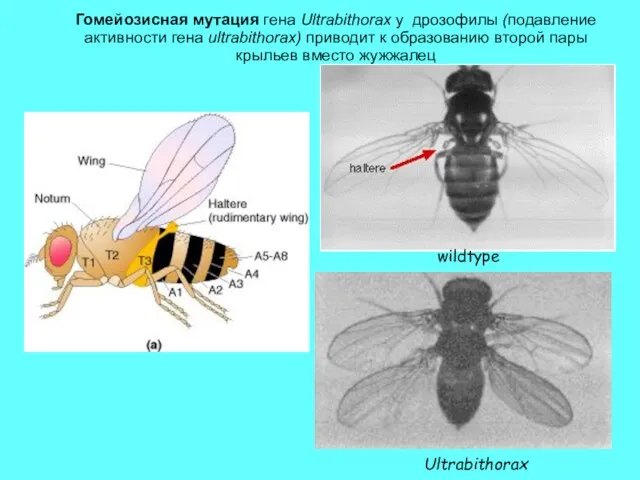
Гомейозисная мутация гена Ultrabithorax у дрозофилы (подавление активности гена ultrabithorax) приводит
Гомейозисная мутация гена Ultrabithorax у дрозофилы (подавление активности гена ultrabithorax) приводит


Эктопическая активность гомеотического гена Antennapedia (gain of function) в головном отделе
Эктопическая активность гомеотического гена Antennapedia (gain of function) в головном отделе
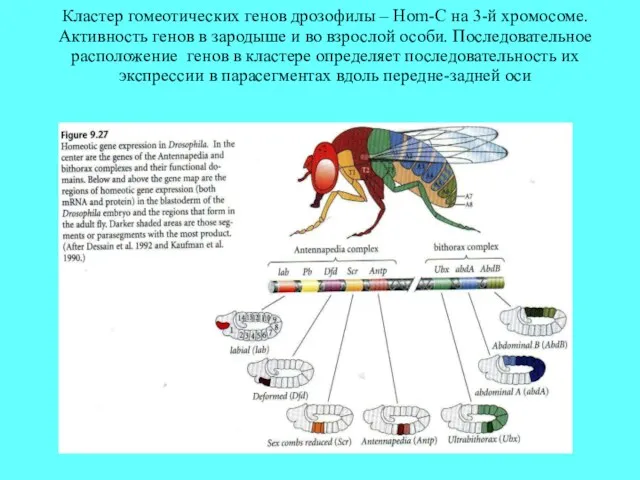
Кластер гомеотических генов дрозофилы – Hom-C на 3-й хромосоме. Активность генов
Кластер гомеотических генов дрозофилы – Hom-C на 3-й хромосоме. Активность генов

8 гомеотических (гомейозисных) генов дрозофилы контролируют идентичность тела как у эмбриональной
8 гомеотических (гомейозисных) генов дрозофилы контролируют идентичность тела как у эмбриональной

Районы экспрессии гомеотических генов в бластодерме зародыша дрозофилы и несколькими часами
Районы экспрессии гомеотических генов в бластодерме зародыша дрозофилы и несколькими часами
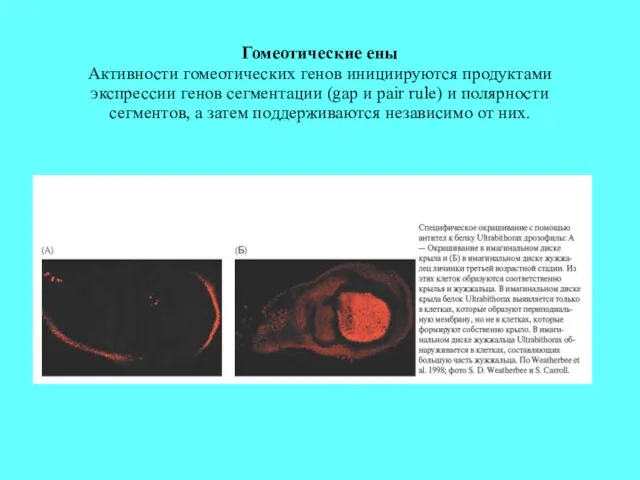
Гомеотические ены
Активности гомеотических генов инициируются продуктами экспрессии генов сегментации (gap и
Гомеотические ены Активности гомеотических генов инициируются продуктами экспрессии генов сегментации (gap и
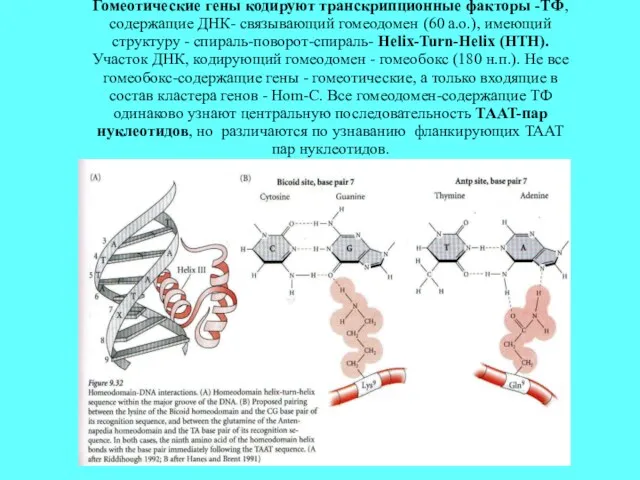
Гомеотические гены кодируют транскрипционные факторы -ТФ, содержащие ДНК- связывающий гомеодомен (60
Гомеотические гены кодируют транскрипционные факторы -ТФ, содержащие ДНК- связывающий гомеодомен (60

Первоначальную экспрессию кластера гомеотических генов инициируют сегментационные гены: gap-гены и гены
Первоначальную экспрессию кластера гомеотических генов инициируют сегментационные гены: gap-гены и гены
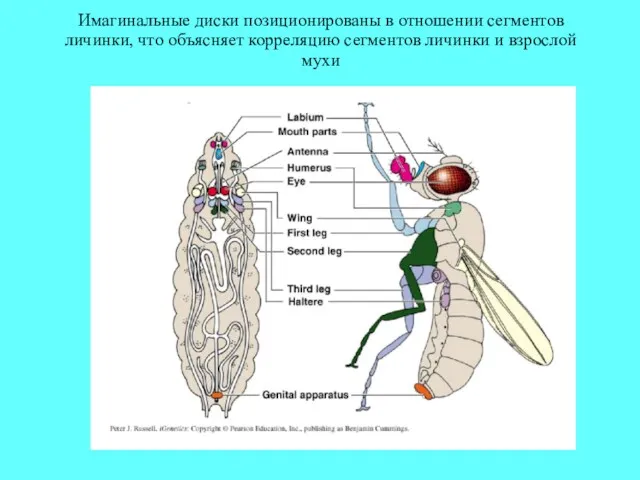
Имагинальные диски позиционированы в отношении сегментов личинки, что объясняет корреляцию сегментов
Имагинальные диски позиционированы в отношении сегментов личинки, что объясняет корреляцию сегментов
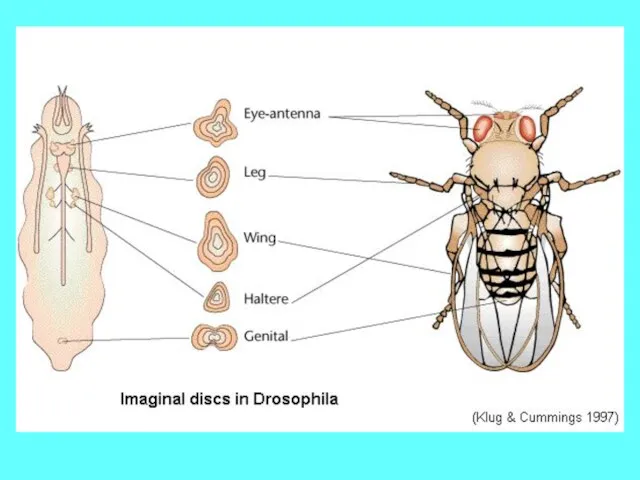
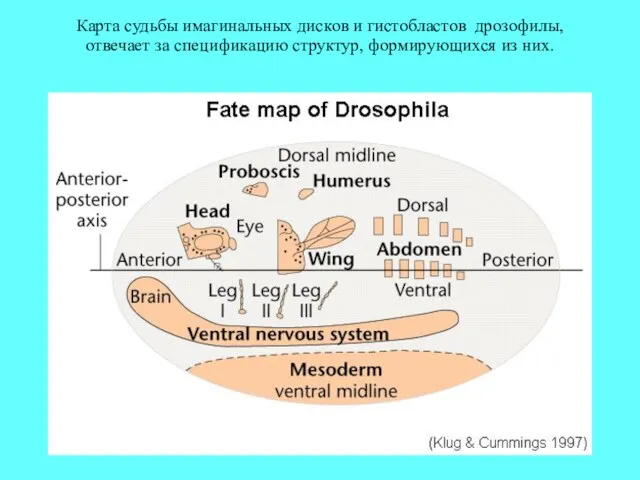
Карта судьбы имагинальных дисков и гистобластов дрозофилы, отвечает за спецификацию структур,
Карта судьбы имагинальных дисков и гистобластов дрозофилы, отвечает за спецификацию структур,
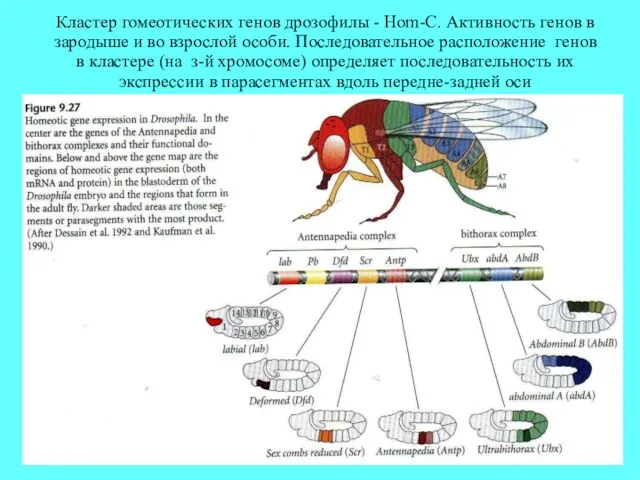
Кластер гомеотических генов дрозофилы - Hom-C. Активность генов в зародыше и
Кластер гомеотических генов дрозофилы - Hom-C. Активность генов в зародыше и
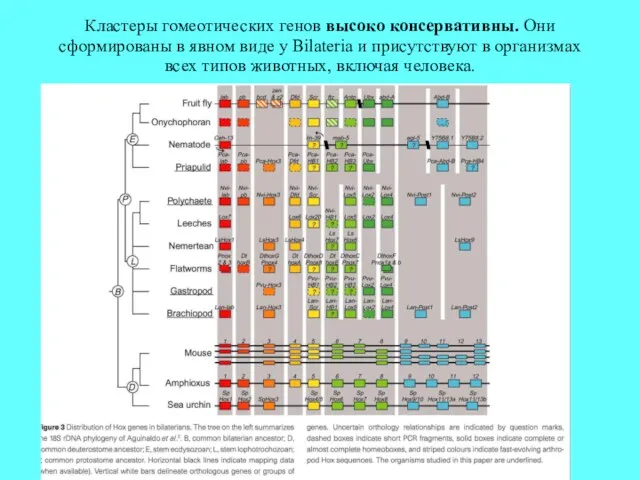
Кластеры гомеотических генов высоко консервативны. Они сформированы в явном виде у
Кластеры гомеотических генов высоко консервативны. Они сформированы в явном виде у

Ортологичные гомеотические гены у всех животных (включая и саму дрозофилу) обозначаются

У позвоночных выявляется не менее четырех кластеров паралогичных гомеотических генов. Число
У позвоночных выявляется не менее четырех кластеров паралогичных гомеотических генов. Число
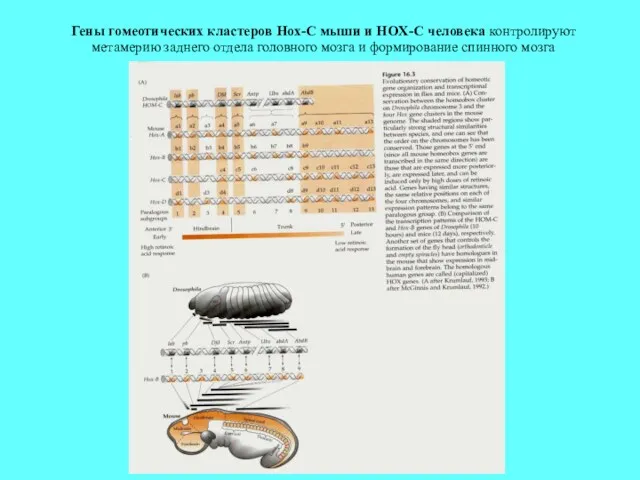
Гены гомеотических кластеров Hox-C мыши и HOX-C человека контролируют метамерию заднего
Гены гомеотических кластеров Hox-C мыши и HOX-C человека контролируют метамерию заднего
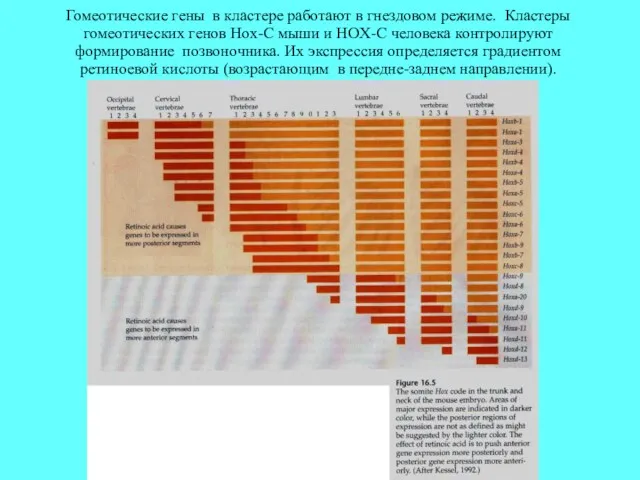
Гомеотические гены в кластере работают в гнездовом режиме. Кластеры гомеотических генов
Гомеотические гены в кластере работают в гнездовом режиме. Кластеры гомеотических генов
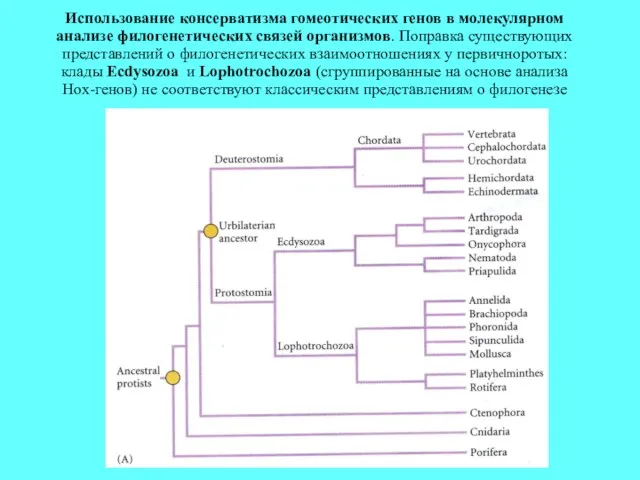
Использование консерватизма гомеотических генов в молекулярном анализе филогенетических связей организмов. Поправка
Использование консерватизма гомеотических генов в молекулярном анализе филогенетических связей организмов. Поправка

Предполагаемая макроэволюционная роль гомейозисных мутаций - вклад биологии развития в эволюционную
Предполагаемая макроэволюционная роль гомейозисных мутаций - вклад биологии развития в эволюционную
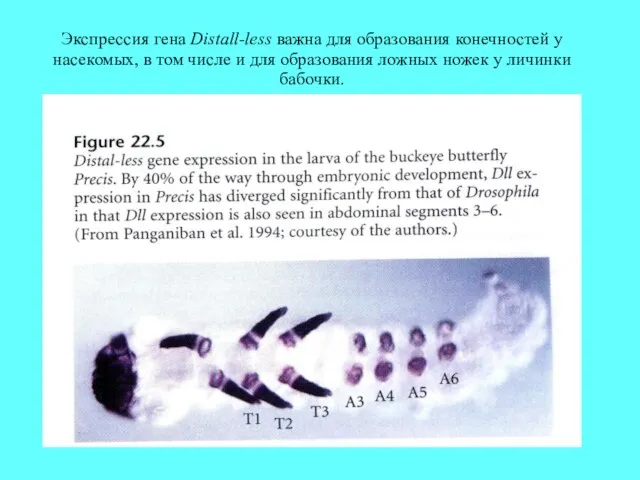
Экспрессия гена Distall-less важна для образования конечностей у насекомых, в том
Экспрессия гена Distall-less важна для образования конечностей у насекомых, в том
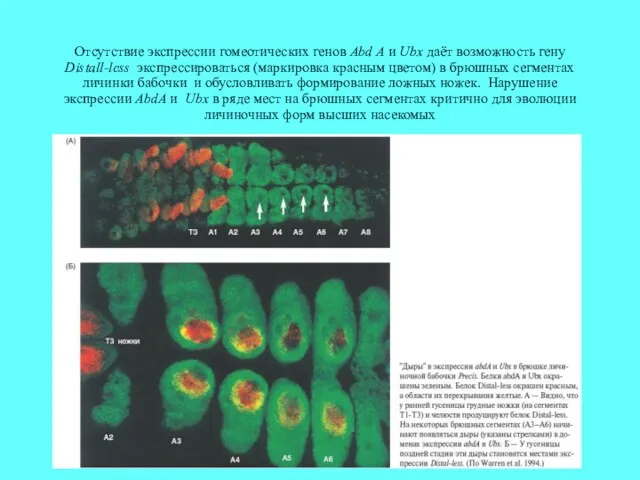
Отсутствие экспрессии гомеотических генов Abd A и Ubx даёт возможность гену
Отсутствие экспрессии гомеотических генов Abd A и Ubx даёт возможность гену

Формирование дорсо-вентральной оси у Drosophila.
Дорсо-вентральная полярность зародыша формируется позднее передне-задней, хотя
Формирование дорсо-вентральной оси у Drosophila. Дорсо-вентральная полярность зародыша формируется позднее передне-задней, хотя

Three general classes of genes specify the body plan:
1. Maternal effect
1. Maternal effect
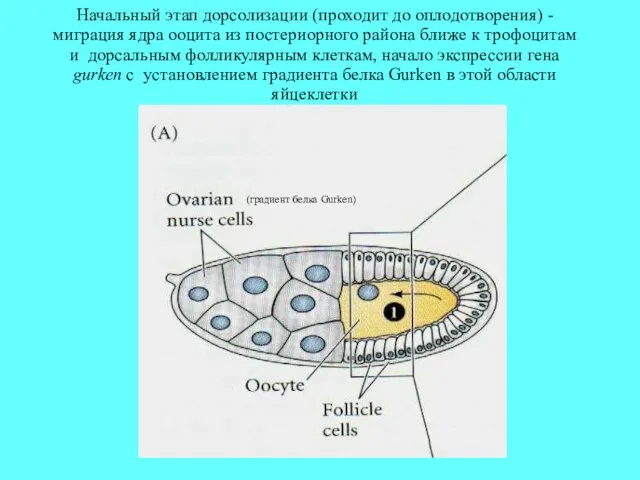
Начальный этап дорсолизации (проходит до оплодотворения) - миграция ядра ооцита из
Начальный этап дорсолизации (проходит до оплодотворения) - миграция ядра ооцита из
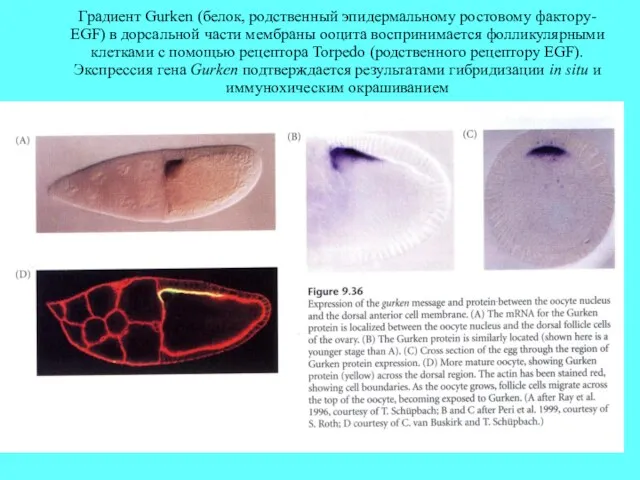
Градиент Gurken (белок, родственный эпидермальному ростовому фактору- EGF) в дорсальной части
Градиент Gurken (белок, родственный эпидермальному ростовому фактору- EGF) в дорсальной части
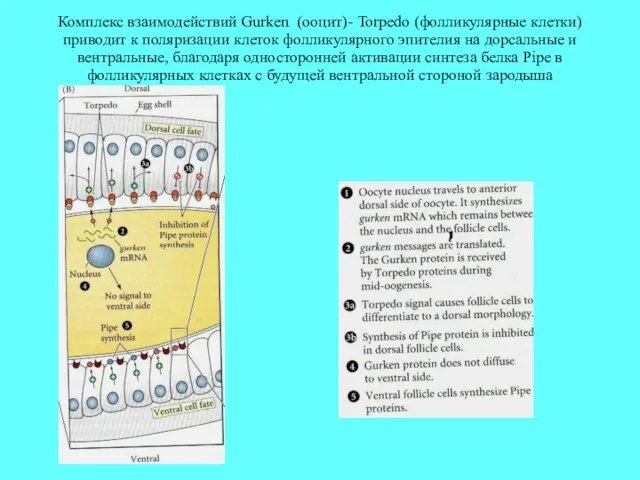
Комплекс взаимодействий Gurken (ооцит)- Torpedo (фолликулярные клетки) приводит к поляризации клеток
Комплекс взаимодействий Gurken (ооцит)- Torpedo (фолликулярные клетки) приводит к поляризации клеток

Сигнал из вентральных фолликулярных клеток в цитоплазму зародыша. События происходят в
Сигнал из вентральных фолликулярных клеток в цитоплазму зародыша. События происходят в
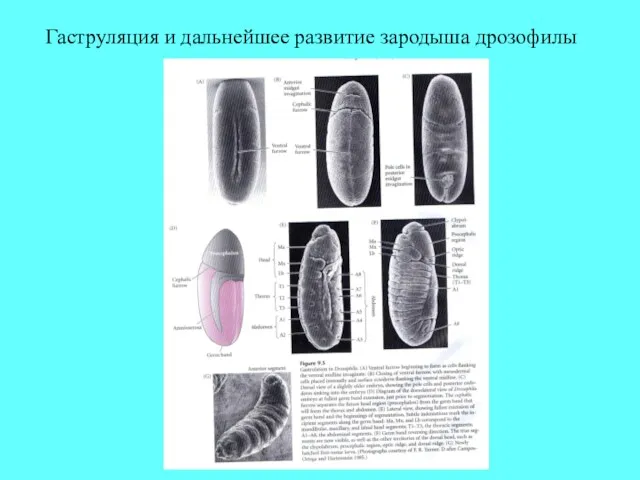
Гаструляция и дальнейшее развитие зародыша дрозофилы
Гаструляция и дальнейшее развитие зародыша дрозофилы
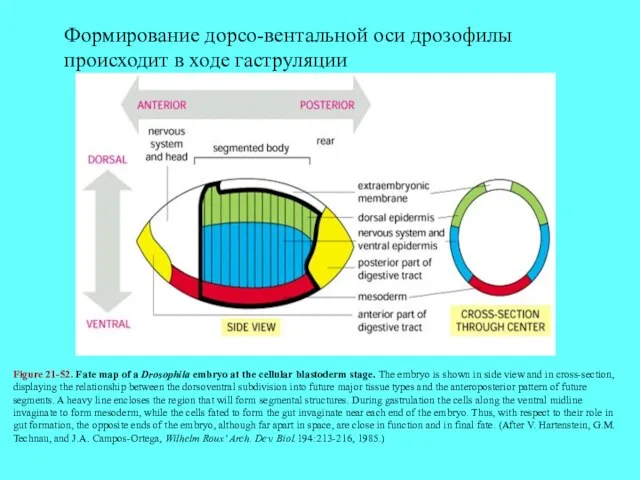
Figure 21-52. Fate map of a Drosophila embryo at the cellular
Figure 21-52. Fate map of a Drosophila embryo at the cellular
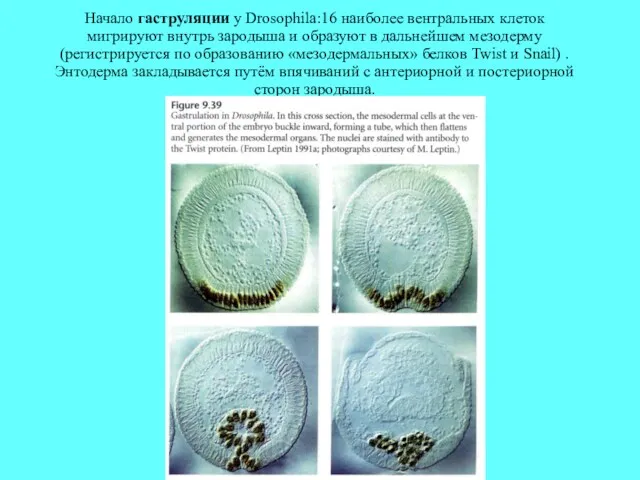
Начало гаструляции у Drosophila:16 наиболее вентральных клеток мигрируют внутрь зародыша и
Начало гаструляции у Drosophila:16 наиболее вентральных клеток мигрируют внутрь зародыша и
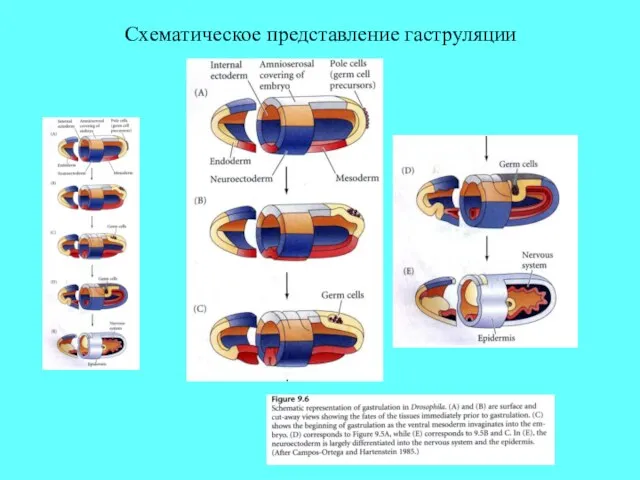
Схематическое представление гаструляции
Схематическое представление гаструляции
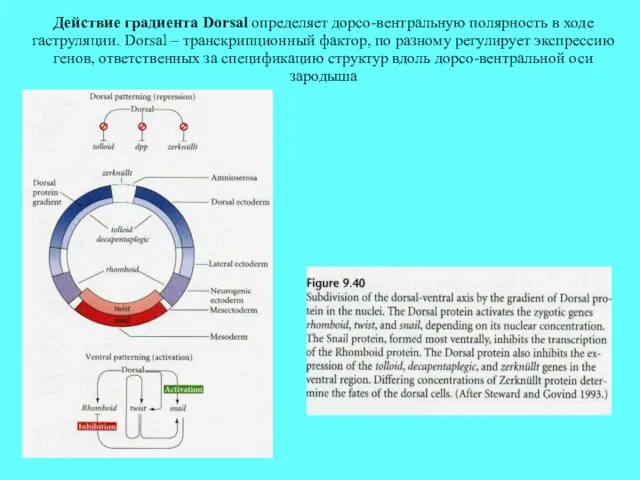
Действие градиента Dorsal определяет дорсо-вентральную полярность в ходе гаструляции. Dorsal –
Действие градиента Dorsal определяет дорсо-вентральную полярность в ходе гаструляции. Dorsal –
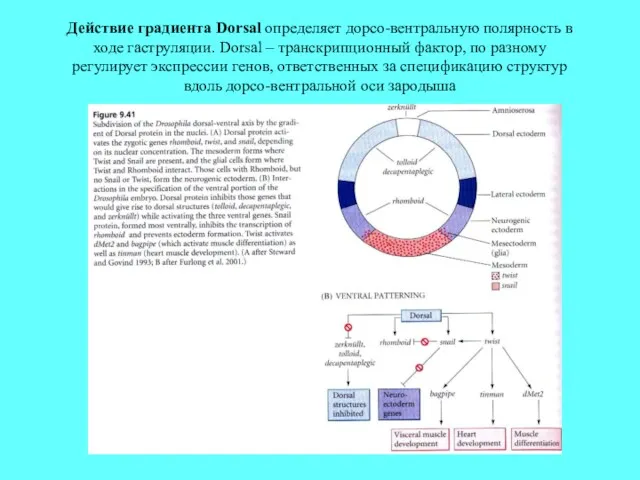
Действие градиента Dorsal определяет дорсо-вентральную полярность в ходе гаструляции. Dorsal –
Действие градиента Dorsal определяет дорсо-вентральную полярность в ходе гаструляции. Dorsal –
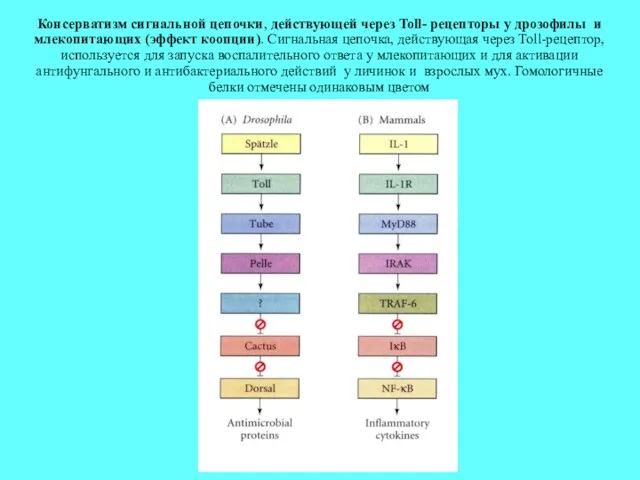
Консерватизм сигнальной цепочки, действующей через Toll- рецепторы у дрозофилы и млекопитающих
Консерватизм сигнальной цепочки, действующей через Toll- рецепторы у дрозофилы и млекопитающих
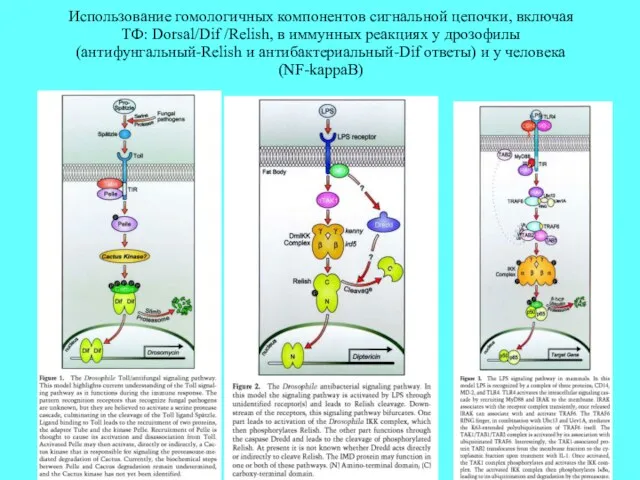
Использование гомологичных компонентов сигнальной цепочки, включая ТФ: Dorsal/Dif /Relish, в иммунных
Использование гомологичных компонентов сигнальной цепочки, включая ТФ: Dorsal/Dif /Relish, в иммунных
 Дигибридтік және полигибридтік будандастыру
Дигибридтік және полигибридтік будандастыру Методы цитологии. Клеточная теория
Методы цитологии. Клеточная теория Основы агрономии. Введение
Основы агрономии. Введение Работа над ошибками при подготовке к ЕГЭ. Решение генетических задач
Работа над ошибками при подготовке к ЕГЭ. Решение генетических задач les systeme de lorganisme
les systeme de lorganisme Внутреннее строение лягушки
Внутреннее строение лягушки Простейшие, или эукариоты. (5 класс)
Простейшие, или эукариоты. (5 класс) Тип членистоногие
Тип членистоногие Властивості та характеристики екосистем. Типи зв'язків між популяціями в екосистемі
Властивості та характеристики екосистем. Типи зв'язків між популяціями в екосистемі Презентация урока 5 класс ФГОС.
Презентация урока 5 класс ФГОС. Чувствительность. Рецепторы
Чувствительность. Рецепторы Оценка качества почвы с помощью растения – биоиндикатора
Оценка качества почвы с помощью растения – биоиндикатора Анатомия опорно-двигательного аппарата
Анатомия опорно-двигательного аппарата Стрекозы
Стрекозы Рост и развитие растений
Рост и развитие растений Биомембраналар
Биомембраналар Гербарии растений, содержащих дубильные вещества
Гербарии растений, содержащих дубильные вещества Строение клетки. Тест - тренажер
Строение клетки. Тест - тренажер Лишайники. Среда обитания лишайников
Лишайники. Среда обитания лишайников Пигментация волос
Пигментация волос Дәм сезу анализаторы,оның бөлімдері
Дәм сезу анализаторы,оның бөлімдері Аффинная хроматография
Аффинная хроматография Витамины: Мал золотник, да дорог
Витамины: Мал золотник, да дорог Дистанционное обучение. Презентация: Наследственные болезни.
Дистанционное обучение. Презентация: Наследственные болезни. Обмен белков
Обмен белков Видообразование как результат микроэволюции
Видообразование как результат микроэволюции Размножение голосеменных растений
Размножение голосеменных растений Экологическая ниша
Экологическая ниша