- Функции центральной нервной системы

Содержание
- 2. Функции ЦНС Управление деятельностью опорно-двигательного аппарата. Регуляция работы внутренних органов осуществляется посредством ВНС и эндокринными железами.
- 3. Эти сложнейшие и жизненно важные задачи решаются с помощью нервных клеток (нейронов), специализированных на 1) восприятии,
- 4. Нейронная теория Синапс В основе современного представления о структуре и функции ЦНС лежит нейронная теория, которая
- 5. Нобелевская премия по физиологии и медицине 1906 г. Камило Гольджи (1844-1926) - профессор гистологии и общей
- 6. Окрашенная по Гольджи нервная ткань из зрительной коры крысы зарисована Рамон-и-Кахалом в 1888 г. Одним из
- 7. Функции клеток ЦНС и ликвора А. Нервная клетка (нейрон): Сома (тело) Отростки: дендриты, аксон Функции нейрона:
- 8. Нейрон Перикарион (сома) - Отростки нейрона - тело нейрона это вырост от тела нейрона дендрит аксон
- 9. Функциональные структуры нейрона: Структура, обеспечивающая синтез макромолекул, которые транспортируются по аксону и дендритам – сома (тело
- 10. НЕЙРОН. СТРОЕНИЕ
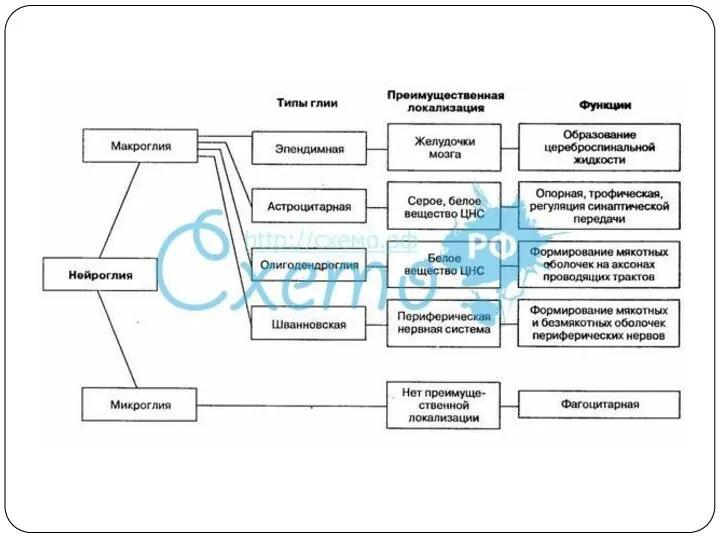
- 11. Б. Глиальные клетки Нейроглия – «нервный клей» 50% объема ЦНС 14*1010 Функции: Опорная Защитная Изолирующая Обменная
- 13. В. Ликвор (цереброспинальная жидкость) 90% воды 10% сухого остатка (2% - орг. в-ва, 8% - неорг.
- 14. Классификация нейронов 1.В зависимости от отдела ЦНС: Нейроны соматической НС Нейроны вегетативной НС 2. По источнику
- 15. Классификация нейронов 6. По морфологии: Униполярные Биполярные Мультиполярные Ложноуниполярные Биполярный нейрон – имеет два отростка –
- 16. 7. По выполняемым функциям: Рецепторные аффекторные, афферентные Вставочные, ассоциативные Двигательные, эффекторные, эфферентные. Двигательный мотонейрон, иннервирующий мышцу
- 17. Нервный отросток – вырост от тела нейрона (дендрит или аксон). Нервное волокно – это отросток нервной
- 18. Стадии образования миелиновой оболочки Шванновской клеткой
- 19. СИНАПСЫ 3 части: Пресинаптическое окончание Постсинаптическое окончание Синаптическая щель
- 20. Пресинапс представляет из себя расширенные концы терминалей аксона (бляшка). Мембрана бляшек называется пресинаптической мембраной (имеет такое
- 21. Синаптическая щель Постсинапс Постсинаптическая мембрана содержит большое количество белков-рецепторов с высоким сродством к медиатору. Синаптическая щель
- 22. Этапы синаптической передачи (трансмиссии) 1) Синтез медиатора 2) Транспорт медиатора 3) Накопление медиатора 4) Высвобождение медиатора
- 23. Этапы синаптической передачи (трансмиссии)
- 25. СИНАПСЫ. ВИДЫ. По знаку действия Возбуждающие Медиатор Ацетилхолин Тормозные Медиатор Гамма-аминомасляная кислота, глицин
- 26. СИНАПСЫ. ВИДЫ. По локализации: Нервно-мышечный Нейро –нейрональный аксосоматические аксодендритические Аксо-аксональные
- 27. Синапсы. виды По способу передачи сигнала Электрические Химические Электро-химические
- 28. В синапсах с химическим механизмом передачи продолжительность синаптической задержки у теплокровных составляет 0,2—0,5 мс. В электрических
- 29. Медиаторы и рецепторы ЦНС А. Ацетилхолин Б. Амины (дофамин, норадреналин, серотонин, гистамин) В. Аминокислоты(глицин, Гамма-аминомасляная кислота)
- 30. ПЛАН: Рефлекс. Виды рефлексов Рефлекторная дуга Торможение, виды. Тема: РЕФЛЕКТОРНАЯ ДЕЯТЕЛЬНОСТЬ ЦНС
- 31. ЦНС осуществляет две функции рефлекторную и проводниковую. Рефлекторная деятельность осуществляется за счет рефлексов. РЕФЛЕКС – реакция
- 32. В естественных условиях рефлекторная реакция происходит при пороговом и надпороговом раздражении входа рефлекторной дуги – рецептивного
- 33. Структурной основой рефлекса является рефлекторная дуга – последовательно соединенная цепочка нервных клеток
- 34. ВРЕМЯ РЕФЛЕКСА - это время, необходимое для осуществления рефлекса, складывается из 5 компонентов: - латентный период
- 35. КЛАССИФИКАЦИЯ РЕФЛЕКСОВ 1. По способу вызывания: - безусловные рефлексы - условные 2. По месту расположения рецептора:
- 36. 3. В зависимости от расположения центров рефлексов уровня активации части мозга: - спинномозговые - бульбарные -
- 37. 4. По биологическому значению - пищевые - оборонительные - половые и др. 5. По характеру ответной
- 38. Рефлекторная деятельность организма во многом определяется общими свойствами нервных центров. Нервный центр – «ансамбль» нейронов, согласованно
- 39. Классификация нервных центров Морфологический критерий (локализация в отделах ЦНС ): Спинальные центры (в спинном мозге); Бульбарные
- 40. Нервные центры В основе нервной деятельности лежат активные и противоположные по своим функциональным свойствам процессы: Возбуждение;
- 41. ОСОБЕННОСТИ РАСПРОСТРАНЕНИЯ ВОЗБУЖДЕНИЯ В ЦНС: В ЦНС, внутри рефлекторной дуги и нейронных цепей возбуждение идет, как
- 42. 2.замедленное проведение Известно, что возбуждение по нервным волокнам (периферия) проводится быстро, а в ЦНС– относительно медленно
- 43. 3. суммация Это свойство впервые описал И.М. Сеченов (1863): При действии ряда подпороговых стимулов на рецептор
- 44. временная суммация А.. В ответ на одиночный раздражитель возникает синаптический ток (затененная область) и синаптический потенциал
- 45. пространственная суммация два или несколько подпороговых импульсов приходят в ЦНС по разным афферентным путям и вызывают
- 46. 4.окклюзия Феномен окклюзии ( Причина феномена – перекрытие путей на вставочных или эфферентных нейронах благодаря конвергенции.
- 47. 5. последействие Причина: Длительное последействие связано с наличием в ЦНС кольцевых связей между нейронами Структурная основа
- 48. 6. проторение (постактивационное облегчение) Проторение (постактивационное облегчение): После возбуждения, вызванного ритмической стимуляцией, последующий стимул вызывает больший
- 49. Свойства нервных центров: Длительное повторное раздражение рецептивного поля рефлекса →ослабление рефлекторной реакции вплоть до полного исчезновения
- 50. На нейронах в ЦНС сходятся синаптические влияния разного функционального значения. Это приводит к трансформации ритма поступающих
- 51. Обусловлена высокой интенсивность обменных процессов: 100 г нервной ткани (головной мозг собаки) использует О2 в 22
- 52. Пластичность – функциональная подвижность нервного центра: возможность его включения в регуляцию различных функций. Тонус – наличие
- 53. Торможение в ЦНС (И.М. Сеченов), его виды и роль. Современное представление о механизмах центрального торможения. Тормозные
- 54. ТОРМОЖЕНИЕ В ЦНС Интегративная деятельность в ЦНС осуществляется при участии возбуждающих и тормозных процессов. Явление торможения
- 55. Современная трактовка опыта И. М. Сеченова (И. М. Сеченов раздражал ретикулярную формацию ствола мозга): возбуждение ретикулярной
- 56. Выделяют два типа торможения: первичное. Для его возникновения необходимо наличие специальных тормозных нейронов. Торможение возникает первично
- 57. Постсинаптическое торможение (лат. post позади, после чего-либо + греч. sinapsis соприкосновение, соединение) - нервный процесс, обусловленный
- 58. Разновидности постсинаптического торможения Возвратное (А): ТН действуют на те же нейроны, которые их активируют. Поэтому развивающееся
- 60. Разновидности постсинаптического торможения (продолжение) 3. Латеральное: ТН активирются импульсами от возбужденного центра и влияют на соседние
- 61. Разновидности постсинаптического торможения (продолжение) 4. Прямое торможение (реципрокное): Оно вызывает угнетение центра антогониста. Обеспечивает согласованную работу
- 62. Пресинаптическое торможение Развивается на пресинаптической мембране. Осуществляется посредством специальных тормозных нейронов. Его структурной основой являются аксо-аксональные
- 63. Передача возбуждения в синапсе на фоне пресинаптического торможения
- 64. Конвергенция «возбуждающего возбуждения» и «тормозящего возбуждения» на теле нейрона
- 65. Виды вторичного торможения: запредельное, возникающее при большом потоке информации, поступающей в клетку. Поток информации лежит за
- 66. Значение торможения Торможение выполняет охранительную роль, его отсутствие привело бы к истощению медиаторов и прекращению деятельности
- 67. Принципы координации нервных центров Координация (лат. сo - вместе+ ordinatio – расположение в порядке) – согласование
- 68. Принципы координации нервных центров: «общий конечный путь» (конвергенция) Выдвинут Ч.С. Шеррингтоном в 1906 г. Конвергенция –
- 69. Принципы координации нервных центров: «общий конечный путь» Согласно этому принципу к одному мотонейрону приходит множество импульсов
- 70. Принципы координации нервных центров: доминанта Принцип доминанты (лат. dominare господствовать) – установлен А. А. Ухтомским (1923).
- 71. Доминанта Свойства доминантного очага: Повышенная возбудимость; Инерционность; Способность к суммации; Способность к торможению центров, функционально несовместимых
- 72. Доминанта А.А. Ухтомский о (+) и (–) доминанты: «… Доминанта, как общая формула, ещё ничего не
- 73. Принципы координации нервных центров: иерархия и субординация В ЦНС имеют место: Иерархические взаимоотношения (греч. hierarchia Субординация
- 74. Принципы координации нервных центров: иррадиация Иррадиация (лат. irradio освещать, озарять) – распространение процессов возбуждения (торможения). Иррадиация
- 76. Скачать презентацию
Функции ЦНС
Управление деятельностью опорно-двигательного аппарата.
Регуляция работы внутренних органов осуществляется посредством ВНС
Функции ЦНС
Управление деятельностью опорно-двигательного аппарата.
Регуляция работы внутренних органов осуществляется посредством ВНС
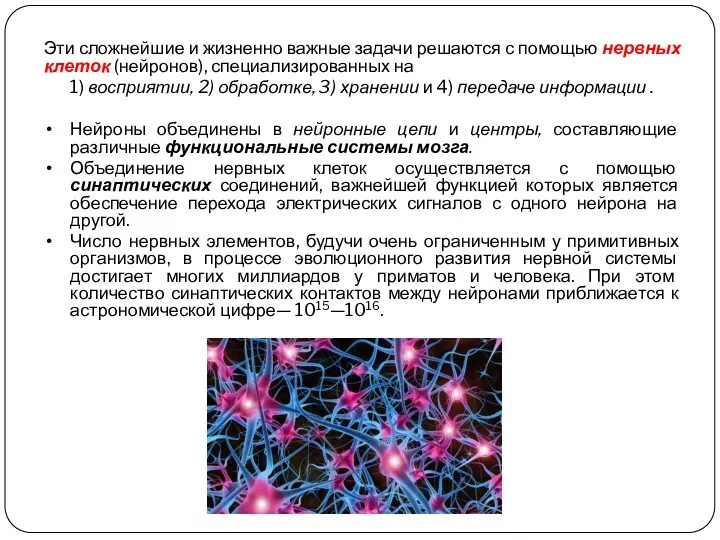
Эти сложнейшие и жизненно важные задачи решаются с помощью нервных клеток
Эти сложнейшие и жизненно важные задачи решаются с помощью нервных клеток

Нейронная теория
Синапс
В основе современного представления о структуре и функции ЦНС лежит
Нейронная теория
Синапс
В основе современного представления о структуре и функции ЦНС лежит

Нобелевская премия по физиологии и медицине 1906 г.
Камило Гольджи (1844-1926)
Нобелевская премия по физиологии и медицине 1906 г.
Камило Гольджи (1844-1926)
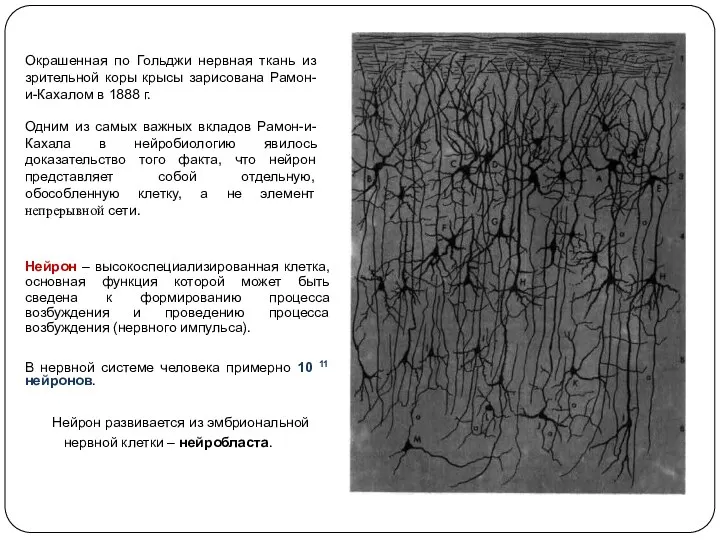
Окрашенная по Гольджи нервная ткань из зрительной коры крысы зарисована Рамон-и-Кахалом
Окрашенная по Гольджи нервная ткань из зрительной коры крысы зарисована Рамон-и-Кахалом
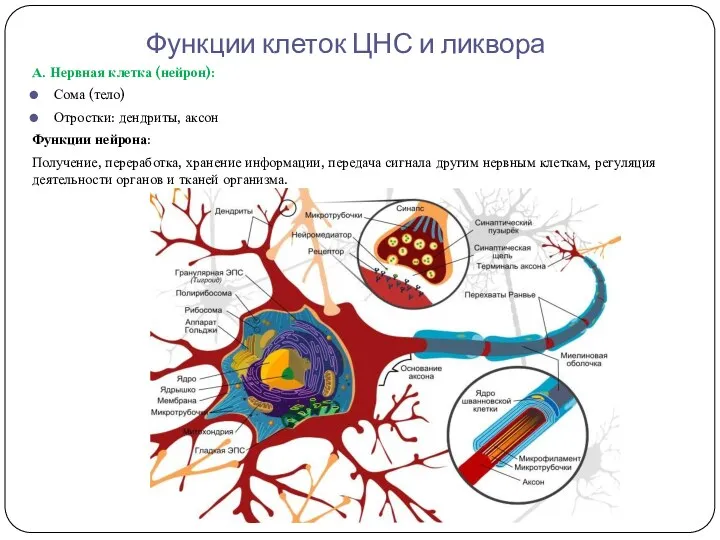
Функции клеток ЦНС и ликвора
А. Нервная клетка (нейрон):
Сома (тело)
Отростки: дендриты, аксон
Функции
Функции клеток ЦНС и ликвора
А. Нервная клетка (нейрон):
Сома (тело)
Отростки: дендриты, аксон
Функции
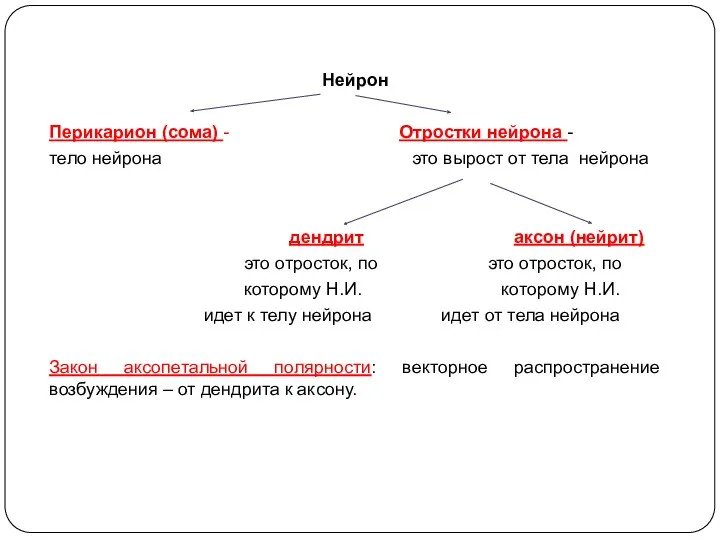
Нейрон
Перикарион (сома) - Отростки нейрона -
тело нейрона это вырост от
Нейрон
Перикарион (сома) - Отростки нейрона -
тело нейрона это вырост от
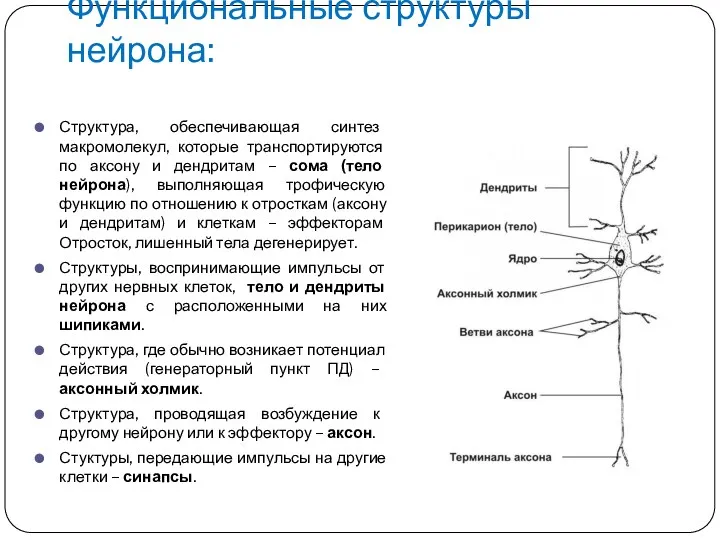
Функциональные структуры нейрона:
Структура, обеспечивающая синтез макромолекул, которые транспортируются по аксону и
Функциональные структуры нейрона:
Структура, обеспечивающая синтез макромолекул, которые транспортируются по аксону и
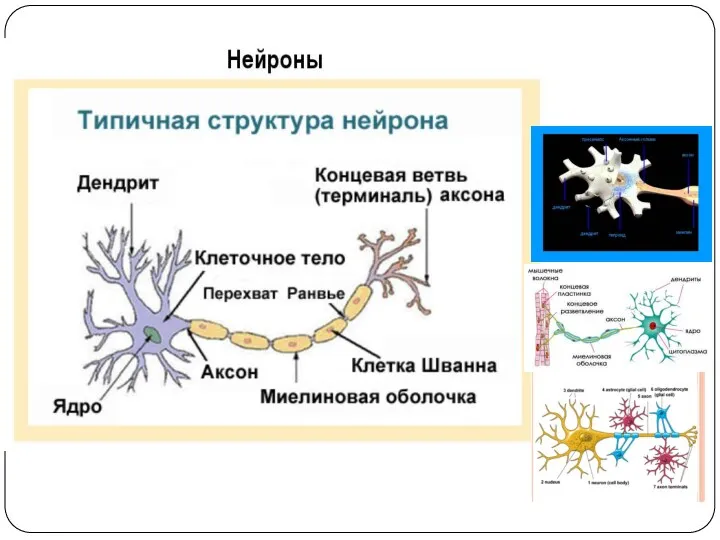
НЕЙРОН. СТРОЕНИЕ
НЕЙРОН. СТРОЕНИЕ
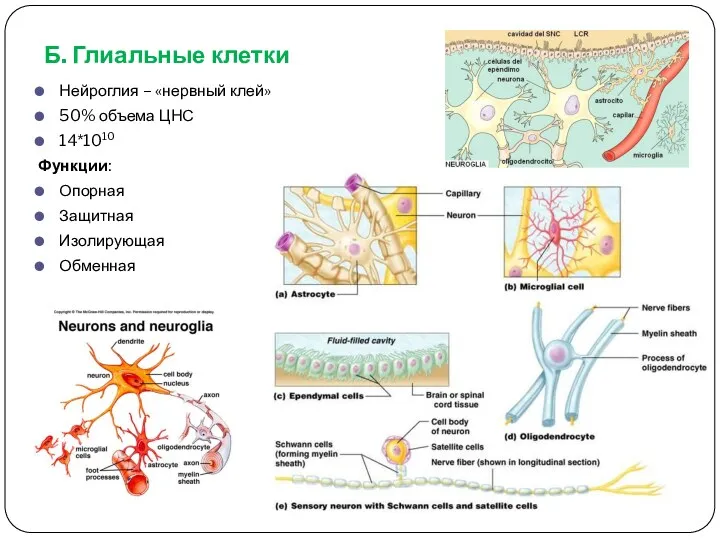
Б. Глиальные клетки
Нейроглия – «нервный клей»
50% объема ЦНС
14*1010
Функции:
Опорная
Защитная
Изолирующая
Обменная
Б. Глиальные клетки
Нейроглия – «нервный клей»
50% объема ЦНС
14*1010
Функции:
Опорная
Защитная
Изолирующая
Обменная
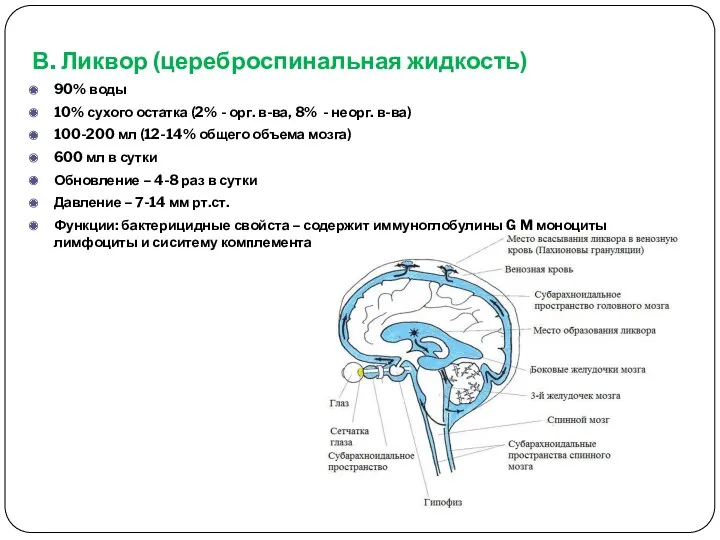
В. Ликвор (цереброспинальная жидкость)
90% воды
10% сухого остатка (2% - орг.
В. Ликвор (цереброспинальная жидкость)
90% воды
10% сухого остатка (2% - орг.

Классификация нейронов
1.В зависимости от отдела ЦНС:
Нейроны соматической НС
Нейроны вегетативной НС
2. По
Классификация нейронов
1.В зависимости от отдела ЦНС:
Нейроны соматической НС
Нейроны вегетативной НС
2. По
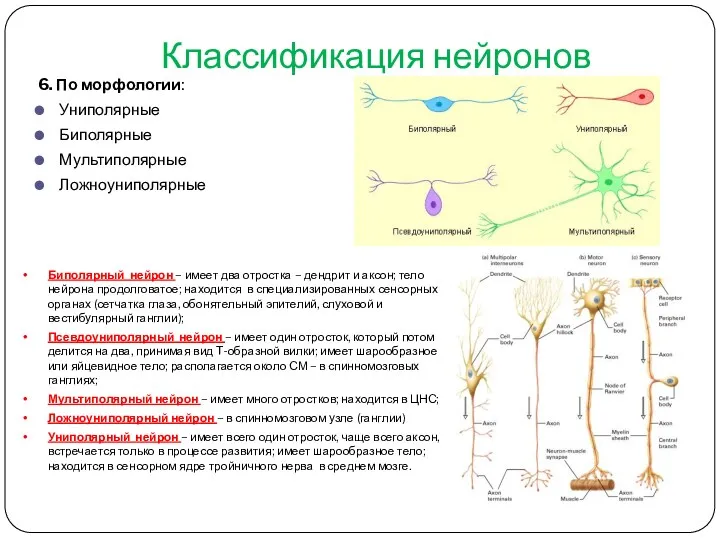
Классификация нейронов
6. По морфологии:
Униполярные
Биполярные
Мультиполярные
Ложноуниполярные
Биполярный нейрон – имеет два отростка – дендрит
Классификация нейронов
6. По морфологии:
Униполярные
Биполярные
Мультиполярные
Ложноуниполярные
Биполярный нейрон – имеет два отростка – дендрит
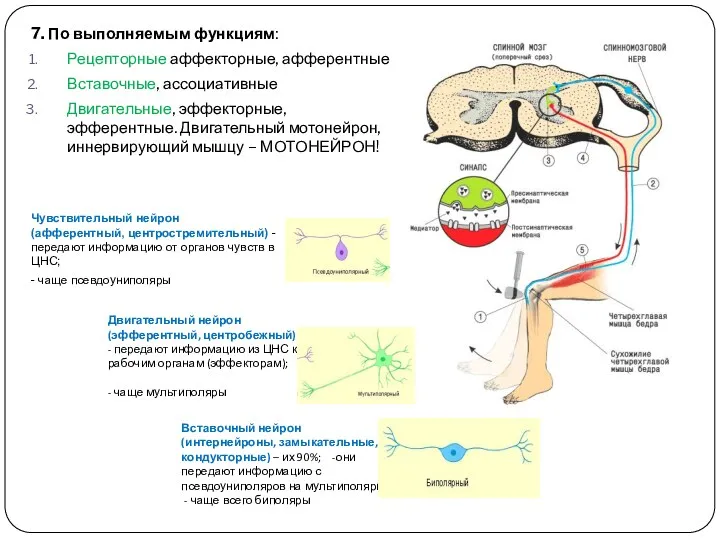
7. По выполняемым функциям:
Рецепторные аффекторные, афферентные
Вставочные, ассоциативные
Двигательные, эффекторные, эфферентные. Двигательный мотонейрон,
7. По выполняемым функциям:
Рецепторные аффекторные, афферентные
Вставочные, ассоциативные
Двигательные, эффекторные, эфферентные. Двигательный мотонейрон,

Нервный отросток – вырост от тела нейрона (дендрит или аксон).
Нервное волокно
Нервный отросток – вырост от тела нейрона (дендрит или аксон).
Нервное волокно
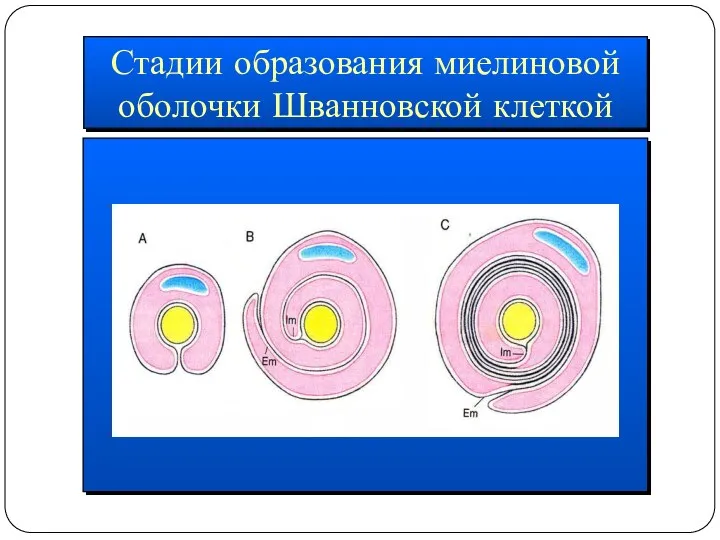
Стадии образования миелиновой
оболочки Шванновской клеткой
Стадии образования миелиновой
оболочки Шванновской клеткой
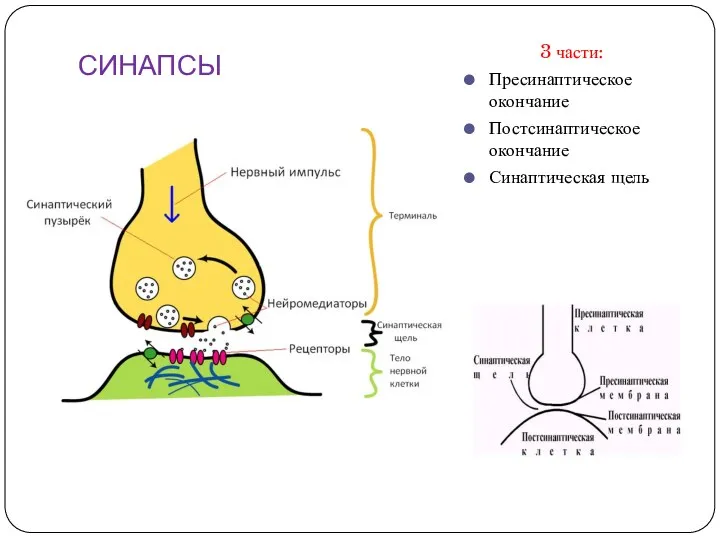
СИНАПСЫ
3 части:
Пресинаптическое окончание
Постсинаптическое окончание
Синаптическая щель
СИНАПСЫ
3 части:
Пресинаптическое окончание
Постсинаптическое окончание
Синаптическая щель
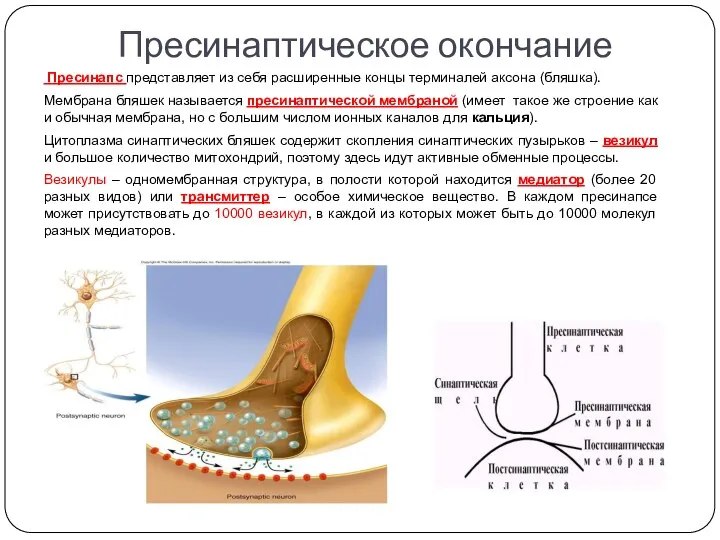
Пресинапс представляет из себя расширенные концы терминалей аксона (бляшка).
Мембрана
Пресинапс представляет из себя расширенные концы терминалей аксона (бляшка).
Мембрана

Синаптическая щель
Постсинапс
Постсинаптическая мембрана содержит большое количество белков-рецепторов с высоким сродством
Синаптическая щель
Постсинапс
Постсинаптическая мембрана содержит большое количество белков-рецепторов с высоким сродством
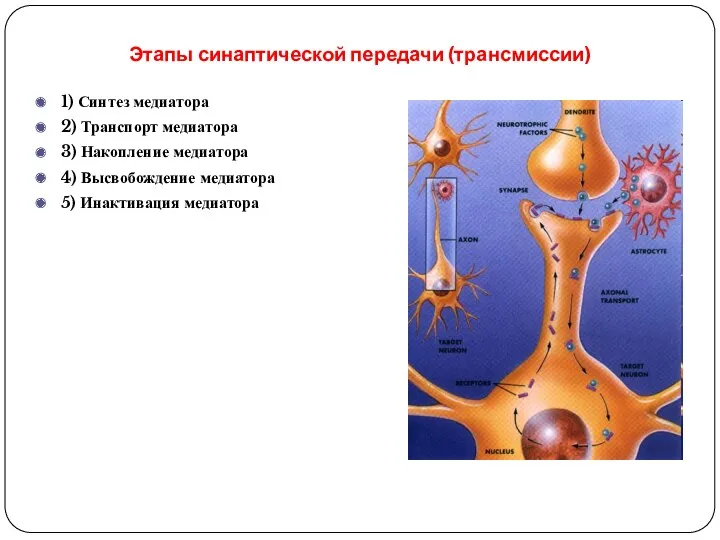
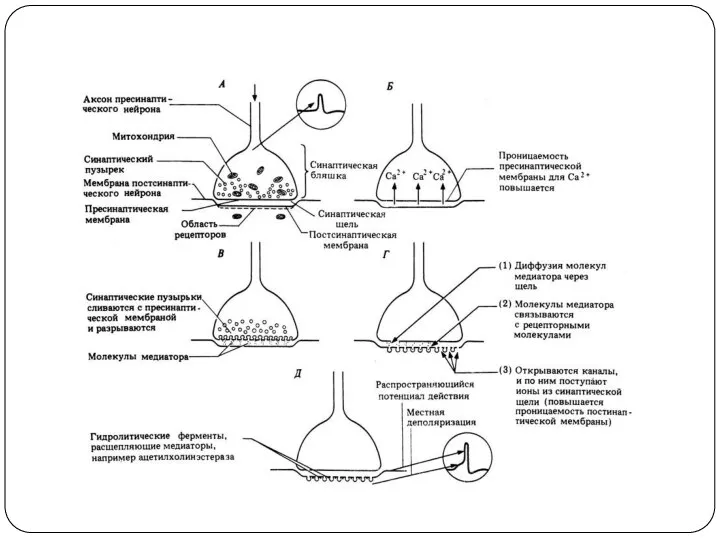
Этапы синаптической передачи (трансмиссии)
1) Синтез медиатора
2) Транспорт медиатора
3) Накопление медиатора
4) Высвобождение
Этапы синаптической передачи (трансмиссии)
1) Синтез медиатора
2) Транспорт медиатора
3) Накопление медиатора
4) Высвобождение
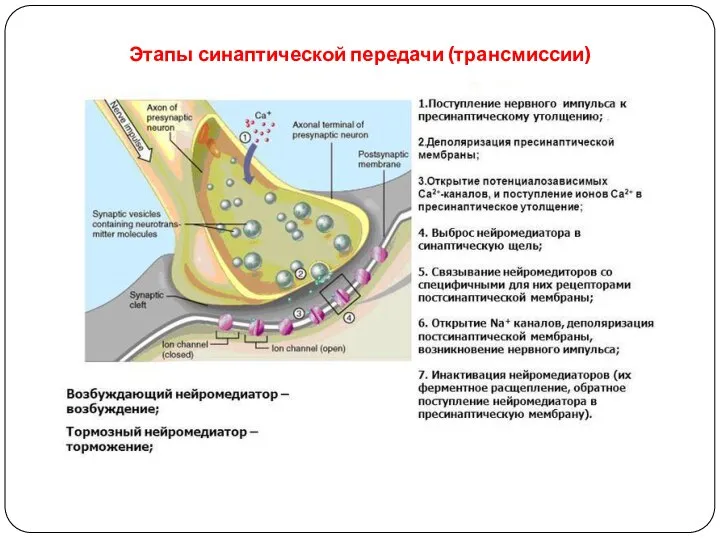
Этапы синаптической передачи (трансмиссии)
Этапы синаптической передачи (трансмиссии)
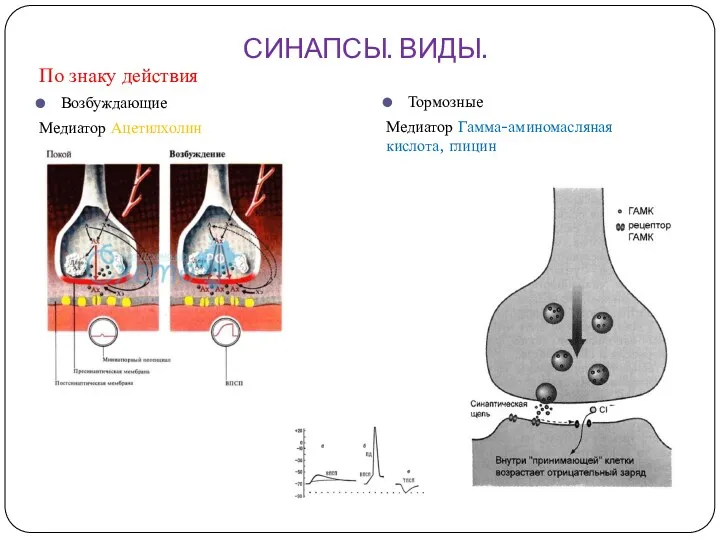
СИНАПСЫ. ВИДЫ.
По знаку действия
Возбуждающие
Медиатор Ацетилхолин
Тормозные
Медиатор Гамма-аминомасляная кислота, глицин
СИНАПСЫ. ВИДЫ.
По знаку действия
Возбуждающие
Медиатор Ацетилхолин
Тормозные
Медиатор Гамма-аминомасляная кислота, глицин
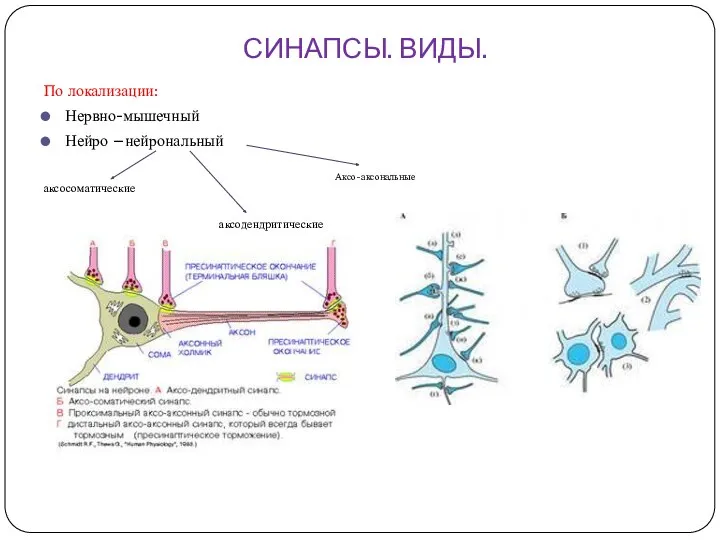
СИНАПСЫ. ВИДЫ.
По локализации:
Нервно-мышечный
Нейро –нейрональный
аксосоматические
аксодендритические
Аксо-аксональные
СИНАПСЫ. ВИДЫ.
По локализации:
Нервно-мышечный
Нейро –нейрональный
аксосоматические
аксодендритические
Аксо-аксональные
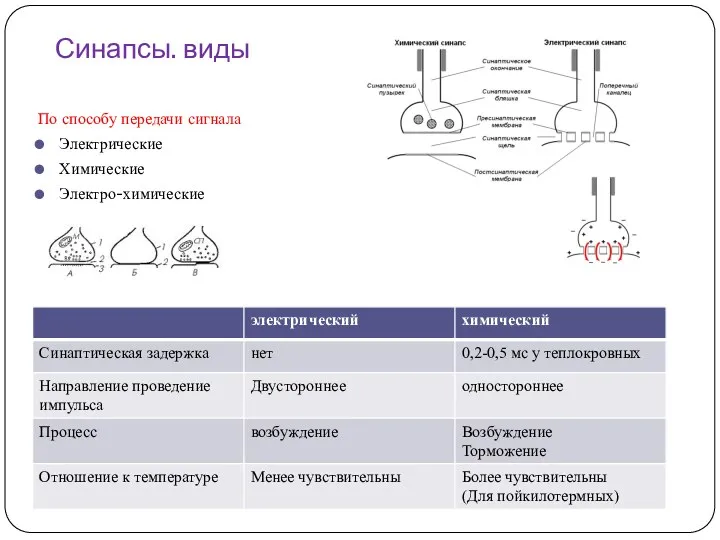
Синапсы. виды
По способу передачи сигнала
Электрические
Химические
Электро-химические
Синапсы. виды
По способу передачи сигнала
Электрические
Химические
Электро-химические

В синапсах с химическим механизмом передачи продолжительность синаптической задержки у теплокровных
В синапсах с химическим механизмом передачи продолжительность синаптической задержки у теплокровных

Медиаторы и рецепторы ЦНС
А. Ацетилхолин
Б. Амины (дофамин, норадреналин, серотонин, гистамин)
В. Аминокислоты(глицин,
Медиаторы и рецепторы ЦНС
А. Ацетилхолин
Б. Амины (дофамин, норадреналин, серотонин, гистамин)
В. Аминокислоты(глицин,

ПЛАН:
Рефлекс.
Виды рефлексов
Рефлекторная дуга
Торможение, виды.
Тема: РЕФЛЕКТОРНАЯ ДЕЯТЕЛЬНОСТЬ ЦНС
ПЛАН:
Рефлекс.
Виды рефлексов
Рефлекторная дуга
Торможение, виды.
Тема: РЕФЛЕКТОРНАЯ ДЕЯТЕЛЬНОСТЬ ЦНС

ЦНС осуществляет две функции
рефлекторную и проводниковую.
Рефлекторная деятельность осуществляется за
ЦНС осуществляет две функции
рефлекторную и проводниковую.
Рефлекторная деятельность осуществляется за
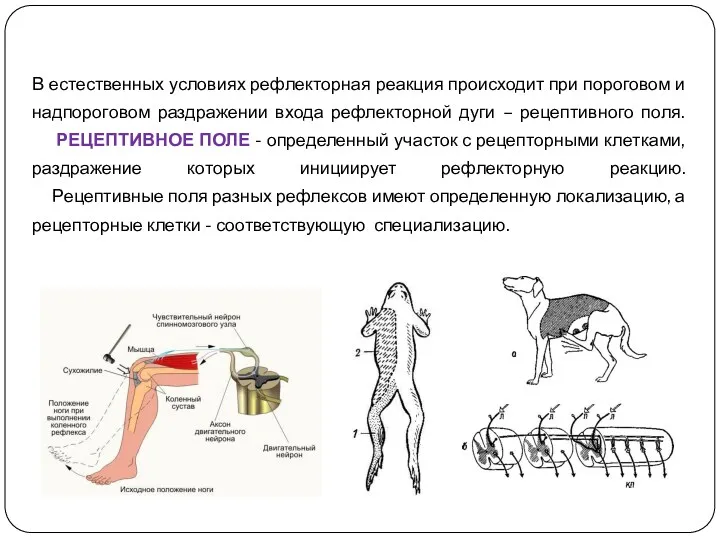
В естественных условиях рефлекторная реакция происходит при пороговом и надпороговом раздражении
В естественных условиях рефлекторная реакция происходит при пороговом и надпороговом раздражении
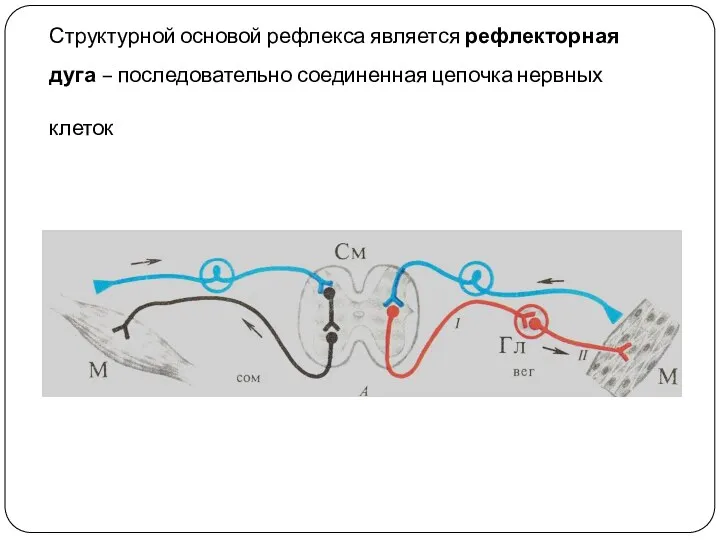
Структурной основой рефлекса является рефлекторная дуга – последовательно соединенная цепочка нервных
Структурной основой рефлекса является рефлекторная дуга – последовательно соединенная цепочка нервных

ВРЕМЯ РЕФЛЕКСА - это время, необходимое для осуществления рефлекса, складывается
ВРЕМЯ РЕФЛЕКСА - это время, необходимое для осуществления рефлекса, складывается

КЛАССИФИКАЦИЯ РЕФЛЕКСОВ
1. По способу вызывания:
- безусловные рефлексы
- условные
2. По месту расположения
КЛАССИФИКАЦИЯ РЕФЛЕКСОВ
1. По способу вызывания:
- безусловные рефлексы
- условные
2. По месту расположения
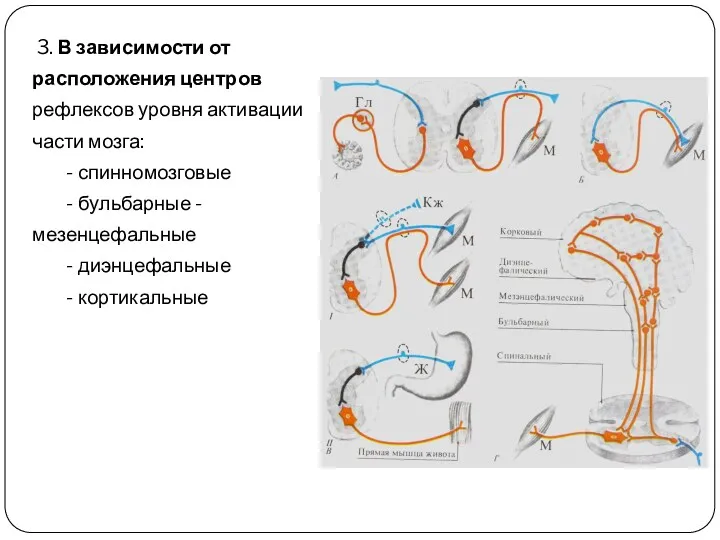
3. В зависимости от расположения центров рефлексов уровня активации части
3. В зависимости от расположения центров рефлексов уровня активации части

4. По биологическому значению
- пищевые
- оборонительные
- половые и
4. По биологическому значению
- пищевые
- оборонительные
- половые и

Рефлекторная деятельность организма во многом определяется общими свойствами нервных центров.
Нервный центр
Рефлекторная деятельность организма во многом определяется общими свойствами нервных центров.
Нервный центр

Классификация нервных центров
Морфологический критерий (локализация в отделах ЦНС ):
Спинальные центры (в
Классификация нервных центров
Морфологический критерий (локализация в отделах ЦНС ):
Спинальные центры (в

Нервные центры
В основе нервной деятельности лежат активные и противоположные по
Нервные центры
В основе нервной деятельности лежат активные и противоположные по
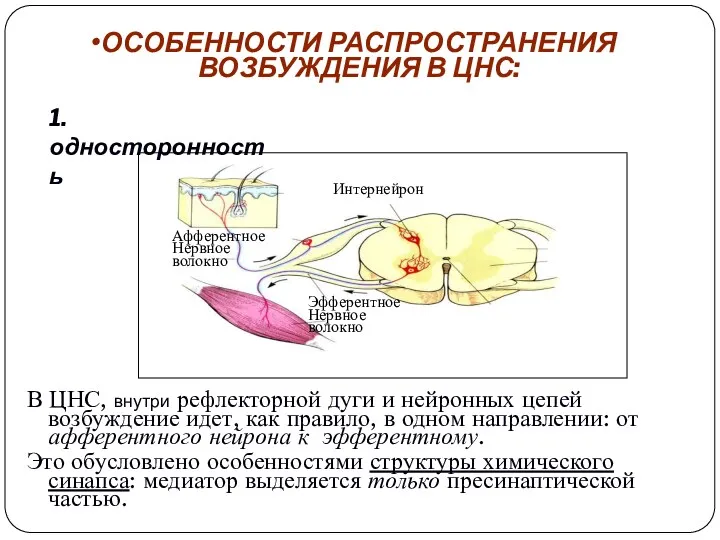
ОСОБЕННОСТИ РАСПРОСТРАНЕНИЯ
ВОЗБУЖДЕНИЯ В ЦНС:
В ЦНС, внутри рефлекторной дуги и нейронных
ОСОБЕННОСТИ РАСПРОСТРАНЕНИЯ
ВОЗБУЖДЕНИЯ В ЦНС:
В ЦНС, внутри рефлекторной дуги и нейронных
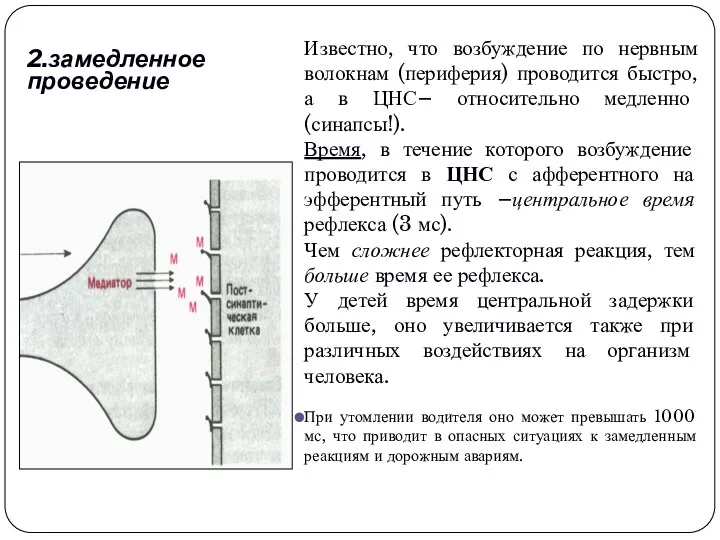
2.замедленное проведение
Известно, что возбуждение по нервным волокнам (периферия) проводится быстро, а
2.замедленное проведение
Известно, что возбуждение по нервным волокнам (периферия) проводится быстро, а

3. суммация
Это свойство впервые описал И.М. Сеченов (1863):
При действии ряда подпороговых
3. суммация
Это свойство впервые описал И.М. Сеченов (1863):
При действии ряда подпороговых
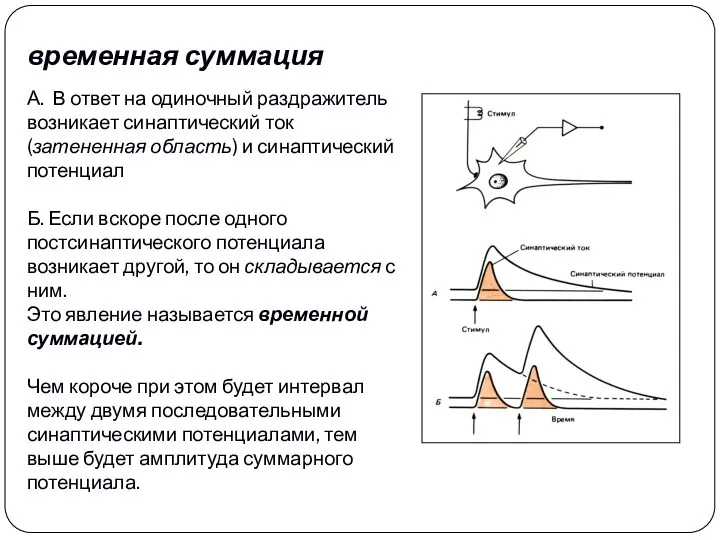
временная суммация
А.. В ответ на одиночный раздражитель возникает синаптический ток
временная суммация
А.. В ответ на одиночный раздражитель возникает синаптический ток
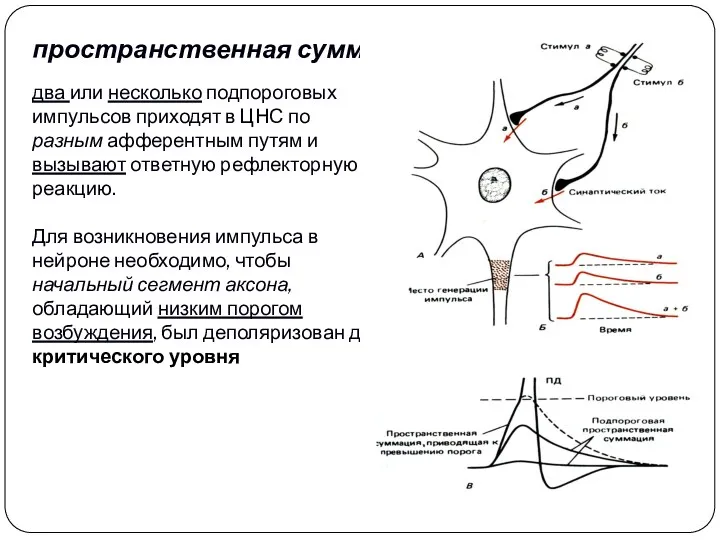
пространственная суммация
два или несколько подпороговых импульсов приходят в ЦНС по разным
пространственная суммация
два или несколько подпороговых импульсов приходят в ЦНС по разным
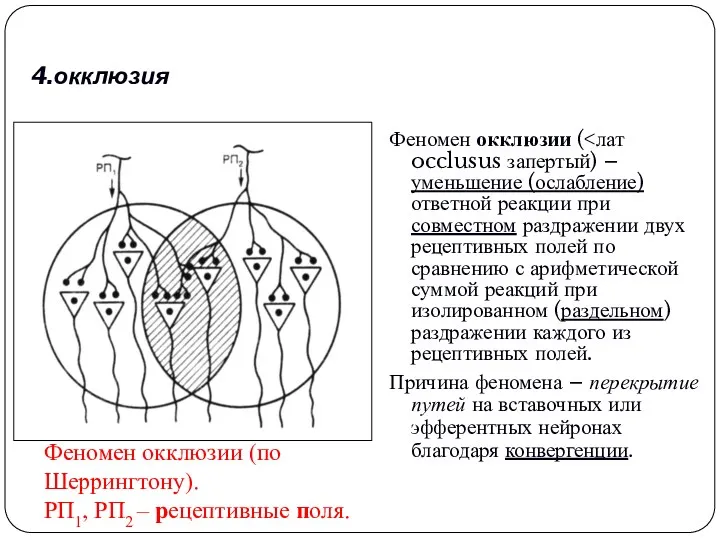
4.окклюзия
Феномен окклюзии (<лат occlusus запертый) – уменьшение (ослабление) ответной реакции при
4.окклюзия
Феномен окклюзии (<лат occlusus запертый) – уменьшение (ослабление) ответной реакции при
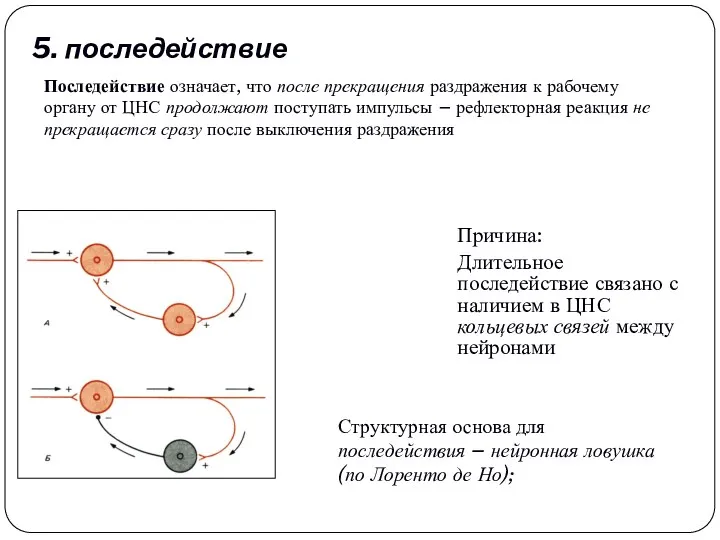
5. последействие
Причина:
Длительное последействие связано с наличием в ЦНС кольцевых связей между
5. последействие
Причина:
Длительное последействие связано с наличием в ЦНС кольцевых связей между

6. проторение (постактивационное облегчение)
Проторение (постактивационное облегчение):
После возбуждения, вызванного ритмической стимуляцией,
6. проторение (постактивационное облегчение)
Проторение (постактивационное облегчение):
После возбуждения, вызванного ритмической стимуляцией,

Свойства нервных центров:
Длительное повторное раздражение рецептивного поля рефлекса →ослабление рефлекторной реакции
Свойства нервных центров:
Длительное повторное раздражение рецептивного поля рефлекса →ослабление рефлекторной реакции
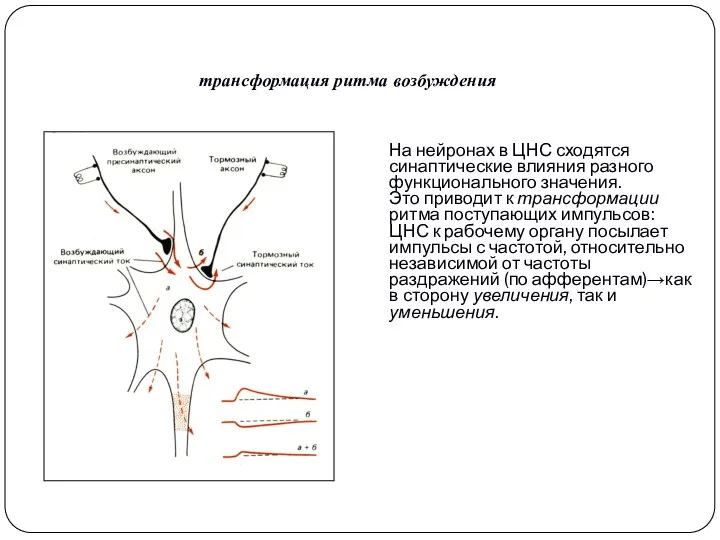
На нейронах в ЦНС сходятся синаптические влияния разного функционального значения.
Это
На нейронах в ЦНС сходятся синаптические влияния разного функционального значения.
Это

Обусловлена высокой интенсивность обменных процессов:
100 г нервной ткани (головной мозг собаки)
Обусловлена высокой интенсивность обменных процессов:
100 г нервной ткани (головной мозг собаки)

Пластичность – функциональная подвижность нервного центра: возможность его включения в регуляцию
Пластичность – функциональная подвижность нервного центра: возможность его включения в регуляцию

Торможение в ЦНС (И.М. Сеченов), его виды и роль. Современное представление
Торможение в ЦНС (И.М. Сеченов), его виды и роль. Современное представление
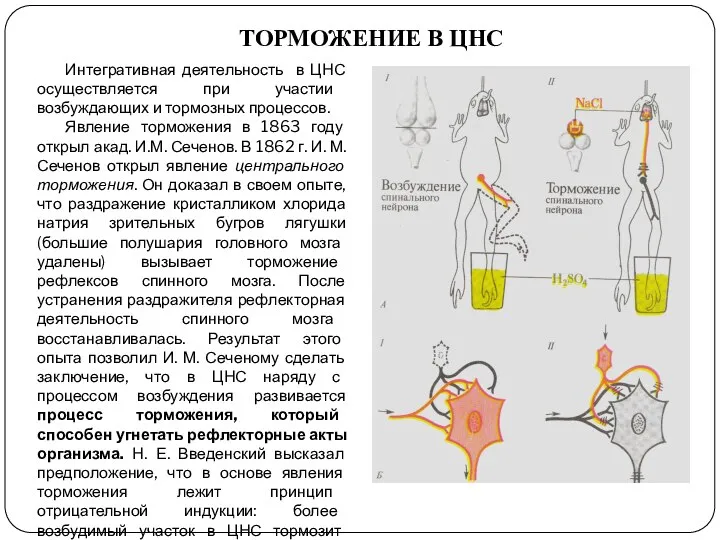
ТОРМОЖЕНИЕ В ЦНС
Интегративная деятельность в ЦНС осуществляется при участии возбуждающих
ТОРМОЖЕНИЕ В ЦНС
Интегративная деятельность в ЦНС осуществляется при участии возбуждающих

Современная трактовка опыта И. М. Сеченова (И. М. Сеченов раздражал ретикулярную
Современная трактовка опыта И. М. Сеченова (И. М. Сеченов раздражал ретикулярную
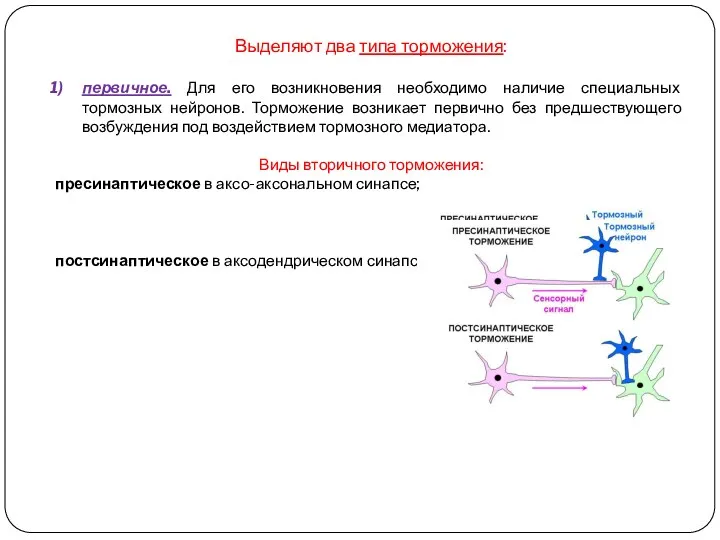
Выделяют два типа торможения:
первичное. Для его возникновения необходимо наличие специальных тормозных
Выделяют два типа торможения:
первичное. Для его возникновения необходимо наличие специальных тормозных

Постсинаптическое торможение
(лат. post позади, после чего-либо + греч. sinapsis соприкосновение,
Постсинаптическое торможение
(лат. post позади, после чего-либо + греч. sinapsis соприкосновение,
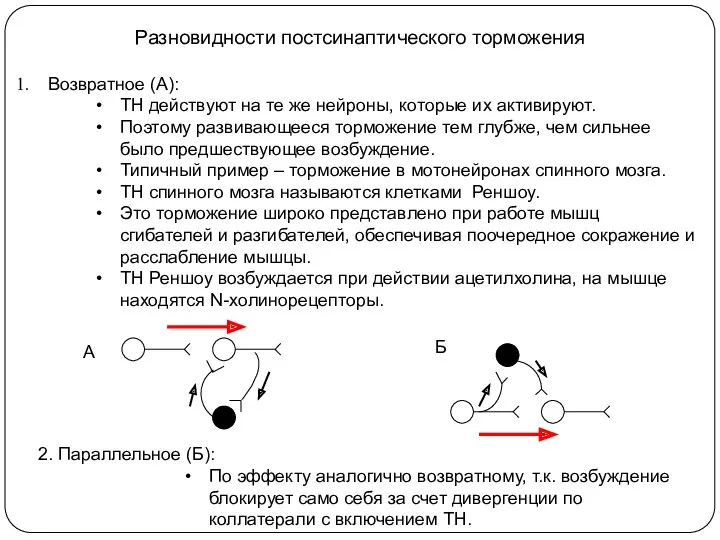
Разновидности постсинаптического торможения
Возвратное (А):
ТН действуют на те же нейроны, которые их
Возвратное (А):
ТН действуют на те же нейроны, которые их
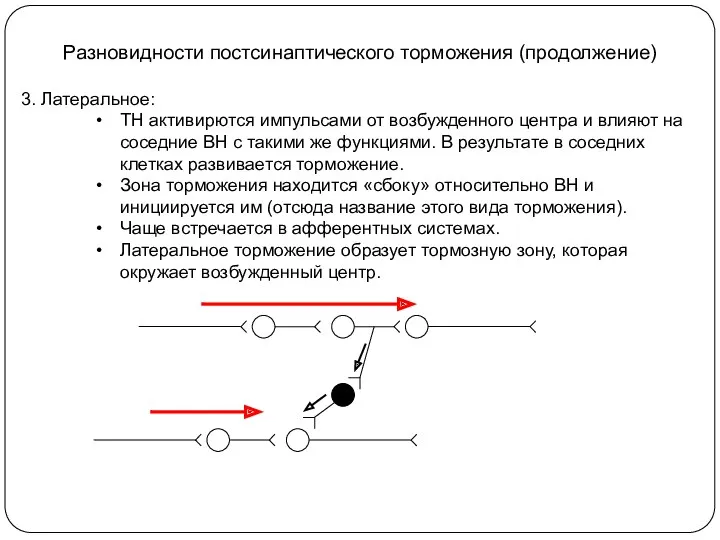
Разновидности постсинаптического торможения (продолжение)
3. Латеральное:
ТН активирются импульсами от возбужденного центра и
3. Латеральное:
ТН активирются импульсами от возбужденного центра и
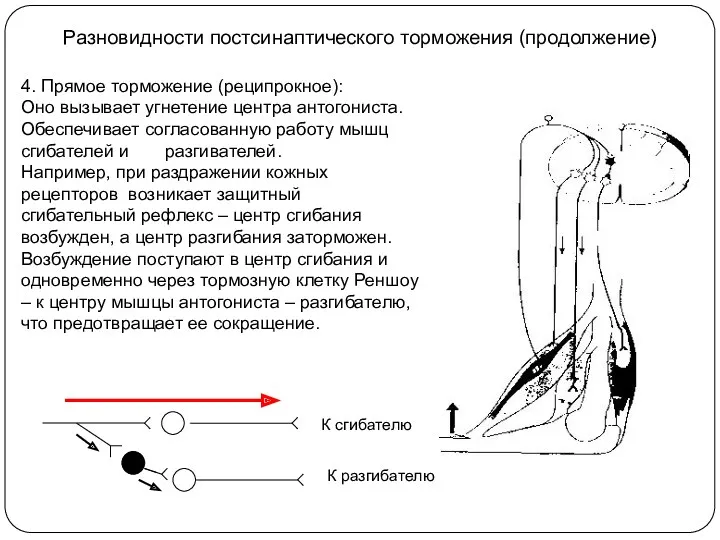
Разновидности постсинаптического торможения (продолжение)
4. Прямое торможение (реципрокное):
Оно вызывает угнетение центра антогониста.
4. Прямое торможение (реципрокное):
Оно вызывает угнетение центра антогониста.

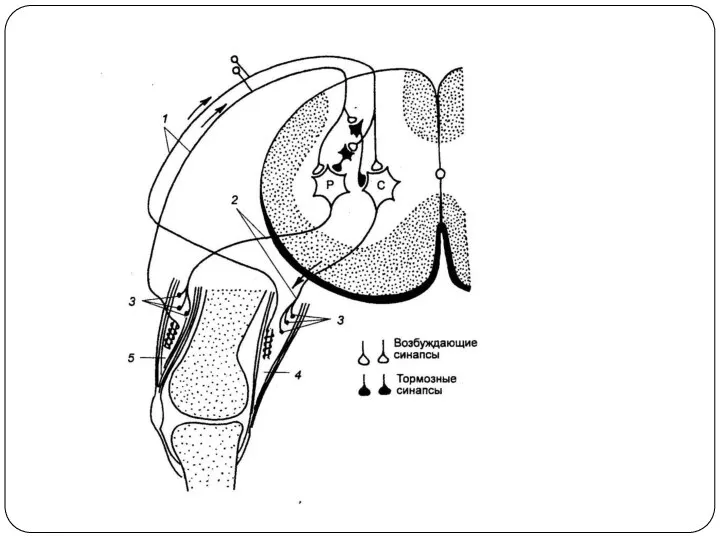
Пресинаптическое торможение
Развивается на пресинаптической мембране.
Осуществляется посредством специальных тормозных нейронов.
Пресинаптическое торможение
Развивается на пресинаптической мембране.
Осуществляется посредством специальных тормозных нейронов.
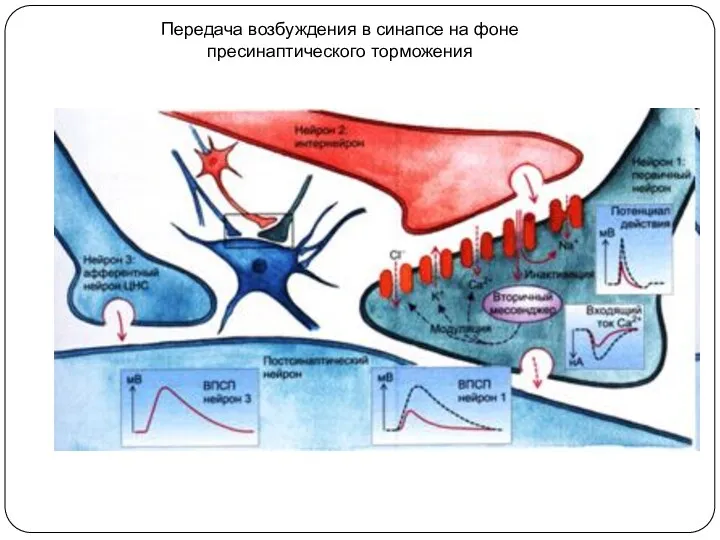
Передача возбуждения в синапсе на фоне пресинаптического торможения
Передача возбуждения в синапсе на фоне пресинаптического торможения
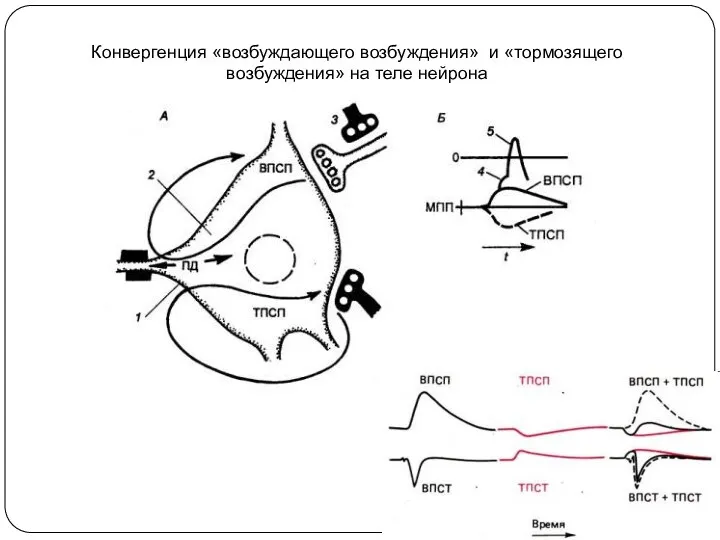
Конвергенция «возбуждающего возбуждения» и «тормозящего возбуждения» на теле нейрона
Конвергенция «возбуждающего возбуждения» и «тормозящего возбуждения» на теле нейрона

Виды вторичного торможения:
запредельное, возникающее при большом потоке информации, поступающей в клетку.
Виды вторичного торможения:
запредельное, возникающее при большом потоке информации, поступающей в клетку.

Значение торможения
Торможение выполняет охранительную роль, его отсутствие привело бы к истощению
Значение торможения
Торможение выполняет охранительную роль, его отсутствие привело бы к истощению

Принципы координации нервных центров
Координация (лат. сo - вместе+ ordinatio – расположение
Принципы координации нервных центров
Координация (лат. сo - вместе+ ordinatio – расположение

Принципы координации нервных центров:
«общий конечный путь» (конвергенция)
Выдвинут Ч.С. Шеррингтоном в
Принципы координации нервных центров:
«общий конечный путь» (конвергенция)
Выдвинут Ч.С. Шеррингтоном в
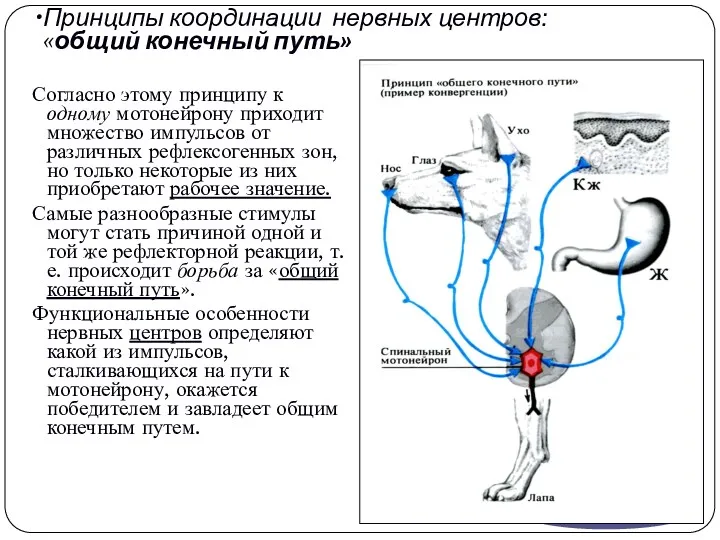
Принципы координации нервных центров:
«общий конечный путь»
Согласно этому принципу к одному
Принципы координации нервных центров:
«общий конечный путь»
Согласно этому принципу к одному

Принципы координации нервных центров: доминанта
Принцип доминанты (лат. dominare господствовать) – установлен
Принципы координации нервных центров: доминанта
Принцип доминанты (лат. dominare господствовать) – установлен

Доминанта
Свойства доминантного очага:
Повышенная возбудимость;
Инерционность;
Способность к суммации;
Способность к торможению центров, функционально несовместимых
Доминанта
Свойства доминантного очага:
Повышенная возбудимость;
Инерционность;
Способность к суммации;
Способность к торможению центров, функционально несовместимых

Доминанта
А.А. Ухтомский о (+) и (–) доминанты:
«… Доминанта, как общая формула,
Доминанта
А.А. Ухтомский о (+) и (–) доминанты:
«… Доминанта, как общая формула,
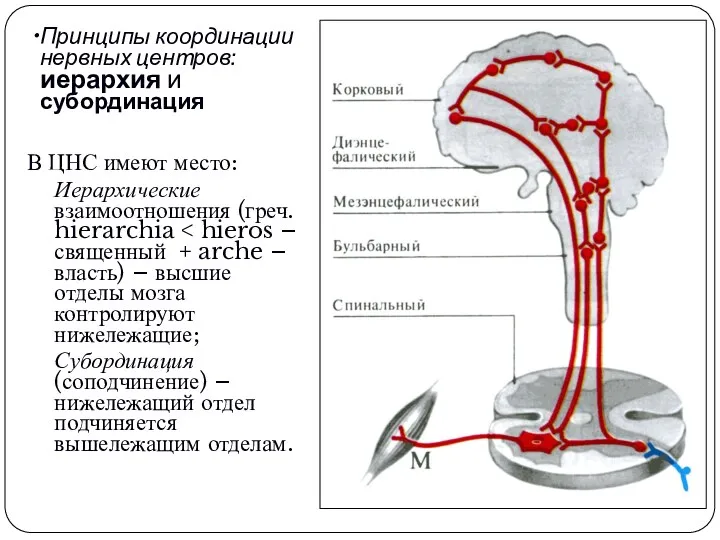
Принципы координации нервных центров: иерархия и субординация
В ЦНС имеют место:
Иерархические взаимоотношения
Принципы координации нервных центров: иерархия и субординация
В ЦНС имеют место:
Иерархические взаимоотношения

Принципы координации нервных центров: иррадиация
Иррадиация (лат. irradio освещать, озарять) –
Принципы координации нервных центров: иррадиация
Иррадиация (лат. irradio освещать, озарять) –
 Гусеводство. Продукция и направление гусеводства
Гусеводство. Продукция и направление гусеводства Экологические аспекты производства биопрепаратов
Экологические аспекты производства биопрепаратов Классификация животных. Основные систематические группы. Влияние человека на животных
Классификация животных. Основные систематические группы. Влияние человека на животных Развитие жизни на Земле в различные периоды
Развитие жизни на Земле в различные периоды Тип Кишечнополостные
Тип Кишечнополостные Анатомия и физиология человека. Органы чувств. Анализаторы
Анатомия и физиология человека. Органы чувств. Анализаторы Антропосоциогенез
Антропосоциогенез Комнатные растения в интерьере помещения
Комнатные растения в интерьере помещения Растения красной книги России
Растения красной книги России Возбудимые ткани
Возбудимые ткани Классификация возрастных периодов
Классификация возрастных периодов Общие принципы анатомического строения и закономерности регуляции функциональных систем организма
Общие принципы анатомического строения и закономерности регуляции функциональных систем организма Тренажёр Птицы, биология 8 класс
Тренажёр Птицы, биология 8 класс Выступление на всероссийском семинаре Личностно- ориентированнный подход при создании проектных работ.
Выступление на всероссийском семинаре Личностно- ориентированнный подход при создании проектных работ. Межклеточная сигнализация. Сигнальные молекулы. Гормоны. (Тема 3)
Межклеточная сигнализация. Сигнальные молекулы. Гормоны. (Тема 3) Опорные системы
Опорные системы Бактериологический анализ
Бактериологический анализ Биосинтез белка
Биосинтез белка Неживая природа
Неживая природа Органы чуств. Регуляция деятельности организма
Органы чуств. Регуляция деятельности организма Обитатели моря
Обитатели моря Углеводы. Строение и функции
Углеводы. Строение и функции Female culex
Female culex Комнатные растения. О чем рассказали листья
Комнатные растения. О чем рассказали листья Опорно-двигательный аппарат. Скелет человека
Опорно-двигательный аппарат. Скелет человека Игра крестики - нолики (природа Марийского края)
Игра крестики - нолики (природа Марийского края) Магистры Тема 4б
Магистры Тема 4б Строение волос человека. Волосы: строение и функции
Строение волос человека. Волосы: строение и функции