- Функциональная морфология биологических мембран

Содержание
- 2. Биологическая мембрана Цитоплазматическая; Плазматическая; ультратонкая пленка, состоящая из двойного слоя липидных молекул, с которым связаны белки
- 3. Строение биологической мембраны Молекулы мембранных липидов – амфифильные молекулы, т.е. имеют полярную («головку») и неполярную («хвост»)
- 4. Структура мембран
- 5. Сборка липидного бислоя Находящиеся в водной фазе фосфолипиды формируют двухслойные структуры, объединяя свои гидрофобные участки. Когда
- 6. Плазматическая мембрана (внешняя клеточная мембрана, цитолемма, плазмолемма) основной, постоянный, универсальный для всех клеток компонент системы поверхностного
- 7. Плазматическая мембрана Структура толщиной 7-10 нм, образованная, главным образом, липидами и белками. Молекулярное строение плазмолеммы описывается
- 8. Поверхности плазматической мембраны наружная (внешняя); Е-поверхность (от англ. еxternal); Р-поверхность (от англ. protoplasmic); цитоплазматическая (внутренняя); Биологическая
- 9. Жирные кислоты и свойства мембраны Чем длиннее алифатический радикал, тем толще липидный бислой (орt-16-20). 2. Чем
- 10. Содержание фосфолипидов в плазматической мембране
- 11. Холестерин и проницаемость биологической мембраны В мембране с преобладанием насыщенных жирных кислот («жесткая» мембрана), молекула холестерола
- 12. Мембранные белки - функционально гетерогенная группа белков (рецепторы, ферменты, переносчики, каналы и др.), обеспечивающая специфические свойства
- 14. Химическая модификация мембранных липидов и белков Молекулы олигосахаридов связываются с липидами (гликолипиды) и мембранными белками (гликопротеины).
- 15. Гликокаликс С наружной стороны плазмолеммы имеется надмембранный слой - гликокаликс (3-4 нм). Он образован углеводными фрагментами
- 16. Субмембранный комплекс Функции: Участвует в поддержании формы клетки. Участвует в формировании межклеточных контактов. Обеспечивает мембранные процессы.
- 17. Мобильность липидов При температуре тела мембрана текуча и липиды свободно перемещаются в ней. Латеральная подвижность, то
- 18. Перемещение мембранных белков Белковые молекулы мозаично распределены в липидном бислое и перемещаться в его плоскости. Перемещение
- 19. Асимметрия биологических мембран Липидный состав монослоев различен. Например, в плазматической мембране фосфатидилхолин преобладает в наружном слое,
- 20. Липидные рафты домены липидного бислоя клеточной мембраны, обогащённые холестерином, и насыщенными фосфолипидами. Это участок плотно упакованного
- 21. Эффект молекулярной памяти липидов (эф-т Л.Д.Бергельсона) Краткосрочные события, протекающие в белковой молекуле клеточной мембраны, влияют на
- 22. Эффект Л.Д. Бергельсона Анулярные липиды – липиды, окружающие молекулу мембран-ного белка и взаимодействующие с ним.
- 23. Заряд биологической мембраны Определяется совокупностью зарядов входящих в их состав молекул липидов, белков и углеводов, а
- 24. Обновление мембран в клетке Баланс эндоцитоз/экзоцитоз для плазмолеммы. Обмен везикулами между органеллами. Синтез de novo компонентов
- 25. Функции плазмолеммы: барьерная; транспорт веществ в клетку и из клетки; взаимодействие с сигнальными молекулами; взаимодействие с
- 26. Рецепторы гликопротеины, способные высокоселективно связываться с определенными молекулами - лигандами (гормон, медиатор). Регулируют проницаемость плазмолеммы. Передают
- 27. Каталитические рецепторы - цитоплазматический домен обладает ферментативной активностью. При связывании лиганда внеклеточным доменом активируется протеинкиназная или
- 28. Рецепторы-каналы После связывания с лигандом, изменяют свою конформацию, что ведет к возникновению в мембране гидрофильного канала
- 29. Рецепторы, связанные с G-белками
- 30. Трансмембранный транспорт веществ прямой (диффузия, осмос, фильтрация); опосредованный: 1. пассивный (с участием белка-переносчика или с участием
- 31. Простая диффузия - переход вещества из области более высокой концентрации в область меньшей. Небольшие нейтральные молекулы
- 32. Осмос движение молекул растворителя через полупроницаемую мембрану из области с низкой концентрацией растворенного вещества в область
- 33. Фильтрация движение веществ через поры под действием избыточного давления.
- 34. Облегчённая диффузия транспорт веществ через мембрану с помощью специальных белков. Поры – отверстия в мембране, образованные
- 35. Эндоцитоз вид активного транспорта, с формированием эндоцитозной везикулы. Всегда рецептор-опосредованный Всегда с участием цитоскелета
- 36. Виды эндоцитоза Фагоцитоз - процесс поглощения клеткой объектов размером >10-6 м, таких как бактерии, вирусы, остатки
- 37. Окаймленные ямки Клатрин — внутриклеточный белок молекулы, которого формируют тримеры. В результате полимеризации тримеров клатрина формируется
- 38. Кавеолы - углубление плазмолеммы, покрытое с цитоплазматической стороны кавеолинами. Кавеолины — группа мембранных белков рецептор-независимого эндоцитоза.
- 39. Динамин - эукариотическая клеточная ГТФ-аза, участвующая в эндоцитозе. Динамин необходим для отделения сформировавшейся везикулы от плазмолеммы
- 40. Экзоцитоз - процесс слияния экзоцитозного пузырька с плазматической мембраной клетки в результате которого его содержимое освобождается
- 41. Первично-активный транспорт перенос веществ через мембрану против градиента концентрации при участии транспортных АТФ-аз. Na+/K+-АТФ-аза, Са++-АТФ-аза, Н+/К+-АТФ-аза.
- 42. Вторично-активный транспорт перенос веществ против градиента концентрации сопряжен с одновременным переносом другого вещества по градиенту концентрации
- 43. Кинки Проницаемость липидного бислоя зависит от степени ненасыщенности жирных кислот мембранных липидов. В липидном бислое имеются
- 44. Межклеточные контакты Механические – контакты, которые обеспечивают механическую связь клеток друг с другом (интердигитации, десмосомы, промежуточные
- 45. Интердигитации Наиболее просто устроенный вид взаимодействия мембран смежных клеток. Представлены выпячиваниями цитоплазмы через плазмолемму одной клетки,
- 46. Плотное соединение соединение наружных листков плазмолемм взаимодействующих клеток, опосредованное трансмембранными белками-окклюдинами. Окклюдины мембраны одной клетки взаимодействуют
- 47. Промежуточное соединение, опоясывающая десмосома Охватывают клетку по периметру в виде пояса. С цитоплазматической стороны плазмолеммы в
- 48. Десмосома (пятно сцепления) Межклеточная щель расширена (25 нм).
- 49. Кадхерины Кадхерины «зоны слипания»: Е-кадхерин (эпителиальный кадхерин, увоморулин, L-CAM). N-кадхерин (нейральный кадхерин, A-CAМ). Р-кадхерин (плацентарный кадхерин).
- 50. Нексус (щелевое соединение) Образован соединением трансмембранных гексамеров – коннексонами (10 нм) одной клетки, с а налогичными
- 51. Интегрины суперсемейство молекул адгезии к молекулам межклеточного матрикса: Коллаген. Фибронектин. Ламинин. Все интегрины гетеродимерные трансмембранные белки,
- 52. Селектины - семейство трансмембранных мономерных гликопротеинов, участвующих в межклеточной адгезии. Каждый рецептор имеет один трансмембранный домен.
- 54. Скачать презентацию

Биологическая мембрана
Цитоплазматическая;
Плазматическая;
ультратонкая пленка, состоящая из двойного слоя
Биологическая мембрана
Цитоплазматическая;
Плазматическая;
ультратонкая пленка, состоящая из двойного слоя
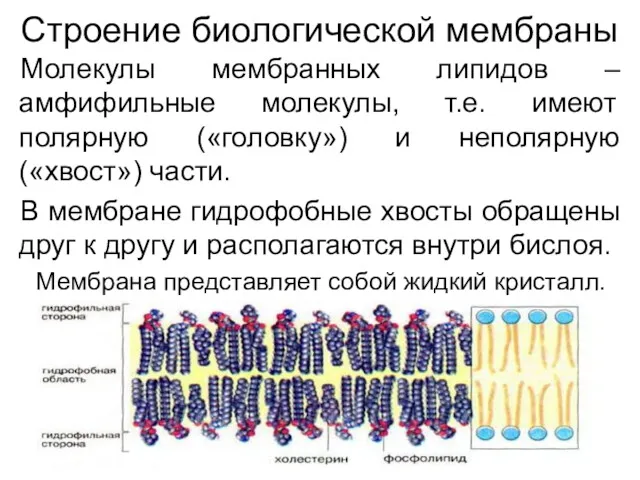
Строение биологической мембраны
Молекулы мембранных липидов – амфифильные молекулы, т.е. имеют полярную
Строение биологической мембраны
Молекулы мембранных липидов – амфифильные молекулы, т.е. имеют полярную
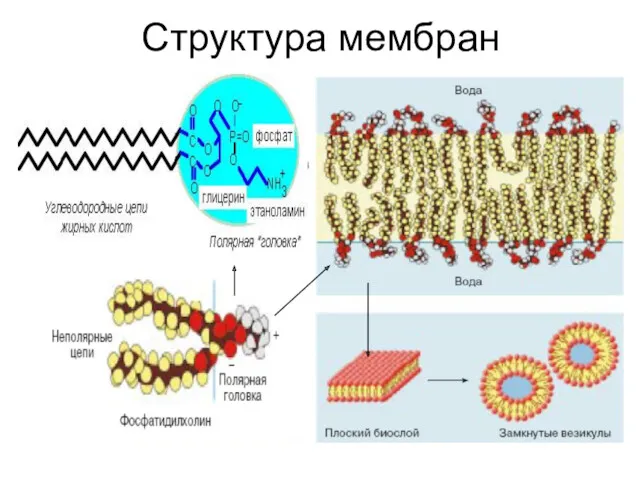
Структура мембран
Структура мембран

Сборка липидного бислоя
Находящиеся в водной фазе фосфолипиды формируют двухслойные структуры, объединяя
Сборка липидного бислоя
Находящиеся в водной фазе фосфолипиды формируют двухслойные структуры, объединяя
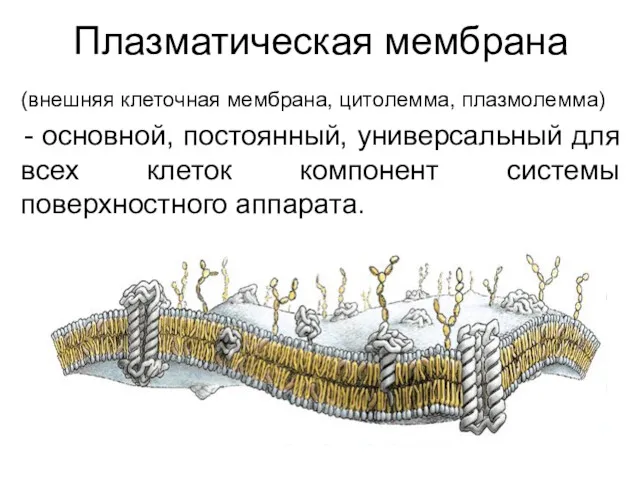
Плазматическая мембрана
(внешняя клеточная мембрана, цитолемма, плазмолемма)
основной, постоянный, универсальный для всех
Плазматическая мембрана
(внешняя клеточная мембрана, цитолемма, плазмолемма)
основной, постоянный, универсальный для всех
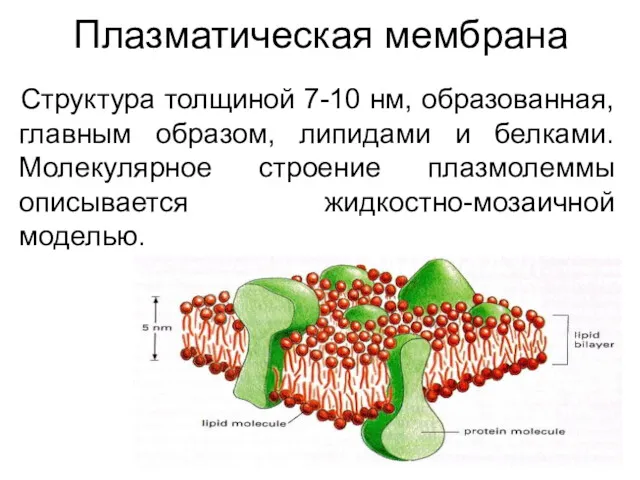
Плазматическая мембрана
Структура толщиной 7-10 нм, образованная, главным образом, липидами и белками.
Молекулярное
Плазматическая мембрана
Структура толщиной 7-10 нм, образованная, главным образом, липидами и белками. Молекулярное
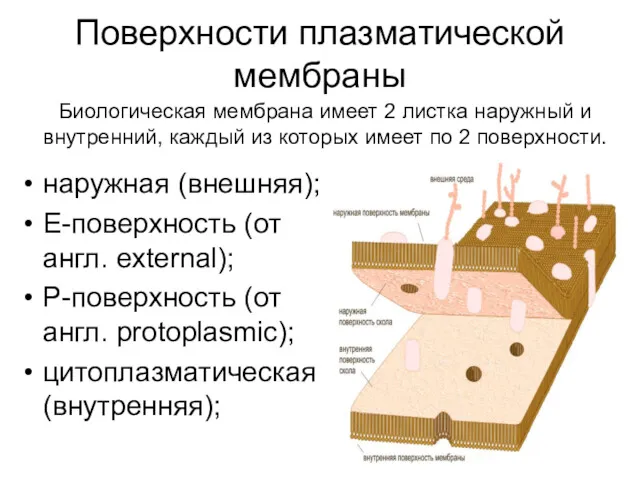
Поверхности плазматической мембраны
наружная (внешняя);
Е-поверхность (от англ. еxternal);
Р-поверхность (от англ. protoplasmic);
цитоплазматическая (внутренняя);
Биологическая
Поверхности плазматической мембраны
наружная (внешняя);
Е-поверхность (от англ. еxternal);
Р-поверхность (от англ. protoplasmic);
цитоплазматическая (внутренняя);
Биологическая
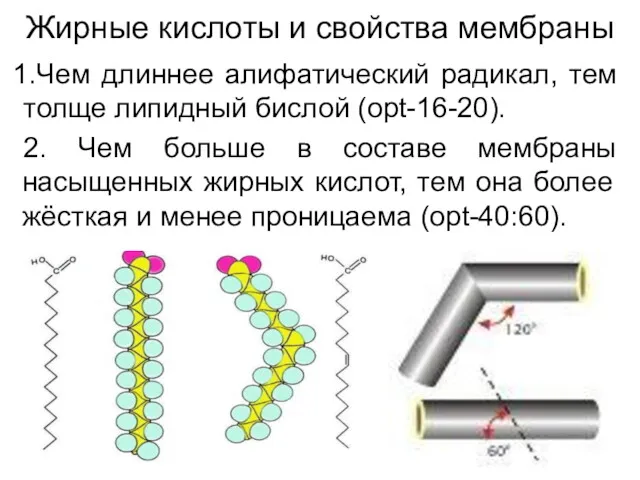
Жирные кислоты и свойства мембраны
Чем длиннее алифатический радикал, тем толще липидный
Жирные кислоты и свойства мембраны
Чем длиннее алифатический радикал, тем толще липидный

Содержание фосфолипидов в плазматической мембране
Содержание фосфолипидов в плазматической мембране
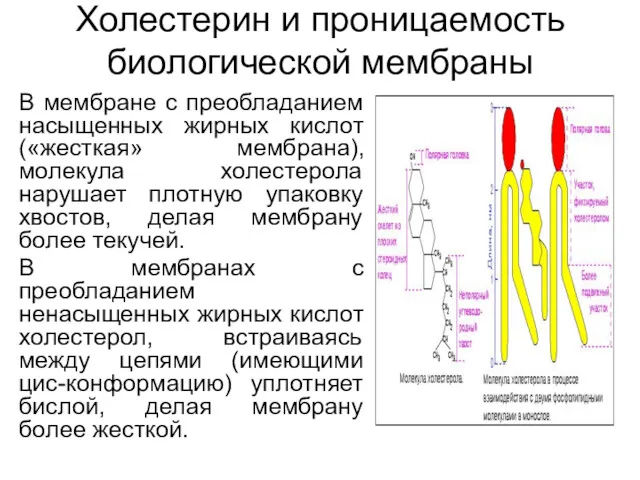
Холестерин и проницаемость биологической мембраны
В мембране с преобладанием насыщенных жирных кислот
Холестерин и проницаемость биологической мембраны
В мембране с преобладанием насыщенных жирных кислот
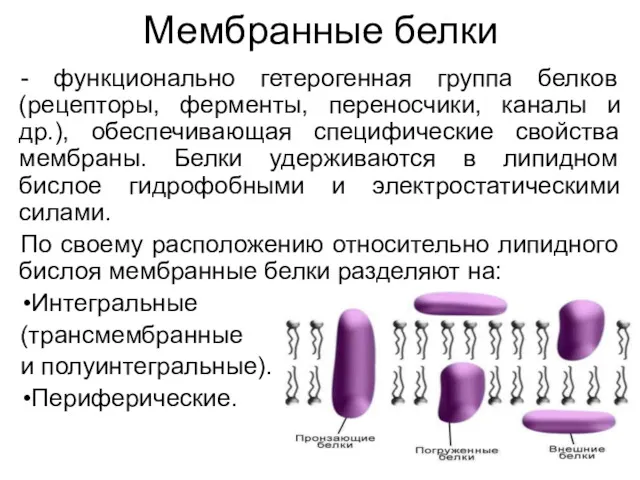
Мембранные белки
- функционально гетерогенная группа белков (рецепторы, ферменты, переносчики, каналы и
Мембранные белки
- функционально гетерогенная группа белков (рецепторы, ферменты, переносчики, каналы и
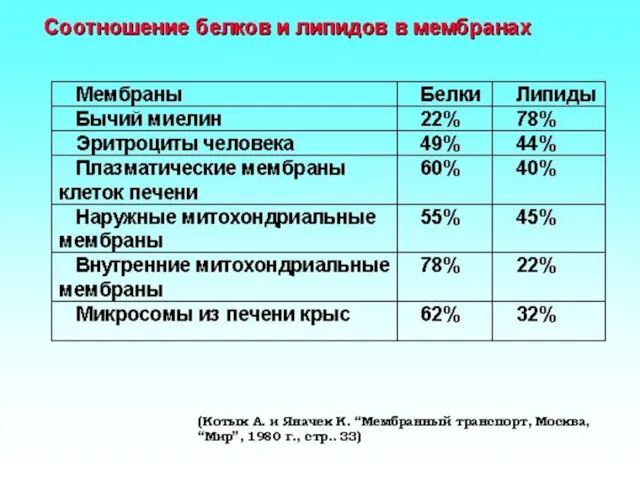

Химическая модификация мембранных липидов и белков
Молекулы олигосахаридов связываются с
липидами (гликолипиды)
Химическая модификация мембранных липидов и белков
Молекулы олигосахаридов связываются с
липидами (гликолипиды)
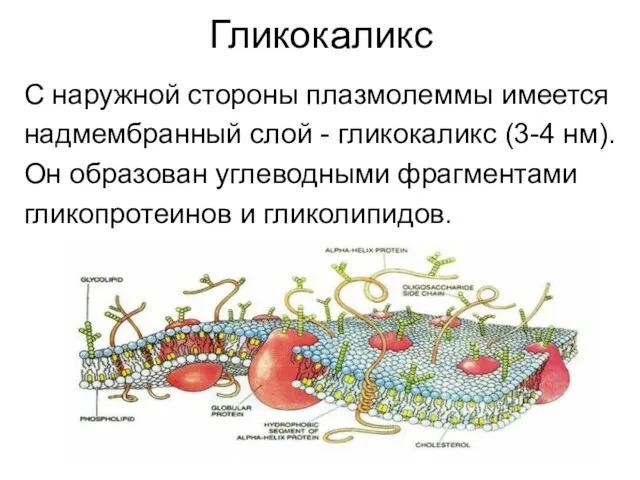
Гликокаликс
С наружной стороны плазмолеммы имеется
надмембранный слой - гликокаликс (3-4 нм).
Он
Гликокаликс
С наружной стороны плазмолеммы имеется
надмембранный слой - гликокаликс (3-4 нм).
Он
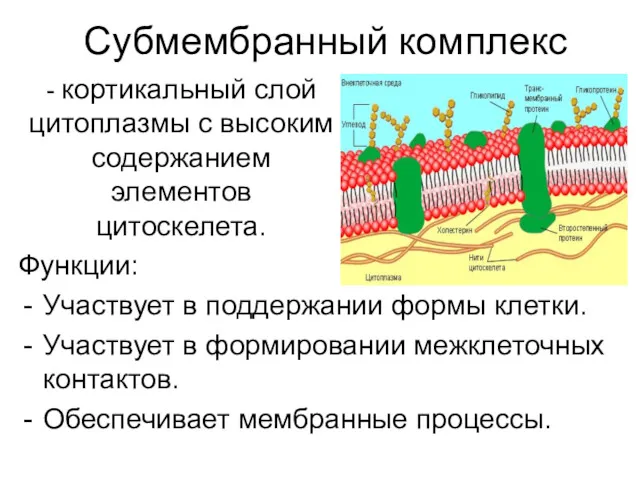
Субмембранный комплекс
Функции:
Участвует в поддержании формы клетки.
Участвует в формировании межклеточных контактов.
Обеспечивает мембранные
Субмембранный комплекс
Функции:
Участвует в поддержании формы клетки.
Участвует в формировании межклеточных контактов.
Обеспечивает мембранные

Мобильность липидов
При температуре тела мембрана текуча и
липиды свободно перемещаются
Мобильность липидов
При температуре тела мембрана текуча и
липиды свободно перемещаются

Перемещение мембранных белков
Белковые молекулы мозаично распределены в липидном бислое и перемещаться
Перемещение мембранных белков
Белковые молекулы мозаично распределены в липидном бислое и перемещаться

Асимметрия биологических мембран
Липидный состав монослоев различен. Например, в плазматической мембране фосфатидилхолин
Асимметрия биологических мембран
Липидный состав монослоев различен. Например, в плазматической мембране фосфатидилхолин
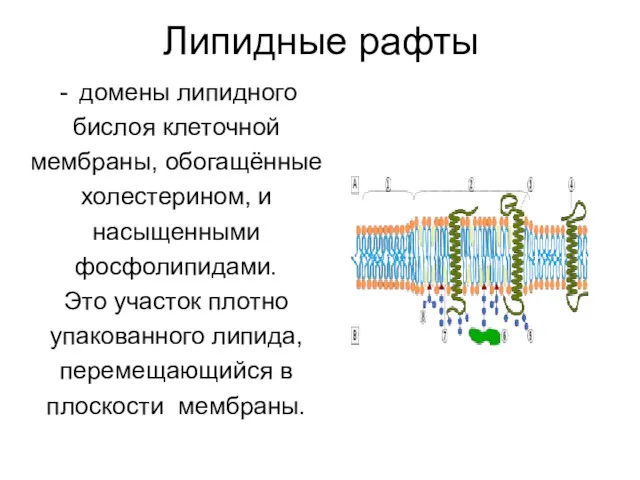
Липидные рафты
домены липидного
бислоя клеточной
мембраны, обогащённые
холестерином, и
насыщенными
фосфолипидами.
Это
Липидные рафты
домены липидного
бислоя клеточной
мембраны, обогащённые
холестерином, и
насыщенными
фосфолипидами.
Это

Эффект молекулярной памяти липидов (эф-т Л.Д.Бергельсона)
Краткосрочные события, протекающие в белковой молекуле
Эффект молекулярной памяти липидов (эф-т Л.Д.Бергельсона)
Краткосрочные события, протекающие в белковой молекуле
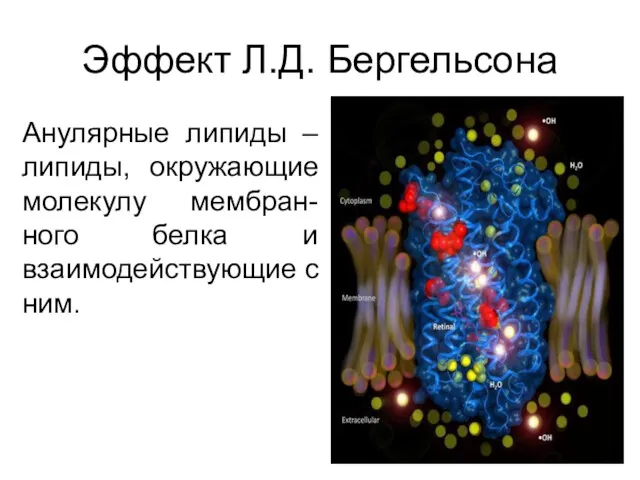
Эффект Л.Д. Бергельсона
Анулярные липиды – липиды, окружающие молекулу мембран-ного белка и
Эффект Л.Д. Бергельсона
Анулярные липиды – липиды, окружающие молекулу мембран-ного белка и

Заряд биологической мембраны
Определяется совокупностью зарядов входящих в их состав молекул липидов,
Заряд биологической мембраны
Определяется совокупностью зарядов входящих в их состав молекул липидов,

Обновление мембран в клетке
Баланс эндоцитоз/экзоцитоз для плазмолеммы.
Обмен везикулами между
Обновление мембран в клетке
Баланс эндоцитоз/экзоцитоз для плазмолеммы.
Обмен везикулами между

Функции плазмолеммы:
барьерная;
транспорт веществ в клетку и из клетки;
взаимодействие с сигнальными молекулами;
взаимодействие
Функции плазмолеммы:
барьерная;
транспорт веществ в клетку и из клетки;
взаимодействие с сигнальными молекулами;
взаимодействие

Рецепторы
гликопротеины, способные высокоселективно связываться с определенными молекулами - лигандами (гормон,
Рецепторы
гликопротеины, способные высокоселективно связываться с определенными молекулами - лигандами (гормон,
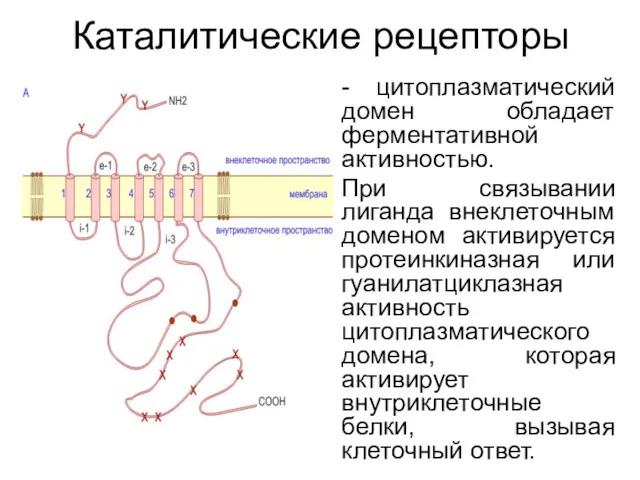
Каталитические рецепторы
- цитоплазматический домен обладает ферментативной активностью.
При связывании лиганда внеклеточным доменом
Каталитические рецепторы
- цитоплазматический домен обладает ферментативной активностью.
При связывании лиганда внеклеточным доменом
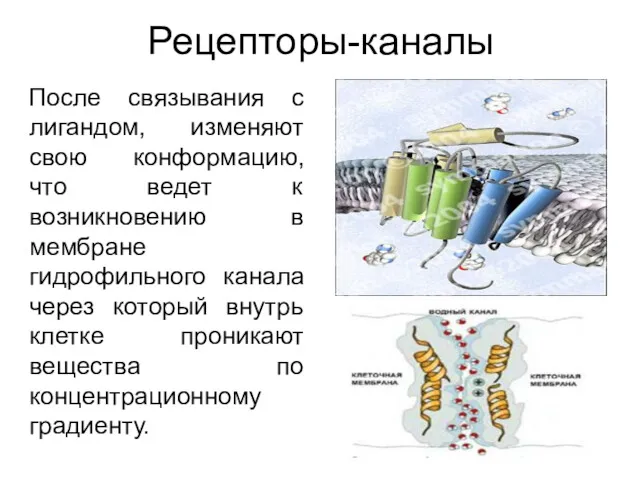
Рецепторы-каналы
После связывания с лигандом, изменяют свою конформацию, что ведет к возникновению
Рецепторы-каналы
После связывания с лигандом, изменяют свою конформацию, что ведет к возникновению
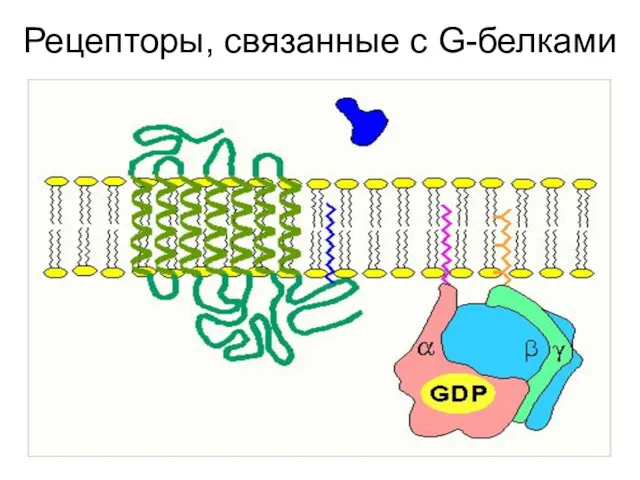
Рецепторы, связанные с G-белками
Рецепторы, связанные с G-белками

Трансмембранный транспорт веществ
прямой (диффузия, осмос, фильтрация);
опосредованный:
1. пассивный (с участием белка-переносчика
Трансмембранный транспорт веществ
прямой (диффузия, осмос, фильтрация);
опосредованный:
1. пассивный (с участием белка-переносчика

Простая диффузия
- переход вещества из области более высокой концентрации в область
Простая диффузия
- переход вещества из области более высокой концентрации в область
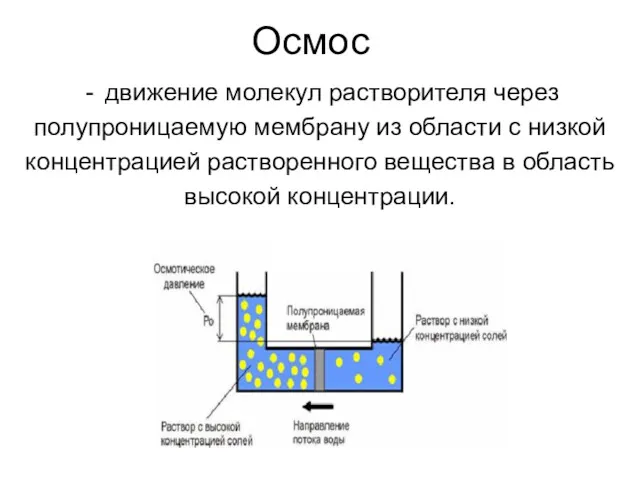
Осмос
движение молекул растворителя через
полупроницаемую мембрану из области с низкой
концентрацией
Осмос
движение молекул растворителя через
полупроницаемую мембрану из области с низкой
концентрацией
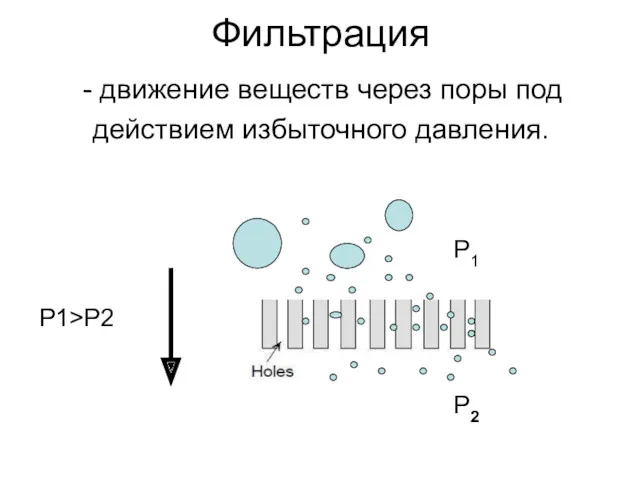
Фильтрация
движение веществ через поры под
действием избыточного давления.
Фильтрация
движение веществ через поры под
действием избыточного давления.

Облегчённая диффузия
транспорт веществ через мембрану с помощью специальных белков.
Облегчённая диффузия
транспорт веществ через мембрану с помощью специальных белков.
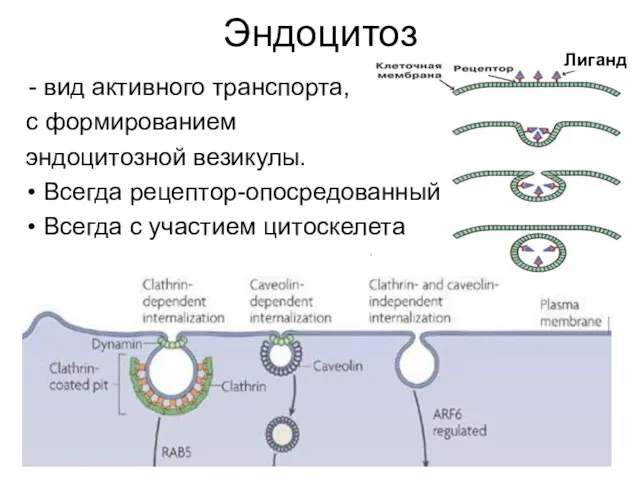
Эндоцитоз
вид активного транспорта,
с формированием
эндоцитозной везикулы.
Всегда рецептор-опосредованный
Всегда
Эндоцитоз
вид активного транспорта,
с формированием
эндоцитозной везикулы.
Всегда рецептор-опосредованный
Всегда

Виды эндоцитоза
Фагоцитоз - процесс поглощения клеткой объектов размером >10-6 м,
Виды эндоцитоза
Фагоцитоз - процесс поглощения клеткой объектов размером >10-6 м,
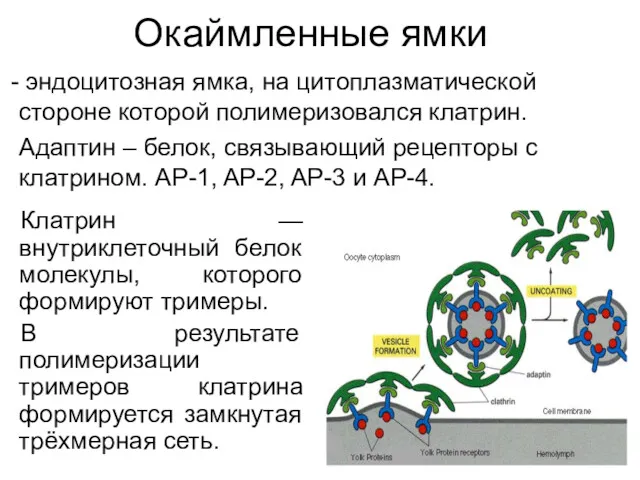
Окаймленные ямки
Клатрин — внутриклеточный белок молекулы, которого формируют тримеры.
В результате полимеризации
Окаймленные ямки
Клатрин — внутриклеточный белок молекулы, которого формируют тримеры.
В результате полимеризации

Кавеолы
- углубление плазмолеммы, покрытое с цитоплазматической стороны кавеолинами.
Кавеолины — группа мембранных
Кавеолы
- углубление плазмолеммы, покрытое с цитоплазматической стороны кавеолинами.
Кавеолины — группа мембранных

Динамин
- эукариотическая клеточная ГТФ-аза, участвующая в эндоцитозе. Динамин необходим для отделения
Динамин
- эукариотическая клеточная ГТФ-аза, участвующая в эндоцитозе. Динамин необходим для отделения
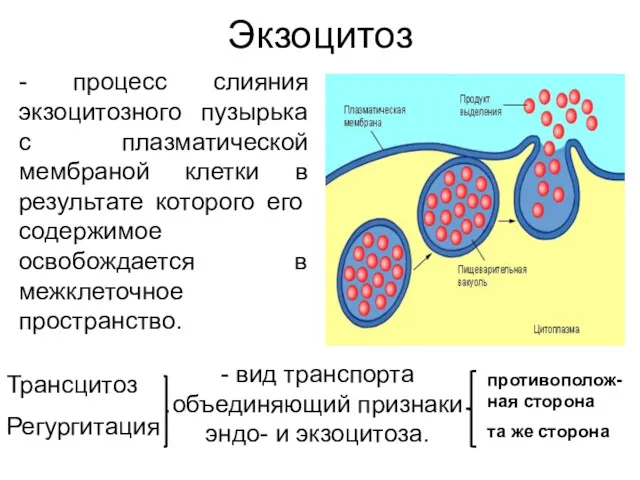
Экзоцитоз
- процесс слияния экзоцитозного пузырька с плазматической мембраной клетки в результате
Экзоцитоз
- процесс слияния экзоцитозного пузырька с плазматической мембраной клетки в результате
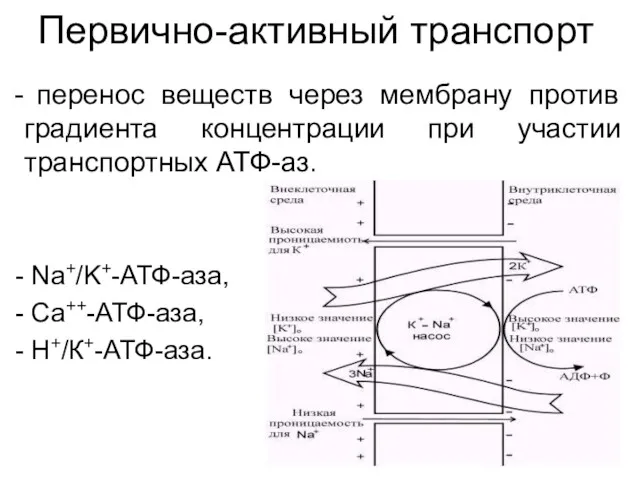
Первично-активный транспорт
перенос веществ через мембрану против градиента концентрации при участии
Первично-активный транспорт
перенос веществ через мембрану против градиента концентрации при участии
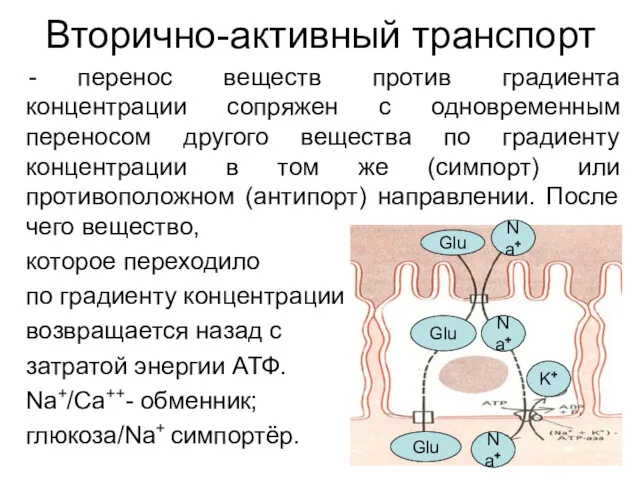
Вторично-активный транспорт
перенос веществ против градиента концентрации сопряжен с одновременным переносом
Вторично-активный транспорт
перенос веществ против градиента концентрации сопряжен с одновременным переносом
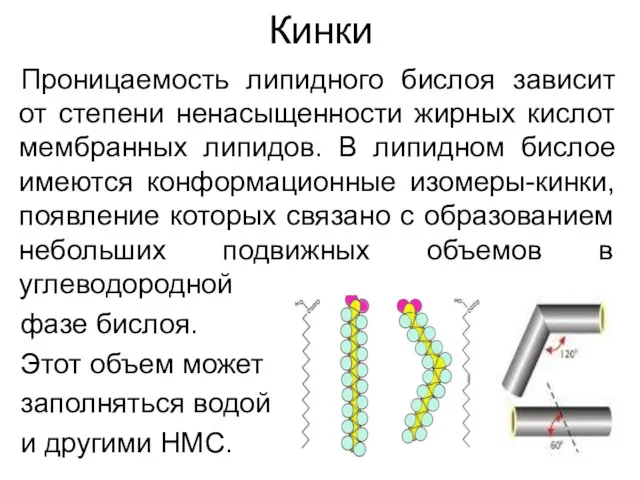
Кинки
Проницаемость липидного бислоя зависит от степени ненасыщенности жирных кислот мембранных липидов.
Кинки
Проницаемость липидного бислоя зависит от степени ненасыщенности жирных кислот мембранных липидов.

Межклеточные контакты
Механические – контакты, которые обеспечивают механическую связь клеток
Межклеточные контакты
Механические – контакты, которые обеспечивают механическую связь клеток

Интердигитации
Наиболее просто устроенный вид взаимодействия мембран смежных клеток. Представлены выпячиваниями цитоплазмы
Интердигитации
Наиболее просто устроенный вид взаимодействия мембран смежных клеток. Представлены выпячиваниями цитоплазмы
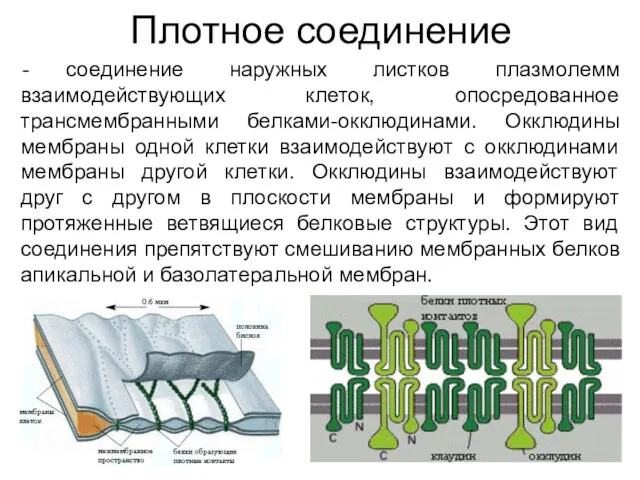
Плотное соединение
соединение наружных листков плазмолемм взаимодействующих клеток, опосредованное трансмембранными
Плотное соединение
соединение наружных листков плазмолемм взаимодействующих клеток, опосредованное трансмембранными

Промежуточное соединение, опоясывающая десмосома
Охватывают клетку по периметру в виде пояса. С
Промежуточное соединение, опоясывающая десмосома
Охватывают клетку по периметру в виде пояса. С
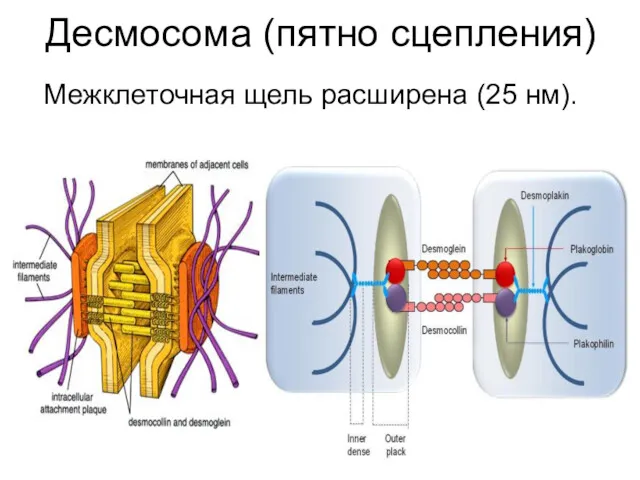
Десмосома (пятно сцепления)
Межклеточная щель расширена (25 нм).
Десмосома (пятно сцепления)
Межклеточная щель расширена (25 нм).

Кадхерины
Кадхерины «зоны слипания»:
Е-кадхерин (эпителиальный кадхерин, увоморулин, L-CAM).
N-кадхерин (нейральный
Кадхерины
Кадхерины «зоны слипания»:
Е-кадхерин (эпителиальный кадхерин, увоморулин, L-CAM).
N-кадхерин (нейральный
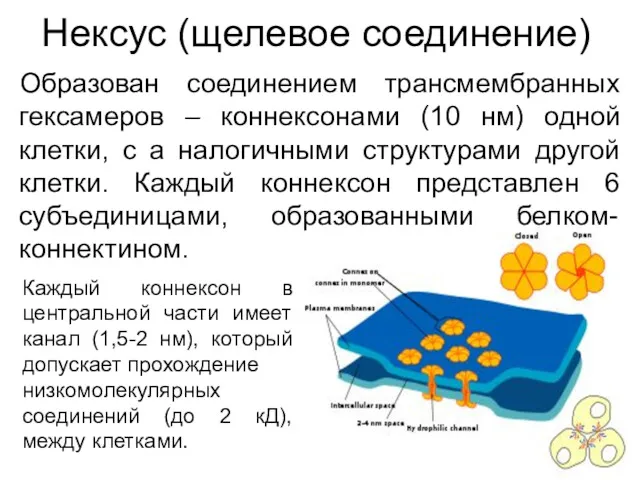
Нексус (щелевое соединение)
Образован соединением трансмембранных гексамеров – коннексонами (10 нм) одной
Нексус (щелевое соединение)
Образован соединением трансмембранных гексамеров – коннексонами (10 нм) одной
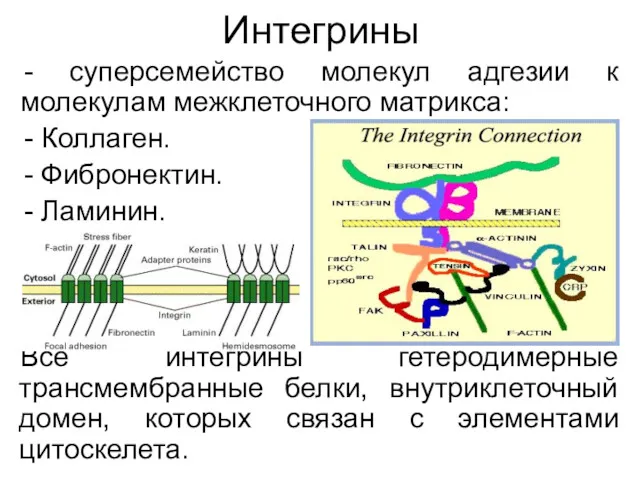
Интегрины
суперсемейство молекул адгезии к молекулам межклеточного матрикса:
Коллаген.
Фибронектин.
Ламинин.
Все
Интегрины
суперсемейство молекул адгезии к молекулам межклеточного матрикса:
Коллаген.
Фибронектин.
Ламинин.
Все
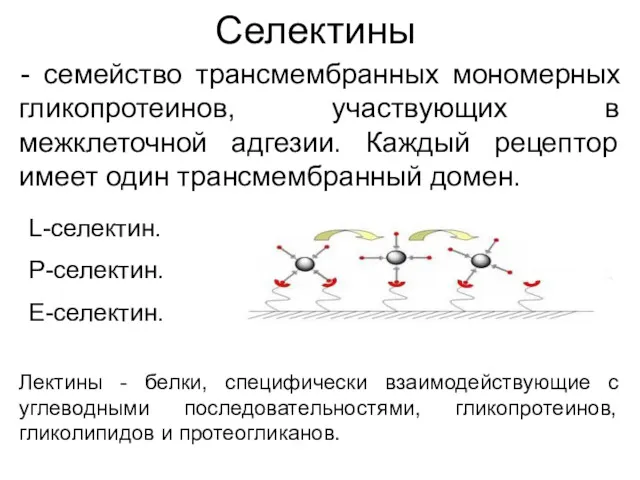
Селектины
- семейство трансмембранных мономерных гликопротеинов, участвующих в межклеточной адгезии. Каждый рецептор
Селектины
- семейство трансмембранных мономерных гликопротеинов, участвующих в межклеточной адгезии. Каждый рецептор
 Биотехнология
Биотехнология Царство protista - простейшие. Надтип sarcodina - саркодовые
Царство protista - простейшие. Надтип sarcodina - саркодовые Паразитические плоские черви. Верно ли утверждение?
Паразитические плоские черви. Верно ли утверждение? Биологическое действие ионизирующих излучений
Биологическое действие ионизирующих излучений Низкомолекулярные органические соединения витамины
Низкомолекулярные органические соединения витамины Древесная лягушка
Древесная лягушка Разработка урока по биологии для 6 класса
Разработка урока по биологии для 6 класса Пищеварение в тонком кишечнике
Пищеварение в тонком кишечнике Неотропикалық патшалық
Неотропикалық патшалық Направления эволюции
Направления эволюции Carbohydrates
Carbohydrates Деление клетки. Митоз
Деление клетки. Митоз Regnul plante
Regnul plante Биохимия гормонов
Биохимия гормонов Сүйек тінінің және тіс тінінің биохимиясы
Сүйек тінінің және тіс тінінің биохимиясы Разнообразие живых организмов. 5 класс
Разнообразие живых организмов. 5 класс Деление клетки. Мейоз
Деление клетки. Мейоз Эндокринная система: железы внешней, внутренней и смешанной секреции
Эндокринная система: железы внешней, внутренней и смешанной секреции Дагестанский заповедник у Кизлярского залива
Дагестанский заповедник у Кизлярского залива Биология. Классическая биология. Физико-химическая биология. Эволюционная биология
Биология. Классическая биология. Физико-химическая биология. Эволюционная биология Новосибирсий зоопарк
Новосибирсий зоопарк Биологиялық жүйелерді зерттеудің дыбыстық әдістері. Акустика. (Дәріс 7)
Биологиялық жүйелерді зерттеудің дыбыстық әдістері. Акустика. (Дәріс 7) Презентация по теме Законы Менделя
Презентация по теме Законы Менделя Органы и системы органов человека
Органы и системы органов человека Нуклеиновые кислоты
Нуклеиновые кислоты Механорецепторы кожи
Механорецепторы кожи Общая характеристика подтипа Позвоночные. Часть1
Общая характеристика подтипа Позвоночные. Часть1 Сердечно-сосудистая система
Сердечно-сосудистая система