- Функциональная морфология синтетического аппарата. Рибосомы

Содержание
- 2. Рибосомы - немембранная органелла клетки (10-30 нм), обеспечивающая трансляцию.
- 3. 80S рибосома Неактивные рибосомы диссоциируют и располагаются в цитозоле в виде отдельных субъединиц. Рибосомальные белки рРНК
- 4. Строение 80S рибосомы
- 5. Синтез рибосом
- 6. Сборка рибосом в цитоплазме В присутствии мРНК малая субъединица связывается с её 5‘- концом, после чего
- 7. Центры рибосомы Аминоацильный – А. Пептидильный – Р. Формируют обе субъединицы. Оба центра включают участок мРНК
- 8. Полисома (полирибосома) несколько рибосом, объединенных общей мРНК. Рибосомы в составе полисомы – транслирующие. Неактивные рибосомы располагаются
- 9. Протеасома белковый комплекс, осуществляющий разрушение цитоплазматических белков. В эукариотических клетках протеасомы содержатся и в ядре и
- 10. Синтез 20S протеасом Коровая частица, 700kDa. Обеспечивает АТФ и убиквитин-независимый протеолиз.
- 11. 26S протеасомы 20S+2*(19S)=26S 19S частица служит для распознавания субстрата и денатурации белка. 26S протеасома обеспечивает АТФ-
- 12. Эндоплазматический ретикулум Описан К. Портером в 1945 г. - постоянная органелла эукариотической клетки, представляющая собой разветвлённую
- 13. Шероховатый ЭПР - ЭПР на цитоплазматической поверхности, которой расположены рибосомы.
- 14. Функции грЭПР Биосинтез всех мембранных и экспортных белков. Посттрансляционные изменения белков (обеспечение фолдинга, химическая модификация).
- 15. Гладкий ЭПР, микросомы представляет собой ЭПР на поверхности которого отсутствуют рибосомы. фрагменты ЭПР (везикулы D 100
- 16. Функции аЭПР Синтез липидов. Метаболизм углеводов. Восстановление кариолеммы в телофазе. Накопление ионов Са++ (кальретикулин, кальсеквестрин). Концентрация
- 17. Метаболизм липидов и углеводов Метаболизм липидов: Синтез холестерола (ГМГ-КоА-редуктаза). Синтез фосфолипидов. Ферменты синтеза глико- и сфинголипидов
- 18. ERGIC Еndoplasmicum Reticulum - Golgi Intermediate Complex совокупность везикул, обеспечивающих взаимодействие между ЭПР и цис- поверхностью
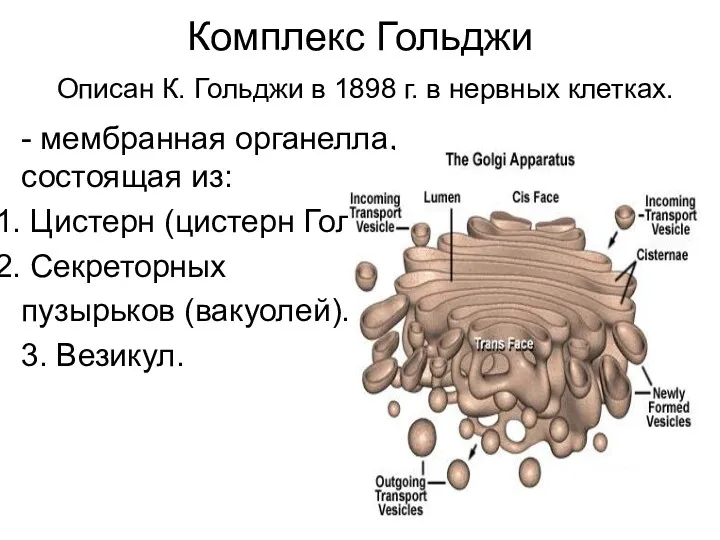
- 19. Комплекс Гольджи Описан К. Гольджи в 1898 г. в нервных клетках. - мембранная органелла, состоящая из:
- 20. Диктиосома (поле Гольджи) - основная часть комплекса Гольджи, представленная уложенными параллельно друг другу цистернами Гольджи. Комплекс
- 21. Полярность диктиосомы Цис (незрелая) - выпуклая, обращенная к ЭПР и связанную с последней системой везикул –
- 22. Строение цистерны Гольджи - изогнутые, уложенные стопкой диски (3-30 шт.) диаметром 0,5-5 мкм разделенные пространством (20-40
- 23. Везикулы и секреторные пузырьки Везикулы - сферические элементы (D 40-80 нм), образующиеся путем отщепления от цистерн.
- 24. Функции комплекса Гольджи Формирование секреторных гранул. Обновление плазмолеммы. Процессинг белков (гликозилирование, фосфорилирование, ацилирование, сульфатирование, частичный протеолиз).
- 25. Транскрипция. Транскриптон. Транскрипция - синтез гяРНК по матрице ДНК. Транскриптон – участок ДНК, ограниченный промотором и
- 26. Инициация-элонгация-терминация транкрипции ТАТА-фактор, взаимодействует с ТАТА-боксом, после чего происходит присоединение РНК-полимеразы. Факторы инициации вызывают раскручивание одного
- 27. Процессинг первичного транскрипта Сплайсинг. Кэпирование. Полиаденилирование. Модификация азотистых оснований. зрелая РНК
- 28. Сплайсинг процесс удаления интронов из гяРНК, с последующим соединением экзонов и формированием мРНК. Сплайсосома – нуклеопротеиновый
- 29. Варианты сплайсинга Неферментативный (аутосплайсинг - вид сплайсинга, при котором интроны являются рибозимами). Ферментативный: 1. С участием
- 30. Кэпирование пре-мРНК Присоединение гуанилилтрансферазой к 5’-концу синтезирующейся пре-мРНК (≈ 30 нуклеотидов) 7-метилгуанозина – кэпа. Биологическая роль:
- 31. Полиаденилирование Присоединение полиаденилатполимеразой к 3’-концу пре-мРНК поли(А)-фрагмента (≈200 адениловых нуклеотидов). Сигналом к полиаденилированию является – ААUААА.
- 32. Ковалентная модификация азотистых оснований РНК Дезаминирование цитозина (урацил). Редактирование пре-мРНК апопротеина В в энтероцитах тонкого кишечника.
- 33. Трансляция синтез полипептида рибосомами, используя матричную РНК. Прочтение мРНК рибосомой идёт в направлении от 5‘- к
- 34. Инициация трансляции Происходит образование комплекса мРНК, рибосомы и инициирующей тРНК (fMet-, Met-тРНК). Участвуют eIF (более 10).
- 35. Элонгация трансляции В Р-центре – AUG-кодон, соединенный с Met-tRNAi, в А-центре – триплет, кодирующий включение первой
- 36. Терминация трансляции В А-центре рибосомы оказывается стор-кодон: UAG, UAA,UGA. Для стоп-кодонов нет соответствующей тРНК. К рибосоме
- 37. Фолдинг процесс принятия функционально активной конформации биополимерами внутри клетки при помощи белков – факторов фолдинга. Факторы
- 38. Шапероны класс белков, придающих, стабилизирующих и восстанавливающие конформацию других белков. Конститутивные Индуцибельные (белки теплового шока, НSP)
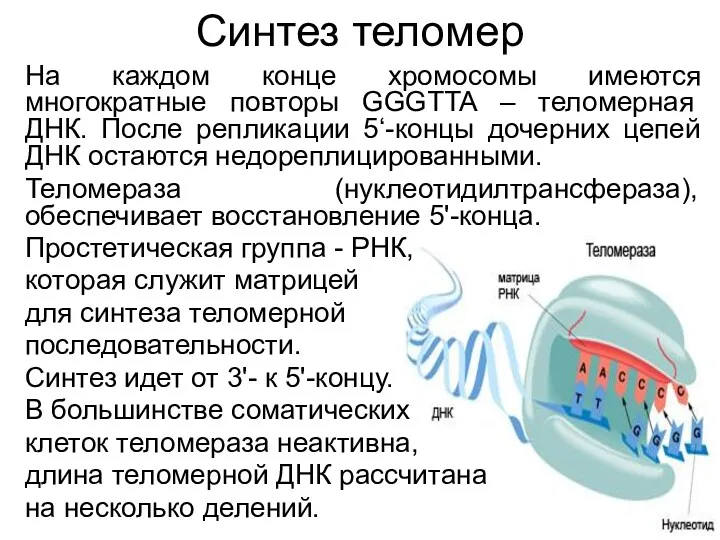
- 39. Синтез теломер На каждом конце хромосомы имеются многократные повторы GGGTTA – теломерная ДНК. После репликации 5‘-концы
- 41. Скачать презентацию
Рибосомы
- немембранная органелла клетки (10-30 нм), обеспечивающая трансляцию.
Рибосомы
- немембранная органелла клетки (10-30 нм), обеспечивающая трансляцию.
80S рибосома
Неактивные рибосомы диссоциируют и располагаются в цитозоле в виде отдельных субъединиц.
Рибосомальные белки
рРНК
80S рибосома
Неактивные рибосомы диссоциируют и располагаются в цитозоле в виде отдельных субъединиц.
Рибосомальные белки
рРНК

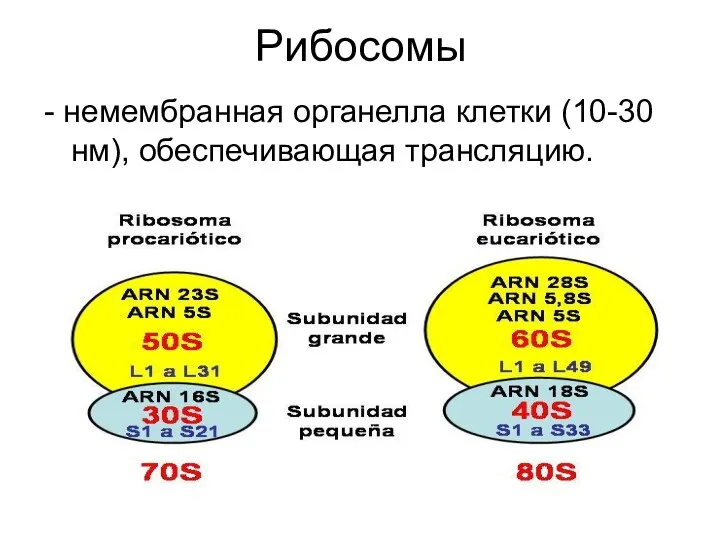
Строение 80S рибосомы
Строение 80S рибосомы
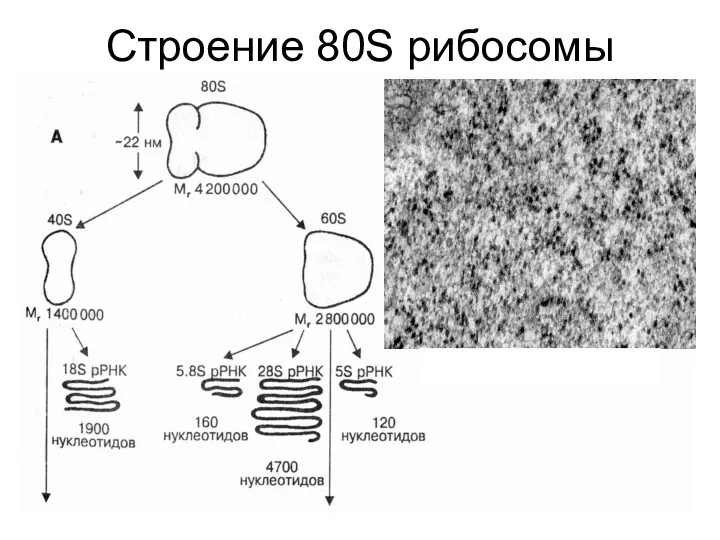
Синтез рибосом
Синтез рибосом
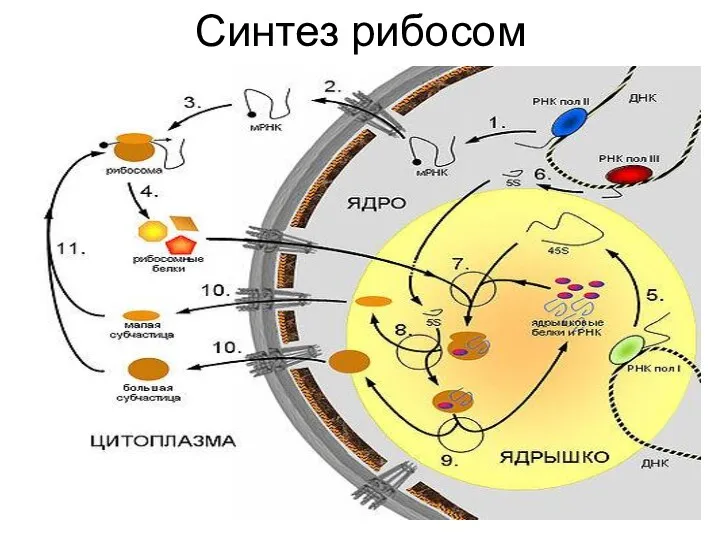
Сборка рибосом в цитоплазме
В присутствии мРНК
малая субъединица
связывается с её 5‘-
концом, после
Сборка рибосом в цитоплазме
В присутствии мРНК
малая субъединица
связывается с её 5‘-
концом, после

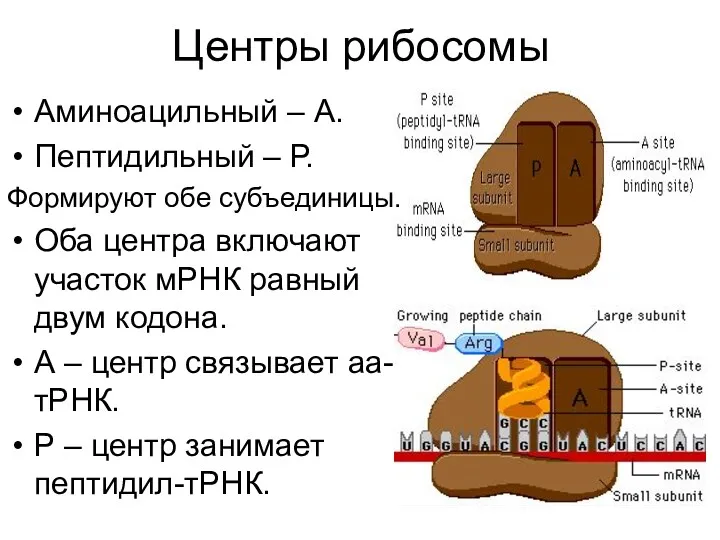
Центры рибосомы
Аминоацильный – А.
Пептидильный – Р.
Формируют обе субъединицы.
Оба центра включают участок мРНК равный
Центры рибосомы
Аминоацильный – А.
Пептидильный – Р.
Формируют обе субъединицы.
Оба центра включают участок мРНК равный
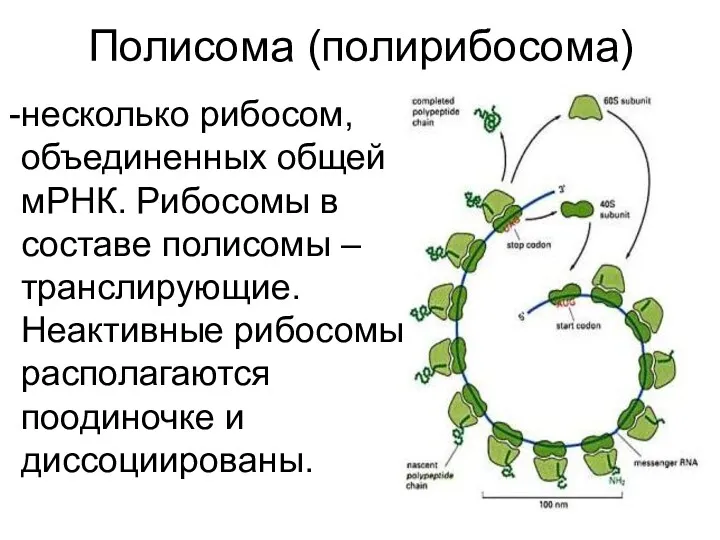
Полисома (полирибосома)
несколько рибосом, объединенных общей мРНК. Рибосомы в составе полисомы – транслирующие. Неактивные
Полисома (полирибосома)
несколько рибосом, объединенных общей мРНК. Рибосомы в составе полисомы – транслирующие. Неактивные
Протеасома
белковый комплекс, осуществляющий разрушение цитоплазматических белков.
В эукариотических клетках протеасомы содержатся и в
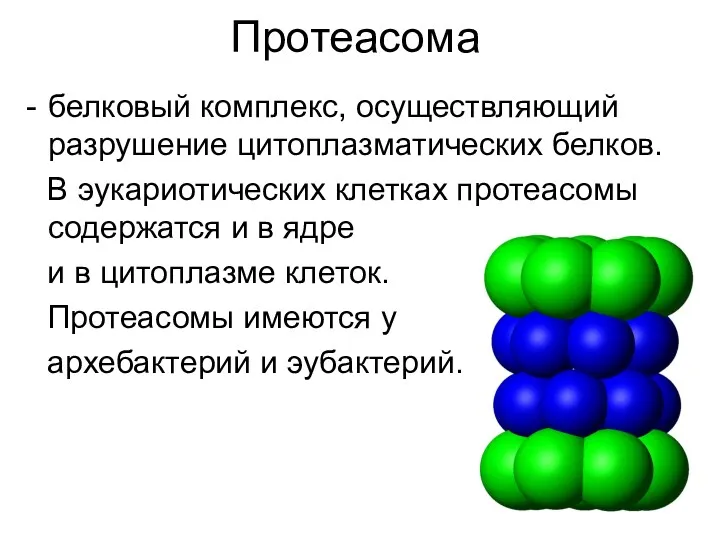
Протеасома
белковый комплекс, осуществляющий разрушение цитоплазматических белков.
В эукариотических клетках протеасомы содержатся и в
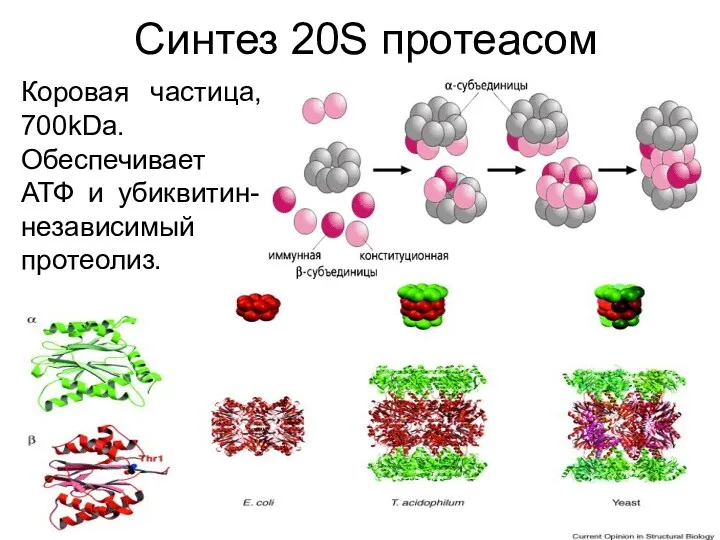
Синтез 20S протеасом
Коровая частица, 700kDa. Обеспечивает АТФ и убиквитин-независимый протеолиз.
Синтез 20S протеасом
Коровая частица, 700kDa. Обеспечивает АТФ и убиквитин-независимый протеолиз.
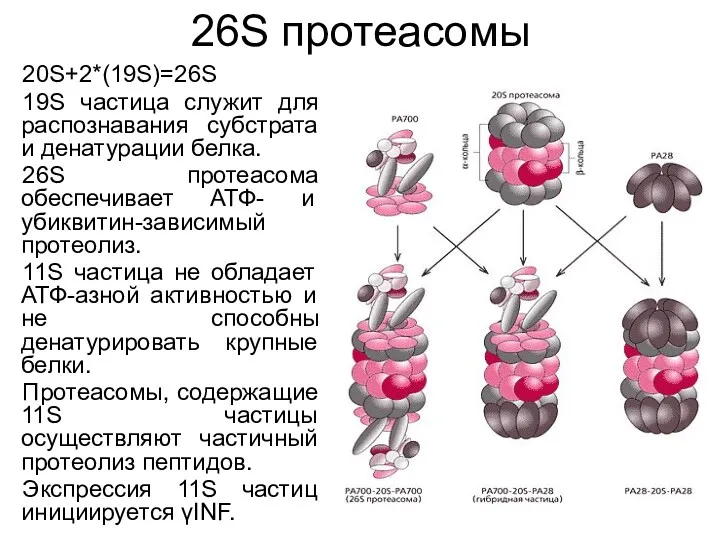
26S протеасомы
20S+2*(19S)=26S
19S частица служит для распознавания субстрата и денатурации белка.
26S протеасома обеспечивает АТФ-
26S протеасомы
20S+2*(19S)=26S
19S частица служит для распознавания субстрата и денатурации белка.
26S протеасома обеспечивает АТФ-
Эндоплазматический ретикулум Описан К. Портером в 1945 г.
- постоянная органелла эукариотической клетки,
Эндоплазматический ретикулум Описан К. Портером в 1945 г.
- постоянная органелла эукариотической клетки,

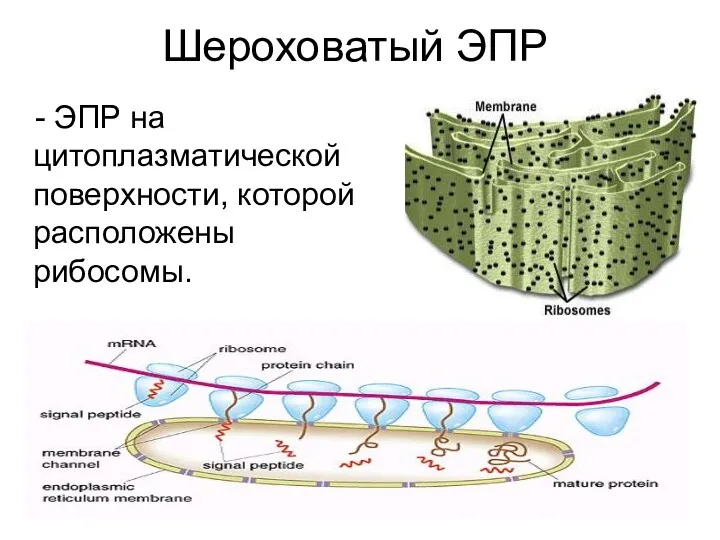
Шероховатый ЭПР
- ЭПР на цитоплазматической поверхности, которой расположены рибосомы.
Шероховатый ЭПР
- ЭПР на цитоплазматической поверхности, которой расположены рибосомы.
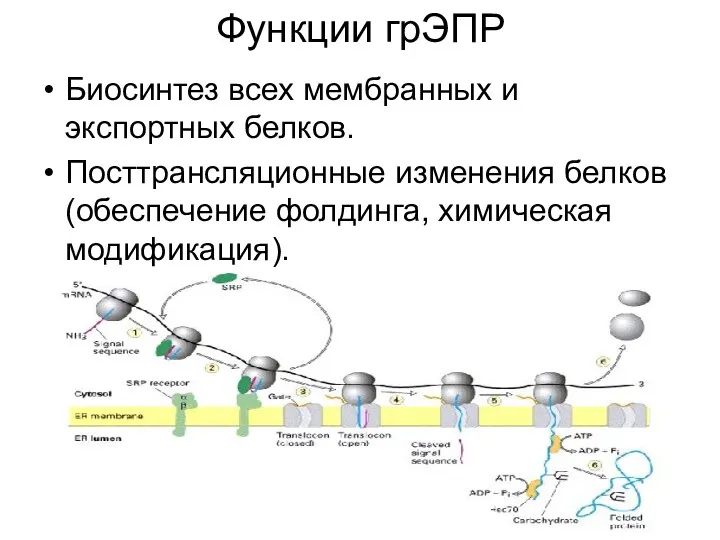
Функции грЭПР
Биосинтез всех мембранных и экспортных белков.
Посттрансляционные изменения белков (обеспечение фолдинга, химическая модификация).
Функции грЭПР
Биосинтез всех мембранных и экспортных белков.
Посттрансляционные изменения белков (обеспечение фолдинга, химическая модификация).
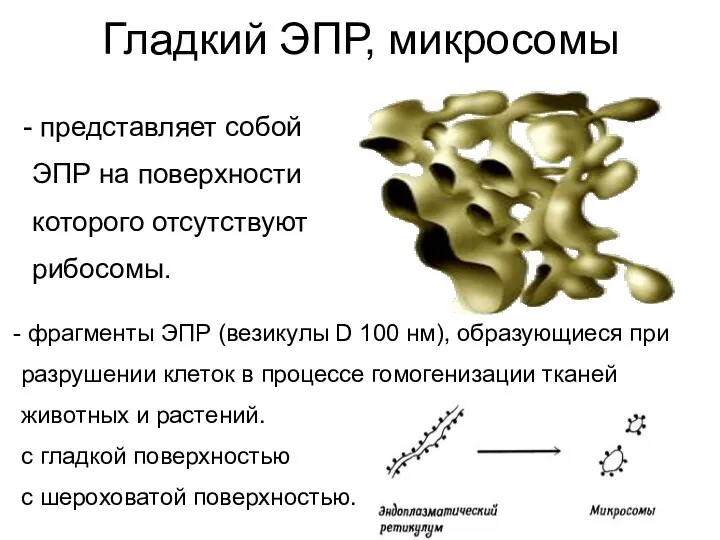
Гладкий ЭПР, микросомы
представляет собой
ЭПР на поверхности
которого отсутствуют
рибосомы.
фрагменты ЭПР (везикулы D
Гладкий ЭПР, микросомы
представляет собой
ЭПР на поверхности
которого отсутствуют
рибосомы.
фрагменты ЭПР (везикулы D
Функции аЭПР
Синтез липидов.
Метаболизм углеводов.
Восстановление кариолеммы в телофазе.
Накопление ионов
Функции аЭПР
Синтез липидов.
Метаболизм углеводов.
Восстановление кариолеммы в телофазе.
Накопление ионов

Метаболизм липидов и углеводов
Метаболизм липидов:
Синтез холестерола (ГМГ-КоА-редуктаза).
Синтез фосфолипидов.
Ферменты
Метаболизм липидов и углеводов
Метаболизм липидов:
Синтез холестерола (ГМГ-КоА-редуктаза).
Синтез фосфолипидов.
Ферменты

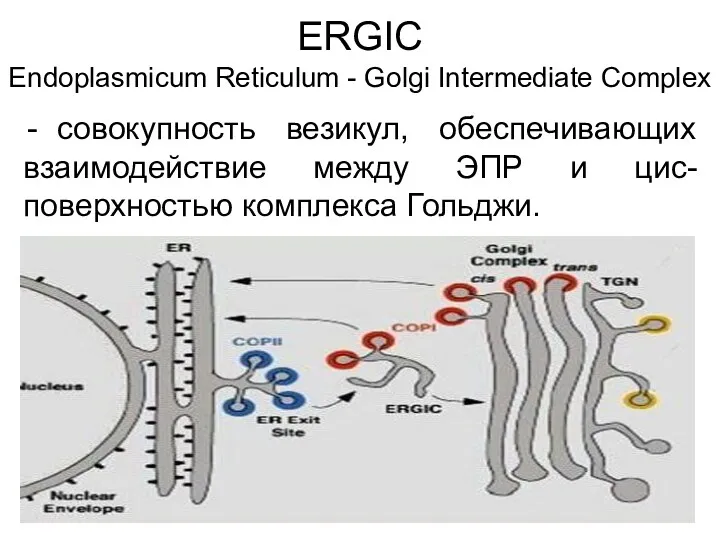
ERGIC
Еndoplasmicum Reticulum - Golgi Intermediate Complex
совокупность везикул, обеспечивающих взаимодействие между ЭПР и
ERGIC
Еndoplasmicum Reticulum - Golgi Intermediate Complex
совокупность везикул, обеспечивающих взаимодействие между ЭПР и
Комплекс Гольджи
Описан К. Гольджи в 1898 г. в нервных клетках.
- мембранная органелла,
Комплекс Гольджи
Описан К. Гольджи в 1898 г. в нервных клетках.
- мембранная органелла,
Диктиосома (поле Гольджи)
- основная часть комплекса Гольджи, представленная уложенными параллельно друг другу цистернами
Диктиосома (поле Гольджи)
- основная часть комплекса Гольджи, представленная уложенными параллельно друг другу цистернами

Полярность диктиосомы
Цис (незрелая) - выпуклая, обращенная к ЭПР и связанную с последней
Полярность диктиосомы
Цис (незрелая) - выпуклая, обращенная к ЭПР и связанную с последней

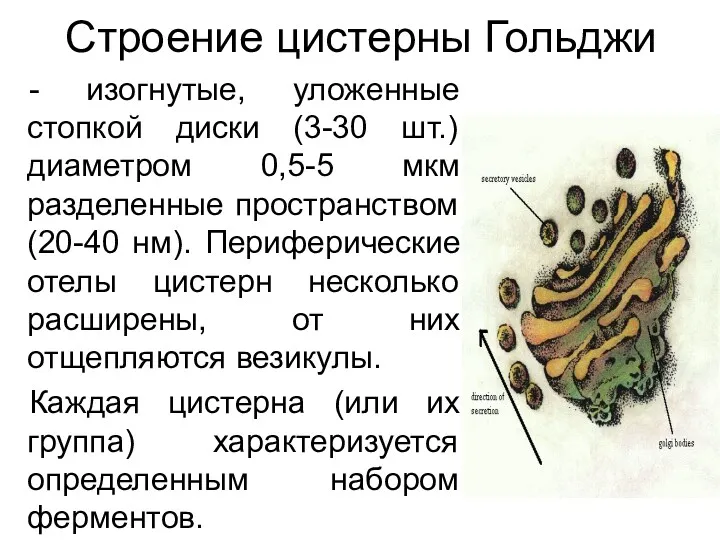
Строение цистерны Гольджи
- изогнутые, уложенные стопкой диски (3-30 шт.) диаметром 0,5-5 мкм разделенные
Строение цистерны Гольджи
- изогнутые, уложенные стопкой диски (3-30 шт.) диаметром 0,5-5 мкм разделенные
Везикулы и секреторные пузырьки
Везикулы - сферические элементы (D 40-80 нм), образующиеся путем отщепления
Везикулы и секреторные пузырьки
Везикулы - сферические элементы (D 40-80 нм), образующиеся путем отщепления

Функции комплекса Гольджи
Формирование секреторных гранул.
Обновление плазмолеммы.
Процессинг белков (гликозилирование, фосфорилирование, ацилирование, сульфатирование, частичный протеолиз).
Синтез
Функции комплекса Гольджи
Формирование секреторных гранул.
Обновление плазмолеммы.
Процессинг белков (гликозилирование, фосфорилирование, ацилирование, сульфатирование, частичный протеолиз).
Синтез

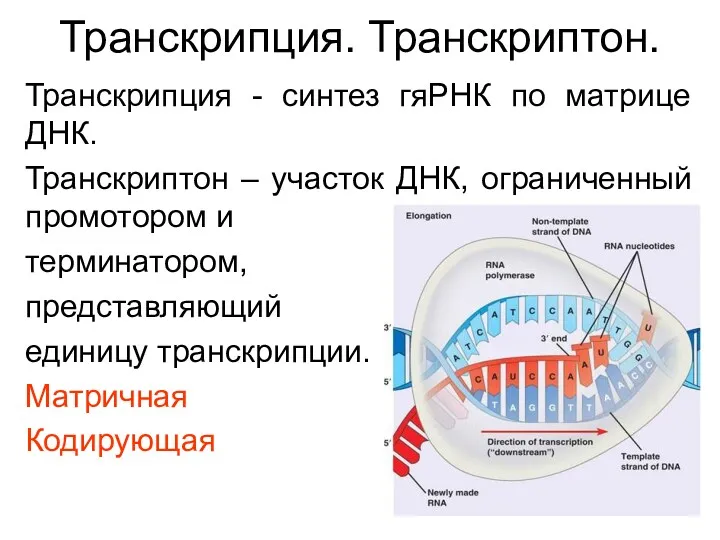
Транскрипция. Транскриптон.
Транскрипция - синтез гяРНК по матрице ДНК.
Транскриптон – участок ДНК, ограниченный промотором
Транскрипция. Транскриптон.
Транскрипция - синтез гяРНК по матрице ДНК.
Транскриптон – участок ДНК, ограниченный промотором
Инициация-элонгация-терминация транкрипции
ТАТА-фактор, взаимодействует с ТАТА-боксом, после чего происходит присоединение РНК-полимеразы. Факторы инициации вызывают
Инициация-элонгация-терминация транкрипции
ТАТА-фактор, взаимодействует с ТАТА-боксом, после чего происходит присоединение РНК-полимеразы. Факторы инициации вызывают

Процессинг первичного транскрипта
Сплайсинг.
Кэпирование.
Полиаденилирование.
Модификация азотистых
оснований.
зрелая РНК
Процессинг первичного транскрипта
Сплайсинг.
Кэпирование.
Полиаденилирование.
Модификация азотистых
оснований.
зрелая РНК
Сплайсинг
процесс удаления интронов из гяРНК, с последующим соединением экзонов и формированием мРНК.
Сплайсосома
Сплайсинг
процесс удаления интронов из гяРНК, с последующим соединением экзонов и формированием мРНК.
Сплайсосома

Варианты сплайсинга
Неферментативный (аутосплайсинг - вид сплайсинга, при котором интроны являются рибозимами).
Ферментативный:
1.
Варианты сплайсинга
Неферментативный (аутосплайсинг - вид сплайсинга, при котором интроны являются рибозимами).
Ферментативный:
1.

Кэпирование пре-мРНК
Присоединение гуанилилтрансферазой к 5’-концу синтезирующейся пре-мРНК (≈ 30 нуклеотидов) 7-метилгуанозина –
Кэпирование пре-мРНК
Присоединение гуанилилтрансферазой к 5’-концу синтезирующейся пре-мРНК (≈ 30 нуклеотидов) 7-метилгуанозина –

Полиаденилирование
Присоединение полиаденилатполимеразой к 3’-концу пре-мРНК поли(А)-фрагмента (≈200 адениловых нуклеотидов). Сигналом к полиаденилированию
Полиаденилирование
Присоединение полиаденилатполимеразой к 3’-концу пре-мРНК поли(А)-фрагмента (≈200 адениловых нуклеотидов). Сигналом к полиаденилированию

Ковалентная модификация азотистых оснований РНК
Дезаминирование цитозина (урацил). Редактирование пре-мРНК апопротеина В в
Ковалентная модификация азотистых оснований РНК
Дезаминирование цитозина (урацил). Редактирование пре-мРНК апопротеина В в

Трансляция
синтез полипептида
рибосомами, используя
матричную РНК.
Прочтение мРНК
рибосомой идёт
в направлении
от 5‘- к
Трансляция
синтез полипептида
рибосомами, используя
матричную РНК.
Прочтение мРНК
рибосомой идёт
в направлении
от 5‘- к
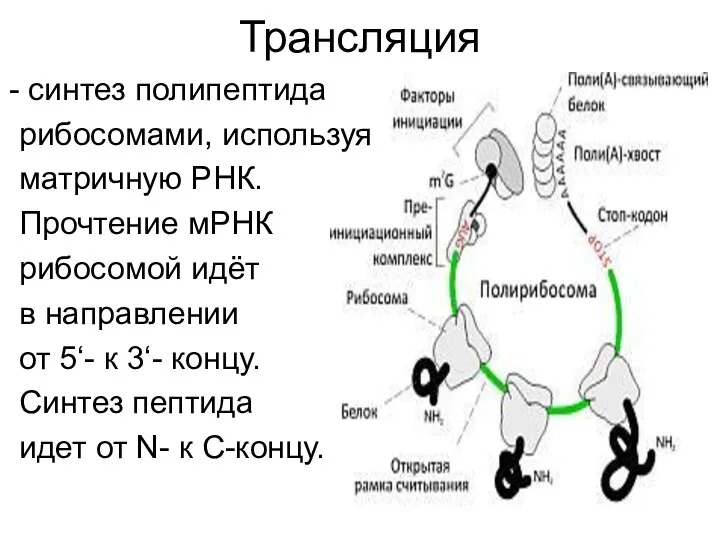
Инициация трансляции
Происходит образование комплекса мРНК, рибосомы и инициирующей тРНК (fMet-, Met-тРНК).
Участвуют eIF (более
Инициация трансляции
Происходит образование комплекса мРНК, рибосомы и инициирующей тРНК (fMet-, Met-тРНК).
Участвуют eIF (более

Элонгация трансляции
В Р-центре – AUG-кодон, соединенный с Met-tRNAi, в А-центре – триплет, кодирующий
Элонгация трансляции
В Р-центре – AUG-кодон, соединенный с Met-tRNAi, в А-центре – триплет, кодирующий

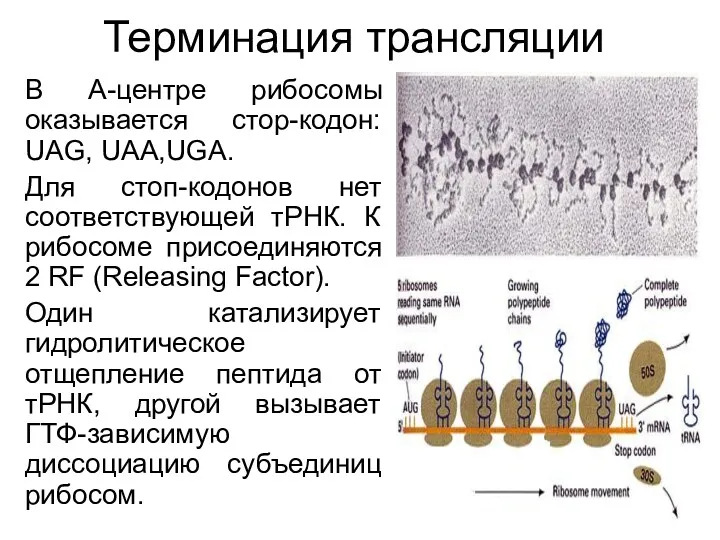
Терминация трансляции
В А-центре рибосомы оказывается стор-кодон: UAG, UAA,UGA.
Для стоп-кодонов нет соответствующей тРНК. К
Терминация трансляции
В А-центре рибосомы оказывается стор-кодон: UAG, UAA,UGA.
Для стоп-кодонов нет соответствующей тРНК. К
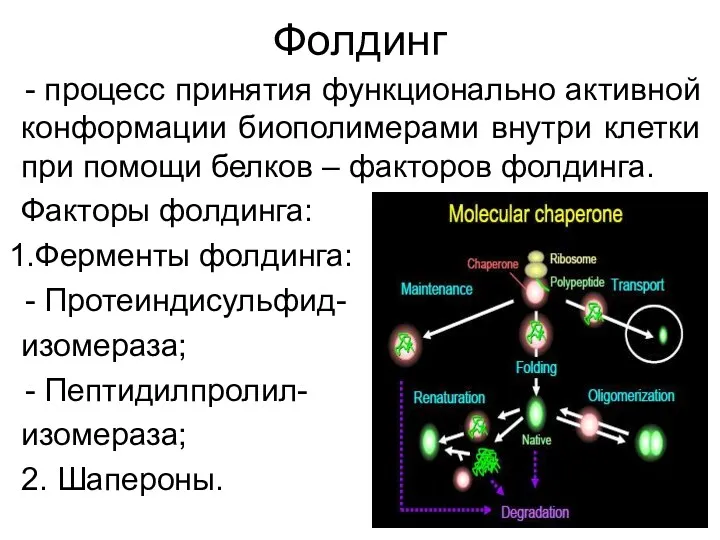
Фолдинг
процесс принятия функционально активной конформации биополимерами внутри клетки при помощи белков –
Фолдинг
процесс принятия функционально активной конформации биополимерами внутри клетки при помощи белков –
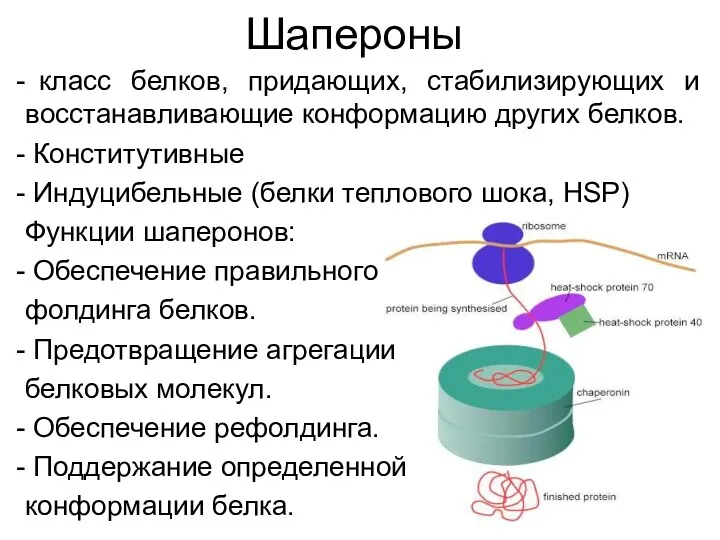
Шапероны
класс белков, придающих, стабилизирующих и восстанавливающие конформацию других белков.
Конститутивные
Индуцибельные (белки
Шапероны
класс белков, придающих, стабилизирующих и восстанавливающие конформацию других белков.
Конститутивные
Индуцибельные (белки
Синтез теломер
На каждом конце хромосомы имеются многократные повторы GGGTTA – теломерная ДНК. После
Синтез теломер
На каждом конце хромосомы имеются многократные повторы GGGTTA – теломерная ДНК. После
 Мышечная система. Кафедра анатомии человека
Мышечная система. Кафедра анатомии человека Гипофиз, гипоталамус гормондары
Гипофиз, гипоталамус гормондары Орган слуха и равновесия
Орган слуха и равновесия Онтогенез
Онтогенез Ќарапайым хордалылар ретінде бас хордалылардыњ µкілі ланцетниктіњ сыртќы жєне ішкі ќ±рылысы
Ќарапайым хордалылар ретінде бас хордалылардыњ µкілі ланцетниктіњ сыртќы жєне ішкі ќ±рылысы Эволюционная теория Жана Батиста Ламарка
Эволюционная теория Жана Батиста Ламарка Белки. Структура белков. Физические и химические свойства
Белки. Структура белков. Физические и химические свойства Органы чувств. Анализаторы
Органы чувств. Анализаторы Жорғалаушылар класы
Жорғалаушылар класы Презентация проекта ЗОЖ.
Презентация проекта ЗОЖ. Растения в русском фольклоре
Растения в русском фольклоре Тип плоские черви. Класс цестоды
Тип плоские черви. Класс цестоды Круговорот углерода в природе
Круговорот углерода в природе Физиология сенсорных систем
Физиология сенсорных систем Міні-проект на тему: цікаві факти про рептилії
Міні-проект на тему: цікаві факти про рептилії Жасуша ядросы
Жасуша ядросы презентация к уроку Голосеменные растения
презентация к уроку Голосеменные растения Жизненный цикл клетки. Митоз
Жизненный цикл клетки. Митоз Бактериологиялық бояулардың түрлерімен танысу
Бактериологиялық бояулардың түрлерімен танысу Организм человека - сложная биосистема
Организм человека - сложная биосистема Испарение и выделение у растений
Испарение и выделение у растений Ямчатость стебля
Ямчатость стебля Гормоны-1
Гормоны-1 Мышечная и нервная ткани. Лекция № 4
Мышечная и нервная ткани. Лекция № 4 Биологически активные добавки
Биологически активные добавки Наука генетика. Краткая история развития классической генетики
Наука генетика. Краткая история развития классической генетики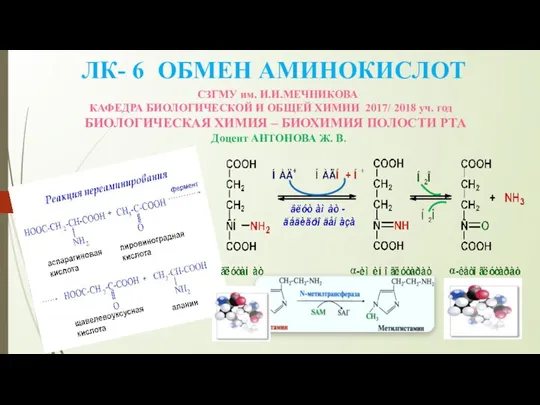 Обмен аминокислот
Обмен аминокислот Клетка. Клеточная теория
Клетка. Клеточная теория