- Функциональная морфология цитоскелета. Функциональная морфология ядра клетки

Содержание
- 2. Цитоскелет Это сложная динамичная (лабильная), трехмерная цитоплазматическая сеть немембранных волокнистых и трубчатых структур различного типа. Элементы
- 4. Компоненты цитоскелета Микротрубочки Промежуточные филаменты Микрофиламенты Микротрабекулы
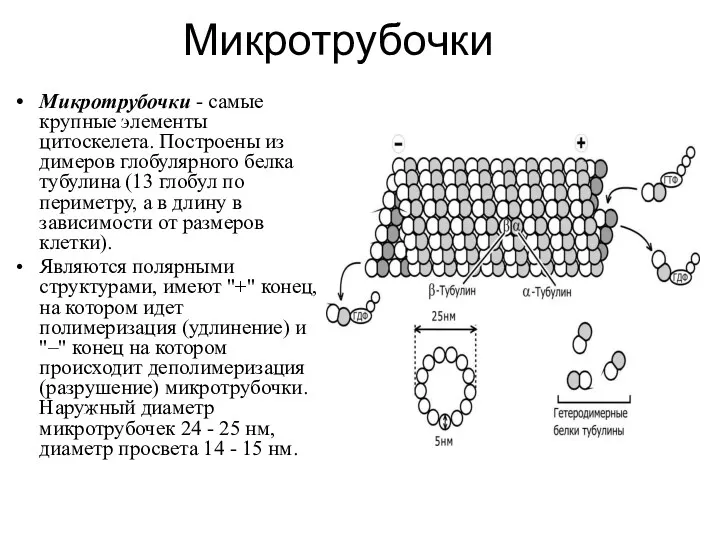
- 6. Микротрубочки Микротрубочки - самые крупные элементы цитоскелета. Построены из димеров глобулярного белка тубулина (13 глобул по
- 7. Лабильность микротрубочек «-» конец связан с белками ЦОМТ. «+» конец свободный, участвует в процессах полимеризации /
- 8. Микротрубочки Работа микротрубочек регулируется специальными белками (динеин, кинезин, МАР-белки, и др.). МАР-белки регулируют процессы полимеризации –
- 9. Динеин и кинезин обеспечивает скольжение микротрубочек относительно друг друга и перемещение мембранных органелл вдоль по поверхности
- 10. Центр организации микротрубочек ЦОМТ, связанный с клеточным центром. ЦОМТ без определенной локализации. Микротрубочки могут образовываться в
- 11. Комплексы микротрубочек В клетке микротрубочки могут формировать комплексы, выполняющие специфические функции. К комплексам, построенным из микротрубочек
- 12. Ресничка На вертикальном разрезе ресничка состоит из корешка, базального тельца (в цитоплазме) и собственно реснички (над
- 13. Ресничка Строение ресничек и жгутиков одинаково - это цилиндрические выросты цитоплазмы, основу которых составляет каркас из
- 14. Неподвижные Все типы клеток (кроме клеток крови, мышечных клеток) в G0-периоде формируют первичные реснички. Рост аксонемы
- 15. Клеточный центр (центросома) в 1888 г.Теодор Бовери - универсальный немембранный органоид всех эукариотических клеток, органелла клеточного
- 16. Клеточный центр (диплосома) Образован двумя центриолями. Одна является материнской, другая – дочерней. Располагаются перпендикулярно друг другу.
- 17. Центросфера Сателлиты/гало Микротрубочки Фокусы схождения микротрубочек Сателлиты – ЦОМТ, состоящие из белковой головки и фибриллярной ножки,
- 18. Центросомный цикл М-фаза: две диплосомы на полюсах клетки, от них отходят нити ахроматинового веретена деления. Материнская
- 19. Базальное тельце (кинетосома) Состоит из 9 триплетов микротрубочек, имеет центральную белковую фибриллу и спицы идущие к
- 20. Функции микротрубочек Поддержание формы клетки Обеспечение внутриклеточного транспорта Формирование органелл Обеспечение подвижности клетки Формирования веретена деления
- 21. Промежуточные филаменты Прочные устойчивые стабильные и самые долгоживущие компоненты цитоскелета. Представляют собой неполярные нити (фибриллы) толщиной
- 22. Строение промежуточных филаментов Димер Тетрамер Протофиламент Волокно
- 23. Классы промежуточных филаментов Тонофиламенты (кератины); Десминовые филаменты (десмин); Виментиновые филаменты (виментин); Нейрофиламенты (NF-L, NF-M, NF-H); Глиальные
- 24. Функции промежуточных филаментов Поддержание формы клетки; Распределение органелл в цитоплазме; Формирование рогового вещества; Формирование остова отростков
- 25. Микрофиламенты Тонкие белковые полярные нити диаметром 5-7 нм, расположенные в цитоплазме поодиночке, пучками или в виде
- 26. Белки микрофиламентов Актин – глобулярный мономерный белок (G-актин), способный к полимеризации (F-актин). Молекула актина имеет вид
- 27. Микро- филаменты
- 28. Актин-связывающие белки Белки, ингибирующие полимеризацию актина (профиллин, ДНКазаI); стабилизирующие белки (тропомиозины); кэпактины (гельзолин/вилин,фрагмин/северин) белки, сшивающие актиновые
- 29. Микроворсинки выросты цитоплазмы, окруженные плазмолеммой, каркас которых образован пучком микрофиламентами. У основания микроворсинки пучок микрофиламентов переходит
- 30. Функции микрофиламентов Обеспечение сокращения клеток Обеспечение движения клеток. Обеспечение мембранных функций. Перемещение в цитозоле органелл, макромолекул.
- 31. Микротрабекулы Наименее изученная часть цитоскелета. Выявляются только высоковольтной (мегавольтной) трансмиссионной электронной микроскопией в виде сети нитей
- 32. Микротрабекулы система тонких белковых нитей, пересекающих цитоплазму в различных направлениях. D 2 нм. С микротрабекулярной системой
- 33. Клеточное ядро - основной компонент эукариотической клетки, содержащий её генетический материал.
- 34. Ядро эукариотической клетки Функция ядра заключается: в хранении и реализации генетической информации (для синтеза белков) и
- 35. Ядро эукариотической клетки Схема строения клеточного ядра. 1 — ядерная оболочка (две мембраны, внутренняя и внешняя,
- 36. Кариолемма Кариолемма состоит из двух биологических мембран (наружной и внутренней) толщиной 6 - 7,5 нм, которые
- 37. Кариолемма Перинуклеарное пространство (люмен) переходит в полости ЭПС. Со стороны цитоплазмы наружная ядерная мембрана окружена сетью
- 38. Внутренняя мембрана кариолеммы ядерная ламина (80-300 нм) - структура, образованная белками-ламинами, к которой прикреплены нити хроматина.
- 39. Функции ламины Поддерживает форму ядра. Участвует в формировании порового комплекса. Отвечает за упорядоченное расположение хроматина в
- 40. Кариолемма В отличие от других органелл, кариолемма содержит поровые комплексы, состоящие из пор и белковых компонентов.
- 41. области перехода внутренней мембраны кариолеммы в наружную. В поре расположен комплекс ядерной поры: Два параллельных белковых
- 42. Комплекс ядерной поры 1— перинуклеарное пространство, 2 — внутренняя ядерная мембрана, 3 — внешняя ядерная мембрана,
- 43. Комплекс ядерной поры Образован - нуклеопоринами
- 44. Комплекс ядерной поры
- 45. Хроматин комплекс ДНК и белков интерфазного ядра, представляющий деспирализованные хромосомы. Гетерохроматин: а. Облигатный; б. Факультативный; Эухроматин;
- 46. Хроматин Хроматин ядра интерфазной клетки - комплекс ДНК и белков. Разделяется на гетерохроматин (конденсированный, заблокированный) и
- 47. Уровни компактизации хроматина
- 48. Негистоновые белки Большое влияние на структуру хроматина и функционирование эукариотических генов оказывают различные негистоновые белки. Белки
- 49. Метафазная хромосома Центромера Теломера Хроматида Плечи (р и q) В области центромеры расположен кинетохор - мультибелковый
- 50. Типы хромосом акроцентрические (палочковидные хромосомы с очень коротким, почти незаметным вторым плечом); субметацентрические (с плечами неравной
- 51. Кариотип Совокупность признаков (числа, величины и морфологии) полного набора хромосом, присущий клеткам данного биологического вида (видовой
- 52. Микроядра - фрагменты хромосом или целые хромосомы, не включенные в состав ядра после митотического деления клетки.
- 53. Ядрышко Ядрышко (от 1 до 5) образованно специализированными участками 13, 14, 15, 21, 22 хромосом (ядрышковыми
- 54. Ядрышко Строение ядрышка: 1 – фибриллярный компонент, 2 – глобулярный компонент, 3 – ядерная оболочка.
- 55. Структура ядрышка Фибриллярный компонент - внутренняя часть ядрышка, состоит из нитей хроматина и первичных транскриптов рРНК.
- 56. Размеры и количество ядрышек В интерфазном ядре на светооптическом уровне – плотная гранула (D 1-3 мкм),
- 57. Ядерный матрикс Кариоскелет построен из негистоновых белков, формирующих разветвленную сеть, взаимодействующую с ядерной ламиной. В клетке
- 58. Кариоплазма Кариоплазма (ядерный сок, кариолимфа) - жидкий компонент ядра, в котором располагаются все внутренние структуры (сходен
- 60. Скачать презентацию

Цитоскелет
Это сложная динамичная (лабильная), трехмерная цитоплазматическая сеть немембранных волокнистых и
Цитоскелет
Это сложная динамичная (лабильная), трехмерная цитоплазматическая сеть немембранных волокнистых и
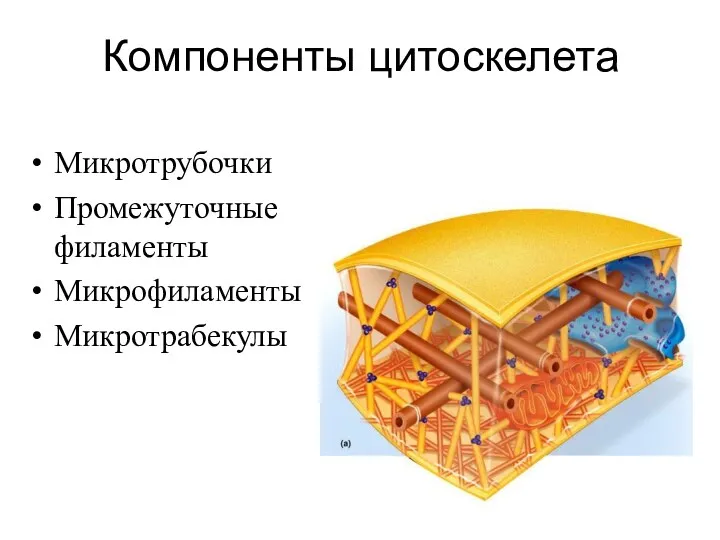
Компоненты цитоскелета
Микротрубочки
Промежуточные филаменты
Микрофиламенты
Микротрабекулы
Компоненты цитоскелета
Микротрубочки
Промежуточные филаменты
Микрофиламенты
Микротрабекулы
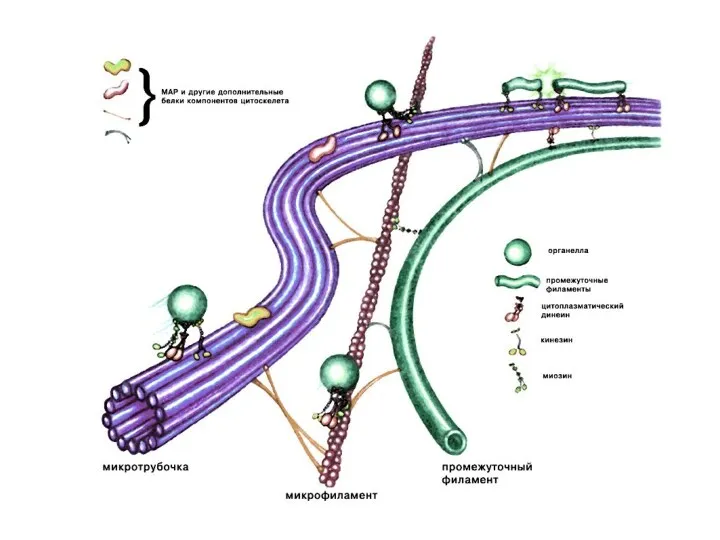
Микротрубочки
Микротрубочки - самые крупные элементы цитоскелета. Построены из димеров глобулярного белка
Микротрубочки
Микротрубочки - самые крупные элементы цитоскелета. Построены из димеров глобулярного белка

Лабильность микротрубочек
«-» конец связан с белками ЦОМТ.
«+» конец свободный, участвует в
Лабильность микротрубочек
«-» конец связан с белками ЦОМТ.
«+» конец свободный, участвует в

Микротрубочки
Работа микротрубочек регулируется специальными белками (динеин, кинезин, МАР-белки, и др.).
МАР-белки
Микротрубочки
Работа микротрубочек регулируется специальными белками (динеин, кинезин, МАР-белки, и др.).
МАР-белки
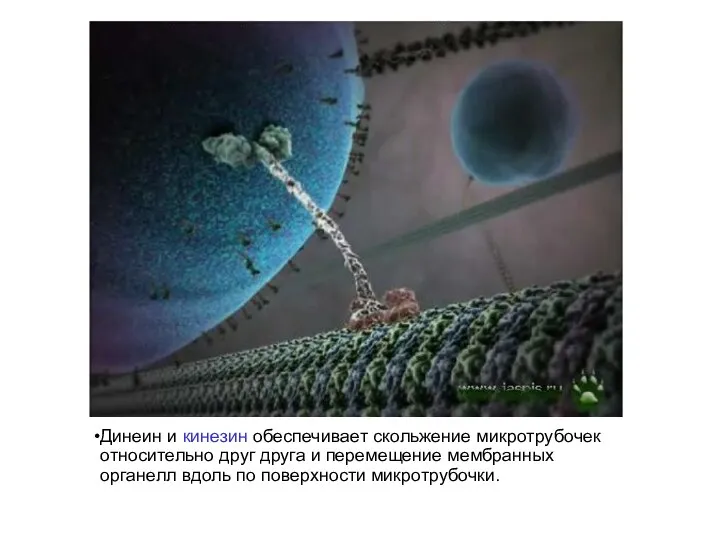
Динеин и кинезин обеспечивает скольжение микротрубочек относительно друг друга и перемещение
Динеин и кинезин обеспечивает скольжение микротрубочек относительно друг друга и перемещение

Центр организации микротрубочек
ЦОМТ, связанный с клеточным центром.
ЦОМТ без определенной
Центр организации микротрубочек
ЦОМТ, связанный с клеточным центром.
ЦОМТ без определенной

Комплексы микротрубочек
В клетке микротрубочки могут формировать комплексы, выполняющие специфические функции.
К
Комплексы микротрубочек
В клетке микротрубочки могут формировать комплексы, выполняющие специфические функции.
К
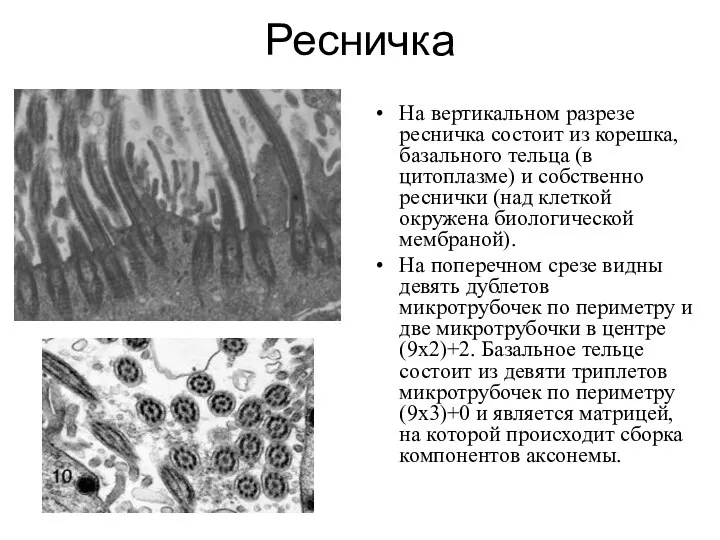
Ресничка
На вертикальном разрезе ресничка состоит из корешка, базального тельца (в цитоплазме)
Ресничка
На вертикальном разрезе ресничка состоит из корешка, базального тельца (в цитоплазме)
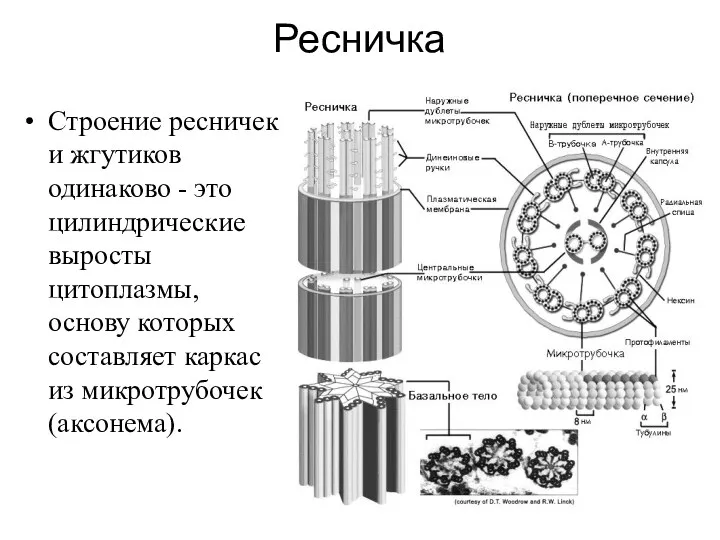
Ресничка
Строение ресничек и жгутиков одинаково - это цилиндрические выросты цитоплазмы, основу
Ресничка
Строение ресничек и жгутиков одинаково - это цилиндрические выросты цитоплазмы, основу
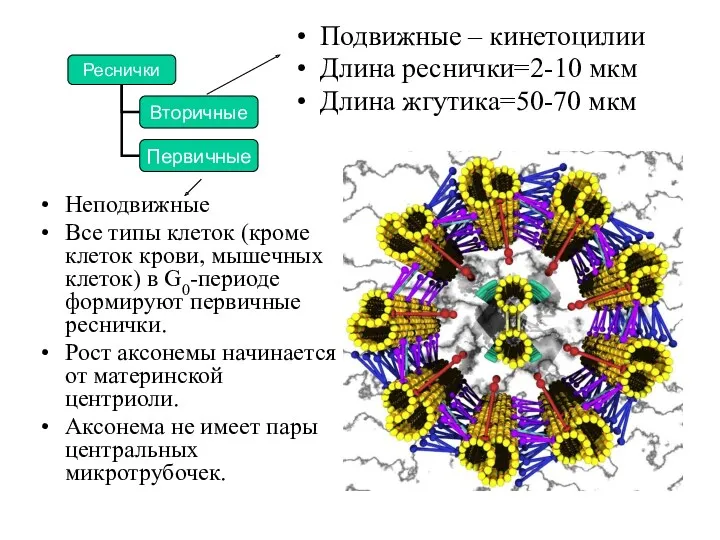
Неподвижные
Все типы клеток (кроме клеток крови, мышечных клеток) в G0-периоде формируют
Неподвижные
Все типы клеток (кроме клеток крови, мышечных клеток) в G0-периоде формируют

Клеточный центр (центросома)
в 1888 г.Теодор Бовери
- универсальный немембранный органоид
Клеточный центр (центросома)
в 1888 г.Теодор Бовери
- универсальный немембранный органоид
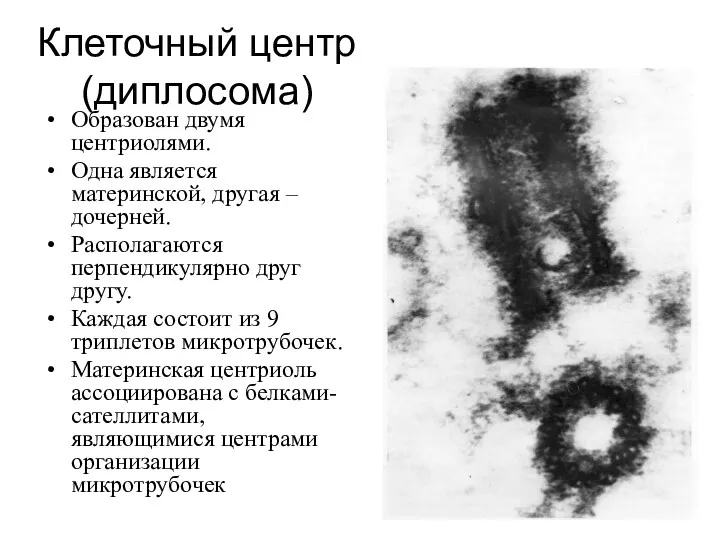
Клеточный центр
(диплосома)
Образован двумя центриолями.
Одна является материнской, другая – дочерней.
Располагаются перпендикулярно друг
Клеточный центр
(диплосома)
Образован двумя центриолями.
Одна является материнской, другая – дочерней.
Располагаются перпендикулярно друг
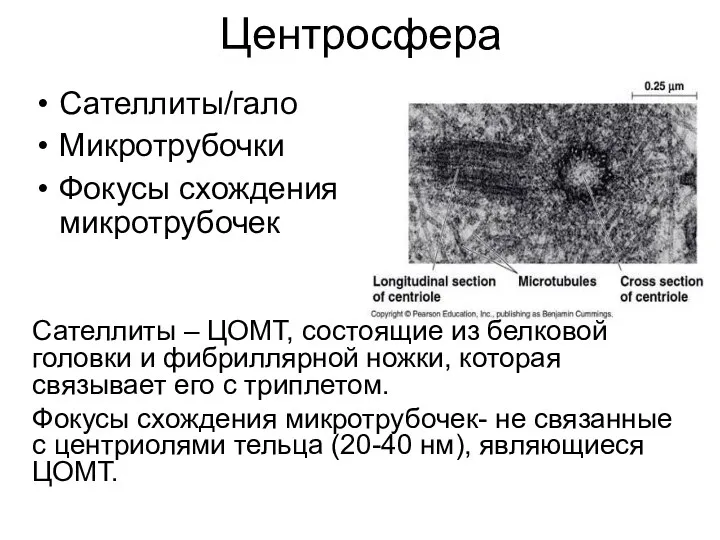
Центросфера
Сателлиты/гало
Микротрубочки
Фокусы схождения микротрубочек
Сателлиты – ЦОМТ, состоящие из белковой головки и фибриллярной
Центросфера
Сателлиты/гало
Микротрубочки
Фокусы схождения микротрубочек
Сателлиты – ЦОМТ, состоящие из белковой головки и фибриллярной

Центросомный цикл
М-фаза: две диплосомы на полюсах клетки, от них отходят нити
Центросомный цикл
М-фаза: две диплосомы на полюсах клетки, от них отходят нити

Базальное тельце (кинетосома)
Состоит из 9 триплетов микротрубочек, имеет центральную
белковую фибриллу
Базальное тельце (кинетосома)
Состоит из 9 триплетов микротрубочек, имеет центральную
белковую фибриллу

Функции микротрубочек
Поддержание формы клетки
Обеспечение внутриклеточного транспорта
Формирование органелл
Обеспечение подвижности клетки
Формирования веретена
Функции микротрубочек
Поддержание формы клетки
Обеспечение внутриклеточного транспорта
Формирование органелл
Обеспечение подвижности клетки
Формирования веретена

Промежуточные филаменты
Прочные устойчивые стабильные и самые долгоживущие компоненты цитоскелета.
Представляют собой
Промежуточные филаменты
Прочные устойчивые стабильные и самые долгоживущие компоненты цитоскелета.
Представляют собой
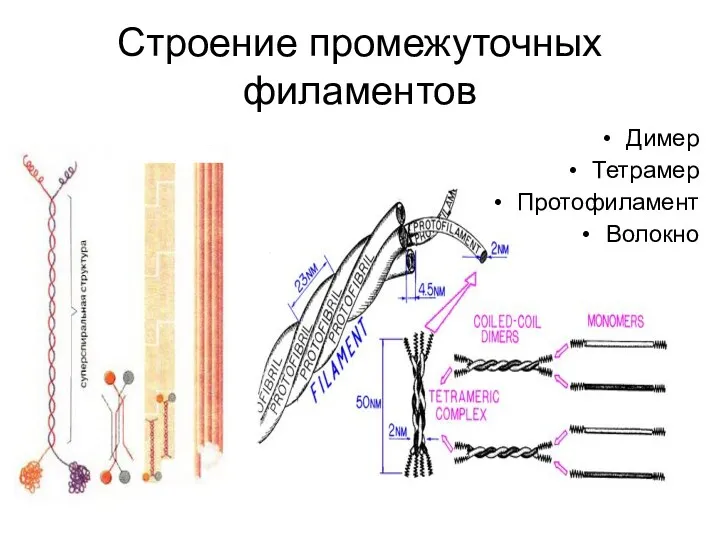
Строение промежуточных филаментов
Димер
Тетрамер
Протофиламент
Волокно
Строение промежуточных филаментов
Димер
Тетрамер
Протофиламент
Волокно

Классы промежуточных филаментов
Тонофиламенты (кератины);
Десминовые филаменты (десмин);
Виментиновые филаменты (виментин);
Нейрофиламенты (NF-L, NF-M, NF-H);
Глиальные
Классы промежуточных филаментов
Тонофиламенты (кератины);
Десминовые филаменты (десмин);
Виментиновые филаменты (виментин);
Нейрофиламенты (NF-L, NF-M, NF-H);
Глиальные

Функции промежуточных филаментов
Поддержание формы клетки;
Распределение органелл в цитоплазме;
Формирование рогового вещества;
Формирование остова
Функции промежуточных филаментов
Поддержание формы клетки;
Распределение органелл в цитоплазме;
Формирование рогового вещества;
Формирование остова

Микрофиламенты
Тонкие белковые полярные нити диаметром 5-7 нм, расположенные в цитоплазме поодиночке,
Микрофиламенты
Тонкие белковые полярные нити диаметром 5-7 нм, расположенные в цитоплазме поодиночке,
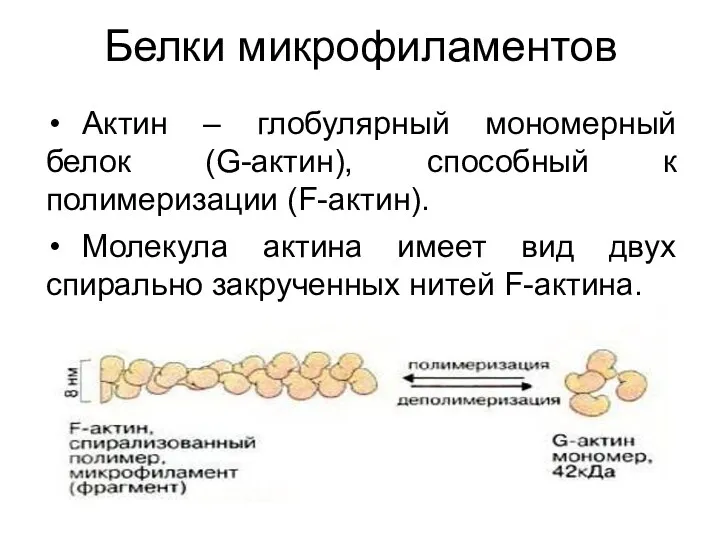
Белки микрофиламентов
Актин – глобулярный мономерный белок (G-актин), способный к полимеризации
Белки микрофиламентов
Актин – глобулярный мономерный белок (G-актин), способный к полимеризации
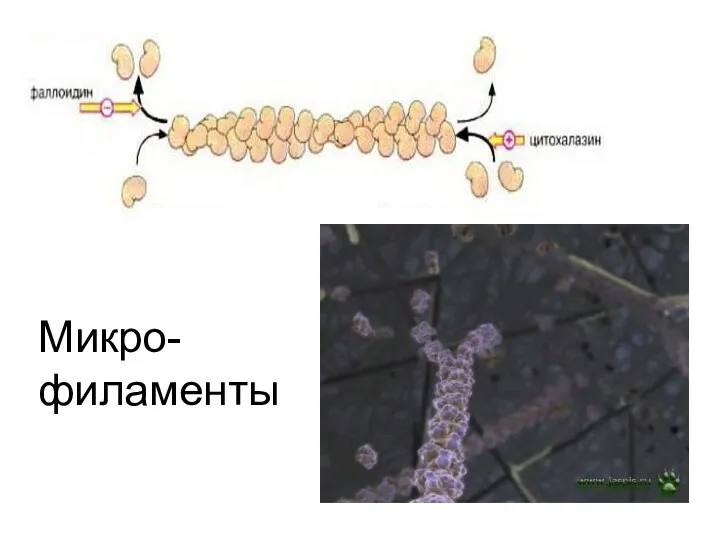
Микро-
филаменты
Микро-
филаменты

Актин-связывающие белки
Белки, ингибирующие полимеризацию актина
(профиллин, ДНКазаI);
стабилизирующие белки (тропомиозины);
Актин-связывающие белки
Белки, ингибирующие полимеризацию актина
(профиллин, ДНКазаI);
стабилизирующие белки (тропомиозины);

Микроворсинки
выросты цитоплазмы, окруженные плазмолеммой, каркас которых образован пучком микрофиламентами.
У основания
Микроворсинки
выросты цитоплазмы, окруженные плазмолеммой, каркас которых образован пучком микрофиламентами.
У основания

Функции микрофиламентов
Обеспечение сокращения клеток
Обеспечение движения клеток.
Обеспечение мембранных функций.
Перемещение в цитозоле органелл,
Функции микрофиламентов
Обеспечение сокращения клеток
Обеспечение движения клеток.
Обеспечение мембранных функций.
Перемещение в цитозоле органелл,

Микротрабекулы
Наименее изученная часть цитоскелета.
Выявляются только высоковольтной (мегавольтной) трансмиссионной электронной микроскопией
Микротрабекулы
Наименее изученная часть цитоскелета.
Выявляются только высоковольтной (мегавольтной) трансмиссионной электронной микроскопией
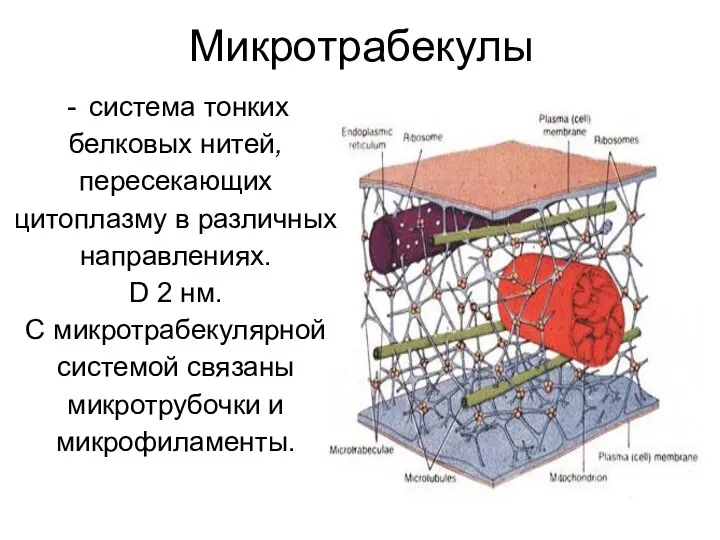
Микротрабекулы
система тонких
белковых нитей,
пересекающих
цитоплазму в различных
направлениях.
D 2
Микротрабекулы
система тонких
белковых нитей,
пересекающих
цитоплазму в различных
направлениях.
D 2
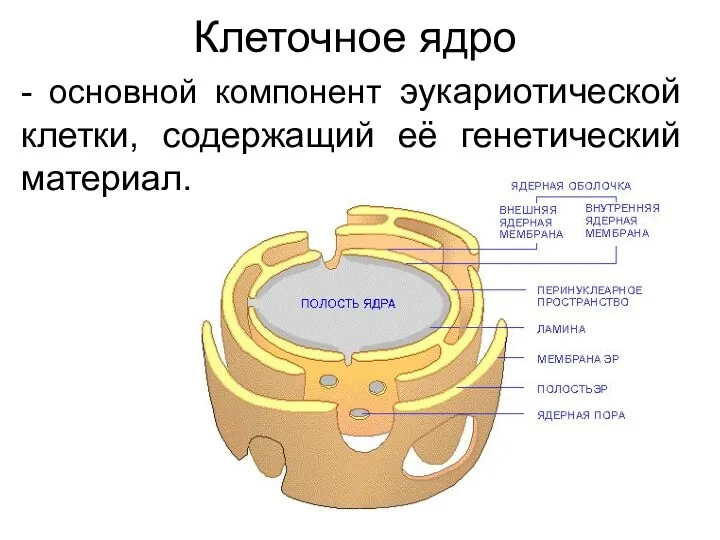
Клеточное ядро
- основной компонент эукариотической клетки, содержащий её генетический материал.
Клеточное ядро
- основной компонент эукариотической клетки, содержащий её генетический материал.

Ядро эукариотической клетки
Функция ядра заключается: в хранении и реализации генетической информации
Ядро эукариотической клетки
Функция ядра заключается: в хранении и реализации генетической информации
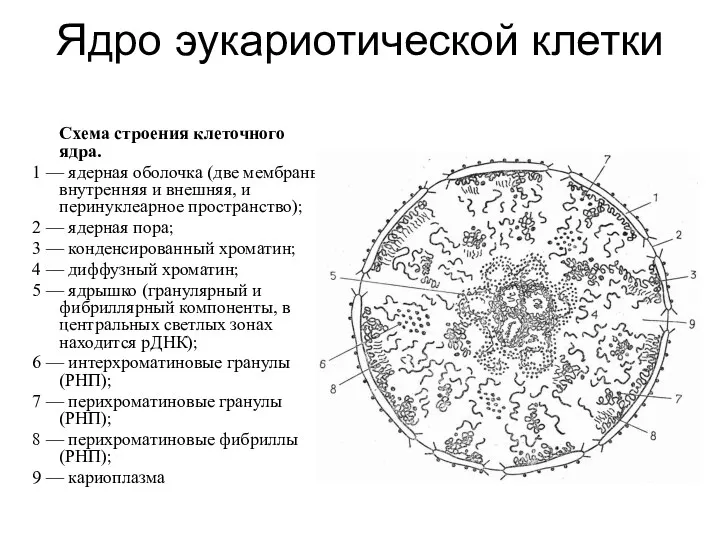
Ядро эукариотической клетки
Схема строения клеточного ядра.
1 — ядерная оболочка
Ядро эукариотической клетки
Схема строения клеточного ядра.
1 — ядерная оболочка
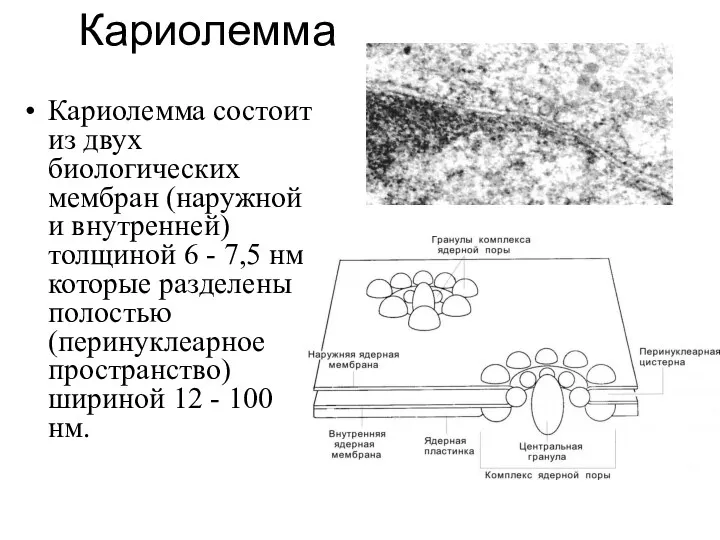
Кариолемма
Кариолемма состоит из двух биологических мембран (наружной и внутренней) толщиной 6
Кариолемма
Кариолемма состоит из двух биологических мембран (наружной и внутренней) толщиной 6
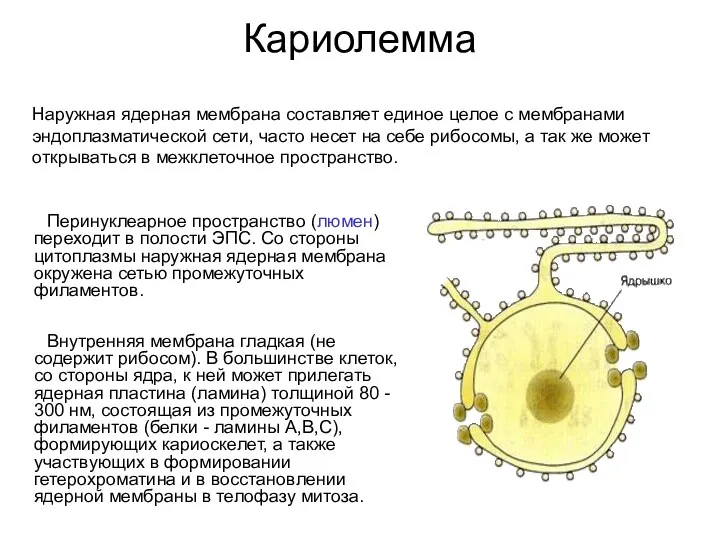
Кариолемма
Перинуклеарное пространство (люмен) переходит в полости ЭПС. Со стороны цитоплазмы
Кариолемма
Перинуклеарное пространство (люмен) переходит в полости ЭПС. Со стороны цитоплазмы
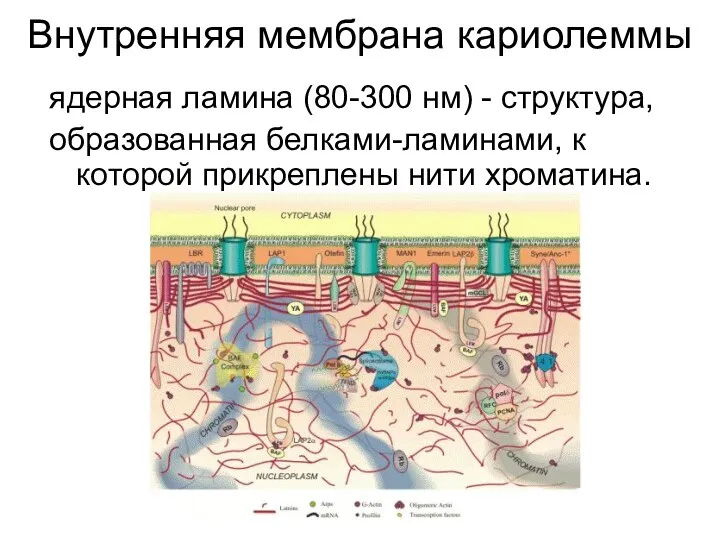
Внутренняя мембрана кариолеммы
ядерная ламина (80-300 нм) - структура,
образованная белками-ламинами, к
Внутренняя мембрана кариолеммы
ядерная ламина (80-300 нм) - структура,
образованная белками-ламинами, к

Функции ламины
Поддерживает форму ядра.
Участвует в формировании порового комплекса.
Отвечает за упорядоченное расположение
Функции ламины
Поддерживает форму ядра.
Участвует в формировании порового комплекса.
Отвечает за упорядоченное расположение

Кариолемма
В отличие от других органелл, кариолемма содержит поровые комплексы, состоящие из
Кариолемма
В отличие от других органелл, кариолемма содержит поровые комплексы, состоящие из

области перехода внутренней мембраны
кариолеммы в наружную.
В поре расположен комплекс ядерной
области перехода внутренней мембраны
кариолеммы в наружную.
В поре расположен комплекс ядерной
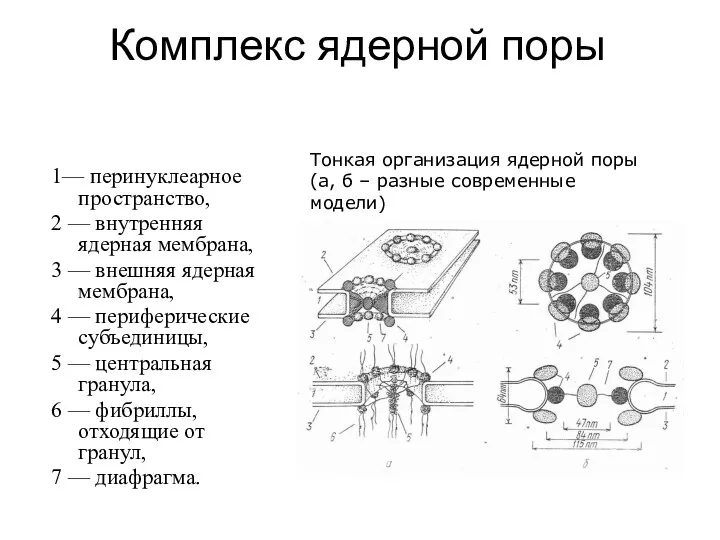
Комплекс ядерной поры
1— перинуклеарное пространство,
2 — внутренняя ядерная мембрана,
3 —
Комплекс ядерной поры
1— перинуклеарное пространство,
2 — внутренняя ядерная мембрана,
3 —
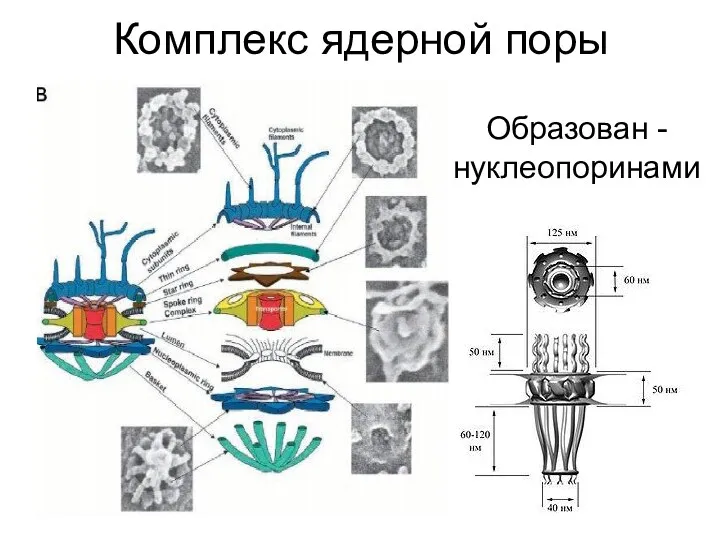
Комплекс ядерной поры
Образован - нуклеопоринами
Комплекс ядерной поры
Образован - нуклеопоринами
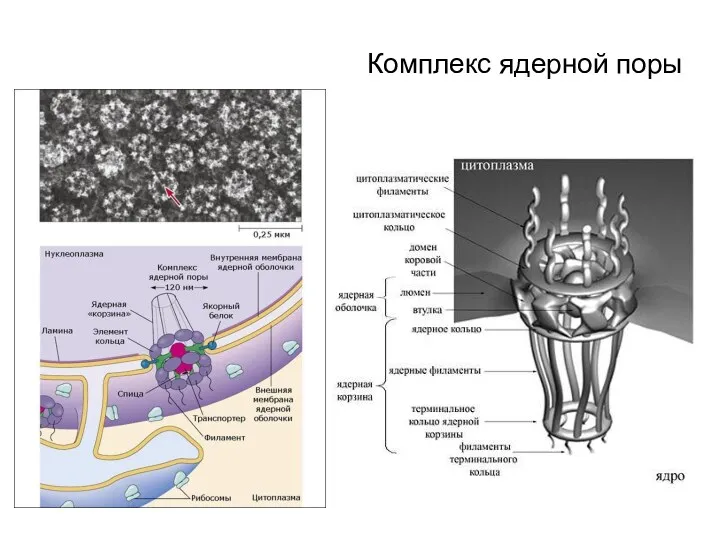
Комплекс ядерной поры
Комплекс ядерной поры
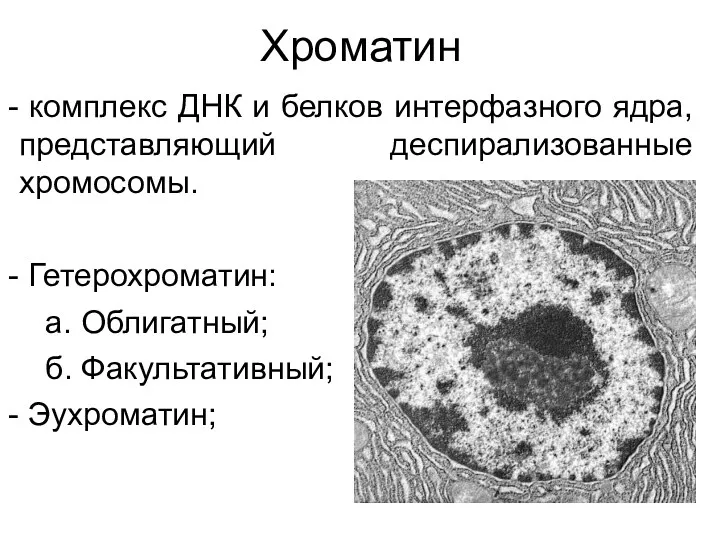
Хроматин
комплекс ДНК и белков интерфазного ядра, представляющий деспирализованные хромосомы.
Гетерохроматин:
Хроматин
комплекс ДНК и белков интерфазного ядра, представляющий деспирализованные хромосомы.
Гетерохроматин:

Хроматин
Хроматин ядра интерфазной клетки - комплекс ДНК и белков.
Разделяется на гетерохроматин
Хроматин
Хроматин ядра интерфазной клетки - комплекс ДНК и белков.
Разделяется на гетерохроматин
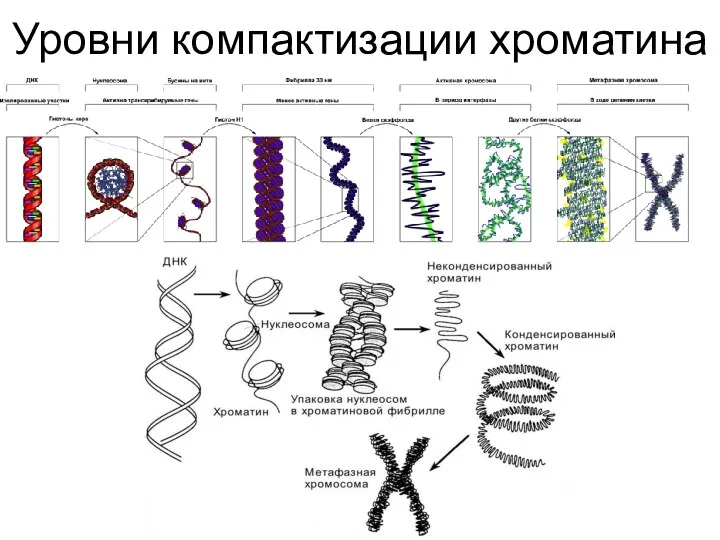
Уровни компактизации хроматина
Уровни компактизации хроматина

Негистоновые белки
Большое влияние на структуру хроматина и функционирование эукариотических генов оказывают
Негистоновые белки
Большое влияние на структуру хроматина и функционирование эукариотических генов оказывают
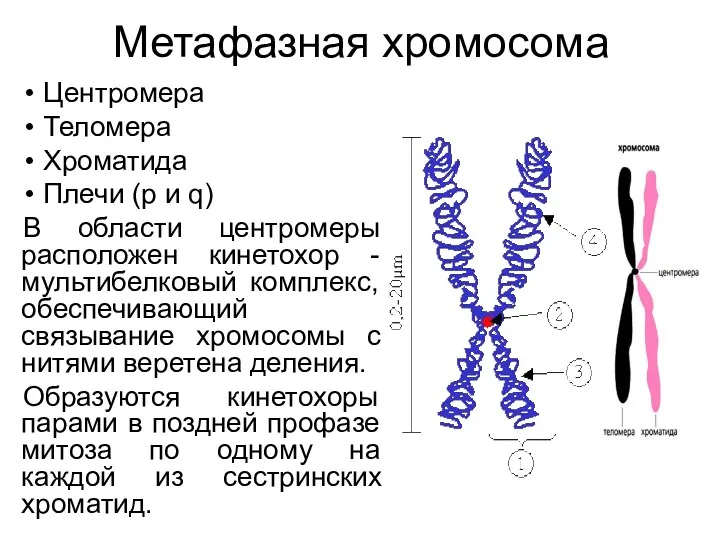
Метафазная хромосома
Центромера
Теломера
Хроматида
Плечи (р и q)
В области центромеры
Метафазная хромосома
Центромера
Теломера
Хроматида
Плечи (р и q)
В области центромеры

Типы хромосом
акроцентрические (палочковидные хромосомы с очень коротким, почти незаметным вторым плечом);
Типы хромосом
акроцентрические (палочковидные хромосомы с очень коротким, почти незаметным вторым плечом);
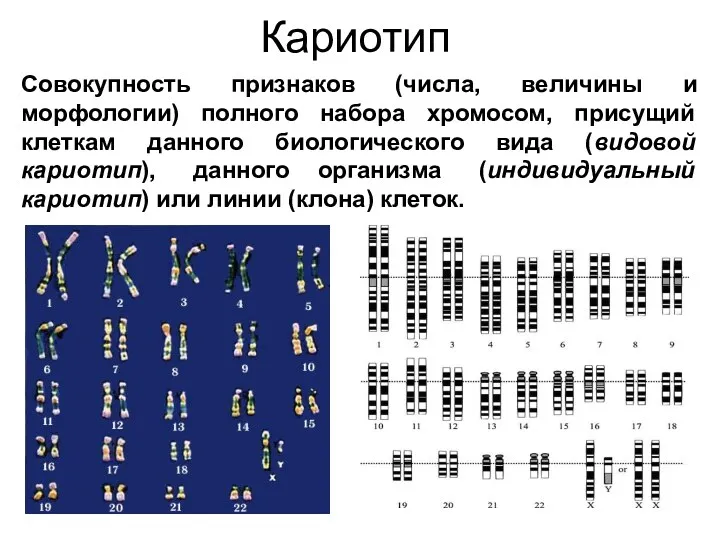
Кариотип
Совокупность признаков (числа, величины и морфологии) полного набора хромосом, присущий клеткам
Кариотип
Совокупность признаков (числа, величины и морфологии) полного набора хромосом, присущий клеткам
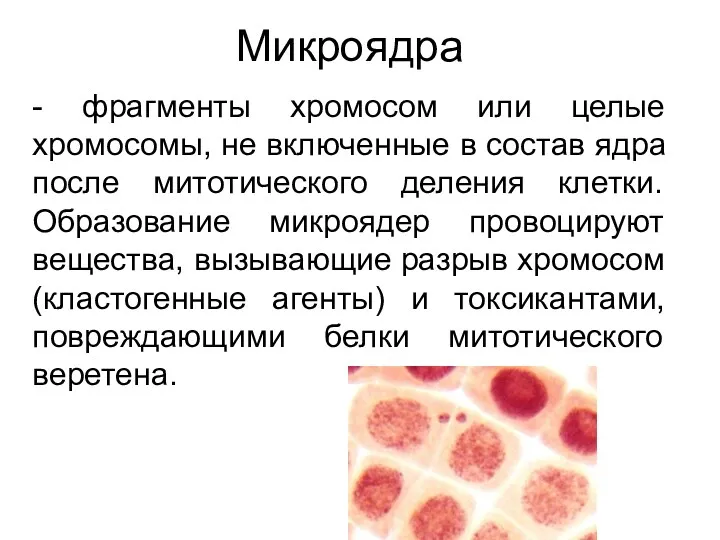
Микроядра
- фрагменты хромосом или целые хромосомы, не включенные в состав ядра
Микроядра
- фрагменты хромосом или целые хромосомы, не включенные в состав ядра
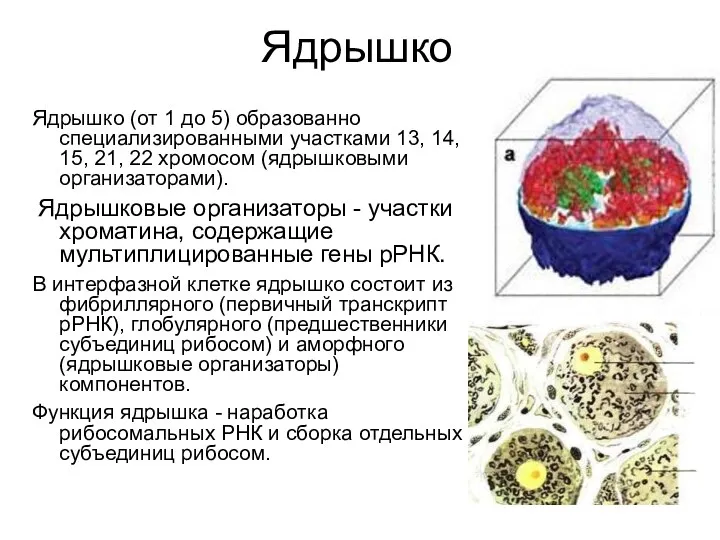
Ядрышко
Ядрышко (от 1 до 5) образованно специализированными участками 13, 14, 15,
Ядрышко
Ядрышко (от 1 до 5) образованно специализированными участками 13, 14, 15,

Ядрышко
Строение ядрышка:
1 – фибриллярный компонент,
2 – глобулярный компонент,
Ядрышко
Строение ядрышка:
1 – фибриллярный компонент,
2 – глобулярный компонент,

Структура ядрышка
Фибриллярный компонент - внутренняя часть ядрышка, состоит из нитей
Структура ядрышка
Фибриллярный компонент - внутренняя часть ядрышка, состоит из нитей

Размеры и количество ядрышек
В интерфазном ядре на светооптическом уровне – плотная
Размеры и количество ядрышек
В интерфазном ядре на светооптическом уровне – плотная

Ядерный матрикс
Кариоскелет построен из негистоновых белков, формирующих разветвленную сеть, взаимодействующую
Ядерный матрикс
Кариоскелет построен из негистоновых белков, формирующих разветвленную сеть, взаимодействующую

Кариоплазма
Кариоплазма (ядерный сок, кариолимфа) - жидкий компонент ядра, в котором располагаются
Кариоплазма
Кариоплазма (ядерный сок, кариолимфа) - жидкий компонент ядра, в котором располагаются
 Анализаторы (органы чувств), их строения и функции, зрительный анализатор
Анализаторы (органы чувств), их строения и функции, зрительный анализатор Среды жизни и места обитания животных. Взаимосвязь животных в природе
Среды жизни и места обитания животных. Взаимосвязь животных в природе Биология – наука о живой природе. Многообразие живых организмов
Биология – наука о живой природе. Многообразие живых организмов Медицинское значение плоских червей
Медицинское значение плоских червей Обмен углеводов
Обмен углеводов Разнообразие животных
Разнообразие животных по биологии Питание и пищеварение
по биологии Питание и пищеварение Видообразование. Микроэволюция
Видообразование. Микроэволюция Растения и животные луга
Растения и животные луга презентация по биологии 7 класс
презентация по биологии 7 класс Биология. Классическая биология. Физико-химическая биология. Эволюционная биология
Биология. Классическая биология. Физико-химическая биология. Эволюционная биология Вегетативное размножение.
Вегетативное размножение. Биохимия соединительной ткани
Биохимия соединительной ткани Вымирающие виды животных в России
Вымирающие виды животных в России Лекарственные растения Воронежской области
Лекарственные растения Воронежской области Презентация к уроку Газообмен в лёгких и тканях
Презентация к уроку Газообмен в лёгких и тканях Чибис - птица 2010 года
Чибис - птица 2010 года Объекты Всемирного наследия
Объекты Всемирного наследия Анатомо-физиологические особенности человека в подростковом возрасте
Анатомо-физиологические особенности человека в подростковом возрасте Організація геному прокаріотів та еукаріотів
Організація геному прокаріотів та еукаріотів Рослини-хижаки
Рослини-хижаки Анатомия и физиология органа зрения (2 часть)
Анатомия и физиология органа зрения (2 часть) Высшие растения. Споровые: мхи, папоротники, хвощи и плауны
Высшие растения. Споровые: мхи, папоротники, хвощи и плауны Генетика – наука про спадковість і мінливість організмів
Генетика – наука про спадковість і мінливість організмів Красота окружающего мира. Насекомые
Красота окружающего мира. Насекомые Анатомия внутренних органов
Анатомия внутренних органов Микробиология зерновых продуктов
Микробиология зерновых продуктов Молекулярные основы наследственности
Молекулярные основы наследственности