- Газообмен. Перенос газов кровью

Содержание
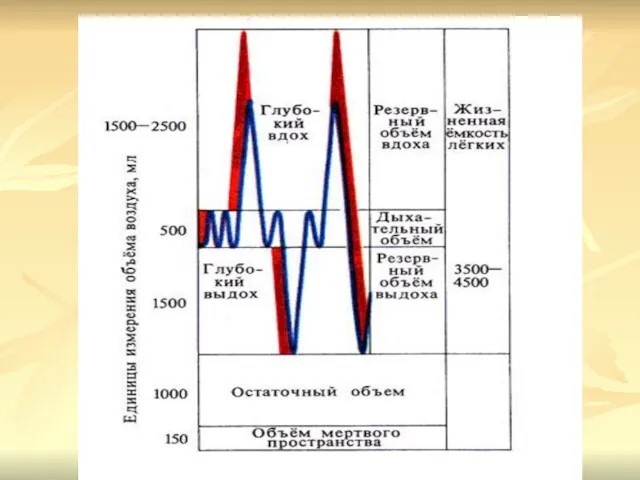
- 2. Объем воздуха, который остается в легких после максимального выдоха и в легких мертвого человека, — остаточный
- 3. Различают следующие емкости легких: 1) общая емкость легких (ОЕЛ) — объем воздуха, находящегося в легких после
- 4. 3) емкость вдоха (Е ) равна сумме дыхательного объема и резервного объема вдоха, составляет в среднем
- 5. Схема вентиляции кровотока и газообмена верхнего и нижнего горизонтальных слоев нормального легкого (по Дж. Весту, 1970)
- 6. Исследование легочных объемов и емкостей как важнейших показателей функционального состояния легких имеет большое медико-физиологическое значение не
- 7. Для сопоставимости результатов измерений газовых объемов и емкостей материалы исследований должны быть приведены к стандартному состоянию
- 8. Воздух, находящийся в воздухоносных путях (полость рта, носа, глотки, трахеи, бронхов и бронхиол), не участвует в
- 10. ГАЗООБМЕН И ТРАНСПОРТ ГАЗОВ Количество кислорода, поступающего в альвеолярное пространство из вдыхаемого воздуха в единицу времени
- 11. Эта основная закономерность легочного газообмена характерна и для углекислого газа: количество этого газа, поступающего в альвеолы
- 12. В тканях всего тела, где происходит внутреннее дыхание, кислород переходит из капилляров в клетки, а углекислота
- 13. У человека в покое разность между содержанием кислорода в артериальной и смешанной венозной крови равна 45-55
- 14. Кровообращение в области альвеолы. А — возможные соотношения между капиллярами и альвеолами; Б — газообмен между
- 15. ОБМЕН ГАЗОВ МЕЖДУ АЛЬВЕОЛЯРНЫМ ВОЗДУХОМ И КРОВЬЮ Альвеолярный воздух и кровь легочных капилляров разделяет так называемая
- 16. БАРЬЕР МЕЖДУ КРОВЬЮ И АЛЬВЕОЛЯРНЫМ ВОЗДУХОМ
- 17. Обмен респираторных газов осуществляется через совокупность субмикроскопических структур, содержащих гемоглобин эритроцитов, плазму крови, капиллярный эндотелий и
- 18. Последний имеет толщину около 50 нм, представляет собой комплекс фосфолипидов, белков и полисахаридов и постоянно вырабатывается
- 19. Переход газов через альвеолокапиллярную мембрану происходит по законам диффузии. Но при растворении газов в жидкости процесс
- 20. Количество газа, проходящее через легочную мембрану в единицу времени, т.е. скорость диффузии, прямо пропорциональна разнице его
- 21. Направление и интенсивность перехода кислорода из альвеолярного воздуха в кровь легочных микрососудов, а углекислого газа -
- 22. Сопротивление диффузии кислорода в легких создают альвеолярно-капиллярная мембрана, слой плазмы в капиллярах, мембрана эритроцита и слой
- 23. Величина диффузионной способности легких зависит от их объема и соответствующей ему площади поверхности газообмена. Этим в
- 24. ТРАНСПОРТ КИСЛОРОДА КРОВЬЮ Кислород в крови находится в растворенном виде и в соединении с гемоглобином. В
- 25. После того как кислород входит в кровь легочных капилляров, он диффундирует из плазмы в эритроциты и
- 26. В 100 мл артериальной крови в норме содержится 19-20 мл кислорода, в таком же объеме венозной
- 27. В каждом литре крови максимально возможное содержание кислорода в химически связанной форме составит 190 - 200
- 28. Зависимость связывания кислорода кровью от его парциального давления можно представить в виде графика, где по оси
- 29. КРИВЫЕ ДИССОЦИАЦИИ ОКСИГЕМОГЛОБИНА: I — КРИВАЯ НАСЫЩЕНИЯ ГЕМОГЛОБИНА КИСЛОРОДОМ ПРИ НОРМАЛЬНОМ СОДЕРЖАНИИ СО2, II — ВЛИЯНИЕ
- 30. Сродство кислорода к гемоглобину и способность отдавать 02 в тканях зависит от метаболических потребностей клеток организма
- 31. К этим факторам относятся: концентрация водородных ионов, температура, парциальное напряжение углекислоты и соединение, которое накапливается в
- 32. Их влияние на кривую диссоциации оксигемоглобина называется эффектом Бора (1904). При повышении концентрации водородных ионов и
- 33. На кривую диссоциации оксигемоглобина оказывает влияние также и температура. Рост температуры значительно увеличивает скорость распада оксигемоглобина
- 34. ПЕРЕНОС ГАЗОВ КРОВЬЮ И ГАЗООБМЕН В ЛЕГКИХ И ТКАНЯХ
- 35. ТРАНСПОРТ УГЛЕКИСЛОГО ГАЗА КРОВЬЮ Диффузия СО2 из крови в альвеолы обеспечивается за счет поступления растворенного в
- 36. Углекислый газ в крови находится в трех фракциях: физически растворенный, химически связанный в виде бикарбонатов химически
- 37. От парциального напряжения физически растворенного углекислого газа зависит процесс связывания СО2 кровью. Углекислота поступает в эритроцит,
- 38. Эритроциты переносят в 3 раза больше СО2, чем плазма. Белки плазмы составляют 8 г на 100
- 39. Кроме физически растворенного в плазме крови молекулярного СО2 из крови в альвеолы легких диффундирует СО2 который
- 40. В плазме находится бикарбонат натрия, а в эритроцитах — бикарбонат калия. Мембрана эритроцитов хорошо проницаема для
- 41. Различные яды, ограничивающие транспорт 02, такие как СО, нитриты, ферроцианиды и многие другие, практически не действуют
- 42. Задержка СО2 приводит к дыхательному ацидозу — уменьшению концентрации бикарбонатов, сдвигу рН крови в кислую сторону.
- 43. ОБМЕН ГАЗОВ В ТКАНЯХ ГАЗООБМЕН В ТКАНЯХ ЗАВИСИТ ОТ ПЯТИ ОСНОВНЫХ ФАКТОРОВ: Площади диффузии Градиента напряжения
- 44. Доставка кислорода к тканям происходит с помощью кровотока путем конвекции. Кровоток и АВР-О2 (артериовенозная разница) зависят
- 45. Это значение – критическое напряжение кислорода в митохондриях, ниже которого окисление становится невозможным. Обычно около митохондрий
- 46. В противоположном направлении, начиная от митохондрий, поступает образующийся при окислении СО2. Обычно в тканях уровень РСО2
- 48. Скачать презентацию

Объем воздуха, который остается в легких после максимального выдоха и в
Объем воздуха, который остается в легких после максимального выдоха и в

Различают следующие емкости легких:
1) общая емкость легких (ОЕЛ) — объем
Различают следующие емкости легких:
1) общая емкость легких (ОЕЛ) — объем

3) емкость вдоха (Е ) равна сумме дыхательного объема и резервного
3) емкость вдоха (Е ) равна сумме дыхательного объема и резервного
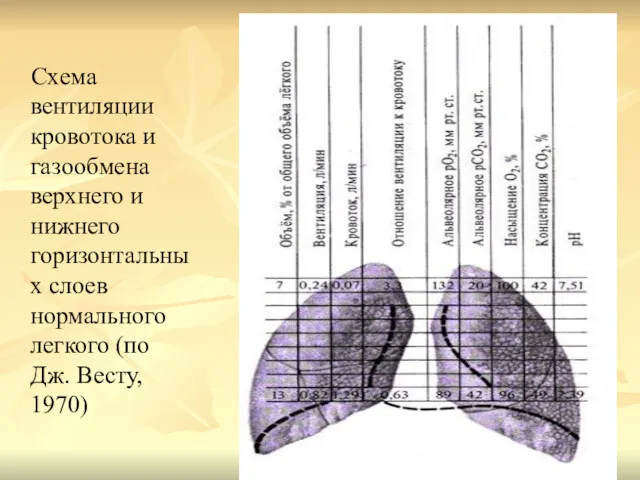
Схема вентиляции кровотока и газообмена верхнего и нижнего горизонтальных
Схема вентиляции кровотока и газообмена верхнего и нижнего горизонтальных

Исследование легочных объемов и емкостей как важнейших показателей функционального состояния
Исследование легочных объемов и емкостей как важнейших показателей функционального состояния

Для сопоставимости результатов измерений газовых объемов и емкостей материалы исследований
Для сопоставимости результатов измерений газовых объемов и емкостей материалы исследований

Воздух, находящийся в воздухоносных путях (полость рта, носа, глотки, трахеи,
Воздух, находящийся в воздухоносных путях (полость рта, носа, глотки, трахеи,

ГАЗООБМЕН И ТРАНСПОРТ ГАЗОВ
Количество кислорода, поступающего в альвеолярное пространство
ГАЗООБМЕН И ТРАНСПОРТ ГАЗОВ
Количество кислорода, поступающего в альвеолярное пространство

Эта основная закономерность легочного газообмена характерна и для углекислого газа:
Эта основная закономерность легочного газообмена характерна и для углекислого газа:

В тканях всего тела, где происходит внутреннее дыхание, кислород переходит
В тканях всего тела, где происходит внутреннее дыхание, кислород переходит

У человека в покое разность между содержанием кислорода в артериальной
У человека в покое разность между содержанием кислорода в артериальной
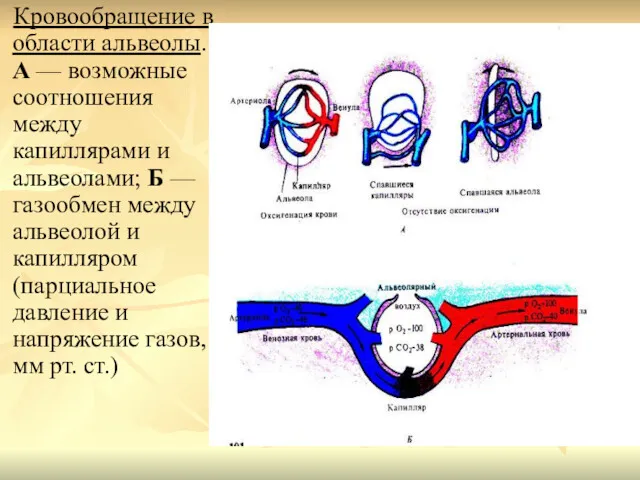
Кровообращение в области альвеолы. А — возможные соотношения между капиллярами
Кровообращение в области альвеолы. А — возможные соотношения между капиллярами

ОБМЕН ГАЗОВ МЕЖДУ АЛЬВЕОЛЯРНЫМ ВОЗДУХОМ И
КРОВЬЮ
Альвеолярный воздух и кровь легочных
ОБМЕН ГАЗОВ МЕЖДУ АЛЬВЕОЛЯРНЫМ ВОЗДУХОМ И
КРОВЬЮ
Альвеолярный воздух и кровь легочных
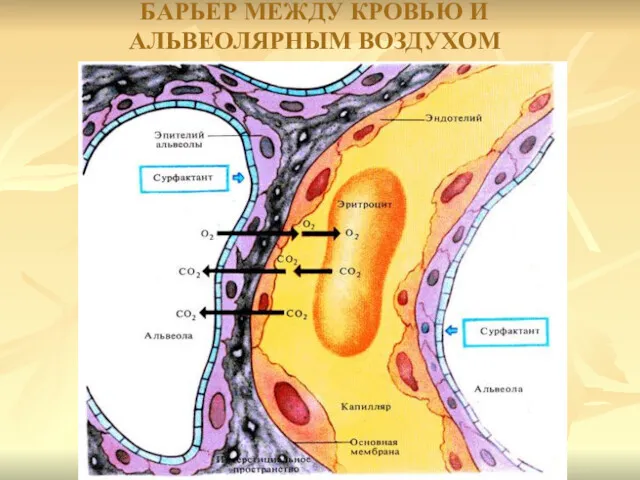
БАРЬЕР МЕЖДУ КРОВЬЮ И АЛЬВЕОЛЯРНЫМ ВОЗДУХОМ
БАРЬЕР МЕЖДУ КРОВЬЮ И АЛЬВЕОЛЯРНЫМ ВОЗДУХОМ

Обмен респираторных газов осуществляется через совокупность субмикроскопических структур, содержащих гемоглобин
Обмен респираторных газов осуществляется через совокупность субмикроскопических структур, содержащих гемоглобин

Последний имеет толщину около 50 нм,
представляет собой комплекс фосфолипидов,
Последний имеет толщину около 50 нм,
представляет собой комплекс фосфолипидов,

Переход газов через альвеолокапиллярную мембрану происходит по законам диффузии. Но
Переход газов через альвеолокапиллярную мембрану происходит по законам диффузии. Но

Количество газа, проходящее через легочную мембрану в единицу времени, т.е.
Количество газа, проходящее через легочную мембрану в единицу времени, т.е.

Направление и интенсивность перехода кислорода из альвеолярного воздуха в кровь
Направление и интенсивность перехода кислорода из альвеолярного воздуха в кровь

Сопротивление диффузии кислорода в легких создают альвеолярно-капиллярная мембрана, слой плазмы
Сопротивление диффузии кислорода в легких создают альвеолярно-капиллярная мембрана, слой плазмы

Величина диффузионной способности легких зависит от их объема и соответствующей
Величина диффузионной способности легких зависит от их объема и соответствующей

ТРАНСПОРТ КИСЛОРОДА КРОВЬЮ
Кислород в крови находится в растворенном виде и
ТРАНСПОРТ КИСЛОРОДА КРОВЬЮ
Кислород в крови находится в растворенном виде и

После того как кислород входит в кровь легочных капилляров, он диффундирует
После того как кислород входит в кровь легочных капилляров, он диффундирует

В 100 мл артериальной крови в норме содержится 19-20 мл
В 100 мл артериальной крови в норме содержится 19-20 мл

В каждом литре крови максимально возможное содержание кислорода в химически
В каждом литре крови максимально возможное содержание кислорода в химически

Зависимость связывания кислорода кровью от его парциального давления можно представить
Зависимость связывания кислорода кровью от его парциального давления можно представить
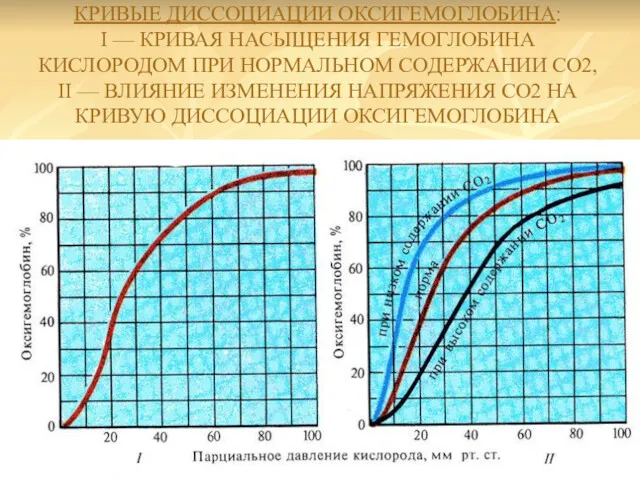
КРИВЫЕ ДИССОЦИАЦИИ ОКСИГЕМОГЛОБИНА:
I — КРИВАЯ НАСЫЩЕНИЯ ГЕМОГЛОБИНА КИСЛОРОДОМ ПРИ НОРМАЛЬНОМ СОДЕРЖАНИИ
КРИВЫЕ ДИССОЦИАЦИИ ОКСИГЕМОГЛОБИНА: I — КРИВАЯ НАСЫЩЕНИЯ ГЕМОГЛОБИНА КИСЛОРОДОМ ПРИ НОРМАЛЬНОМ СОДЕРЖАНИИ

Сродство кислорода к гемоглобину и способность отдавать 02 в тканях
Сродство кислорода к гемоглобину и способность отдавать 02 в тканях

К этим факторам относятся: концентрация водородных ионов, температура, парциальное напряжение
К этим факторам относятся: концентрация водородных ионов, температура, парциальное напряжение

Их влияние на кривую диссоциации оксигемоглобина называется эффектом Бора (1904).
Их влияние на кривую диссоциации оксигемоглобина называется эффектом Бора (1904).

На кривую диссоциации оксигемоглобина оказывает влияние также и температура. Рост
На кривую диссоциации оксигемоглобина оказывает влияние также и температура. Рост
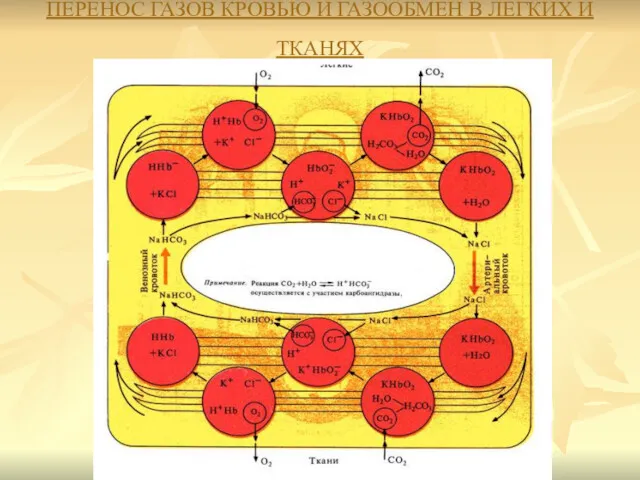
ПЕРЕНОС ГАЗОВ КРОВЬЮ И ГАЗООБМЕН В ЛЕГКИХ И ТКАНЯХ
ПЕРЕНОС ГАЗОВ КРОВЬЮ И ГАЗООБМЕН В ЛЕГКИХ И ТКАНЯХ

ТРАНСПОРТ УГЛЕКИСЛОГО ГАЗА КРОВЬЮ
Диффузия СО2 из крови в альвеолы обеспечивается
ТРАНСПОРТ УГЛЕКИСЛОГО ГАЗА КРОВЬЮ
Диффузия СО2 из крови в альвеолы обеспечивается

Углекислый газ в крови находится в трех фракциях:
физически растворенный,
Углекислый газ в крови находится в трех фракциях:
физически растворенный,

От парциального напряжения физически растворенного углекислого газа зависит процесс связывания
От парциального напряжения физически растворенного углекислого газа зависит процесс связывания

Эритроциты переносят в 3 раза больше СО2, чем плазма. Белки
Эритроциты переносят в 3 раза больше СО2, чем плазма. Белки

Кроме физически растворенного в плазме крови молекулярного СО2 из крови
Кроме физически растворенного в плазме крови молекулярного СО2 из крови

В плазме находится бикарбонат натрия, а в эритроцитах — бикарбонат
В плазме находится бикарбонат натрия, а в эритроцитах — бикарбонат

Различные яды, ограничивающие транспорт 02, такие как СО, нитриты, ферроцианиды
Различные яды, ограничивающие транспорт 02, такие как СО, нитриты, ферроцианиды

Задержка СО2 приводит к дыхательному ацидозу — уменьшению концентрации бикарбонатов,
Задержка СО2 приводит к дыхательному ацидозу — уменьшению концентрации бикарбонатов,

ОБМЕН ГАЗОВ В ТКАНЯХ
ГАЗООБМЕН В ТКАНЯХ ЗАВИСИТ ОТ ПЯТИ ОСНОВНЫХ
ОБМЕН ГАЗОВ В ТКАНЯХ
ГАЗООБМЕН В ТКАНЯХ ЗАВИСИТ ОТ ПЯТИ ОСНОВНЫХ

Доставка кислорода к тканям происходит с помощью кровотока путем конвекции.
Доставка кислорода к тканям происходит с помощью кровотока путем конвекции.

Это значение – критическое напряжение кислорода в митохондриях, ниже которого
Это значение – критическое напряжение кислорода в митохондриях, ниже которого

В противоположном направлении, начиная от митохондрий, поступает образующийся при окислении
В противоположном направлении, начиная от митохондрий, поступает образующийся при окислении
 Стресс у растений. Особенности проявления стрессовых реакций и механизмы устойчивости к стрессовым воздействиям. (Лекция 1-2)
Стресс у растений. Особенности проявления стрессовых реакций и механизмы устойчивости к стрессовым воздействиям. (Лекция 1-2) Жиры, Белки и Углеводы человека
Жиры, Белки и Углеводы человека Ядовитые растения России
Ядовитые растения России Взаимодействие генов
Взаимодействие генов Видоизменения побегов
Видоизменения побегов Семенные растения. Характеристика отделов голосеменные и покрытосеменные
Семенные растения. Характеристика отделов голосеменные и покрытосеменные Строение ядра. Хромосомы
Строение ядра. Хромосомы Презентация Фотосинтез. Необыкновенные приключения хлорофилла
Презентация Фотосинтез. Необыкновенные приключения хлорофилла Животные леса
Животные леса Организм – открытая живая система. 9 класс
Организм – открытая живая система. 9 класс Нейрон и нейроглия. Их виды
Нейрон и нейроглия. Их виды Развитие с полным и неполным превращением. 6 класс
Развитие с полным и неполным превращением. 6 класс Зрительный анализатор. Анатомия и физиология глаза
Зрительный анализатор. Анатомия и физиология глаза Исчезающие растения и животные России
Исчезающие растения и животные России Биосфера и организм
Биосфера и организм Чудо-доктора. Лекарственные растения Лутугинского района
Чудо-доктора. Лекарственные растения Лутугинского района Бесполое размножение (агамогенез)
Бесполое размножение (агамогенез) Химический состав клетки
Химический состав клетки Отряд: Зайцеобразные
Отряд: Зайцеобразные Осенние явления в жизни растений
Осенние явления в жизни растений Строение стебля
Строение стебля Изменчивость и генетика человека (лекция 8,9)
Изменчивость и генетика человека (лекция 8,9) Механические ткани в органах растения
Механические ткани в органах растения Листок і фотосинтез
Листок і фотосинтез Дыхание растений
Дыхание растений Метаболизм. Строение и функции митохондрий
Метаболизм. Строение и функции митохондрий Бурый медведь
Бурый медведь Вроджені системні захворювання скелету
Вроджені системні захворювання скелету