- Генерация ПД в нейронах

Содержание
- 2. Первые записи ионных токов, протекающие через отдельные каналы
- 3. Первые записи ионных токов, протекающие через отдельные каналы
- 4. Ионные токи, протекающие через отдельные каналы Возможность исследования локальных ионных токов, протекающих через отдельные каналы мембран
- 5. Метод локальной фиксации участка мембраны (англ., patch clamp)
- 6. Ионные токи, протекающие через отдельные каналы Открытие и закрытие ионных каналов отражаются в виде прямоугольных токовых
- 7. Ионные токи, протекающие через отдельные каналы Для сравнения приведен Na+-ток в гигантском аксоне Открытие и закрытие
- 8. Ионные токи, протекающие через отдельные каналы Для сравнения приведены Na+- и К+-токи в гигантском аксоне Na+-
- 9. Ионные токи, протекающие через отдельные каналы Для сравнения приведены Na+- и К+-токи в гигантском аксоне Na+-
- 10. Ионные токи, протекающие через отдельные каналы Для сравнения приведены Na+- и К+-токи в гигантском аксоне Na+-
- 11. Ионные токи, протекающие через отдельные Na+-каналы
- 12. Ионные токи, протекающие через отдельные K+-каналы
- 13. Ионные токи, протекающие через отдельные каналы Для сравнения приведены Na+- и К+-токи в гигантском аксоне (!!!
- 15. Ионные токи, протекающие через отдельные каналы С использованием метода локальной фиксации в мембранах нейронов были открыты
- 16. Ионные токи, протекающие через отдельные каналы мышечного волокна предсердия The atrial potential and time course of
- 17. Schematic representation of a human ventricular action potential (top panel). Numbers denote the different phases of
- 18. Функциональная роль потенциал-зависимых токов Потенциал-зависимые каналы (и пропускаемые ими токи) разделяют по различным свойствам (критериям): 1)
- 21. Инактивация Na+-каналов может изменяться в зависимости от их строения Смещение кривой инактивации в сторону гиперполяризации вызвано
- 22. Инактивация Na+-каналов
- 23. Натриевые токи INa,leak NALCH (NALeakCHannels) (ток утечки) является потенциал-независимым током, протекает по неинактивирующимся каналам и обеспечивает
- 25. Кальциевые токи Са2+-токи вносят несущественный вклад в передний фронт ПД из-за их медленной кинетики активации. Но
- 26. Кальциевые токи Большинство Ca2+-каналов активируется при высоком уровне деполяризации мембраны от –20 до –10 мВ. Эти
- 27. Высокопороговые кальциевые токи Высокопороговые (порог от –20 до –10 мВ) Ca2+-токи добавляют порцию деполяризации при генерации
- 28. ICa,L, L-type Ca2+ current
- 29. Высокопороговые кальциевые токи IP/Q (Purkinje) впервые зарегистрирован в клетках Пуркинье. Проводится по двум разновидностям (P и
- 32. Высокопороговые кальциевые токи IR (“Residual”). Изначально канал, проводящий этот ток, был отнесен к промежуточному типу, который
- 33. Инактивация высокопороговых Са2+-токов Инактивация Са2+-каналов Инактивация Na+-каналов
- 34. Низкопороговые кальциевые токи IT (transient, фазический) Единственный низкопороговый Са2+-ток , который обеспечивает ритмический разряд (пачки) ПД.
- 35. ICa,T, T-type Ca2+ current
- 36. Кинетика низко- и высокопороговых кальциевых токов IL, IN, IP/Q и IR IT
- 38. Норма Са2+ Са2+-ток инактивирован
- 39. Калиевые токи Функции К+-каналов многообразны и выходят далеко за пределы процессов межнейронной сигнализации, в частности, в
- 40. Калиевые токи IK (К+-ток задержанного выпрямления) является главным током в мембранах большинства возбудимых клеток. При достижении
- 41. Са2+-активируемые калиевые токи IK,Ca активируются при деполяризации в условиях повышения внутриклеточной концентрации Са2+. IC увеличивается с
- 42. Норма К+
- 43. Са2+-активируемые калиевые токи IK,Ca активируются при деполяризации в условиях повышения внутриклеточной концентрации Са2+. IAHP (от англ.,
- 44. Nа+-активируемые калиевые токи Иногда (например, в отсутствие или при низкой концентрации внутриклеточного Са2+) некоторые Са2+-активируемые К+-токи
- 46. Калиевые токи IA (фазический К+-ток) выполняет свою роль в клетках наряду с IK. IA в меньшей
- 47. Норма
- 48. Разделение IK и IA
- 49. Активация и инактивация IA
- 50. Активация и инактивация IA (канал Kv4-типа) активация инактивация
- 51. Активация и инактивация IA «активный» диапазон IA тока смещен в сторону гиперполяризации по сравнению с INa
- 52. Калиевые токи IM (мускарин-чувствительный К+-ток) активируется при –65 мВ, не инактивируется со временем и блокируется стимуляцией
- 53. Норма
- 54. Калиевые токи IK,leak (ток утечки) является потенциал-независимым током, протекает по неинактивирующимся каналам и обеспечивает вклад в
- 55. Смешанные токи Эти токи представлены одним током Ih, который обозначают также как Iq (от англ, queer,
- 56. Смешанные токи Superimposed current records from a SGN in the standard solution, in the Na+-free solution,
- 58. Schematic representation of a human ventricular action potential (top panel). Numbers denote the different phases of
- 60. Моделирование эффектов влияния различных токов на паттерн импульсной активности в нейронах ЦНС млекопитающих А - ритмический
- 66. Скачать презентацию

Первые записи ионных токов, протекающие через отдельные каналы
Первые записи ионных токов, протекающие через отдельные каналы

Первые записи ионных токов, протекающие через отдельные каналы
Первые записи ионных токов, протекающие через отдельные каналы
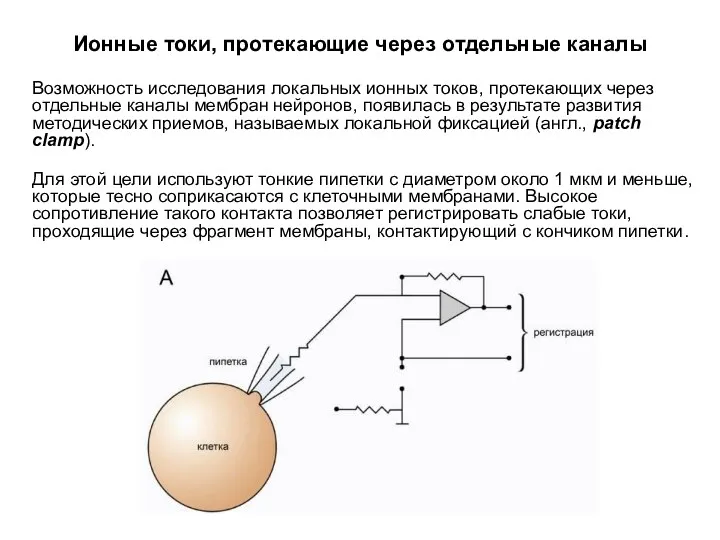
Ионные токи, протекающие через отдельные каналы
Возможность исследования локальных ионных токов,
Ионные токи, протекающие через отдельные каналы
Возможность исследования локальных ионных токов,
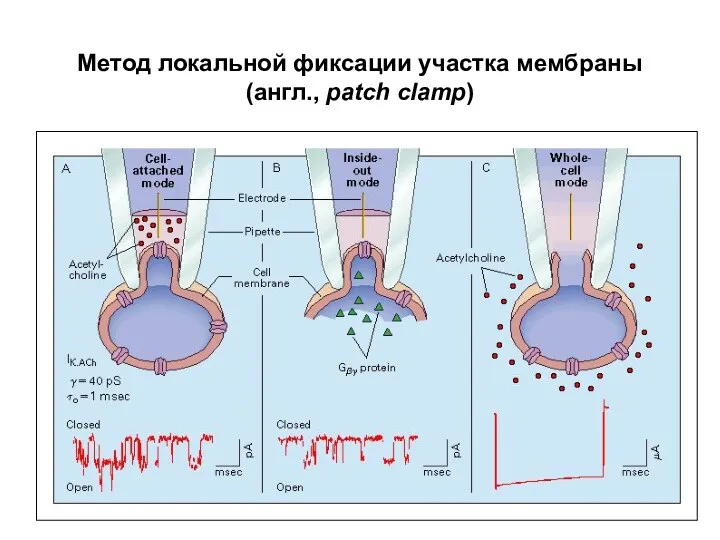
Метод локальной фиксации участка мембраны
(англ., patch clamp)
Метод локальной фиксации участка мембраны
(англ., patch clamp)
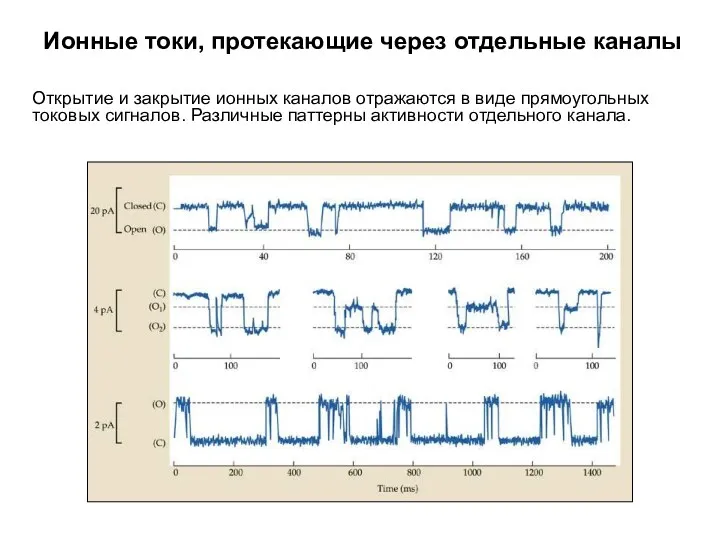
Ионные токи, протекающие через отдельные каналы
Открытие и закрытие ионных каналов
Ионные токи, протекающие через отдельные каналы
Открытие и закрытие ионных каналов
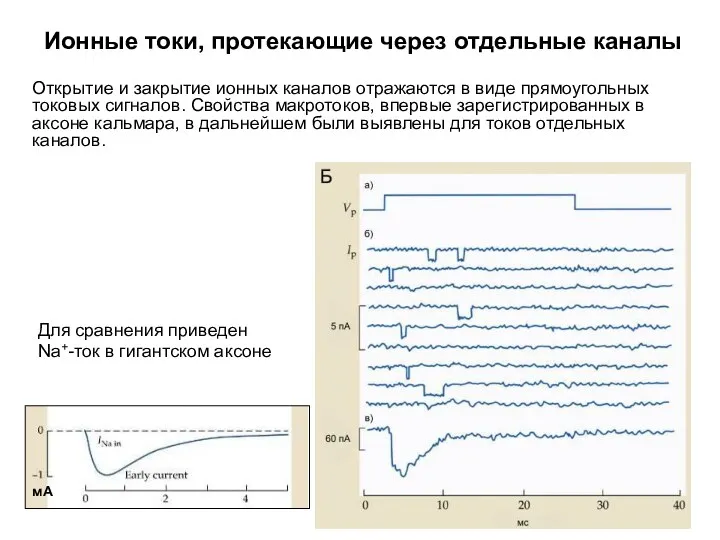
Ионные токи, протекающие через отдельные каналы
Для сравнения приведен Na+-ток в
Ионные токи, протекающие через отдельные каналы
Для сравнения приведен Na+-ток в
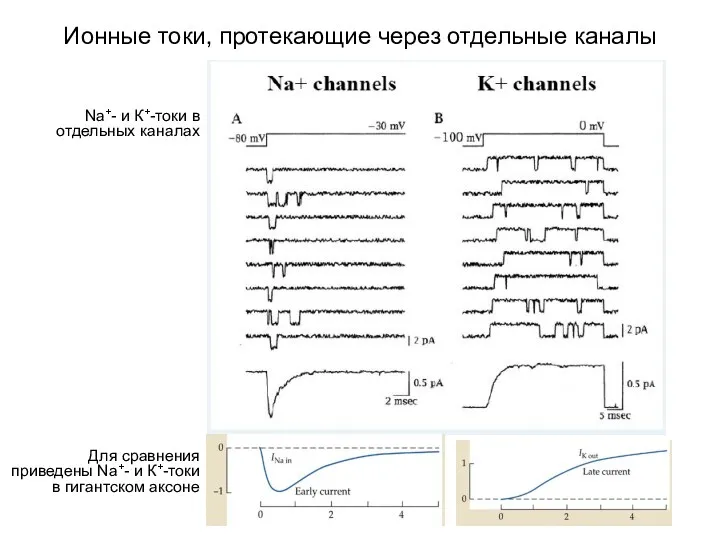
Ионные токи, протекающие через отдельные каналы
Для сравнения приведены Na+- и К+-токи
Ионные токи, протекающие через отдельные каналы
Для сравнения приведены Na+- и К+-токи
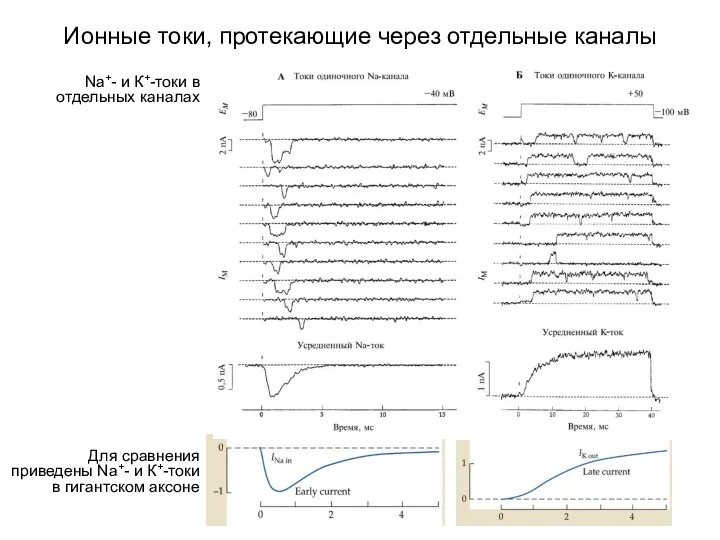
Ионные токи, протекающие через отдельные каналы
Для сравнения приведены Na+- и К+-токи
Ионные токи, протекающие через отдельные каналы
Для сравнения приведены Na+- и К+-токи
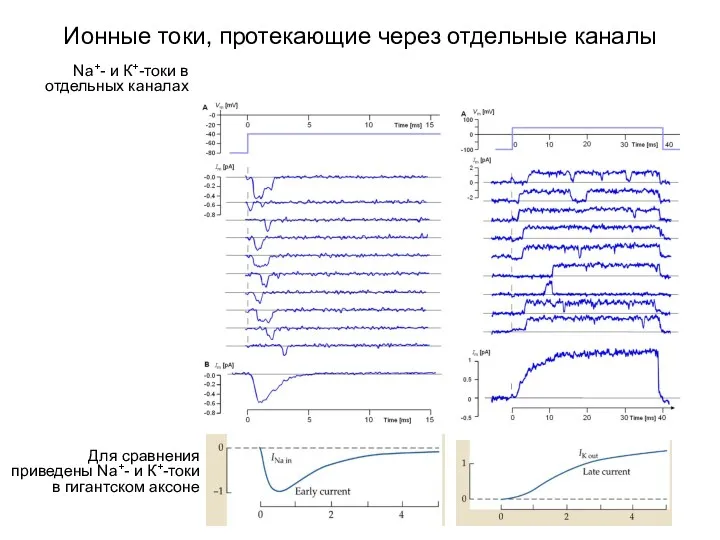
Ионные токи, протекающие через отдельные каналы
Для сравнения приведены Na+- и К+-токи
Ионные токи, протекающие через отдельные каналы
Для сравнения приведены Na+- и К+-токи

Ионные токи, протекающие через отдельные Na+-каналы
Ионные токи, протекающие через отдельные Na+-каналы
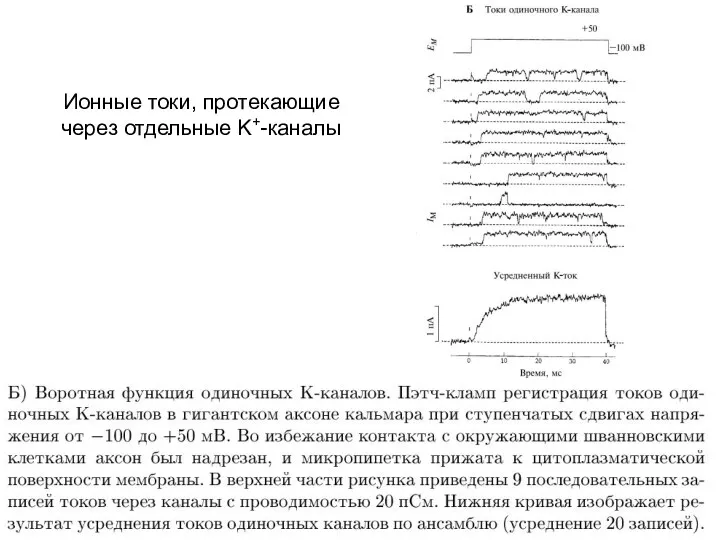
Ионные токи, протекающие через отдельные K+-каналы
Ионные токи, протекающие через отдельные K+-каналы
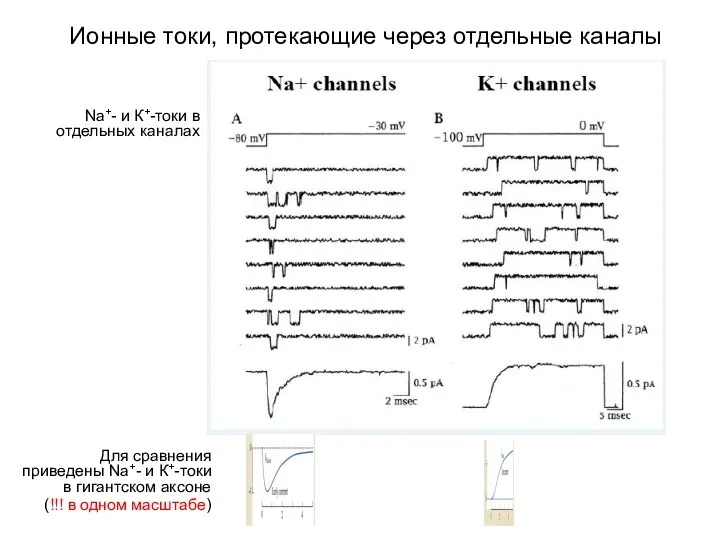
Ионные токи, протекающие через отдельные каналы
Для сравнения приведены Na+- и К+-токи
Ионные токи, протекающие через отдельные каналы
Для сравнения приведены Na+- и К+-токи

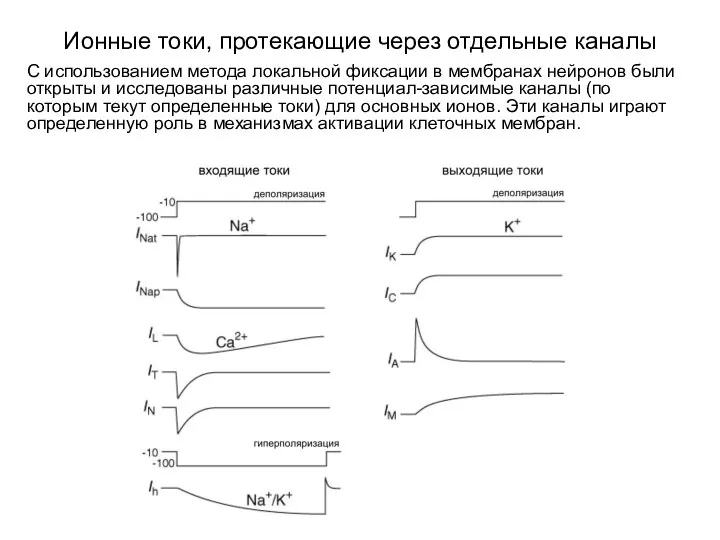
Ионные токи, протекающие через отдельные каналы
С использованием метода локальной фиксации в
Ионные токи, протекающие через отдельные каналы
С использованием метода локальной фиксации в
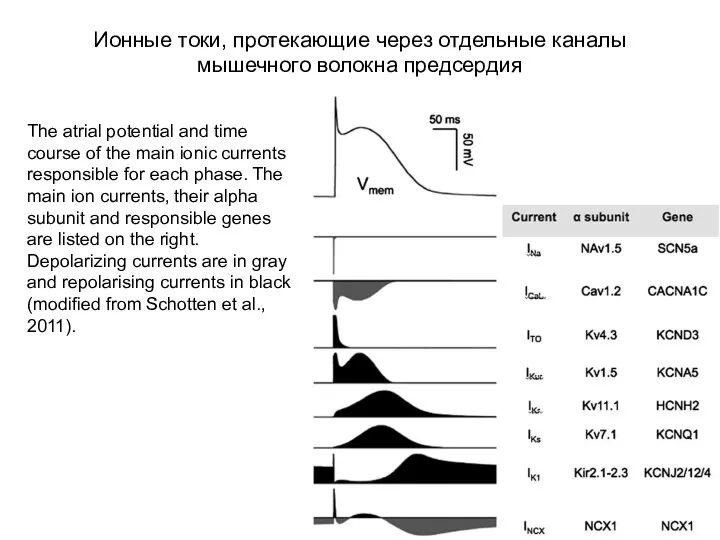
Ионные токи, протекающие через отдельные каналы
мышечного волокна предсердия
The atrial potential and
Ионные токи, протекающие через отдельные каналы
мышечного волокна предсердия
The atrial potential and
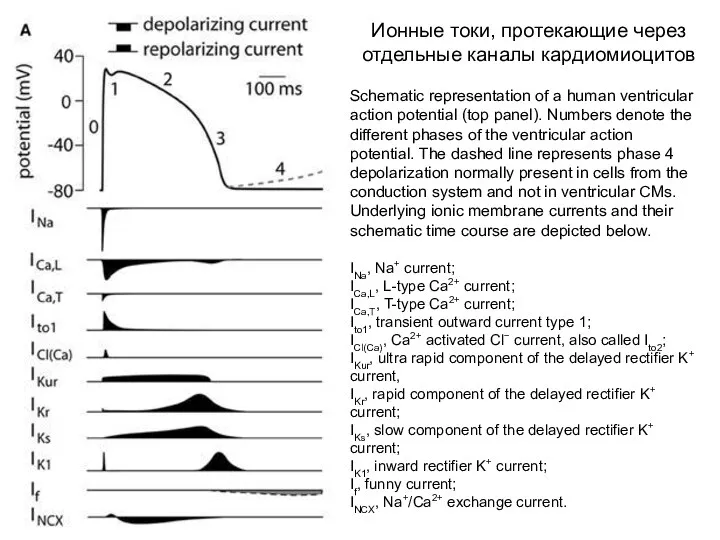
Schematic representation of a human ventricular action potential (top panel). Numbers
Schematic representation of a human ventricular action potential (top panel). Numbers

Функциональная роль потенциал-зависимых токов
Потенциал-зависимые каналы (и пропускаемые ими токи) разделяют
по
Функциональная роль потенциал-зависимых токов
Потенциал-зависимые каналы (и пропускаемые ими токи) разделяют
по

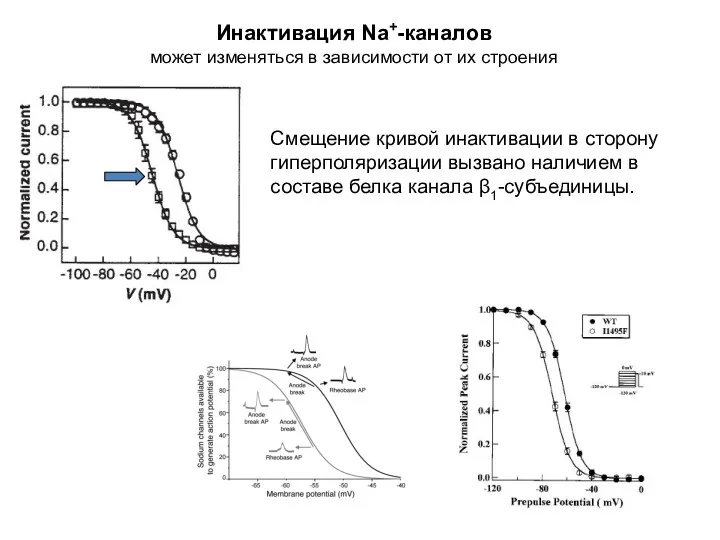
Инактивация Na+-каналов
может изменяться в зависимости от их строения
Смещение кривой инактивации в
Инактивация Na+-каналов
может изменяться в зависимости от их строения
Смещение кривой инактивации в
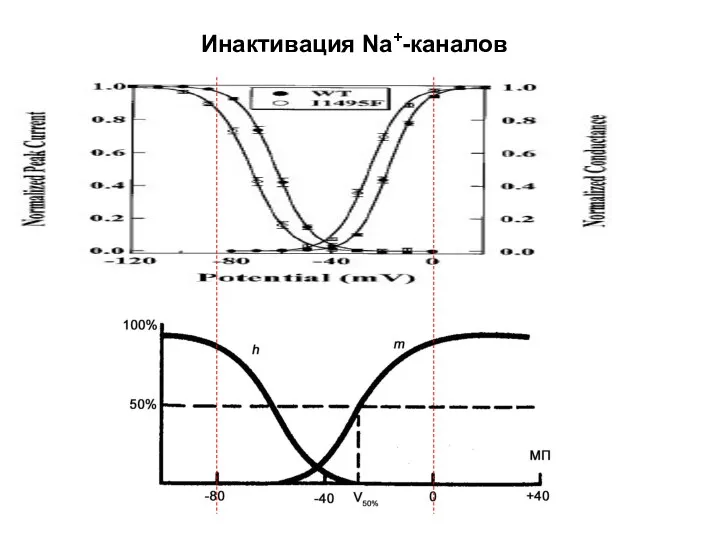
Инактивация Na+-каналов
Инактивация Na+-каналов
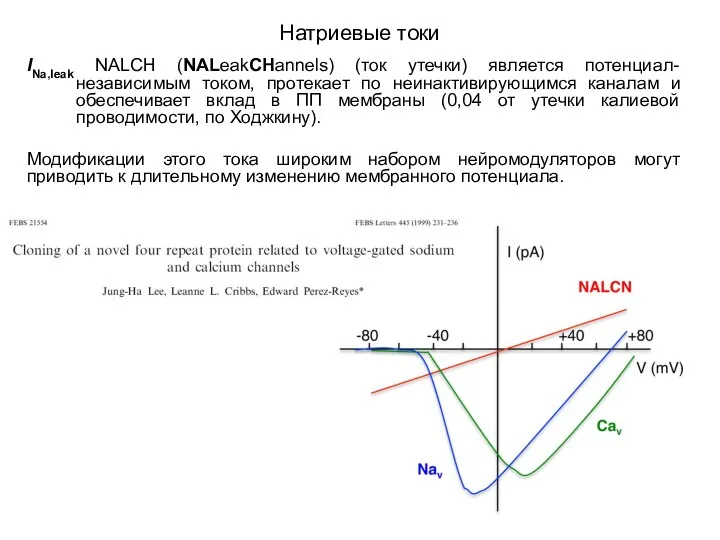
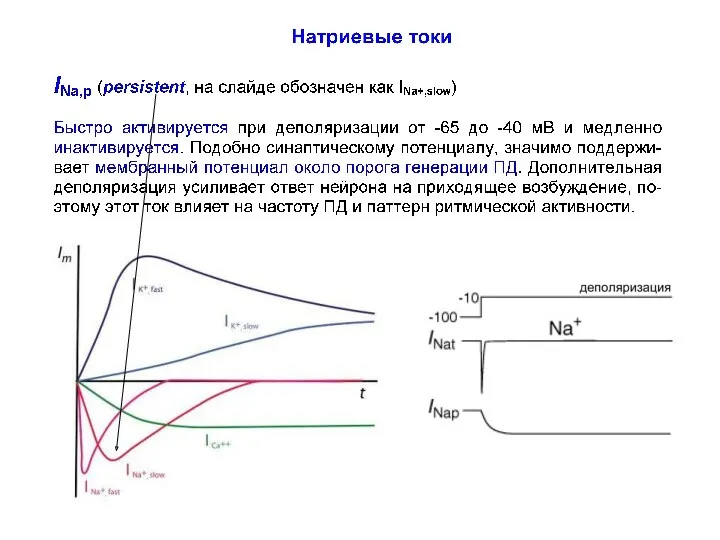
Натриевые токи
INa,leak NALCH (NALeakCHannels) (ток утечки) является потенциал-независимым током, протекает по
Натриевые токи
INa,leak NALCH (NALeakCHannels) (ток утечки) является потенциал-независимым током, протекает по

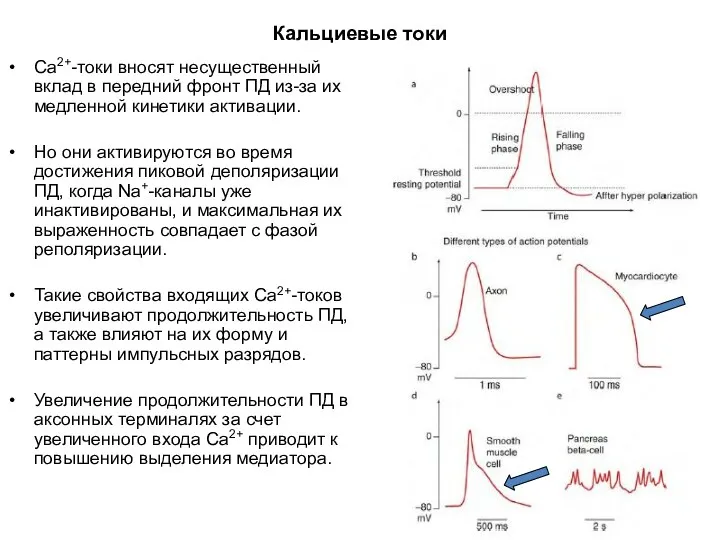
Кальциевые токи
Са2+-токи вносят несущественный вклад в передний фронт ПД из-за их
Кальциевые токи
Са2+-токи вносят несущественный вклад в передний фронт ПД из-за их

Кальциевые токи
Большинство Ca2+-каналов активируется при высоком уровне деполяризации мембраны от –20
Кальциевые токи
Большинство Ca2+-каналов активируется при высоком уровне деполяризации мембраны от –20

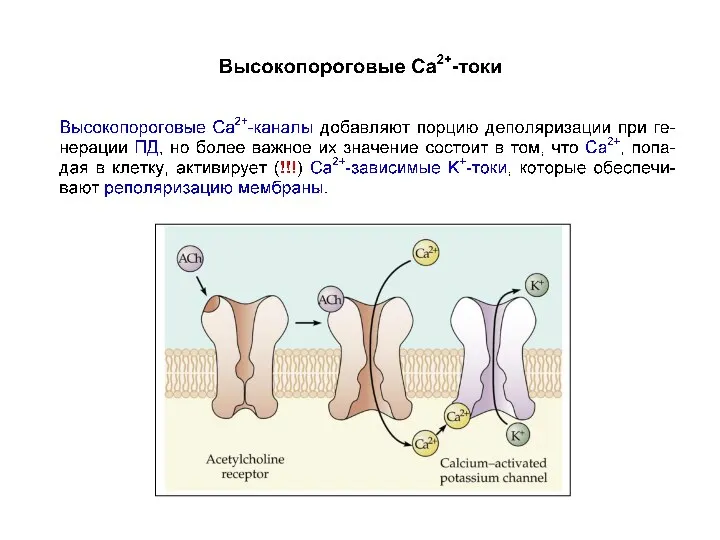
Высокопороговые кальциевые токи
Высокопороговые (порог от –20 до –10 мВ) Ca2+-токи добавляют
Высокопороговые кальциевые токи
Высокопороговые (порог от –20 до –10 мВ) Ca2+-токи добавляют
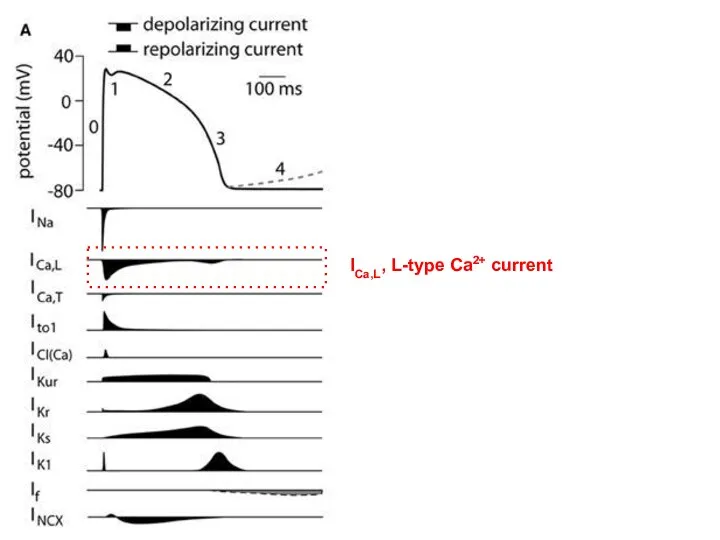
ICa,L, L-type Ca2+ current
ICa,L, L-type Ca2+ current
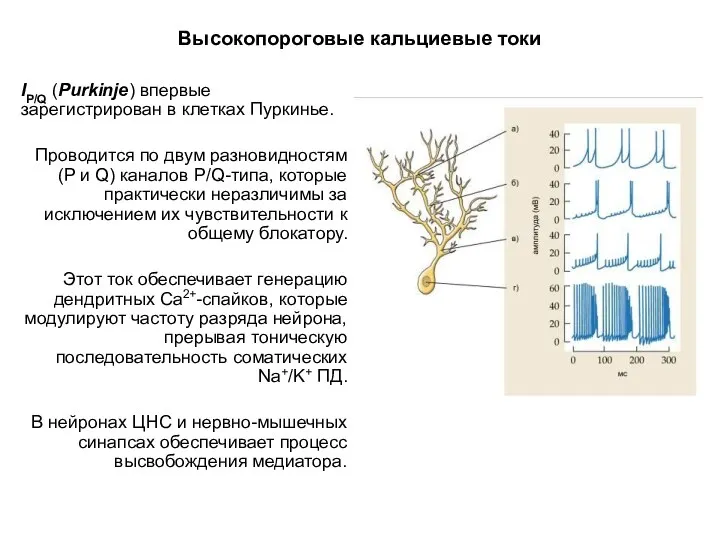
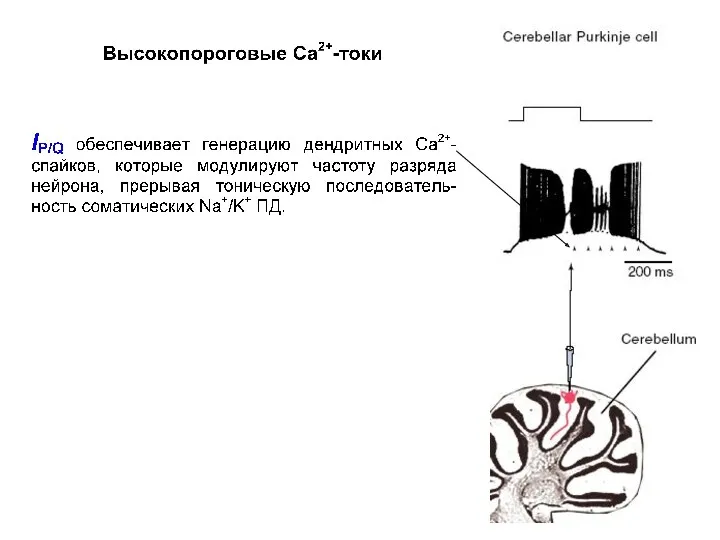
Высокопороговые кальциевые токи
IP/Q (Purkinje) впервые зарегистрирован в клетках Пуркинье.
Проводится по двум
Высокопороговые кальциевые токи
IP/Q (Purkinje) впервые зарегистрирован в клетках Пуркинье.
Проводится по двум

Высокопороговые кальциевые токи
IR (“Residual”).
Изначально канал, проводящий этот ток, был отнесен
Высокопороговые кальциевые токи
IR (“Residual”).
Изначально канал, проводящий этот ток, был отнесен
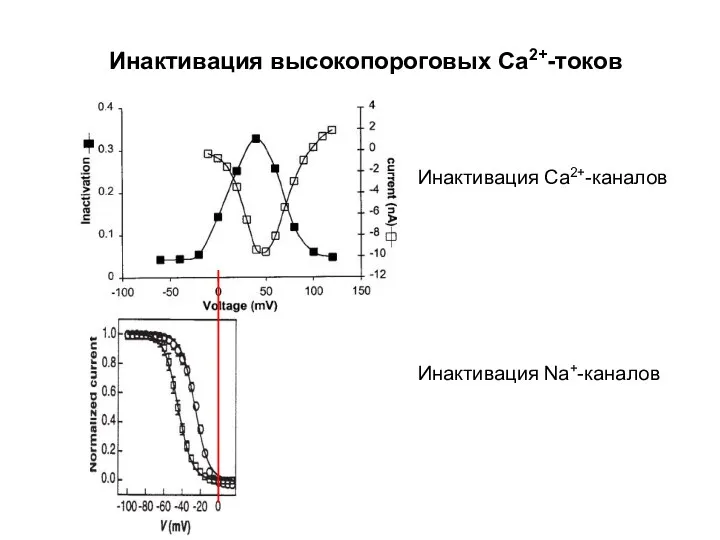
Инактивация высокопороговых Са2+-токов
Инактивация Са2+-каналов
Инактивация Na+-каналов
Инактивация высокопороговых Са2+-токов
Инактивация Са2+-каналов
Инактивация Na+-каналов
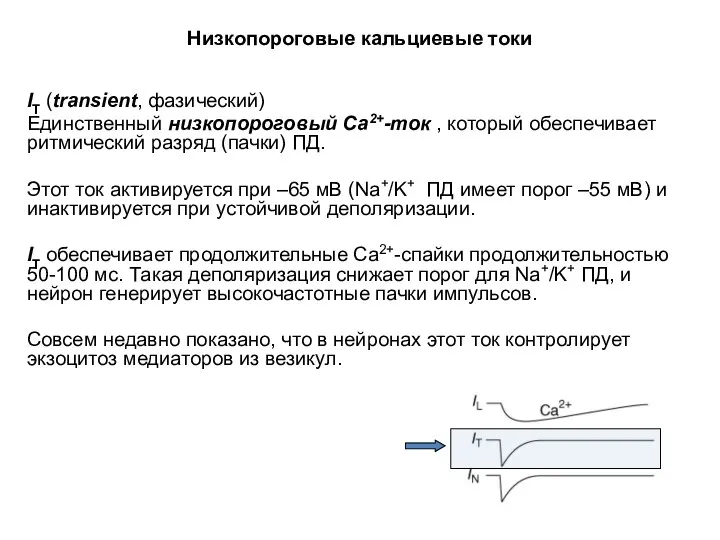
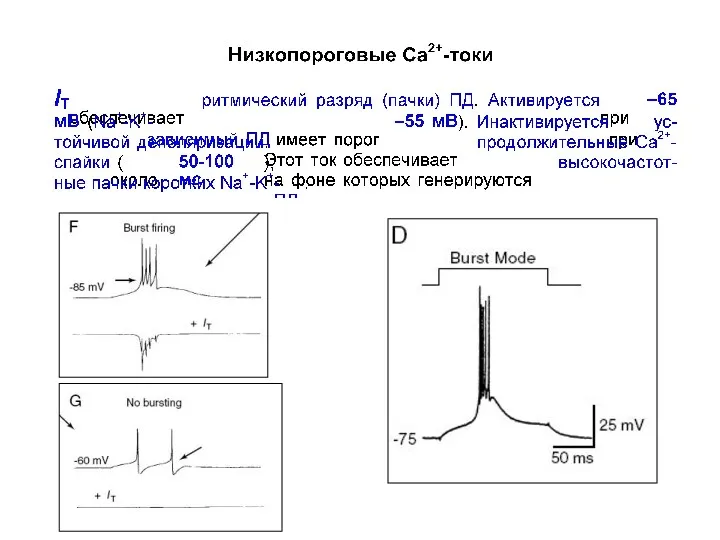
Низкопороговые кальциевые токи
IT (transient, фазический)
Единственный низкопороговый Са2+-ток , который обеспечивает ритмический
Низкопороговые кальциевые токи
IT (transient, фазический)
Единственный низкопороговый Са2+-ток , который обеспечивает ритмический
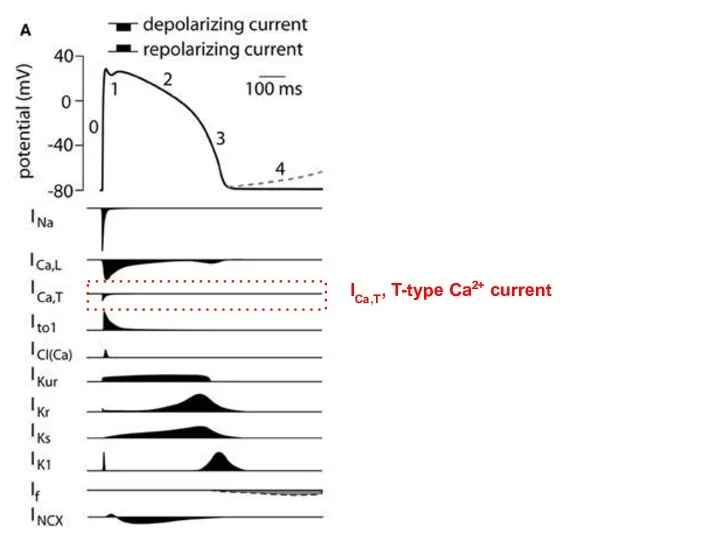
ICa,T, T-type Ca2+ current
ICa,T, T-type Ca2+ current
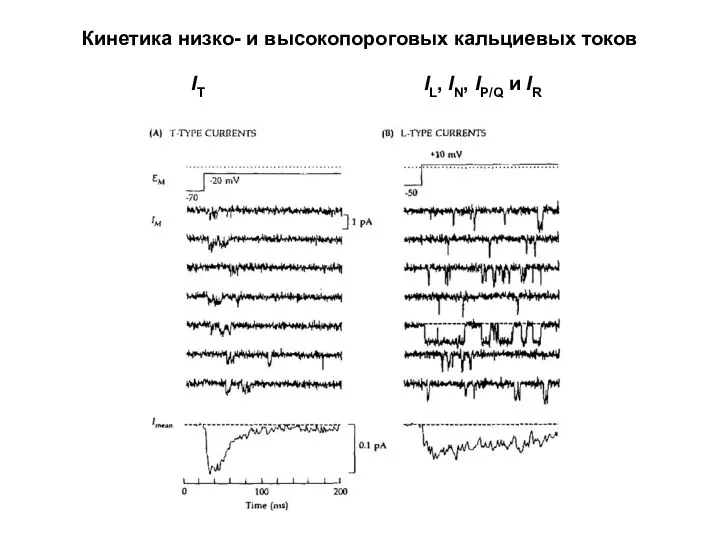
Кинетика низко- и высокопороговых кальциевых токов
IL, IN, IP/Q и IR
IT
Кинетика низко- и высокопороговых кальциевых токов
IL, IN, IP/Q и IR
IT
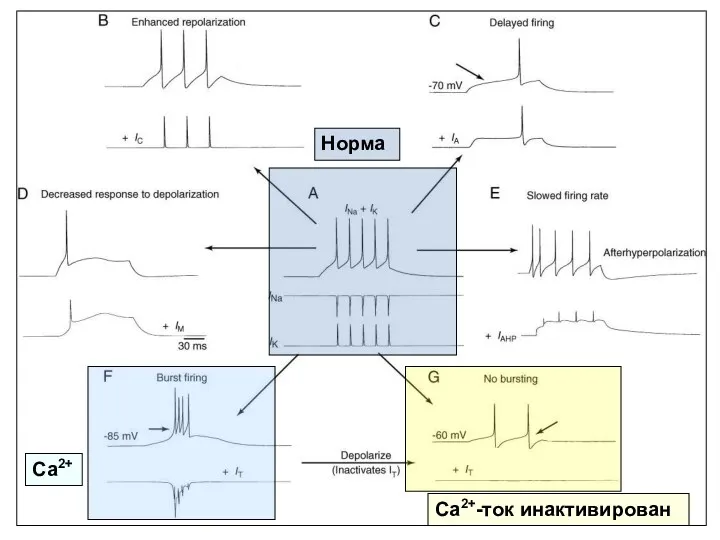
Норма
Са2+
Са2+-ток инактивирован
Норма
Са2+
Са2+-ток инактивирован

Калиевые токи
Функции К+-каналов многообразны и выходят далеко за пределы процессов межнейронной
Калиевые токи
Функции К+-каналов многообразны и выходят далеко за пределы процессов межнейронной
Калиевые токи
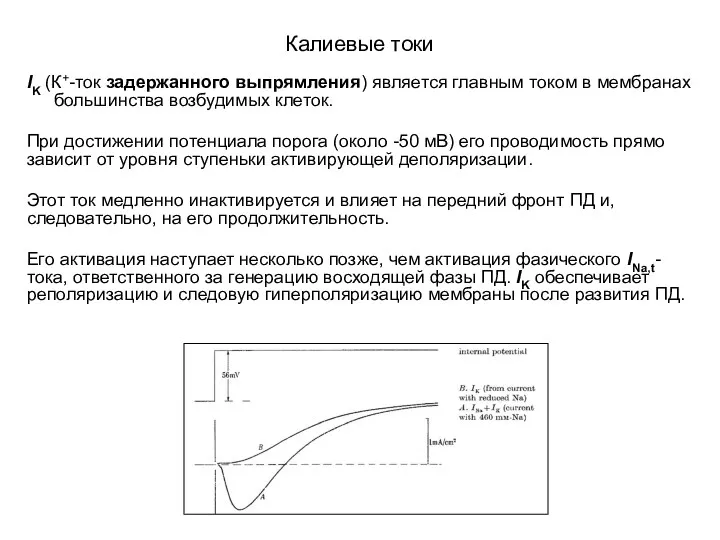
IK (К+-ток задержанного выпрямления) является главным током в мембранах большинства
Калиевые токи
IK (К+-ток задержанного выпрямления) является главным током в мембранах большинства

Са2+-активируемые калиевые токи
IK,Ca активируются при деполяризации в условиях повышения внутриклеточной концентрации
Са2+-активируемые калиевые токи
IK,Ca активируются при деполяризации в условиях повышения внутриклеточной концентрации
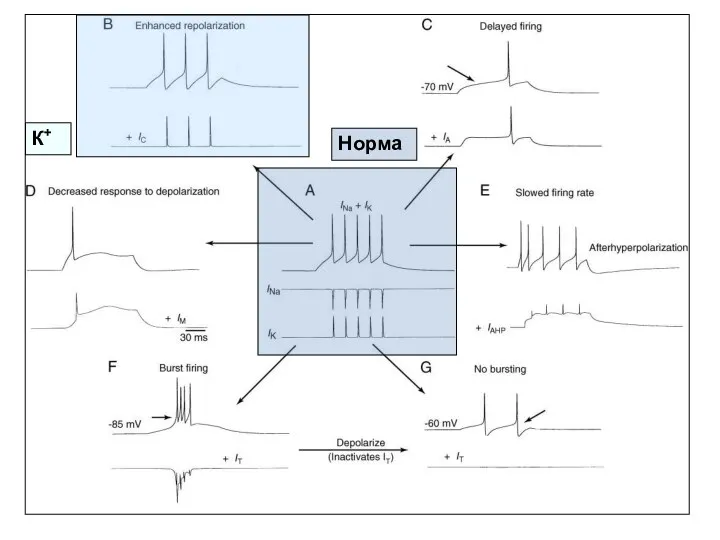
Норма
К+
Норма
К+
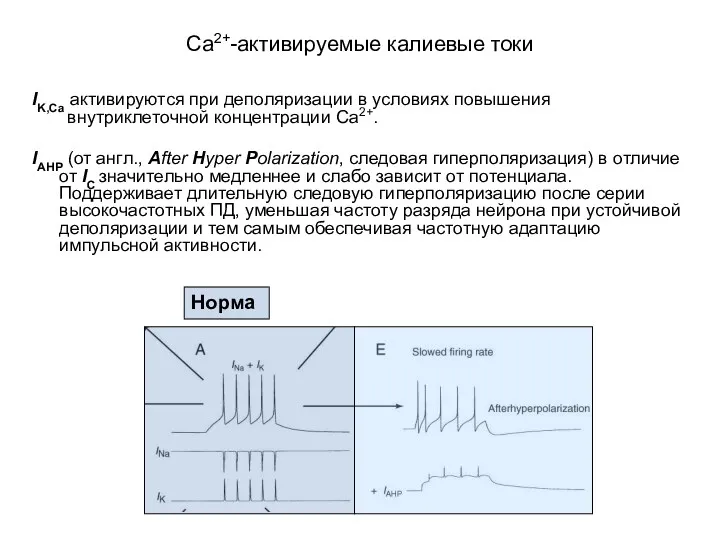
Са2+-активируемые калиевые токи
IK,Ca активируются при деполяризации в условиях повышения внутриклеточной концентрации
Са2+-активируемые калиевые токи
IK,Ca активируются при деполяризации в условиях повышения внутриклеточной концентрации

Nа+-активируемые калиевые токи
Иногда (например, в отсутствие или при низкой концентрации внутриклеточного
Nа+-активируемые калиевые токи
Иногда (например, в отсутствие или при низкой концентрации внутриклеточного
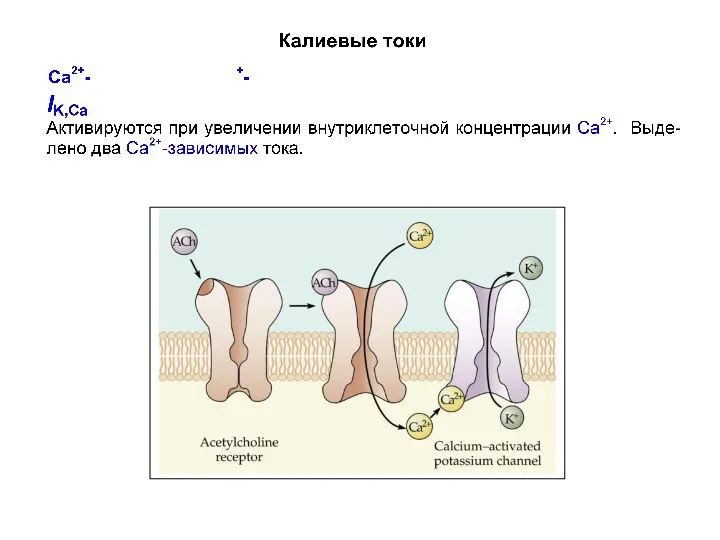
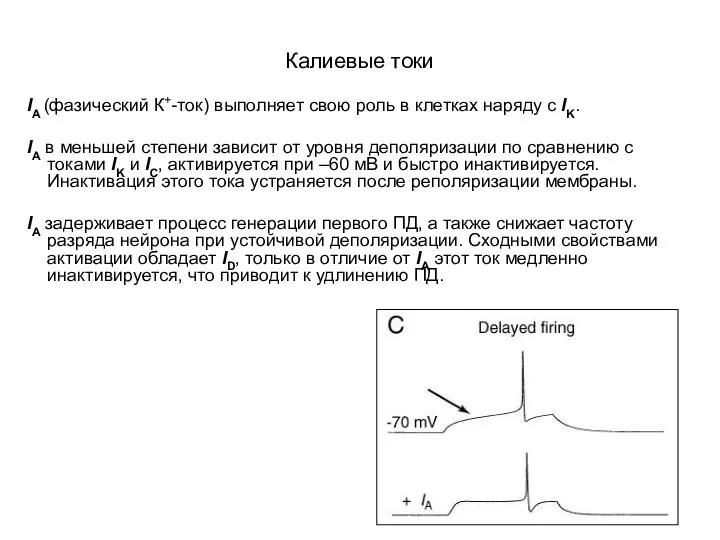
Калиевые токи
IA (фазический К+-ток) выполняет свою роль в клетках наряду с
Калиевые токи
IA (фазический К+-ток) выполняет свою роль в клетках наряду с
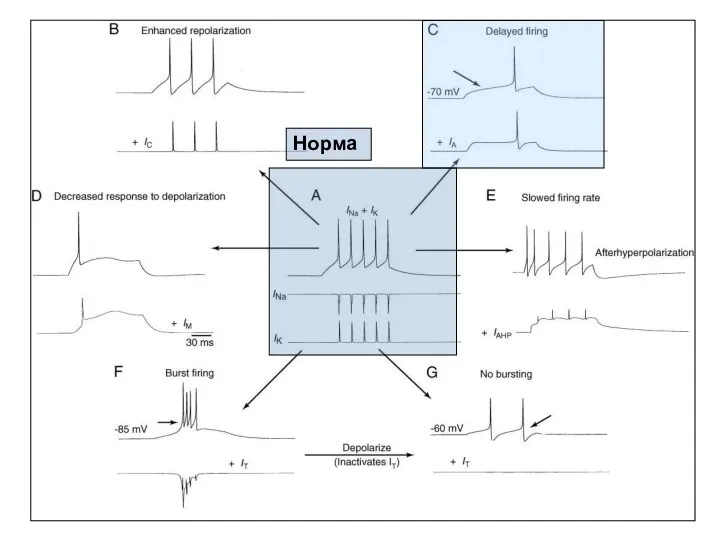
Норма
Норма
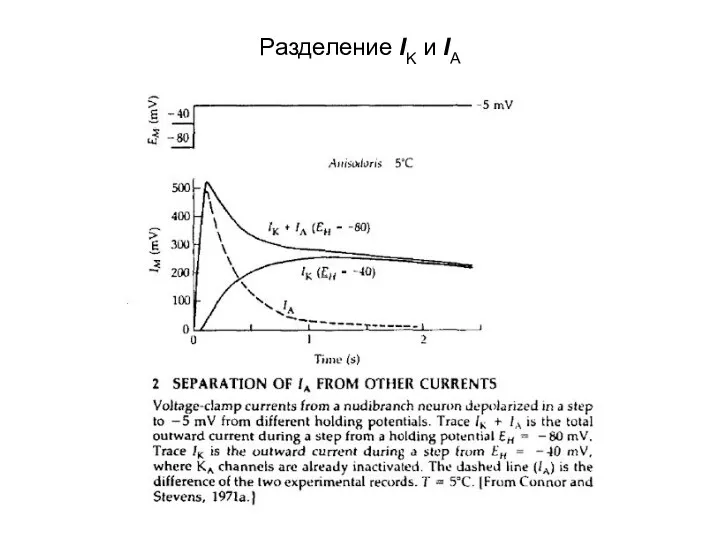
Разделение IK и IA
Разделение IK и IA
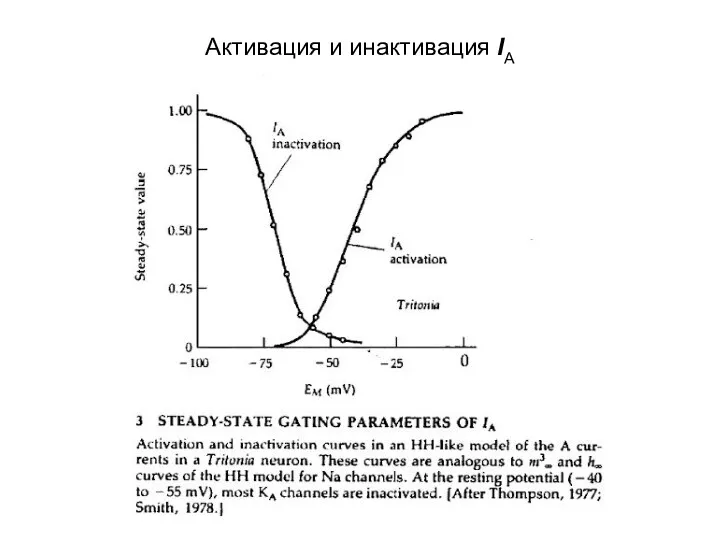
Активация и инактивация IA
Активация и инактивация IA
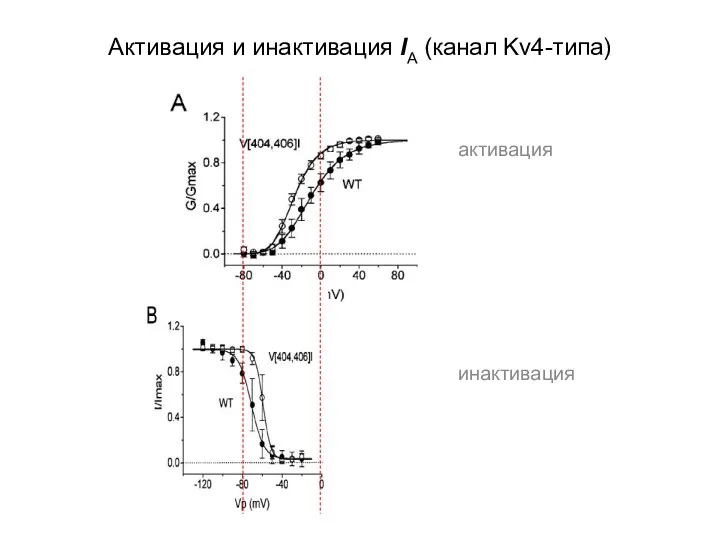
Активация и инактивация IA (канал Kv4-типа)
активация
инактивация
Активация и инактивация IA (канал Kv4-типа)
активация
инактивация
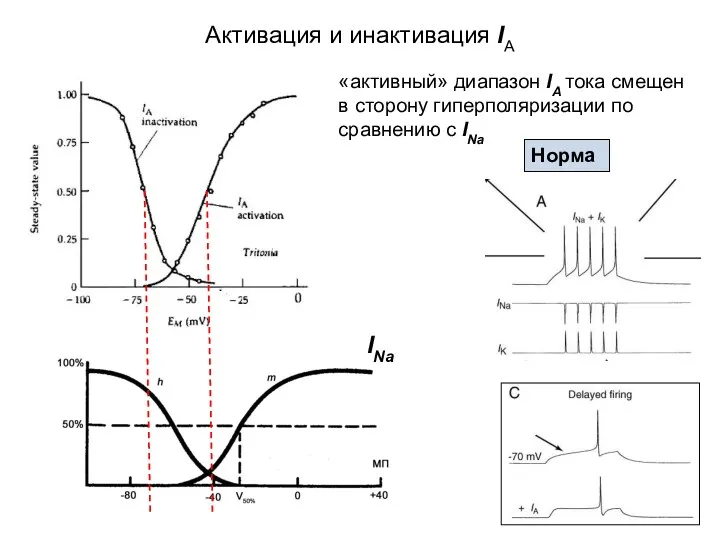
Активация и инактивация IA
«активный» диапазон IA тока смещен в сторону гиперполяризации
Активация и инактивация IA
«активный» диапазон IA тока смещен в сторону гиперполяризации
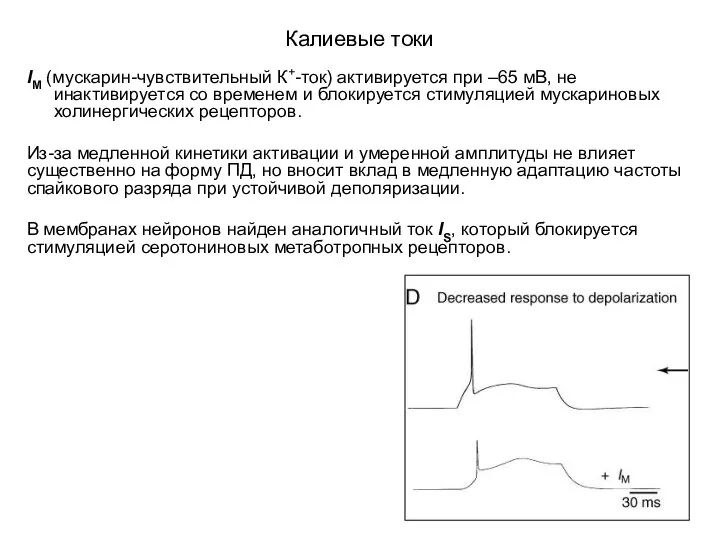
Калиевые токи
IM (мускарин-чувствительный К+-ток) активируется при –65 мВ, не инактивируется со
Калиевые токи
IM (мускарин-чувствительный К+-ток) активируется при –65 мВ, не инактивируется со
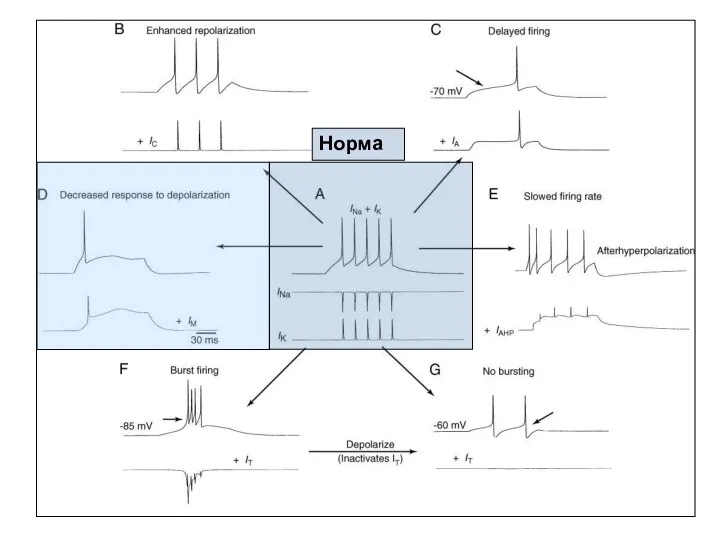
Норма
Норма
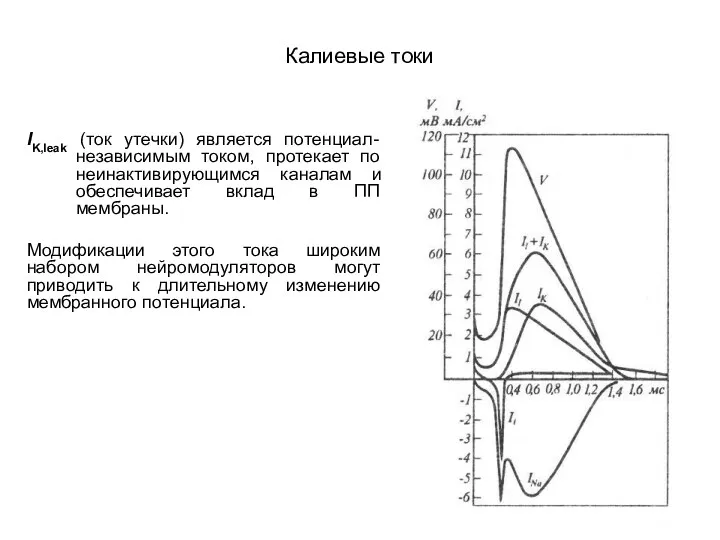
Калиевые токи
IK,leak (ток утечки) является потенциал-независимым током, протекает по неинактивирующимся каналам
Калиевые токи
IK,leak (ток утечки) является потенциал-независимым током, протекает по неинактивирующимся каналам

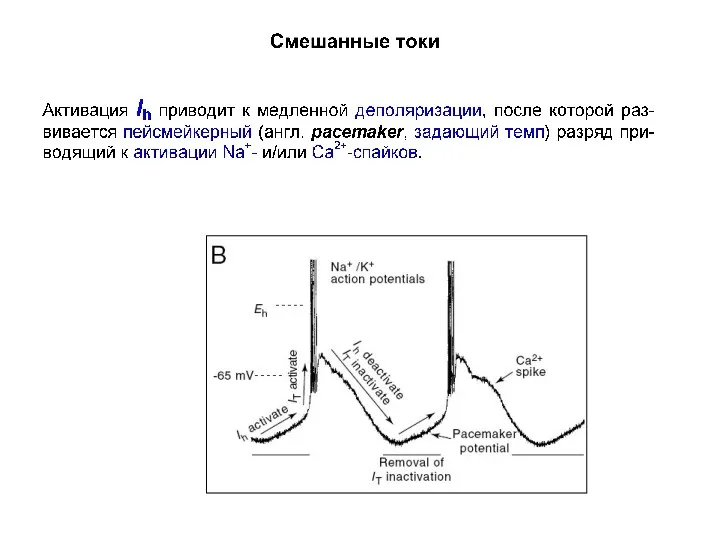
Смешанные токи
Эти токи представлены одним током Ih, который обозначают также как
Смешанные токи
Эти токи представлены одним током Ih, который обозначают также как
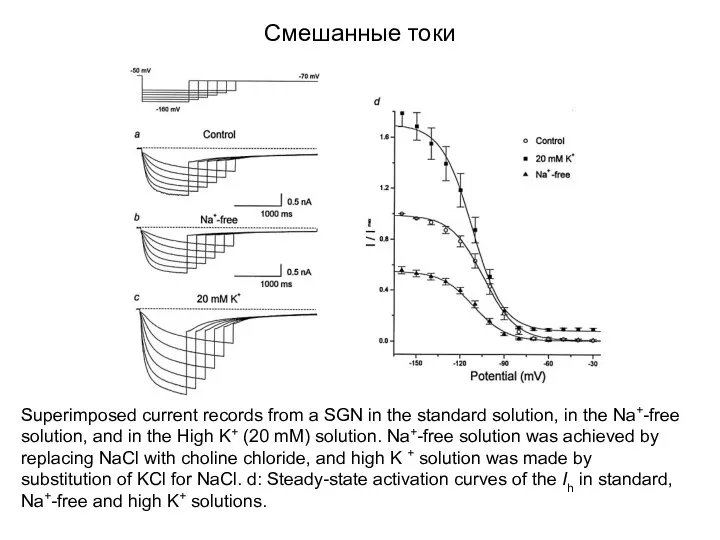
Смешанные токи
Superimposed current records from a SGN in the standard solution,
Смешанные токи
Superimposed current records from a SGN in the standard solution,
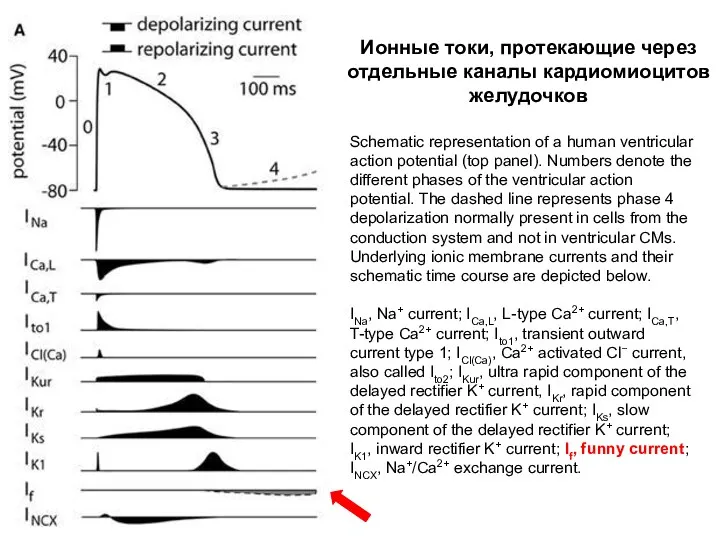
Schematic representation of a human ventricular action potential (top panel). Numbers
Schematic representation of a human ventricular action potential (top panel). Numbers
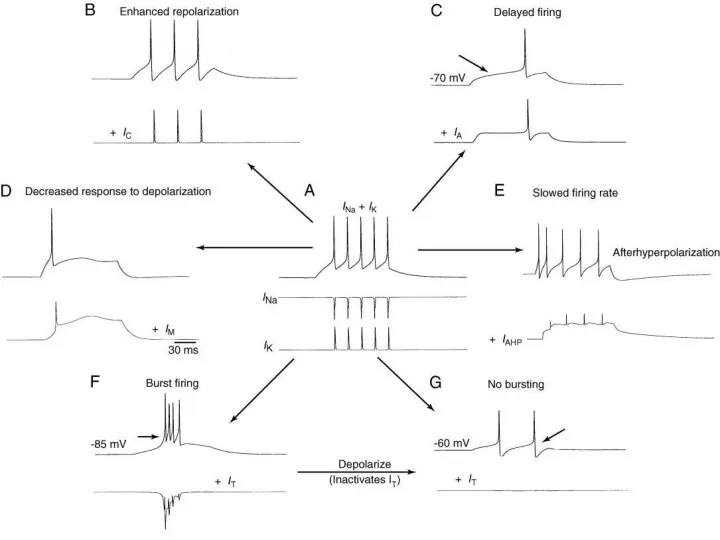

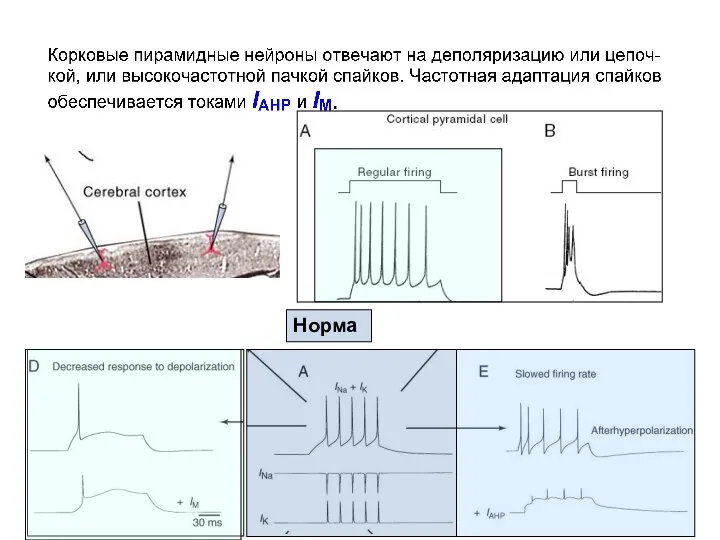
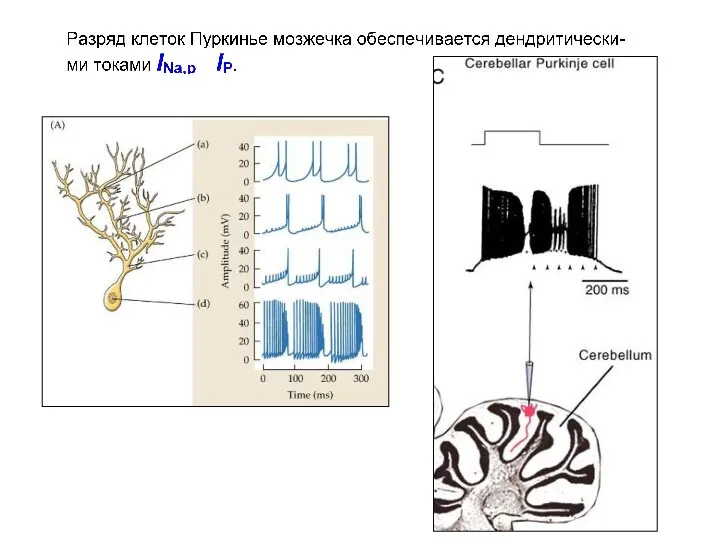
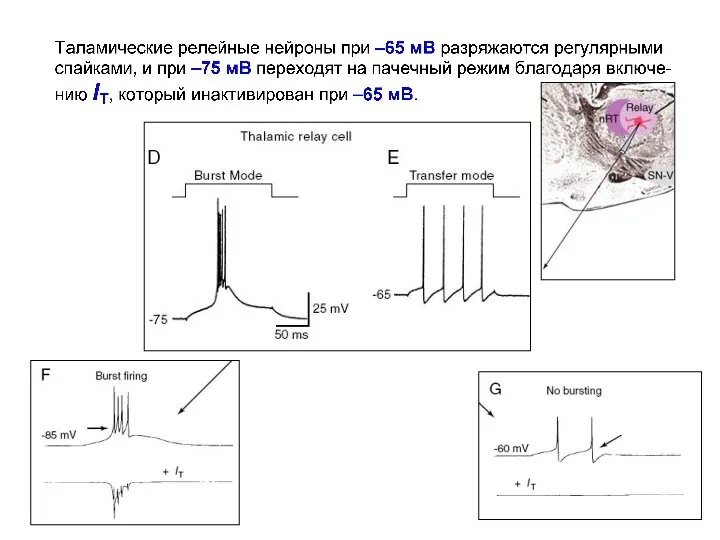
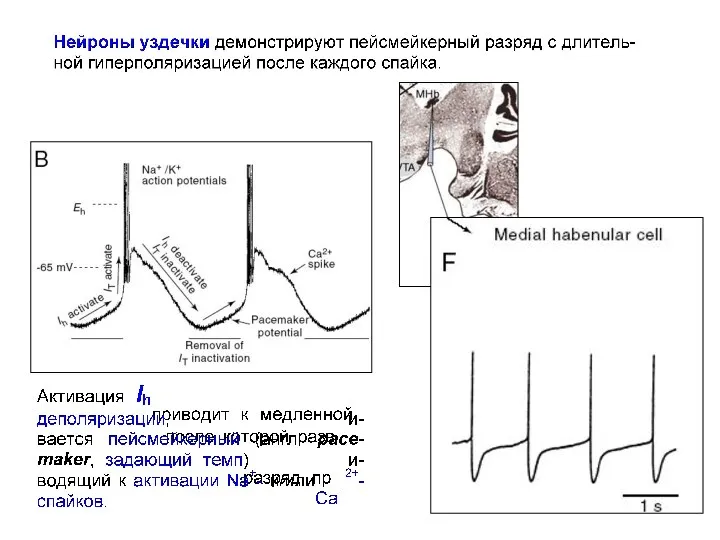
Моделирование эффектов влияния различных токов на паттерн импульсной активности в нейронах
Моделирование эффектов влияния различных токов на паттерн импульсной активности в нейронах
 Генотипическая изменчивость. Мутации
Генотипическая изменчивость. Мутации Изоляция
Изоляция Методы биологических исследований
Методы биологических исследований Презентация к уроку: Тип Плоские черви: многообразие, образ жизни и поведение. Значение в природе и жизни человека.
Презентация к уроку: Тип Плоские черви: многообразие, образ жизни и поведение. Значение в природе и жизни человека. Пути передачи сигнала. Мембранные рецепторы
Пути передачи сигнала. Мембранные рецепторы Жизнь пресного водоёма
Жизнь пресного водоёма Тип Sarcomastigophora
Тип Sarcomastigophora Трематоды и цестоды
Трематоды и цестоды разработка урока в 5 классе по теме: Грибы
разработка урока в 5 классе по теме: Грибы Использование дрожжевых грибов для производства разных групп продуктов
Использование дрожжевых грибов для производства разных групп продуктов Многообразие земноводных
Многообразие земноводных Ферменты. Специфические свойства
Ферменты. Специфические свойства Высшие жирные кислоты. Липиды
Высшие жирные кислоты. Липиды Обмен углеводов в организме человека
Обмен углеводов в организме человека Дополнительная презентация Что такое фитодизайн?к элективному курсу Основы фитодизайна, 9 класс
Дополнительная презентация Что такое фитодизайн?к элективному курсу Основы фитодизайна, 9 класс Онтогенез. Типы онтогенеза
Онтогенез. Типы онтогенеза Загальна характеристика харчування і травлення
Загальна характеристика харчування і травлення Изменчивость, ее формы и проявления. Наследственное многообразие человека
Изменчивость, ее формы и проявления. Наследственное многообразие человека Невероятные факты о животных
Невероятные факты о животных Дыхание животных. 4 класс
Дыхание животных. 4 класс Основы молекулярной биологии
Основы молекулярной биологии Современная эмбриология
Современная эмбриология Зима в жизни животных
Зима в жизни животных Углеводы и липиды
Углеводы и липиды Надотряд типичные птицы или новонёбные
Надотряд типичные птицы или новонёбные Введение в биохимию. Ферменты. Лекция № 1
Введение в биохимию. Ферменты. Лекция № 1 Красная Книга России
Красная Книга России Бактериофаги
Бактериофаги