- Пути передачи сигнала. Мембранные рецепторы

Содержание
- 2. Пути передачи сигнала
- 3. Пути сигнальной трансдукции Первым этапом действия на клетку внеклеточного химического сигнала является его связывание со специфичным
- 4. Молекулы вторичных посредников и их предшественников можно разделить на три основных типа: • Гидрофобные молекулы: нерастворимые
- 5. Общие свойства вторичных посредников • они очень быстро синтезируются и так же быстро удаляются или обезвреживаются
- 7. Выделяют несколько систем вторых посредников: Система аденилатциклаза – цАМФ Система гуанилатциклаза – цГМФ Система фосфолипаза С
- 9. Синтез вторичных посредников в клетке регулируется специфическими мембранными ферментами, активность которых, в свою очередь, связана во
- 11. В общих чертах пути сигнальной трансдукции с участием G-белков – протеинкиназ включает следующие этапы: 1. Лиганд
- 14. Система вторичных посредников «аденилатциклаза – цАМФ»
- 15. Некоторые эффекты, осуществляемые через цАМФ. Через цАМФ гипоталамические либерины (рилизинг-факторы) действуют на секреторный ответ аденогипофиза: АКТГ,
- 16. Система вторичных посредников «фосфолипаза С – инозитолтрифосфат (ИФ3) и диацилглицерол (ДАГ)»
- 17. Физиология мышц
- 18. Скелетная мускулатура Скелетные мышцы прикреплены к костям, осуществляя их поддержку и движение. Они обеспечивают передвижение в
- 19. Гладкая мускулатура Окружает полые и трубчатые органы. Гладкие мышцы обеспечивают механическую активность внутренних органов: передвижение крови
- 20. Сердечная мышца (миокард) Обеспечивает работу сердца. Благодаря ее сокращениям кровь циркулирует в сосудистой системе. Определенные ее
- 22. Гексагональное распределение толстых и тонких филаментов миофибриллы в зоне их перекрывания. Каждый толстый филамент окружен шестью
- 23. Поперечные мостики толстых филаментов, связываясь с актином тонких филаментов, подвергаются конформационному изменению, благодаря которому тонкие филаменты
- 24. Строение актина. А ‐ принцип организации молекул актина. Б – структура актинового комплекса с тропонином и
- 28. Электромеханическое сопряжение – это последовательность процессов, в результате которых потенциал действия плазматической мембраны мышечного волокна приводит
- 30. Три ресурса для образования АТФ во время мышечного сокращения: (1) креатинфосфат, (2) окислительное фосфорилирование, (3) гликолиз.
- 31. Мотонейрон вместе со всеми мышечными волокнами, которые он иннервирует, составляет двигательную единицу
- 32. Типы мышечного сокращения В результате сокращения в мышечных волокнах возникает напряжение. Если мышца укорачивается, а нагрузка
- 33. Увеличение мышечного напряжения при последовательно приходящих потенциалах действия, возникающих до окончания фазы механической активности, называется суммацией.
- 36. Участие G-белков в сопряжении гормон-рецепторного комплекса (RH) с эффекторными системами
- 37. Классификация межклеточных контактов Плотный контакт Промежуточный контакт Десмосома Септированный контакт Щелевой контакт
- 38. Структура щелевых контактов
- 39. Структура щелевых контактов
- 40. Проведение эл. тока через щелевой контакт (электрический синапс)
- 41. Механизм передачи через электрический синапс В нервном волокне потенциал действия возникает за счет разности потен‑ циалов
- 42. Основные функции электрических синапсов Основные функции электрических синапсов сводятся к следующим: 1) Быстродействие, что позволяет обеспечивать
- 43. Химические синапсы Понятие синапс ввел в 1897 году английский физиолог Cherrington Когда потенциал действия достигает пресинаптиче‑
- 44. Химические синапсы
- 45. Химические синапсы
- 46. Ионотропный рецептор
- 47. Метаботропный рецептор
- 48. Ионотропный VS Метаботропный рецептор
- 49. Типы постсинаптических потенциалов
- 50. Рецепторы и нейромедиаторы
- 51. Холинергические синапсы – синтез ацетилхолина
- 52. Холинергические синапсы - ацетилхолин
- 53. Холинергические синапсы
- 54. Адренергические рецепторы – синтез норадреналина
- 55. Адренергические рецепторы
- 56. Другие виды рецепторов
- 58. Скачать презентацию
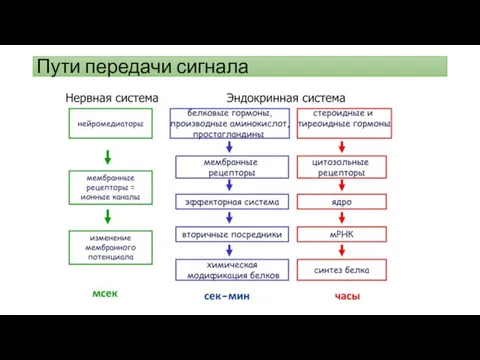
Пути передачи сигнала
Пути передачи сигнала

Пути сигнальной трансдукции
Первым этапом действия на клетку внеклеточного химического сигнала является
Пути сигнальной трансдукции
Первым этапом действия на клетку внеклеточного химического сигнала является

Молекулы вторичных посредников и их предшественников можно разделить на три основных
Молекулы вторичных посредников и их предшественников можно разделить на три основных

Общие свойства вторичных посредников
• они очень быстро синтезируются и так же
Общие свойства вторичных посредников
• они очень быстро синтезируются и так же


Выделяют несколько систем вторых посредников:
Система аденилатциклаза – цАМФ
Система гуанилатциклаза
Выделяют несколько систем вторых посредников:
Система аденилатциклаза – цАМФ
Система гуанилатциклаза


Синтез вторичных посредников в клетке регулируется специфическими мембранными ферментами, активность которых,
Синтез вторичных посредников в клетке регулируется специфическими мембранными ферментами, активность которых,
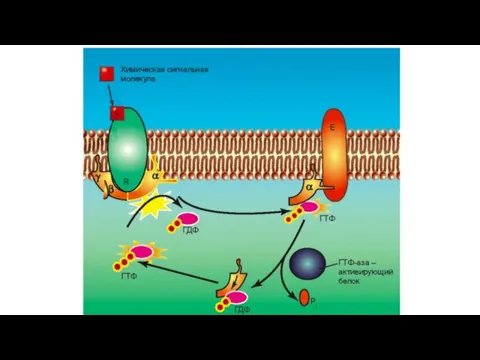

В общих чертах пути сигнальной трансдукции с участием G-белков – протеинкиназ
В общих чертах пути сигнальной трансдукции с участием G-белков – протеинкиназ

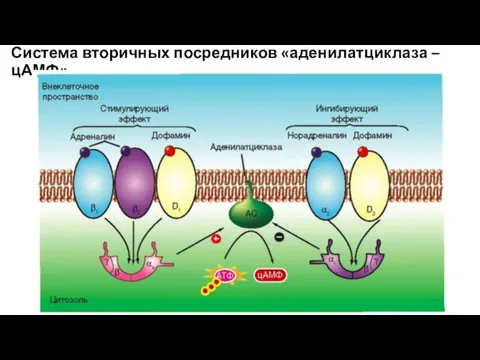
Система вторичных посредников «аденилатциклаза – цАМФ»
Система вторичных посредников «аденилатциклаза – цАМФ»

Некоторые эффекты, осуществляемые через цАМФ.
Через цАМФ гипоталамические либерины
(рилизинг-факторы) действуют
Некоторые эффекты, осуществляемые через цАМФ.
Через цАМФ гипоталамические либерины
(рилизинг-факторы) действуют
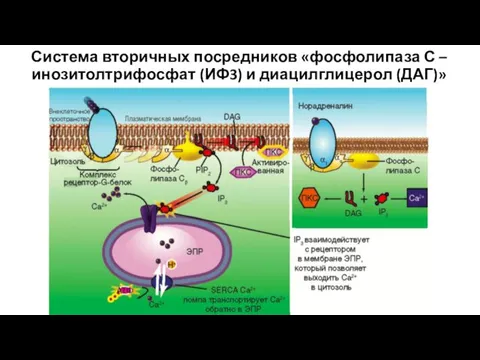
Система вторичных посредников «фосфолипаза С – инозитолтрифосфат (ИФ3) и диацилглицерол (ДАГ)»
Система вторичных посредников «фосфолипаза С – инозитолтрифосфат (ИФ3) и диацилглицерол (ДАГ)»

Физиология мышц
Физиология мышц

Скелетная мускулатура
Скелетные мышцы прикреплены к костям, осуществляя их поддержку и движение.
Скелетная мускулатура
Скелетные мышцы прикреплены к костям, осуществляя их поддержку и движение.

Гладкая мускулатура
Окружает полые и трубчатые органы. Гладкие мышцы обеспечивают механическую активность
Гладкая мускулатура
Окружает полые и трубчатые органы. Гладкие мышцы обеспечивают механическую активность

Сердечная мышца (миокард)
Обеспечивает работу сердца. Благодаря ее сокращениям кровь циркулирует в
Сердечная мышца (миокард)
Обеспечивает работу сердца. Благодаря ее сокращениям кровь циркулирует в
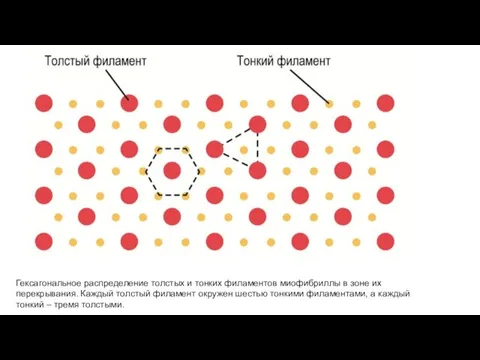
Гексагональное распределение толстых и тонких филаментов миофибриллы в зоне их перекрывания.
Гексагональное распределение толстых и тонких филаментов миофибриллы в зоне их перекрывания.
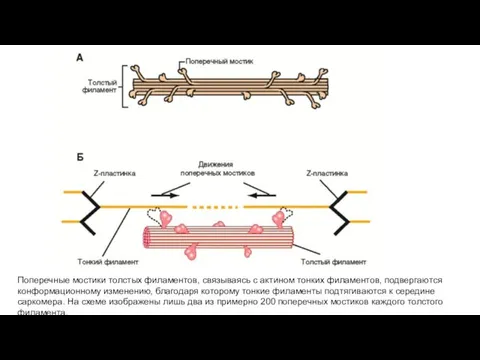
Поперечные мостики толстых филаментов, связываясь с актином тонких филаментов, подвергаются конформационному
Поперечные мостики толстых филаментов, связываясь с актином тонких филаментов, подвергаются конформационному
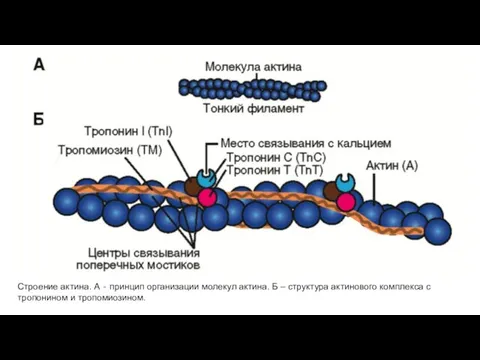
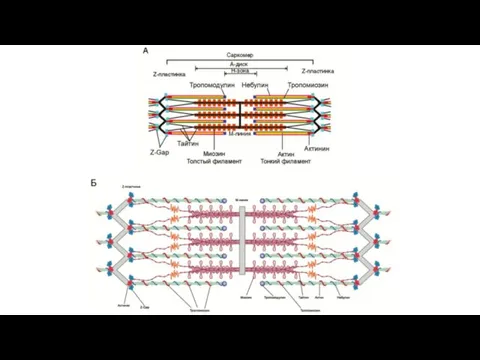
Строение актина. А ‐ принцип организации молекул актина. Б – структура
Строение актина. А ‐ принцип организации молекул актина. Б – структура
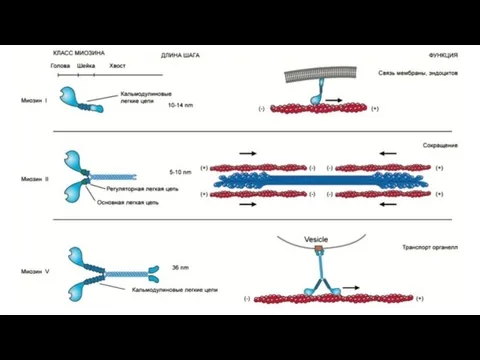
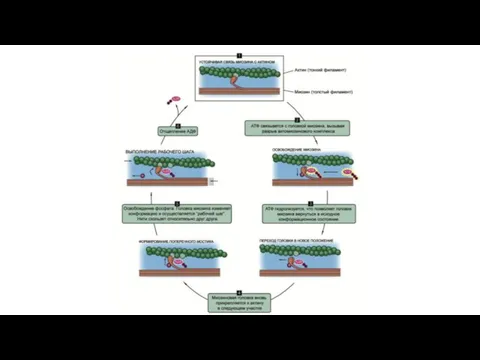
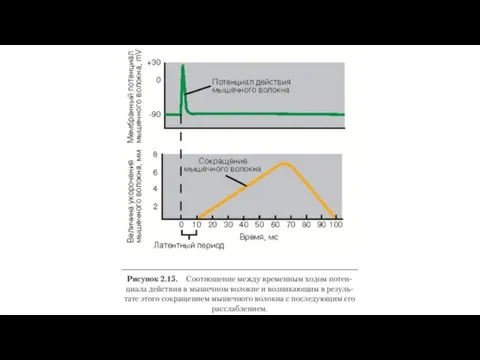

Электромеханическое сопряжение – это последовательность процессов, в результате которых потенциал действия
Электромеханическое сопряжение – это последовательность процессов, в результате которых потенциал действия

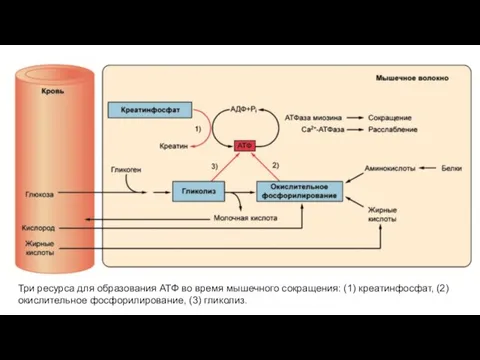
Три ресурса для образования АТФ во время мышечного сокращения: (1) креатинфосфат,
Три ресурса для образования АТФ во время мышечного сокращения: (1) креатинфосфат,
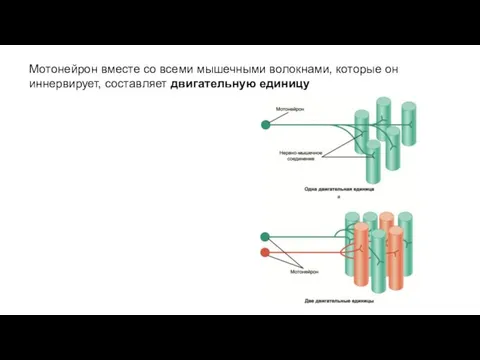
Мотонейрон вместе со всеми мышечными волокнами, которые он иннервирует, составляет двигательную
Мотонейрон вместе со всеми мышечными волокнами, которые он иннервирует, составляет двигательную

Типы мышечного сокращения
В результате сокращения в мышечных волокнах возникает напряжение.
Типы мышечного сокращения
В результате сокращения в мышечных волокнах возникает напряжение.

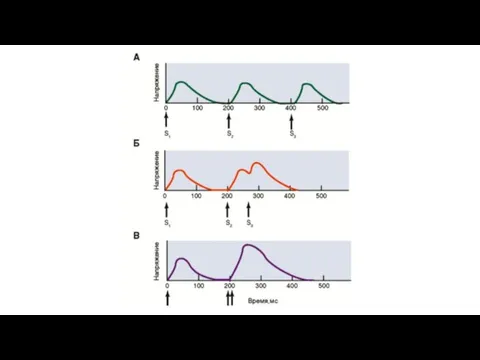
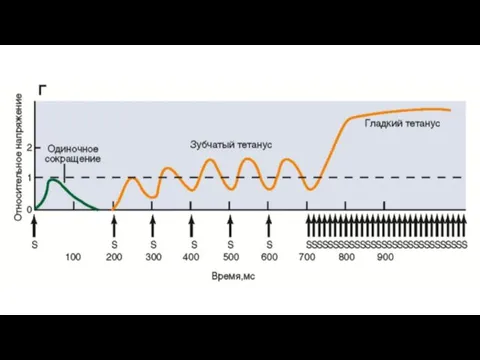
Увеличение мышечного напряжения при последовательно приходящих потенциалах действия, возникающих до окончания
Увеличение мышечного напряжения при последовательно приходящих потенциалах действия, возникающих до окончания
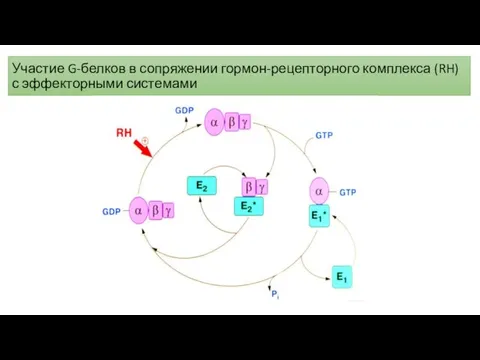
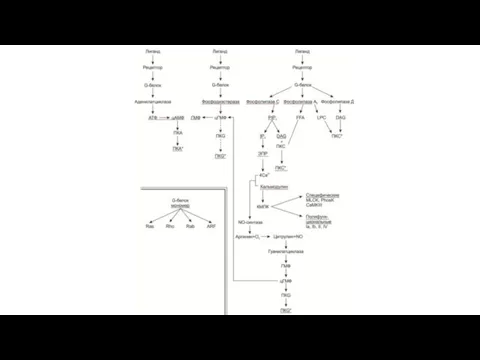
Участие G-белков в сопряжении гормон-рецепторного комплекса (RH) с эффекторными системами
Участие G-белков в сопряжении гормон-рецепторного комплекса (RH) с эффекторными системами
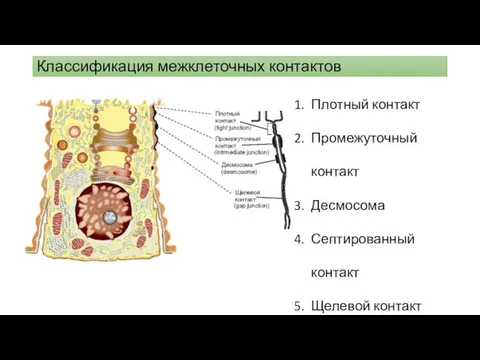
Классификация межклеточных контактов
Плотный контакт
Промежуточный контакт
Десмосома
Септированный контакт
Щелевой контакт
Классификация межклеточных контактов
Плотный контакт
Промежуточный контакт
Десмосома
Септированный контакт
Щелевой контакт
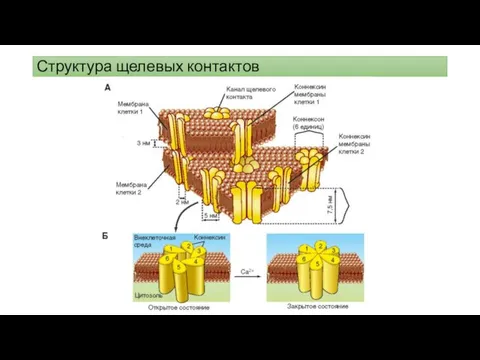
Структура щелевых контактов
Структура щелевых контактов
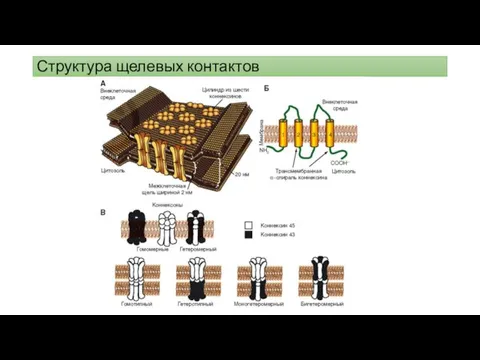
Структура щелевых контактов
Структура щелевых контактов
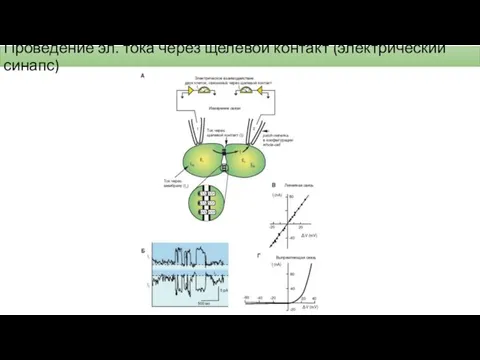
Проведение эл. тока через щелевой контакт (электрический синапс)
Проведение эл. тока через щелевой контакт (электрический синапс)

Механизм передачи через электрический синапс
В нервном волокне
потенциал действия возникает за счет
Механизм передачи через электрический синапс
В нервном волокне потенциал действия возникает за счет

Основные функции электрических синапсов
Основные функции электрических синапсов
сводятся к следующим:
1) Быстродействие, что
Основные функции электрических синапсов
Основные функции электрических синапсов
сводятся к следующим:
1) Быстродействие, что

Химические синапсы
Понятие синапс ввел в 1897 году английский физиолог Cherrington
Когда
Химические синапсы
Понятие синапс ввел в 1897 году английский физиолог Cherrington
Когда
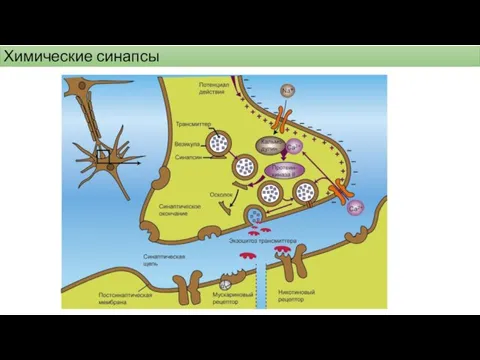
Химические синапсы
Химические синапсы
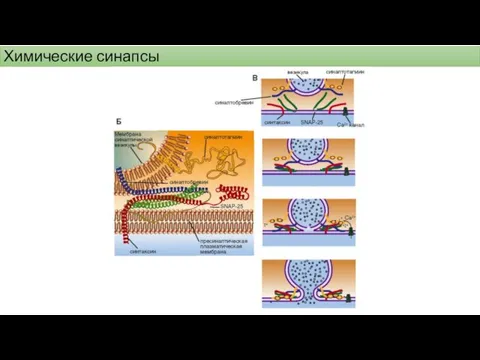
Химические синапсы
Химические синапсы
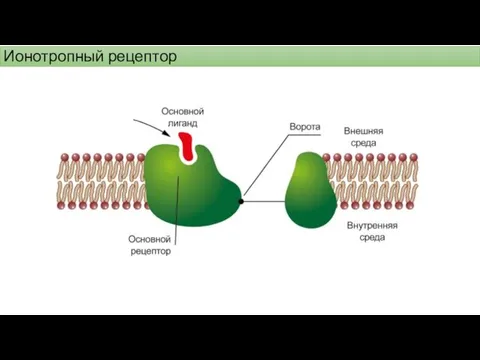
Ионотропный рецептор
Ионотропный рецептор
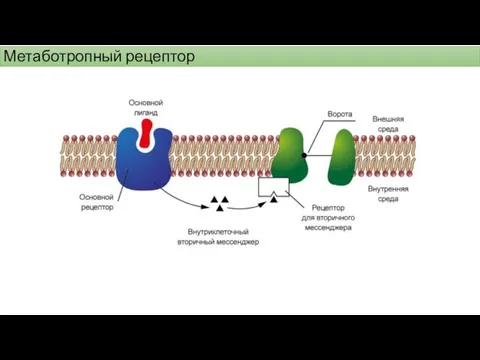
Метаботропный рецептор
Метаботропный рецептор
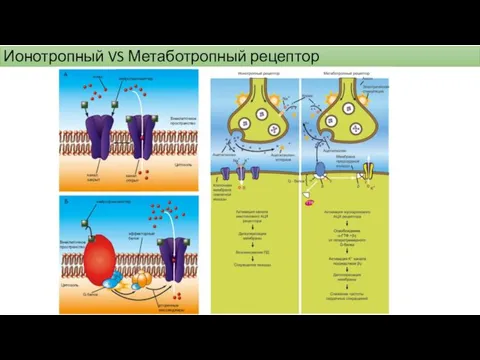
Ионотропный VS Метаботропный рецептор
Ионотропный VS Метаботропный рецептор
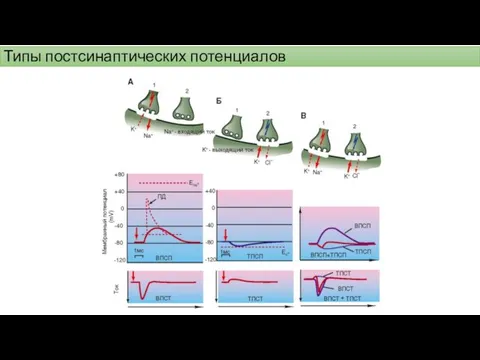
Типы постсинаптических потенциалов
Типы постсинаптических потенциалов
Рецепторы и нейромедиаторы
Рецепторы и нейромедиаторы

Холинергические синапсы – синтез ацетилхолина
Холинергические синапсы – синтез ацетилхолина

Холинергические синапсы - ацетилхолин
Холинергические синапсы - ацетилхолин
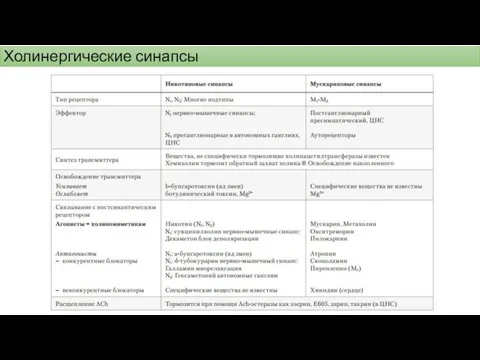
Холинергические синапсы
Холинергические синапсы
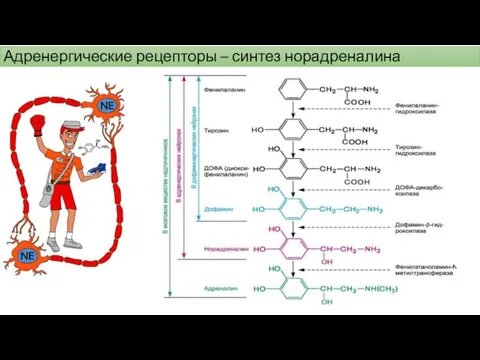
Адренергические рецепторы – синтез норадреналина
Адренергические рецепторы – синтез норадреналина

Адренергические рецепторы
Адренергические рецепторы

Другие виды рецепторов
Другие виды рецепторов
 Investigate DNA Extraction
Investigate DNA Extraction Как питаются паразиты? 1 часть. 5 класс
Как питаются паразиты? 1 часть. 5 класс Врожденные пороки сердца
Врожденные пороки сердца Урок-игра Кровь и кровообращение
Урок-игра Кровь и кровообращение Экологическая игра Подводный мир
Экологическая игра Подводный мир Кто хочет стать биологом. Игра (1)
Кто хочет стать биологом. Игра (1) Метаболизм клетки. Клеточное дыхание. Фотосинтез, хемосинтез
Метаболизм клетки. Клеточное дыхание. Фотосинтез, хемосинтез Биологическое и социальное в человеке
Биологическое и социальное в человеке Қан және лимфа
Қан және лимфа Подтип Хелицеровые (Сhelicerata)
Подтип Хелицеровые (Сhelicerata) Размножение растений
Размножение растений Клетка как структурная основа живых организмов. 10 класс
Клетка как структурная основа живых организмов. 10 класс Maduva spinarii
Maduva spinarii Введение 01 02 2016
Введение 01 02 2016 Құстар класыи- Aves
Құстар класыи- Aves Использование микробов в пищевой промышленности
Использование микробов в пищевой промышленности 1 апреля – международный день птиц!
1 апреля – международный день птиц! Идиоадаптации. 2 часть
Идиоадаптации. 2 часть Ant Life Cycle
Ant Life Cycle Нерыбные объекты водного промысла
Нерыбные объекты водного промысла Клеточное строение растений
Клеточное строение растений Безусловные и условные рефлексы
Безусловные и условные рефлексы Анатомия и физиология сердечно-сосудистой системы
Анатомия и физиология сердечно-сосудистой системы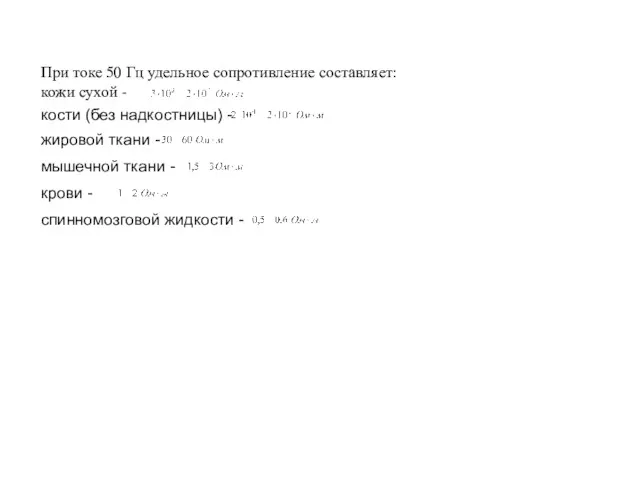 Сопротивление тела человека
Сопротивление тела человека Учение Чарльза Дарвина об искусственном отборе
Учение Чарльза Дарвина об искусственном отборе Систематические группы птиц
Систематические группы птиц Отряды млекопитающих
Отряды млекопитающих Что помогло фасолинке вырасти?
Что помогло фасолинке вырасти?