- Генетическая инженерия в современной селекции и семеноводстве растений

Содержание
- 2. Содержание лекции: 1. Геномная и хромосомная инженерия 2. Генная инженерия М.М.П. 5. СМсспк
- 3. 5.1 1. Геномная и хромосомная инженерия М.М.П. СМсспк
- 4. Поэтому понятие генетической инженерии объединяет понятия геномной, хромосомной и генной инженерии. Под генетической инженерией понимают использование
- 5. 5.1 Сущность геномной ДНК-технологии (геномной инженерии) заключается в целенаправленной перестройке генома того или иного вида растений,
- 6. 5.1 М.М.П. Хромосомная инженерия связана с замещением хромосом у растений на внутривидовом, межвидовом и даже межродовом
- 7. 5.1 Природная геномная и хромосомная инженерия создала полиплоидные комплексы видов растений. При этом в ходе видообразования
- 8. 5.1 Известно, что в клетках каждого диплоидного организма имеются пары гомологичных хромосом. Такой организм называют дисомиком.
- 9. Другой методический прием хромосомной инженерии состоит не в замещении в геноме каких-либо хромосом, а во введении
- 10. Примером геномной инженерии является создание нового рода культурных злаков – тритикале. Получена эта культура путём отдалённой
- 11. М.М.П. Схема получения октоплоидных и гексаплоидных форм тритикале 5.1 СМсспк
- 12. В хромосомной инженерии очень перспективен метод гаплоидов, основанный на выращивании гаплоидных растений с последующим удвоением хромосом.
- 13. МЕТОД ПОЛУЧЕНИЯ ДИГАПЛОИДНЫХ ГОМОЗИГОТНЫХ ЛИНИЙ (ВНИИ сахарной свеклы) Метод позволяет получать гомозиготный материал из обогащенных в
- 14. Технология получения гомозиготных линий злаков (ГНУ НИИСХ Юго-Востока) 1. Отбор донорных растений и культивирование пыльников на
- 15. 5.2 2. Генная инженерия М.М.П. СМсспк
- 16. М.М.П. 5.2 ГЕННАЯ ИНЖЕНЕРИЯ, или технология рекомбинантных ДНК — это изменение с помощью биохимических и генетических
- 17. Генетическая инженерия (генная инженерия) — совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК, выделения
- 18. Под генной инженерией обычно понимают искусственный перенос нужных генов от одного вида живых организмов (бактерий, животных,
- 19. Генная инженерия – это отрасль молекулярной биологии и генетики, целью которой является получение с помощью лабораторных
- 20. Генная инженерия основана на получении гибридных молекул ДНК и введении этих молекул в клетки других организмов,
- 21. Однако эта схема эффективна лишь для двудольных растений. Для однодольных, в основном злаковых растений, разработаны другие
- 22. М.М.П. 5.2 СМсспк
- 23. Чтобы осуществить перенос генов (т.е. трансгенез), необходимо выполнить следующие сложные операции: выделение из клеток бактерий, животных
- 24. М.М.П. 5.2 Общая схема генно-инженерной программы: В качестве донора или реципиента может быть использована любая клетка
- 25. М.М.П. 5.2 Векторы ДНК – это небольшие молекулы ДНК, способные проникать в другие клетки и реплицироваться
- 26. М.М.П. 5.2 В качестве вектора обычно используют бактериальную плазмиду, бактериофаг (вирус бактерий), вирус растений или животных.
- 27. Основные этапы решения генноинженерной задачи следующие: 1. Получение изолированного гена. 2. Введение гена в вектор для
- 28. 5.2 М.М.П. Выделение и идентификация отдельных генов (соответствующих фрагментов ДНК или РНК) Получение генетических конструкций путем
- 29. Процесс синтеза генов в настоящее время разработан очень хорошо и даже в значительной степени автоматизирован. Существуют
- 30. Получила распространение техника, позволяющая использовать для синтеза ДНК, в том числе мутантной, полимеразную цепную реакцию (ПЦР).
- 31. Чтобы встроить ген в вектор, используют ферменты — рестриктазы и лигазы, также являющиеся полезным инструментом генной
- 32. 5.2 М.М.П. Создание рекомбинантной плазмиды СМсспк
- 33. М.М.П. 5.2 СМсспк
- 34. Схема получения гена, кодирующего нужный в производстве белок М.М.П. 5.2 СМсспк
- 35. На первом этапе из клеток выделяют и-РНК. Затем на ней, как на матрице, синтезируют нить комплементарной
- 36. После удаления РНК из этой молекулы на оставшейся одноцепочечной ДНК осуществляют синтез второй нити. В результате
- 37. Используя специальные ферменты, ее встраивают в кольцевую ДНК плазмид (внехромосомных молекул ДНК), которые выполняют роль переносчика
- 38. На последнем этапе плазмиды со вставкой встраиваются в бактериальную хромосому. В ней перенесенный ген человека, животного,
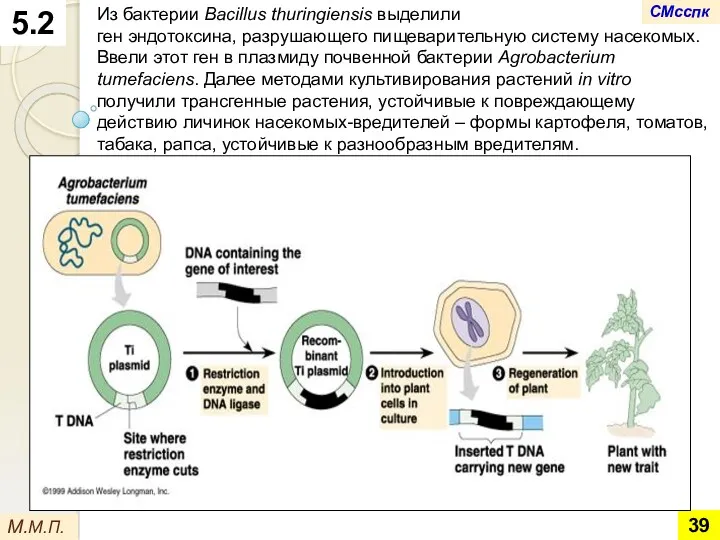
- 39. 5.2 М.М.П. Из бактерии Bacillus thuringiensis выделили ген эндотоксина, разрушающего пищеварительную систему насекомых. Ввели этот ген
- 40. Получение трансгенных растений хлопка с геном bt , несущим устойчивость к насекомым. Ген bt (Bacillus thuringiensis
- 41. М.М.П. 5.2 СМсспк
- 42. 5.2 М.М.П. Во всех рассмотренных примерах хорошо видны основные этапы получения геномодифицированных растений Получение стерильной культуры
- 43. 1. Генная инженерия в корне отличается от выведения новых сортов и пород. Искусственное добавление чужеродных генов
- 44. 3. В результате искусственного добавления чужеродного гена непредвиденно могут образоваться опасные вещества. В худшем случае это
- 45. 6. Созданные до настоящего времени с помощью генной инженерии продукты питания не имеют сколько-нибудь значительной ценности
- 46. 8. Могут возникнуть новые и опасные вирусы. Экспериментально показано, что встроенные в геном гены вирусов могут
- 47. 11. Геномодифицированные растения могут аккумулировать гербициды и пестициды, к которым они устойчивы, что опасно не только
- 48. 5.2 М.М.П. К пищевым рискам относят: Непосредственное действие токсичных и аллергенных трансгенных белков ГМО. Риски, опосредованные
- 49. 5.2 М.М.П. Экологические риски: Снижение генетического разнообразия возделываемых культур вследствие массового применения ГМО, полученных из ограниченного
- 50. 5.2 М.М.П. Агротехнические риски: Риски непредсказуемых изменений нецелевых свойств и признаков модифицированных сортов, связанные с плейотропным
- 52. Скачать презентацию

Содержание лекции:
1. Геномная и хромосомная инженерия
2. Генная инженерия
М.М.П.
5.
СМсспк
Содержание лекции:
1. Геномная и хромосомная инженерия
2. Генная инженерия
М.М.П.
5.
СМсспк

5.1
1. Геномная и хромосомная инженерия
М.М.П.
СМсспк
5.1
1. Геномная и хромосомная инженерия
М.М.П.
СМсспк

Поэтому понятие генетической инженерии объединяет понятия геномной, хромосомной и генной инженерии.
Под
Поэтому понятие генетической инженерии объединяет понятия геномной, хромосомной и генной инженерии.
Под

5.1
Сущность геномной ДНК-технологии
(геномной инженерии) заключается в целенаправленной перестройке генома того или
5.1
Сущность геномной ДНК-технологии (геномной инженерии) заключается в целенаправленной перестройке генома того или
5.1
М.М.П.
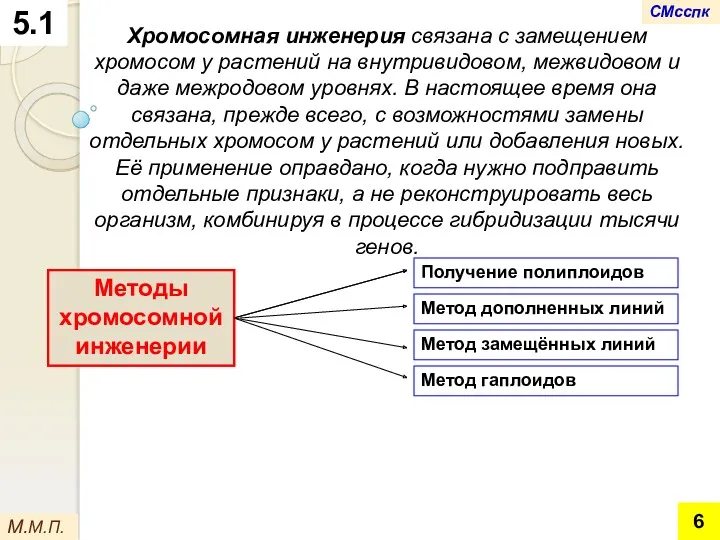
Хромосомная инженерия связана с замещением хромосом у растений на внутривидовом, межвидовом
5.1
М.М.П.
Хромосомная инженерия связана с замещением хромосом у растений на внутривидовом, межвидовом

5.1
Природная геномная и хромосомная инженерия создала полиплоидные комплексы видов растений.
При этом
5.1
Природная геномная и хромосомная инженерия создала полиплоидные комплексы видов растений.
При этом

5.1
Известно, что в клетках каждого диплоидного
организма имеются пары гомологичных хромосом. Такой
5.1
Известно, что в клетках каждого диплоидного организма имеются пары гомологичных хромосом. Такой

Другой методический прием хромосомной инженерии состоит не в замещении в геноме
Другой методический прием хромосомной инженерии состоит не в замещении в геноме

Примером геномной инженерии является
создание нового рода культурных злаков – тритикале.
Получена
Примером геномной инженерии является
создание нового рода культурных злаков – тритикале.
Получена
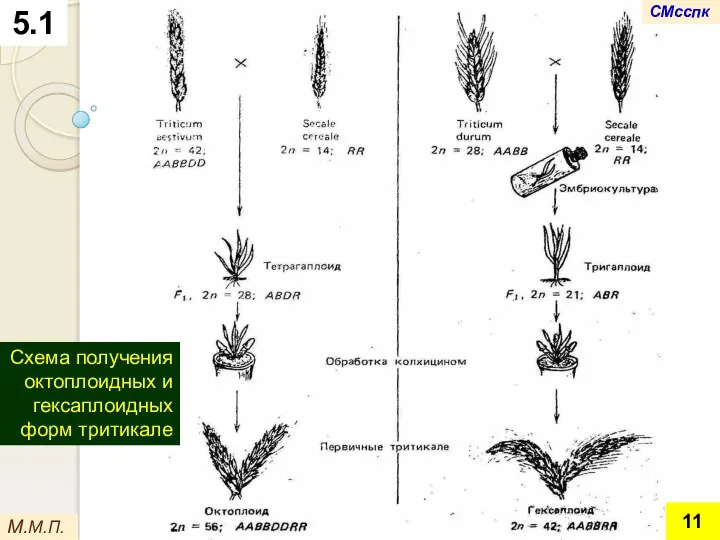
М.М.П.
Схема получения октоплоидных и гексаплоидных форм тритикале
5.1
СМсспк
М.М.П.
Схема получения октоплоидных и гексаплоидных форм тритикале
5.1
СМсспк

В хромосомной инженерии очень перспективен метод гаплоидов, основанный на выращивании гаплоидных
В хромосомной инженерии очень перспективен метод гаплоидов, основанный на выращивании гаплоидных

МЕТОД ПОЛУЧЕНИЯ ДИГАПЛОИДНЫХ ГОМОЗИГОТНЫХ ЛИНИЙ (ВНИИ сахарной свеклы)
Метод позволяет получать гомозиготный
МЕТОД ПОЛУЧЕНИЯ ДИГАПЛОИДНЫХ ГОМОЗИГОТНЫХ ЛИНИЙ (ВНИИ сахарной свеклы)
Метод позволяет получать гомозиготный
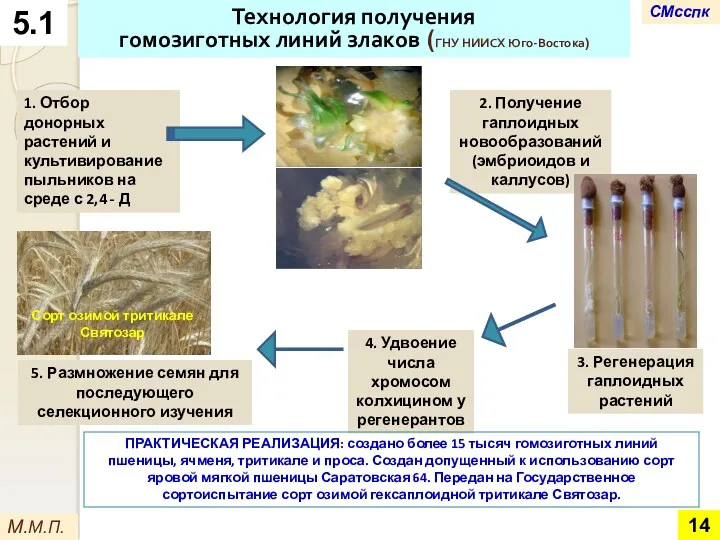
Технология получения
гомозиготных линий злаков (ГНУ НИИСХ Юго-Востока)
1. Отбор донорных растений и
Технология получения
гомозиготных линий злаков (ГНУ НИИСХ Юго-Востока)
1. Отбор донорных растений и

5.2
2. Генная инженерия
М.М.П.
СМсспк
5.2
2. Генная инженерия
М.М.П.
СМсспк

М.М.П.
5.2
ГЕННАЯ ИНЖЕНЕРИЯ, или технология рекомбинантных ДНК — это изменение с помощью
М.М.П.
5.2
ГЕННАЯ ИНЖЕНЕРИЯ, или технология рекомбинантных ДНК — это изменение с помощью

Генетическая инженерия (генная инженерия) — совокупность приёмов, методов и технологий получения рекомбинантных
Генетическая инженерия (генная инженерия) — совокупность приёмов, методов и технологий получения рекомбинантных

Под генной инженерией обычно понимают искусственный перенос нужных генов от одного
Под генной инженерией обычно понимают искусственный перенос нужных генов от одного

Генная инженерия – это отрасль молекулярной биологии и генетики, целью которой
Генная инженерия – это отрасль молекулярной биологии и генетики, целью которой

Генная инженерия основана на получении гибридных молекул ДНК и введении этих
Генная инженерия основана на получении гибридных молекул ДНК и введении этих

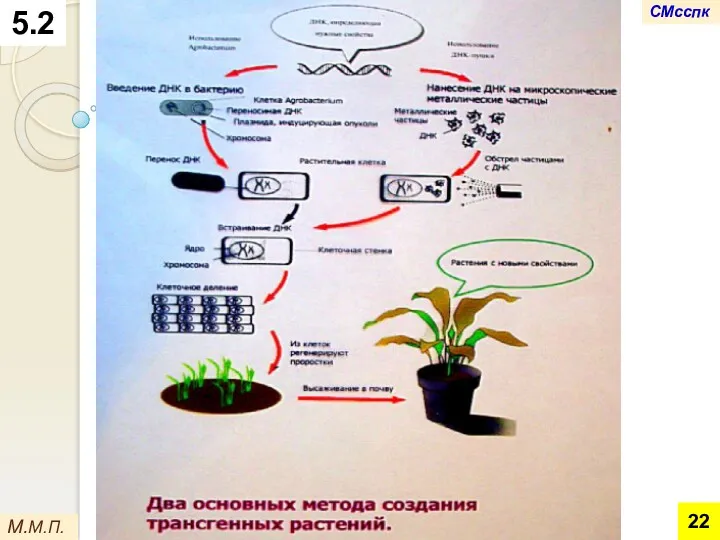
Однако эта схема эффективна лишь для двудольных растений.
Для однодольных, в основном
Однако эта схема эффективна лишь для двудольных растений.
Для однодольных, в основном
М.М.П.
5.2
СМсспк
М.М.П.
5.2
СМсспк

Чтобы осуществить перенос генов (т.е. трансгенез), необходимо выполнить следующие сложные операции:
выделение
Чтобы осуществить перенос генов (т.е. трансгенез), необходимо выполнить следующие сложные операции:
выделение
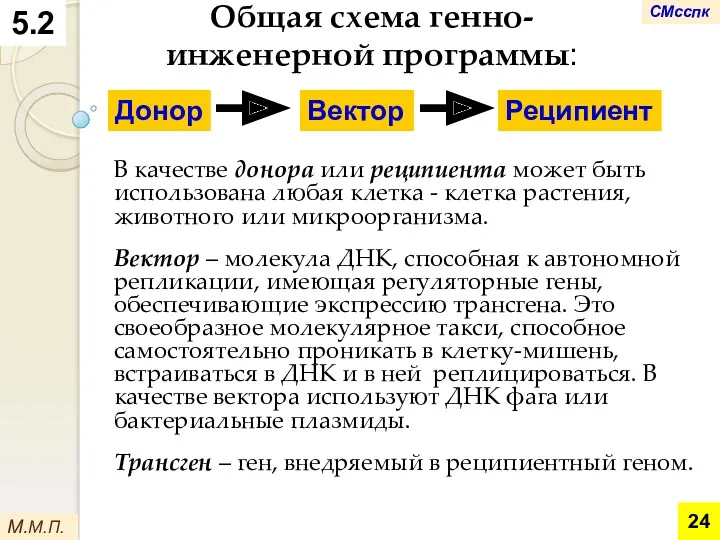
М.М.П.
5.2
Общая схема генно-инженерной программы:
В качестве донора или реципиента может быть использована
М.М.П.
5.2
Общая схема генно-инженерной программы:
В качестве донора или реципиента может быть использована

М.М.П.
5.2
Векторы ДНК – это небольшие молекулы ДНК, способные проникать в другие
М.М.П.
5.2
Векторы ДНК – это небольшие молекулы ДНК, способные проникать в другие

М.М.П.
5.2
В качестве вектора обычно используют
бактериальную плазмиду, бактериофаг (вирус бактерий),
вирус растений или
М.М.П.
5.2
В качестве вектора обычно используют
бактериальную плазмиду, бактериофаг (вирус бактерий),
вирус растений или
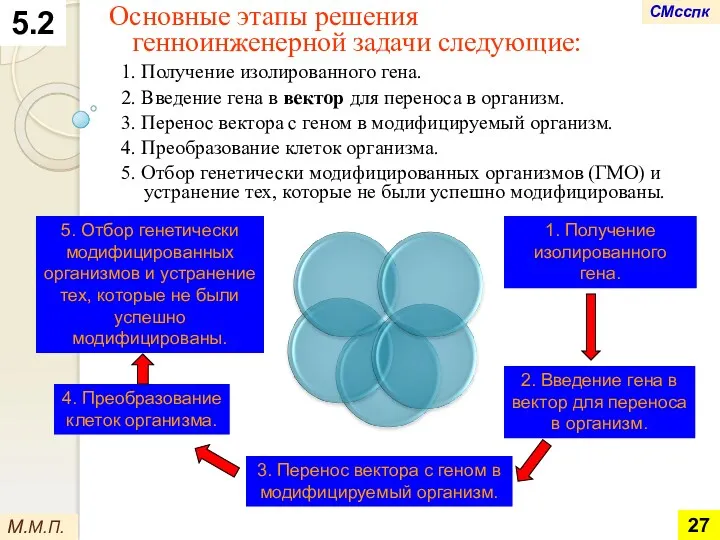
Основные этапы решения
генноинженерной задачи следующие:
1. Получение изолированного гена.
2. Введение гена
Основные этапы решения
генноинженерной задачи следующие:
1. Получение изолированного гена.
2. Введение гена

5.2
М.М.П.
Выделение и идентификация отдельных генов (соответствующих фрагментов ДНК или РНК)
Получение генетических
5.2
М.М.П.
Выделение и идентификация отдельных генов (соответствующих фрагментов ДНК или РНК)
Получение генетических

Процесс синтеза генов в настоящее время разработан очень хорошо и даже
Процесс синтеза генов в настоящее время разработан очень хорошо и даже

Получила распространение техника, позволяющая использовать для синтеза ДНК, в том числе
Получила распространение техника, позволяющая использовать для синтеза ДНК, в том числе

Чтобы встроить ген в вектор, используют ферменты — рестриктазы и лигазы, также
Чтобы встроить ген в вектор, используют ферменты — рестриктазы и лигазы, также
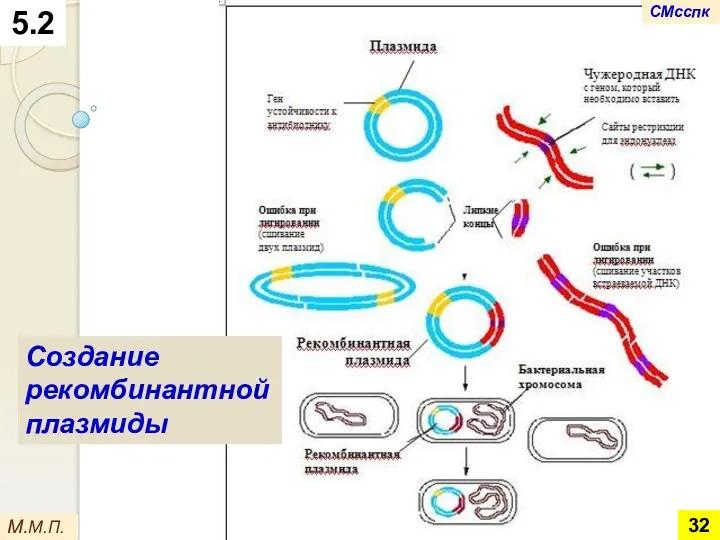
5.2
М.М.П.
Создание рекомбинантной плазмиды
СМсспк
5.2
М.М.П.
Создание рекомбинантной плазмиды
СМсспк

М.М.П.
5.2
СМсспк
М.М.П.
5.2
СМсспк
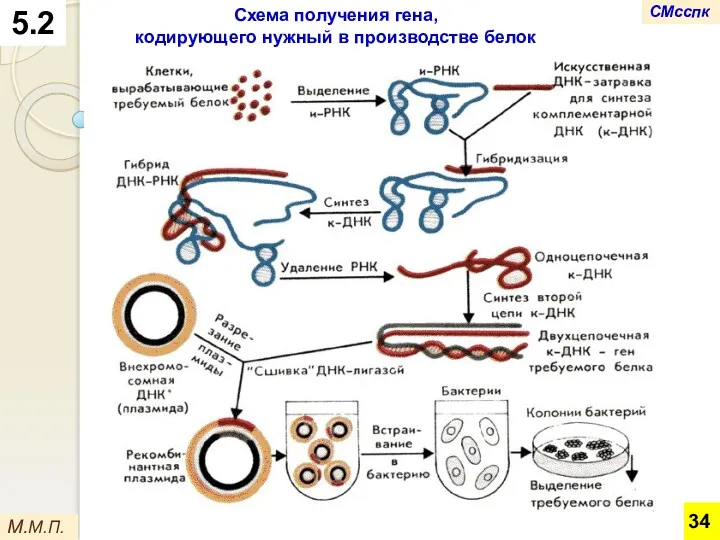
Схема получения гена,
кодирующего нужный в производстве белок
М.М.П.
5.2
СМсспк
Схема получения гена,
кодирующего нужный в производстве белок
М.М.П.
5.2
СМсспк
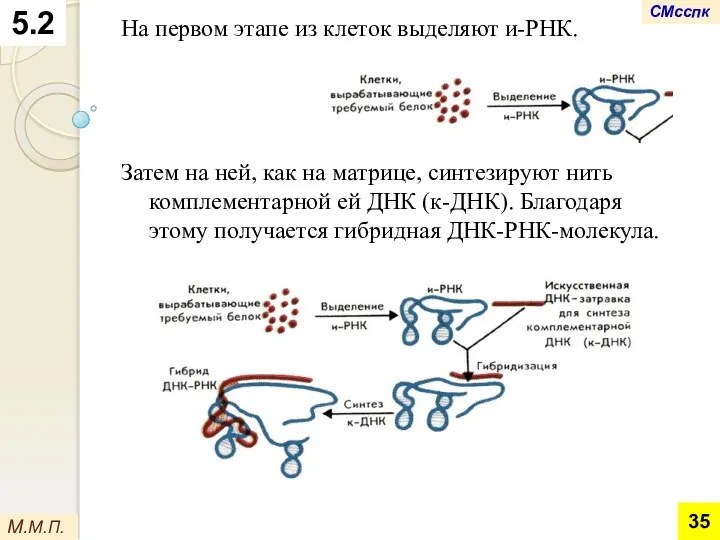
На первом этапе из клеток выделяют и-РНК.
Затем на ней, как на
На первом этапе из клеток выделяют и-РНК.
Затем на ней, как на
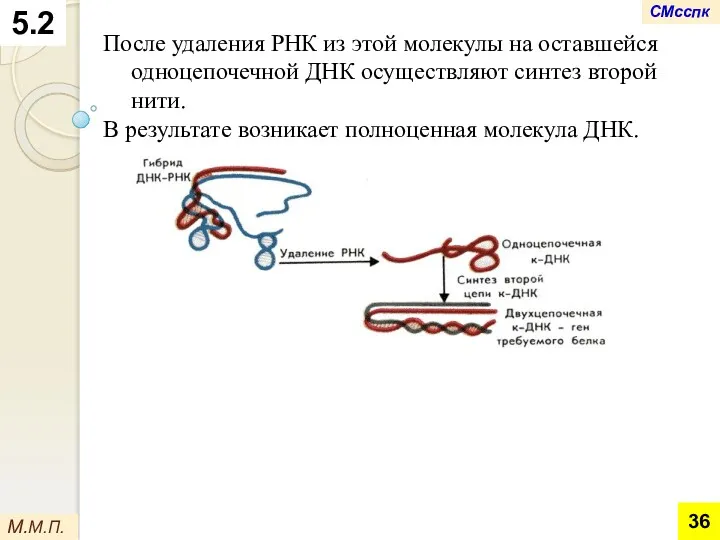
После удаления РНК из этой молекулы на оставшейся одноцепочечной ДНК осуществляют
После удаления РНК из этой молекулы на оставшейся одноцепочечной ДНК осуществляют
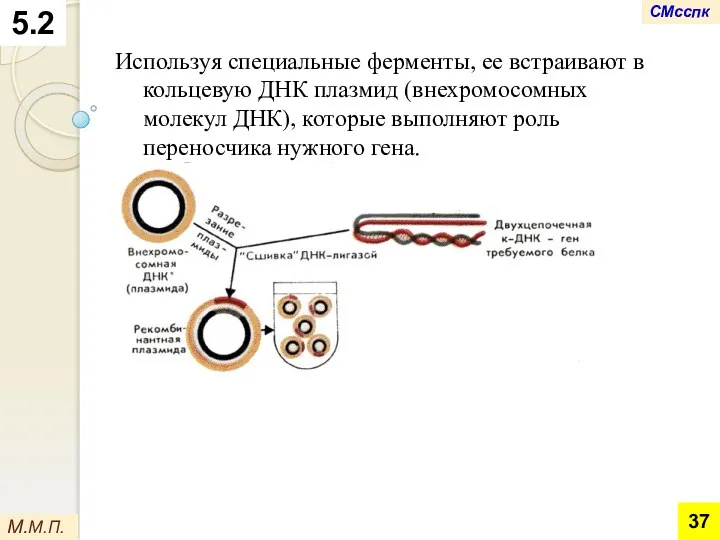
Используя специальные ферменты, ее встраивают в кольцевую ДНК плазмид (внехромосомных молекул
Используя специальные ферменты, ее встраивают в кольцевую ДНК плазмид (внехромосомных молекул
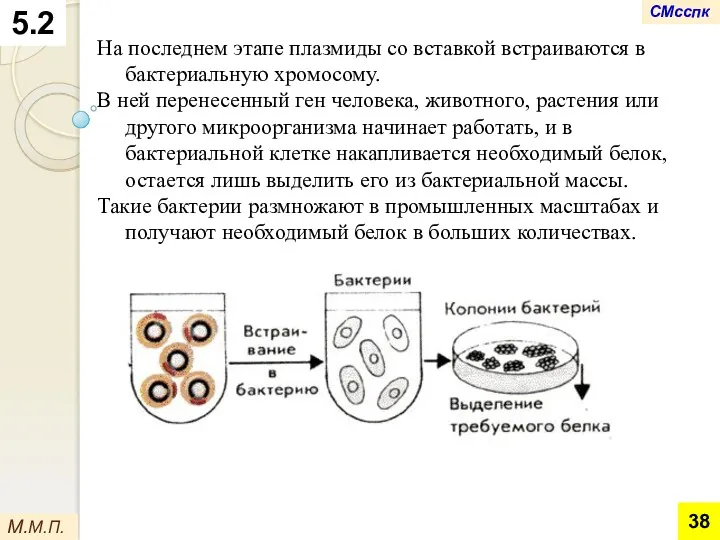
На последнем этапе плазмиды со вставкой встраиваются в бактериальную хромосому.
В ней
На последнем этапе плазмиды со вставкой встраиваются в бактериальную хромосому.
В ней
5.2
М.М.П.
Из бактерии Bacillus thuringiensis выделили
ген эндотоксина, разрушающего пищеварительную систему насекомых. Ввели
5.2
М.М.П.
Из бактерии Bacillus thuringiensis выделили ген эндотоксина, разрушающего пищеварительную систему насекомых. Ввели
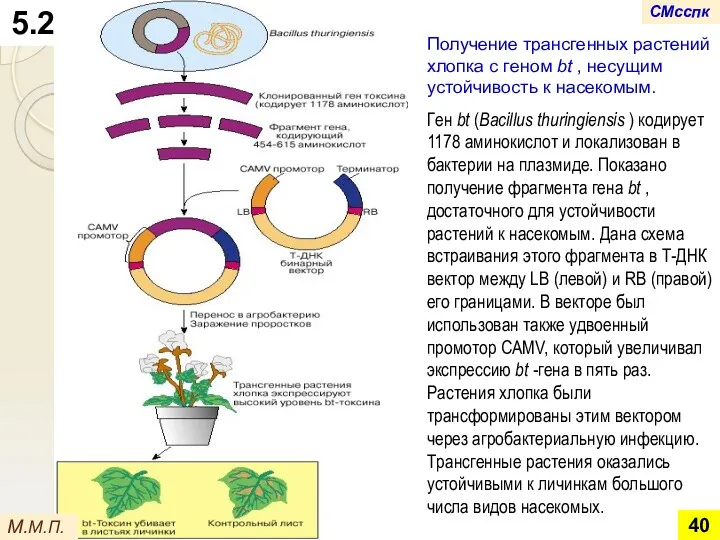
Получение трансгенных растений хлопка с геном bt , несущим устойчивость к
Получение трансгенных растений хлопка с геном bt , несущим устойчивость к
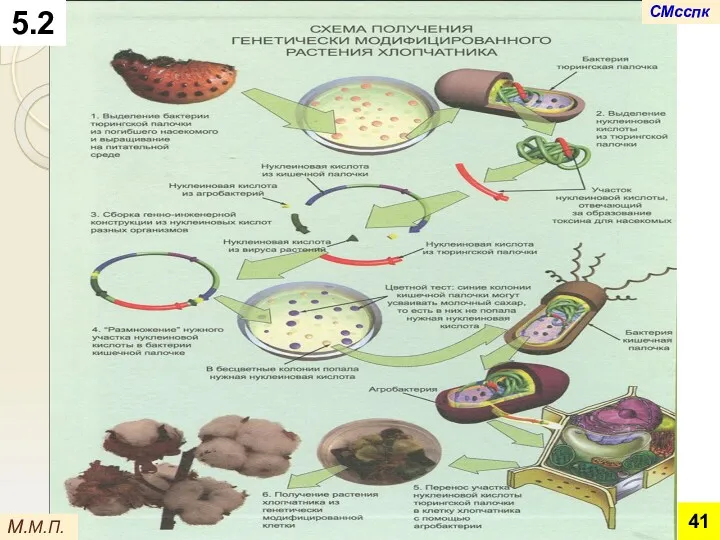
М.М.П.
5.2
СМсспк
М.М.П.
5.2
СМсспк

5.2
М.М.П.
Во всех рассмотренных примерах
хорошо видны основные этапы получения геномодифицированных растений
Получение стерильной
5.2
М.М.П.
Во всех рассмотренных примерах
хорошо видны основные этапы получения геномодифицированных растений
Получение стерильной

1. Генная инженерия в корне отличается от выведения новых сортов и
1. Генная инженерия в корне отличается от выведения новых сортов и

3. В результате искусственного добавления чужеродного гена непредвиденно могут образоваться опасные
3. В результате искусственного добавления чужеродного гена непредвиденно могут образоваться опасные

6. Созданные до настоящего времени с помощью генной инженерии продукты питания
6. Созданные до настоящего времени с помощью генной инженерии продукты питания

8. Могут возникнуть новые и опасные вирусы. Экспериментально показано, что встроенные
8. Могут возникнуть новые и опасные вирусы. Экспериментально показано, что встроенные

11. Геномодифицированные растения могут аккумулировать гербициды и пестициды, к которым они
11. Геномодифицированные растения могут аккумулировать гербициды и пестициды, к которым они

5.2
М.М.П.
К пищевым рискам относят:
Непосредственное действие токсичных и аллергенных трансгенных белков ГМО.
5.2
М.М.П.
К пищевым рискам относят:
Непосредственное действие токсичных и аллергенных трансгенных белков ГМО.

5.2
М.М.П.
Экологические риски:
Снижение генетического разнообразия возделываемых культур вследствие массового применения ГМО, полученных
5.2
М.М.П.
Экологические риски:
Снижение генетического разнообразия возделываемых культур вследствие массового применения ГМО, полученных

5.2
М.М.П.
Агротехнические риски:
Риски непредсказуемых изменений нецелевых свойств и признаков модифицированных сортов, связанные
5.2
М.М.П.
Агротехнические риски:
Риски непредсказуемых изменений нецелевых свойств и признаков модифицированных сортов, связанные
 Бактериальные болезни растений. (Лекция 7)
Бактериальные болезни растений. (Лекция 7) Методы психогенетики
Методы психогенетики Клеточный цикл. Деление и рост растительной клетки
Клеточный цикл. Деление и рост растительной клетки Вирусы (2). Лекция 10
Вирусы (2). Лекция 10 Взаимодействие генов
Взаимодействие генов Соматический и автономный (вегетативный) отделы нервной системы
Соматический и автономный (вегетативный) отделы нервной системы Дикие животные леса. Для дошкольников. (Часть 1)
Дикие животные леса. Для дошкольников. (Часть 1) Вітаміни. Їх роль у життєдіяльності людини
Вітаміни. Їх роль у життєдіяльності людини Сүйектің байланысу түрлері
Сүйектің байланысу түрлері The muscular system
The muscular system Сүйекті жемістер мен жидектерді сақтау және сапасын сараптау
Сүйекті жемістер мен жидектерді сақтау және сапасын сараптау Презентация “В здоровом теле – здоровый дух “
Презентация “В здоровом теле – здоровый дух “ Красная книга Самарской области
Красная книга Самарской области Организация проектирования ботанических садов
Организация проектирования ботанических садов Ретинол
Ретинол Соцветия. Биологическое значение соцветий. Классификация соцветий. Происхождение соцветия
Соцветия. Биологическое значение соцветий. Классификация соцветий. Происхождение соцветия Трюфель – самый вкусный и самый дорогой гриб в мире
Трюфель – самый вкусный и самый дорогой гриб в мире Транскрипция. Биосинтез белка
Транскрипция. Биосинтез белка Физиология дыхания
Физиология дыхания Презентация Бактерии 5 класс
Презентация Бактерии 5 класс Углеводы. Классификация углеводов
Углеводы. Классификация углеводов Блок 3. Птицы
Блок 3. Птицы Бурые и зеленые водоросли
Бурые и зеленые водоросли Семейство розоцветные
Семейство розоцветные Генетика. Решение задач
Генетика. Решение задач Перелітні та осілі птахи Уманщини
Перелітні та осілі птахи Уманщини Умеют ли животные общаться
Умеют ли животные общаться блок_4_Зоо_Простейшие
блок_4_Зоо_Простейшие