- Генетика бактерий

Содержание
- 2. ОРГАНИЗАЦИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА У БАКТЕРИЙ ДНК = нуклеоид (бактериальная «хромосома») – кодирует жизненно важные признаки внехромосомные
- 3. Единицей наследственности является ГЕН = участок ДНК, в котором зашифрована последовательность аминокислот в полипептидной цепочке, контролирующая
- 4. Гены: Структурные = обуславливают синтез определенного белка (фермента), при мутации образуется белок измененного состава, Ген-регулятор =
- 5. Совокупность генов, сосредоточенных в нуклеоиде («Хромосоме») бактерий называется генотип. Фенотип – совокупность всех признаков микроорганизма, сформировавшаяся
- 6. ДНК
- 7. ДНК («хромосома») двухцепочечная кольцевая молекула, сод-т до 5 тыс. генов, имеет молекулярную массу 1,7-2,8х109 дальтон, включает
- 8. Генетическая карта = это схематическое изображение всех генов микроорганизма. Гены, отвечающие за определенный признак, обозначают строчными
- 9. ВНЕХРОМОСОМНЫЕ ФАКТОРЫ НАСЛЕДСТВЕННОСТИ автономные – являются репликоном плазмиды неавтономные - реплицируются только в составе репликона (нуклеоида
- 10. ВСТРАИВАНИЕ В НУКЛЕОИД ВНЕХРОМОСОМНЫХ ФАКТОРОВ НАСЛЕДСТВЕННОСТИ в гомологичных участках Плазмиды, Умеренные фаги. в любых участках Транспозоны,
- 11. ПЛАЗМИДЫ внехромосомные факторы наследственности у бактерий, двухцепочечные молекулы ДНК, несут 40-50 генов, не являются жизненно важными
- 12. ПЛАЗМИДЫ функции регуляторная – компенсирует нарушение функции ДНК нуклеоида, кодирующая – вносит в генотип новую информацию.
- 13. ПЛАЗМИДЫ контроль репликации плазмид со стороны нуклеоида строгий (делятся синхронно с нуклеоидом) ⇒ 1-2 копии на
- 14. ФУНКЦИИ TRA-ОПЕРОНА детерминирует образование конъюгативных пилей, мобилизирует на перенос: саму конъюгативную плазмиду (F+), другую, неконъюгативную, плазмиду
- 15. Фенотипические признаки, сообщаемые бактерии плазмидами устойчивость к антибиотикам, образование бактериоцинов, продукция факторов патогенности, способность к синтезу
- 16. Наиболее изучены плазмиды: F- плазмида = половой фактор – контролирует синтез половых ворсинок, = бывает: -
- 17. Наиболее изучены плазмиды: Col-плазмиды - обусловливают синтез бактериоцинов ( = белки, задерживающие рост других штаммов бактерий
- 18. ТРАНСПОЗОНЫ определение = нуклеотидные последовательности (от 2 000 до 20000 пар нуклеотидов), способные менять место своей
- 19. ТРАНСПОЗОНЫ Состав: особые концевые структуры, которые отличают транспозон от др. фрагментов ДНК (маркеры транспозона), гены транспозиции,
- 20. IS-ПОСЛЕДОВАТЕЛЬНОСТИ определение =вставки нуклеотидных последовательностей (порядка 1 000 пар нуклеотидов), содержат только гены, необходимые для собственного
- 21. IS-ПОСЛЕДОВАТЕЛЬНОСТИ отличия от транспозонов содержат только гены транспозиции, не обнаружены в свободном состоянии.
- 22. Функции IS-ПОСЛЕДОВАТЕЛЬНОСТЕЙ координация взаимодействия внехромосомных факторов наследственности между собой и с бактериальной хромосомой для обеспечения их
- 23. Изменчивость микроорганизмов Модификационная = ненаследуемая, Генотипическая = наследуемая.
- 24. Изменчивость микроорганизмов Модификационная = ненаследуемая, фенотипическая, адаптационная, – возникает как приспособительная реакция организма на условия среды,
- 25. МОДИФИКАЦИИ У БАКТЕРИЙ Фенотипические изменения у бактерий не сопровождаются изменениями первичной структуры ДНК, они выражаются: -
- 26. Изменчивость микроорганизмов Наследуемая = генотипическая – изменения затрагивают лишь отдельные клетки, – приобретенные признаки передаются потомству
- 27. МУТАЦИИ У БАКТЕРИЙ Определение Изменения в первичной структуре ДНК, которые выражаются в наследственно закреплённой утрате или
- 28. Классификация мутаций по происхождению спонтанные – трудно или невозможно связать с действием определённого фактора (мутагена) ошибки
- 29. Классификация мутаций по количеству мутировавших генов: Генные затрагивают один ген: - замена одной пары азотистых оснований
- 30. Хромосомные мутации делеции – потеря гена, инверсия – поворот участка хромосомы или нарушение порядка гена, дупликации
- 31. Классификация мутаций по направленности прямые – потеря или изменение признака, обратные (реверсии) – восстановление признака: истинные
- 32. SR-ДИССОЦИАЦИИ = появление в чистой культуре 2 видов бактериальных клеток, которые отличаются по характеру образуемых колоний
- 33. SR-ДИССОЦИАЦИИ механизм Это инсерционная мутация, приводящая к утрате генов, контролирующих синтез полисахаридных звеньев ЛПС наружной мембраны
- 34. МУТАГЕНЫ Мутагены – факторы, вызывающие мутации. Различают: физические мутагены – ультрафиолетовые лучи, ионизирующие излучения, магнитные поля,
- 35. МУТАГЕНЫ Классификация по механизму действия: аналоги азотистых оснований ⇨ замена пар оснований, акридиновые красители ⇨ выпадения
- 36. РЕПАРАЦИИ Определение Процесс восстановления повреждённой ДНК ферментами репарационных систем Различают 2 типа репарационных систем: Система фотореактивации
- 37. Система фотореактивации УФ-лучи ⇩ мутация тиминовые димеры ⇩ видимый свет ⇩ активация фермента репарация ⇩ расщепление
- 38. Этапы темновой репарации: установление места повреждения ДНК = эндонуклеаза, «вырезание» поврежденного фрагмента = полимераза 1, синтез
- 39. Система темновой репарации УФ-лучи ⇩ тиминовые димеры ⇩ темнота ⇩
- 40. Система темновой репарации обнаружение и нарезание повреждённого участка (эндонуклеаза) ⇩
- 41. Система темновой репарации удаление повреждённого участка (ДНК-полимераза I) ⇩
- 42. Система темновой репарации синтез на матрице второй нити ДНК нового, не содержащего мутации, участка (ДНК-полимераза I
- 43. Система темновой репарации «вшивание» нового участка в цепь ДНК (лигаза)
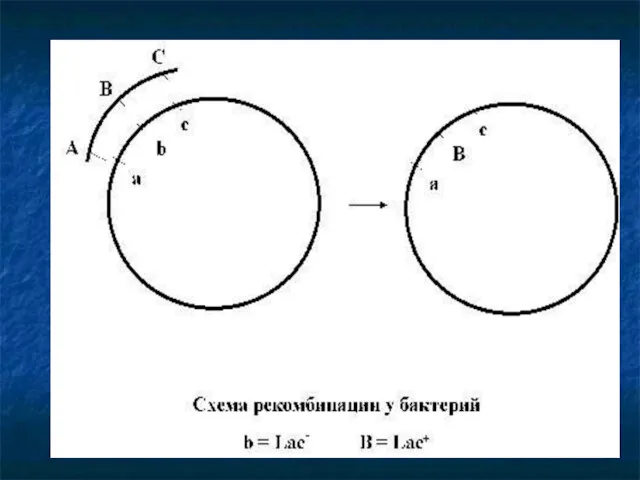
- 44. Генетические рекомбинации = перераспределение генетического материала родителей в потомстве, обусловливающее комбинативную изменчивость организмов, = взаимодействие между
- 45. Механизм рекомбинаций клетки=доноры ⇩ передают информацию ⇩ клеткам-реципиентам ⇩ рекомбинат генотип рекомбинанта = генотип реципиента+ часть
- 47. ВИДЫ РЕКОМБИНАТИВНОЙ ИЗМЕНЧИВОСТИ У БАКТЕРИЙ Трансформация – непосредственная передача генетического материала от донорской к реципиентной клетке
- 48. Трансформация = способ передачи генетической информации путем внедрения свободной ДНК донора в бактерию-реципиент Трансформация эффективно происходит
- 49. Трансформация Клетки, способные принимать донорскую ДНК, называются компетентными. Состояние компетентности возникает в период роста клетки и
- 50. Процесс трансформации состоит из фаз: адсорбция ДНК донора на клетке-реципиенте, проникновение ДНК внутрь клетки-реципиента с последующей
- 51. Схема трансформации
- 52. Конъюгация – перенос генетического материала из клетки-донора в клетку реципиента при тесном контакте. Донорами генетического материала
- 53. Конъюгация 2 вида конъюгации: Если F-плазмида автономна→ бактерия наз-ся F+ штаммом 2.Если F-плазмида интегрирована в ДНК
- 54. 1 Если F-плазмида автономна: 1. Прикрепление клетки донора к реципиенту с помощью половых ворсинок. 2. Между
- 55. Схема конъюгации у бактерий (если F-плазмида автономна)
- 56. 1 Если F-плазмида автономна: 2.3. белок способствует замыканию линейной нити в клетке-реципиенте, 2.4. одноцепочечные нити достраиваются
- 57. Схема конъюгации у бактерий (если F-плазмида автономна)
- 58. 2.Если F-плазмида встроена в хромосому бактерии = Hfr-штамм: Происходит разрыв одной нити ДНК при участии эндонуклеазы
- 59. Образование Hfr-штамма
- 60. Схема конъюгации Hfr-штамма разрыв одной нити ДНК при участии эндонуклеазы в точке О, расположенной в месте
- 61. Схема конъюгации Hfr-штамма Проксимальный конец ДНК через конъюгационный мостик проникает в клетку-реципиент и сразу же достраивается
- 62. Схема конъюгации Hfr-штамма Двунитевой фрагмент ДНК встраивается в геном клетки-реципиента; Плазмида осталась в клетке-доноре (Hfr-штамм)
- 63. Трансдукция – передача генетического материала от одной бактерии к другой при помощи фагов. Различают: 1) общую
- 64. Общая = неспецифическая трансдукция – когда в клетку–реципиент вместе с фаговой ДНК переносится любой ген донора.
- 65. Специфическая трансдукция – фаг переносит специфические гены от бактерии-донора к бактерии-реципиенту: При выходе из ДНК лизогенной
- 66. Абортивная трансдукция = принесенный фагом фрагмент ДНК бактерии-донора не включается в хромосому бактерии-реципиента, а располагается в
- 67. Генетическая рекомбинация = обмен между гомологичными участками геномов двух вирусов, – чаще встречается у ДНК-содержащих вирусов,
- 68. Генетическая реактивация = обмен между геномами родственных вирусов, у которых мутации произошли в разных генах →
- 69. Комплементация = обмен, когда один из двух вирусов в результате мутации синтезирует неполноценный белок. Немутантный вирус
- 70. Фенотипическое смешивание при смешанном заражении двумя вирусами часть потомства приобретает фенотипические признаки, присущие обоим вирусам при
- 71. Фенотипическое смешивание вирус 1 + вирус 2 ⇒ в одной клетке вирус 1 вирус 2 НК
- 72. ГЕННАЯ ИНЖЕНЕРИЯ В МЕДИЦИНСКОЙ МИКРОБИОЛОГИИ Продукты, получаемые генно-инженерным способом с помощью рекомбинантных штаммов бактерий вакцины гормоны
- 73. ГЕННАЯ ИНЖЕНЕРИЯ В МЕДИЦИНСКОЙ МИКРОБИОЛОГИИ Получение рекомбинантной вакцины для профилактики гепатита В встраивание гена вируса гепатита
- 74. ГЕНЕТИЧЕСКИЕ МЕТОДЫ, ПРИМЕНЯЕМЫЕ В МИКРОБИОЛОГИЧЕСКОЙ ДИАГНОСТИКЕ процентное содержание Г+Ц в бактериальном геноме метод молекулярной гибридизации полимеразная
- 75. МЕТОД МОЛЕКУЛЯРНОЙ ГИБРИДИЗАЦИИ Цель Выявления степени сходства различных ДНК (при идентификации микроорганизмов – сравнение ДНК выделенного
- 76. МЕТОД МОЛЕКУЛЯРНОЙ ГИБРИДИЗАЦИИ Принцип осуществления исследуемая ДНК ⇩ нагрев в щелочной среде ⇩ расплетение на две
- 77. ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ Цели обнаружение в патологическом материале конкретного вида микроорганизма без выделения чистой культуры идентификация
- 78. ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ Принцип осуществления патологический материал или штамм микроорганизма ⇩ выделение ДНК ⇩ нагрев ⇩
- 79. ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ Принцип осуществления ⇩ добавление ДНК-полимеразы и нуклеотидов ⇩ нуклеотиды присоединяются к 3’-концам праймеров
- 80. При нагревании две комплементарные нити ДНК расходятся – она плавится
- 81. ПЦР
- 82. Рестрикционный анализ Расщепление ДНК микроорганизмов на фрагменты при помощи рестриктаз (эндонуклеаз), От бактерий выделено 175 рестриктаз,
- 84. Скачать презентацию

ОРГАНИЗАЦИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА У БАКТЕРИЙ
ДНК = нуклеоид (бактериальная «хромосома») – кодирует
ОРГАНИЗАЦИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА У БАКТЕРИЙ
ДНК = нуклеоид (бактериальная «хромосома») – кодирует

Единицей наследственности является ГЕН = участок ДНК, в котором зашифрована последовательность

Гены:
Структурные = обуславливают синтез определенного белка (фермента), при мутации образуется белок
Гены:
Структурные = обуславливают синтез определенного белка (фермента), при мутации образуется белок

Совокупность генов, сосредоточенных в нуклеоиде («Хромосоме») бактерий называется генотип.
Фенотип – совокупность
Совокупность генов, сосредоточенных в нуклеоиде («Хромосоме») бактерий называется генотип.
Фенотип – совокупность
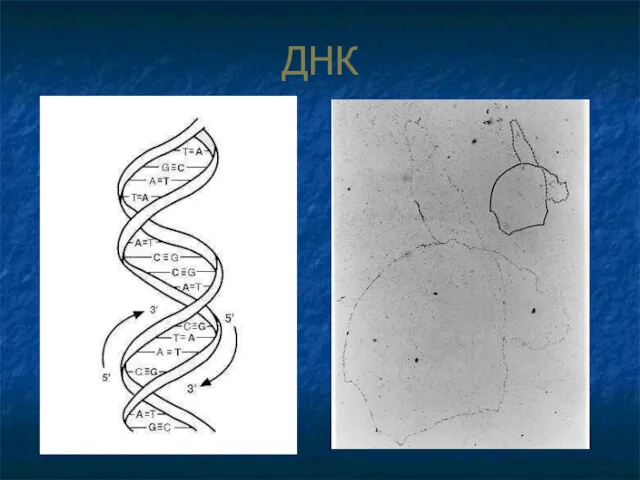
ДНК
ДНК

ДНК («хромосома»)
двухцепочечная кольцевая молекула,
сод-т до 5 тыс. генов,
имеет
ДНК («хромосома»)
двухцепочечная кольцевая молекула,
сод-т до 5 тыс. генов,
имеет

Генетическая карта
= это схематическое изображение всех генов микроорганизма.
Гены, отвечающие за определенный
Генетическая карта
= это схематическое изображение всех генов микроорганизма.
Гены, отвечающие за определенный

ВНЕХРОМОСОМНЫЕ ФАКТОРЫ НАСЛЕДСТВЕННОСТИ
автономные – являются репликоном
плазмиды
неавтономные - реплицируются только в составе
ВНЕХРОМОСОМНЫЕ ФАКТОРЫ НАСЛЕДСТВЕННОСТИ
автономные – являются репликоном
плазмиды
неавтономные - реплицируются только в составе

ВСТРАИВАНИЕ В НУКЛЕОИД ВНЕХРОМОСОМНЫХ ФАКТОРОВ НАСЛЕДСТВЕННОСТИ
в гомологичных участках
Плазмиды,
Умеренные фаги.
в любых участках
Транспозоны,
IS-последовательности.
ВСТРАИВАНИЕ В НУКЛЕОИД ВНЕХРОМОСОМНЫХ ФАКТОРОВ НАСЛЕДСТВЕННОСТИ
в гомологичных участках
Плазмиды,
Умеренные фаги.
в любых участках
Транспозоны,
IS-последовательности.

ПЛАЗМИДЫ
внехромосомные факторы наследственности у бактерий,
двухцепочечные молекулы ДНК,
несут 40-50 генов,
не
ПЛАЗМИДЫ
внехромосомные факторы наследственности у бактерий,
двухцепочечные молекулы ДНК,
несут 40-50 генов,
не

ПЛАЗМИДЫ
функции
регуляторная – компенсирует нарушение функции ДНК нуклеоида,
кодирующая – вносит в генотип
ПЛАЗМИДЫ
функции
регуляторная – компенсирует нарушение функции ДНК нуклеоида,
кодирующая – вносит в генотип

ПЛАЗМИДЫ
контроль репликации плазмид со стороны нуклеоида
строгий (делятся синхронно с нуклеоидом) ⇒
ПЛАЗМИДЫ
контроль репликации плазмид со стороны нуклеоида
строгий (делятся синхронно с нуклеоидом) ⇒

ФУНКЦИИ TRA-ОПЕРОНА
детерминирует образование конъюгативных пилей,
мобилизирует на перенос:
саму конъюгативную плазмиду (F+),
другую, неконъюгативную,
ФУНКЦИИ TRA-ОПЕРОНА
детерминирует образование конъюгативных пилей,
мобилизирует на перенос:
саму конъюгативную плазмиду (F+),
другую, неконъюгативную,

Фенотипические признаки, сообщаемые бактерии плазмидами
устойчивость к антибиотикам,
образование бактериоцинов,
продукция факторов патогенности,
способность
Фенотипические признаки, сообщаемые бактерии плазмидами
устойчивость к антибиотикам,
образование бактериоцинов,
продукция факторов патогенности,
способность

Наиболее изучены плазмиды:
F- плазмида = половой фактор – контролирует синтез половых
Наиболее изучены плазмиды:
F- плазмида = половой фактор – контролирует синтез половых

Наиболее изучены плазмиды:
Col-плазмиды - обусловливают синтез бактериоцинов ( = белки, задерживающие
Наиболее изучены плазмиды:
Col-плазмиды - обусловливают синтез бактериоцинов ( = белки, задерживающие

ТРАНСПОЗОНЫ
определение
= нуклеотидные последовательности (от 2 000 до 20000 пар нуклеотидов), способные
ТРАНСПОЗОНЫ
определение
= нуклеотидные последовательности (от 2 000 до 20000 пар нуклеотидов), способные

ТРАНСПОЗОНЫ
Состав:
особые концевые структуры, которые отличают транспозон от др. фрагментов ДНК (маркеры
ТРАНСПОЗОНЫ
Состав:
особые концевые структуры, которые отличают транспозон от др. фрагментов ДНК (маркеры

IS-ПОСЛЕДОВАТЕЛЬНОСТИ
определение
=вставки нуклеотидных последовательностей (порядка 1 000 пар нуклеотидов),
содержат только гены, необходимые
IS-ПОСЛЕДОВАТЕЛЬНОСТИ
определение
=вставки нуклеотидных последовательностей (порядка 1 000 пар нуклеотидов),
содержат только гены, необходимые

IS-ПОСЛЕДОВАТЕЛЬНОСТИ
отличия от транспозонов
содержат только гены транспозиции,
не обнаружены в свободном состоянии.
IS-ПОСЛЕДОВАТЕЛЬНОСТИ
отличия от транспозонов
содержат только гены транспозиции,
не обнаружены в свободном состоянии.

Функции IS-ПОСЛЕДОВАТЕЛЬНОСТЕЙ
координация взаимодействия внехромосомных факторов наследственности между собой и с
Функции IS-ПОСЛЕДОВАТЕЛЬНОСТЕЙ
координация взаимодействия внехромосомных факторов наследственности между собой и с

Изменчивость микроорганизмов
Модификационная = ненаследуемая,
Генотипическая = наследуемая.
Изменчивость микроорганизмов
Модификационная = ненаследуемая,
Генотипическая = наследуемая.

Изменчивость микроорганизмов
Модификационная = ненаследуемая, фенотипическая, адаптационная,
– возникает как приспособительная реакция
Изменчивость микроорганизмов
Модификационная = ненаследуемая, фенотипическая, адаптационная,
– возникает как приспособительная реакция

МОДИФИКАЦИИ У БАКТЕРИЙ
Фенотипические изменения у бактерий
не сопровождаются изменениями первичной структуры ДНК,
они
МОДИФИКАЦИИ У БАКТЕРИЙ
Фенотипические изменения у бактерий
не сопровождаются изменениями первичной структуры ДНК,
они

Изменчивость микроорганизмов
Наследуемая = генотипическая
– изменения затрагивают лишь отдельные клетки,
Изменчивость микроорганизмов
Наследуемая = генотипическая
– изменения затрагивают лишь отдельные клетки,

МУТАЦИИ У БАКТЕРИЙ
Определение
Изменения в первичной структуре ДНК, которые выражаются в
МУТАЦИИ У БАКТЕРИЙ
Определение
Изменения в первичной структуре ДНК, которые выражаются в

Классификация мутаций по происхождению
спонтанные – трудно или невозможно связать с действием
Классификация мутаций по происхождению
спонтанные – трудно или невозможно связать с действием

Классификация мутаций по количеству мутировавших генов:
Генные затрагивают один ген:
- замена
Классификация мутаций по количеству мутировавших генов:
Генные затрагивают один ген:
- замена

Хромосомные мутации
делеции – потеря гена,
инверсия – поворот участка хромосомы или
Хромосомные мутации
делеции – потеря гена,
инверсия – поворот участка хромосомы или

Классификация мутаций по направленности
прямые – потеря или изменение признака,
обратные (реверсии) –
Классификация мутаций по направленности
прямые – потеря или изменение признака,
обратные (реверсии) –

SR-ДИССОЦИАЦИИ
= появление в чистой культуре 2 видов бактериальных клеток, которые отличаются
SR-ДИССОЦИАЦИИ
= появление в чистой культуре 2 видов бактериальных клеток, которые отличаются

SR-ДИССОЦИАЦИИ
механизм
Это инсерционная мутация, приводящая к утрате генов, контролирующих синтез полисахаридных звеньев
SR-ДИССОЦИАЦИИ
механизм
Это инсерционная мутация, приводящая к утрате генов, контролирующих синтез полисахаридных звеньев

МУТАГЕНЫ
Мутагены – факторы, вызывающие мутации.
Различают:
физические мутагены – ультрафиолетовые лучи, ионизирующие
МУТАГЕНЫ
Мутагены – факторы, вызывающие мутации.
Различают:
физические мутагены – ультрафиолетовые лучи, ионизирующие

МУТАГЕНЫ
Классификация по механизму действия:
аналоги азотистых оснований ⇨ замена пар оснований,
акридиновые красители
МУТАГЕНЫ
Классификация по механизму действия:
аналоги азотистых оснований ⇨ замена пар оснований,
акридиновые красители

РЕПАРАЦИИ
Определение
Процесс восстановления повреждённой ДНК ферментами репарационных систем
Различают 2 типа репарационных систем:
Система
РЕПАРАЦИИ
Определение
Процесс восстановления повреждённой ДНК ферментами репарационных систем
Различают 2 типа репарационных систем:
Система
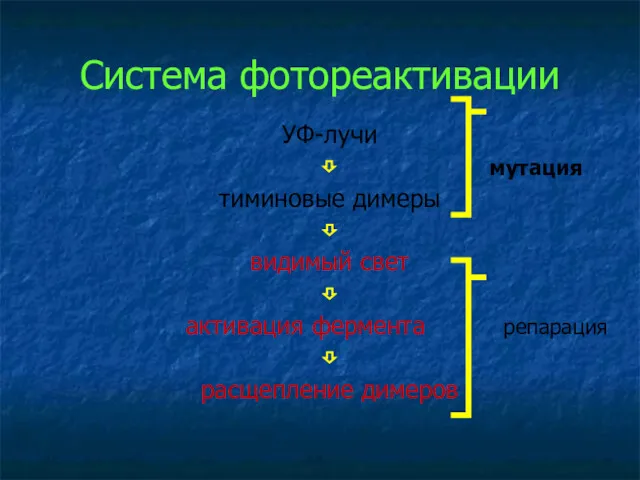
Система фотореактивации
УФ-лучи
⇩ мутация
тиминовые димеры
⇩
видимый свет
⇩
активация фермента репарация
⇩
расщепление димеров
Система фотореактивации
УФ-лучи
⇩ мутация
тиминовые димеры
⇩
видимый свет
⇩
активация фермента репарация
⇩
расщепление димеров

Этапы темновой репарации:
установление места повреждения ДНК = эндонуклеаза,
«вырезание»
Этапы темновой репарации:
установление места повреждения ДНК = эндонуклеаза,
«вырезание»

Система темновой репарации
УФ-лучи
⇩
тиминовые димеры
⇩
темнота
⇩
Система темновой репарации
УФ-лучи
⇩
тиминовые димеры
⇩
темнота
⇩

Система темновой репарации
обнаружение и нарезание повреждённого участка
(эндонуклеаза)
⇩
Система темновой репарации
обнаружение и нарезание повреждённого участка
(эндонуклеаза)
⇩

Система темновой репарации
удаление повреждённого участка
(ДНК-полимераза I)
⇩
Система темновой репарации
удаление повреждённого участка
(ДНК-полимераза I)
⇩

Система темновой репарации
синтез на матрице второй нити ДНК нового, не содержащего
Система темновой репарации
синтез на матрице второй нити ДНК нового, не содержащего

Система темновой репарации
«вшивание» нового участка в цепь ДНК
(лигаза)
Система темновой репарации
«вшивание» нового участка в цепь ДНК
(лигаза)

Генетические рекомбинации
= перераспределение генетического материала родителей в потомстве, обусловливающее комбинативную изменчивость
Генетические рекомбинации
= перераспределение генетического материала родителей в потомстве, обусловливающее комбинативную изменчивость

Механизм рекомбинаций
клетки=доноры
⇩
передают информацию
⇩
клеткам-реципиентам
⇩
рекомбинат
генотип рекомбинанта
Механизм рекомбинаций
клетки=доноры
⇩
передают информацию
⇩
клеткам-реципиентам
⇩
рекомбинат
генотип рекомбинанта

ВИДЫ РЕКОМБИНАТИВНОЙ ИЗМЕНЧИВОСТИ У БАКТЕРИЙ
Трансформация – непосредственная передача генетического материала от
ВИДЫ РЕКОМБИНАТИВНОЙ ИЗМЕНЧИВОСТИ У БАКТЕРИЙ
Трансформация – непосредственная передача генетического материала от

Трансформация
= способ передачи генетической информации путем внедрения свободной ДНК донора в
Трансформация
= способ передачи генетической информации путем внедрения свободной ДНК донора в

Трансформация
Клетки, способные принимать донорскую ДНК, называются компетентными.
Состояние компетентности возникает в
Трансформация
Клетки, способные принимать донорскую ДНК, называются компетентными.
Состояние компетентности возникает в

Процесс трансформации состоит из фаз:
адсорбция ДНК донора на клетке-реципиенте,
проникновение ДНК внутрь
Процесс трансформации состоит из фаз:
адсорбция ДНК донора на клетке-реципиенте,
проникновение ДНК внутрь
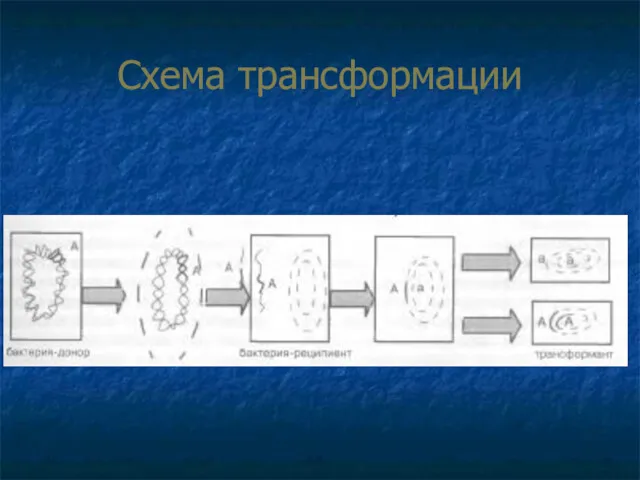
Схема трансформации
Схема трансформации

Конъюгация
– перенос генетического материала из клетки-донора в клетку реципиента при тесном
Конъюгация
– перенос генетического материала из клетки-донора в клетку реципиента при тесном

Конъюгация
2 вида конъюгации:
Если F-плазмида автономна→ бактерия наз-ся F+ штаммом
2.Если F-плазмида
Конъюгация
2 вида конъюгации:
Если F-плазмида автономна→ бактерия наз-ся F+ штаммом
2.Если F-плазмида

1 Если F-плазмида автономна:
1. Прикрепление клетки донора к реципиенту с помощью
1 Если F-плазмида автономна:
1. Прикрепление клетки донора к реципиенту с помощью
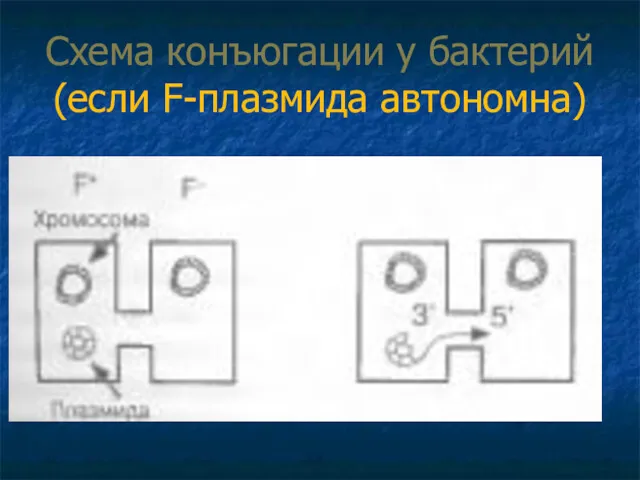
Схема конъюгации у бактерий (если F-плазмида автономна)
Схема конъюгации у бактерий (если F-плазмида автономна)

1 Если F-плазмида автономна:
2.3. белок способствует замыканию линейной нити в
1 Если F-плазмида автономна:
2.3. белок способствует замыканию линейной нити в
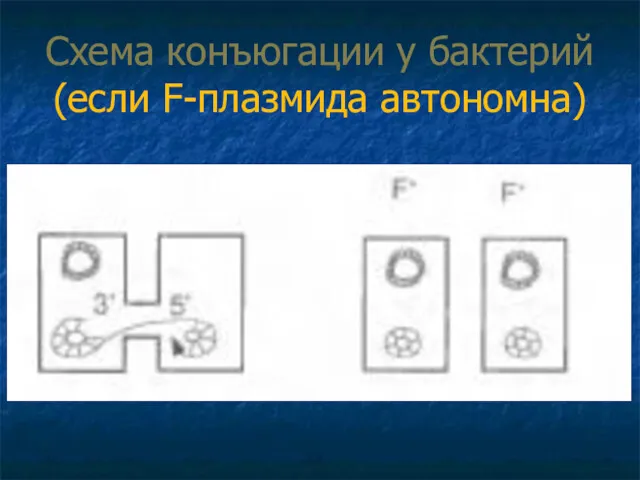
Схема конъюгации у бактерий (если F-плазмида автономна)
Схема конъюгации у бактерий (если F-плазмида автономна)

2.Если F-плазмида встроена в хромосому бактерии = Hfr-штамм:
Происходит разрыв одной
2.Если F-плазмида встроена в хромосому бактерии = Hfr-штамм:
Происходит разрыв одной
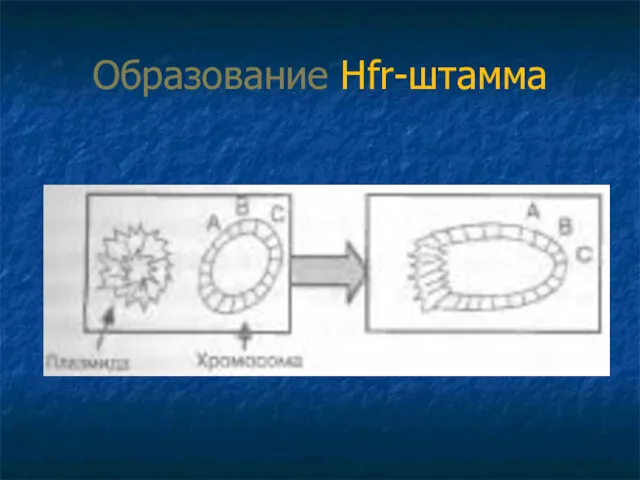
Образование Hfr-штамма
Образование Hfr-штамма

Схема конъюгации Hfr-штамма
разрыв одной нити ДНК при участии эндонуклеазы в
Схема конъюгации Hfr-штамма
разрыв одной нити ДНК при участии эндонуклеазы в

Схема конъюгации Hfr-штамма
Проксимальный конец ДНК через конъюгационный мостик проникает в
Схема конъюгации Hfr-штамма
Проксимальный конец ДНК через конъюгационный мостик проникает в
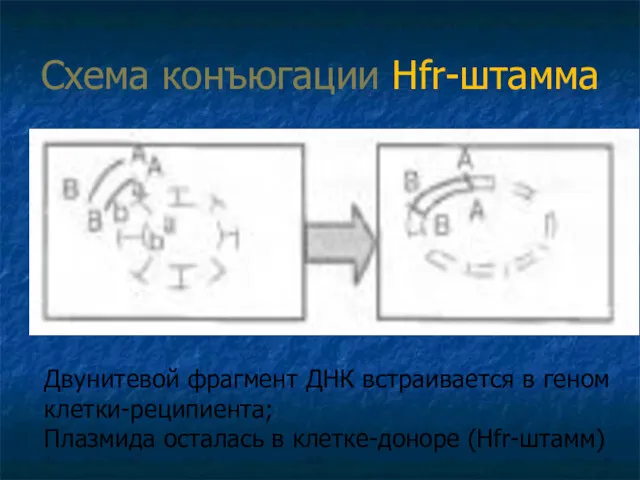
Схема конъюгации Hfr-штамма
Двунитевой фрагмент ДНК встраивается в геном клетки-реципиента;
Плазмида осталась
Схема конъюгации Hfr-штамма
Двунитевой фрагмент ДНК встраивается в геном клетки-реципиента;
Плазмида осталась

Трансдукция
– передача генетического материала от одной бактерии к другой
Трансдукция
– передача генетического материала от одной бактерии к другой

Общая = неспецифическая трансдукция
– когда в клетку–реципиент вместе с фаговой ДНК
Общая = неспецифическая трансдукция
– когда в клетку–реципиент вместе с фаговой ДНК

Специфическая трансдукция
– фаг переносит специфические гены от бактерии-донора к бактерии-реципиенту:
При
Специфическая трансдукция
– фаг переносит специфические гены от бактерии-донора к бактерии-реципиенту:
При

Абортивная трансдукция
= принесенный фагом фрагмент ДНК бактерии-донора не включается в хромосому
Абортивная трансдукция
= принесенный фагом фрагмент ДНК бактерии-донора не включается в хромосому
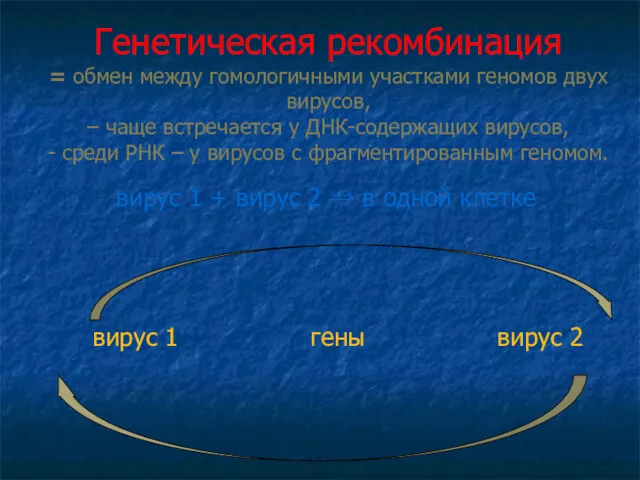
Генетическая рекомбинация
= обмен между гомологичными участками геномов двух вирусов,
– чаще
Генетическая рекомбинация = обмен между гомологичными участками геномов двух вирусов, – чаще

Генетическая реактивация
= обмен между геномами родственных вирусов, у которых мутации произошли
Генетическая реактивация = обмен между геномами родственных вирусов, у которых мутации произошли

Комплементация = обмен, когда один из двух вирусов в результате мутации
Комплементация = обмен, когда один из двух вирусов в результате мутации

Фенотипическое смешивание
при смешанном заражении двумя вирусами часть потомства приобретает фенотипические признаки,
Фенотипическое смешивание
при смешанном заражении двумя вирусами часть потомства приобретает фенотипические признаки,

Фенотипическое смешивание
вирус 1 + вирус 2 ⇒ в одной клетке
вирус
Фенотипическое смешивание
вирус 1 + вирус 2 ⇒ в одной клетке
вирус

ГЕННАЯ ИНЖЕНЕРИЯ В МЕДИЦИНСКОЙ МИКРОБИОЛОГИИ
Продукты, получаемые генно-инженерным способом с помощью рекомбинантных
ГЕННАЯ ИНЖЕНЕРИЯ В МЕДИЦИНСКОЙ МИКРОБИОЛОГИИ
Продукты, получаемые генно-инженерным способом с помощью рекомбинантных

ГЕННАЯ ИНЖЕНЕРИЯ В МЕДИЦИНСКОЙ МИКРОБИОЛОГИИ
Получение рекомбинантной вакцины для профилактики гепатита В
встраивание
ГЕННАЯ ИНЖЕНЕРИЯ В МЕДИЦИНСКОЙ МИКРОБИОЛОГИИ
Получение рекомбинантной вакцины для профилактики гепатита В
встраивание

ГЕНЕТИЧЕСКИЕ МЕТОДЫ, ПРИМЕНЯЕМЫЕ В МИКРОБИОЛОГИЧЕСКОЙ ДИАГНОСТИКЕ
процентное содержание Г+Ц в бактериальном геноме
метод
ГЕНЕТИЧЕСКИЕ МЕТОДЫ, ПРИМЕНЯЕМЫЕ В МИКРОБИОЛОГИЧЕСКОЙ ДИАГНОСТИКЕ
процентное содержание Г+Ц в бактериальном геноме
метод

МЕТОД МОЛЕКУЛЯРНОЙ ГИБРИДИЗАЦИИ
Цель
Выявления степени сходства различных ДНК (при идентификации микроорганизмов –
МЕТОД МОЛЕКУЛЯРНОЙ ГИБРИДИЗАЦИИ
Цель
Выявления степени сходства различных ДНК (при идентификации микроорганизмов –

МЕТОД МОЛЕКУЛЯРНОЙ ГИБРИДИЗАЦИИ
Принцип осуществления
исследуемая ДНК
⇩
нагрев в щелочной среде
⇩
расплетение на две отдельные
МЕТОД МОЛЕКУЛЯРНОЙ ГИБРИДИЗАЦИИ
Принцип осуществления
исследуемая ДНК
⇩
нагрев в щелочной среде
⇩
расплетение на две отдельные

ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ
Цели
обнаружение в патологическом материале конкретного вида микроорганизма без выделения
ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ
Цели
обнаружение в патологическом материале конкретного вида микроорганизма без выделения

ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ
Принцип осуществления
патологический материал или штамм микроорганизма
⇩
выделение ДНК
⇩
нагрев
⇩
расплетение ДНК на
ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ
Принцип осуществления
патологический материал или штамм микроорганизма
⇩
выделение ДНК
⇩
нагрев
⇩
расплетение ДНК на

ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ
Принцип осуществления
⇩
добавление ДНК-полимеразы и нуклеотидов
⇩
нуклеотиды присоединяются к 3’-концам праймеров
⇩
повторение
ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ
Принцип осуществления
⇩
добавление ДНК-полимеразы и нуклеотидов
⇩
нуклеотиды присоединяются к 3’-концам праймеров
⇩
повторение
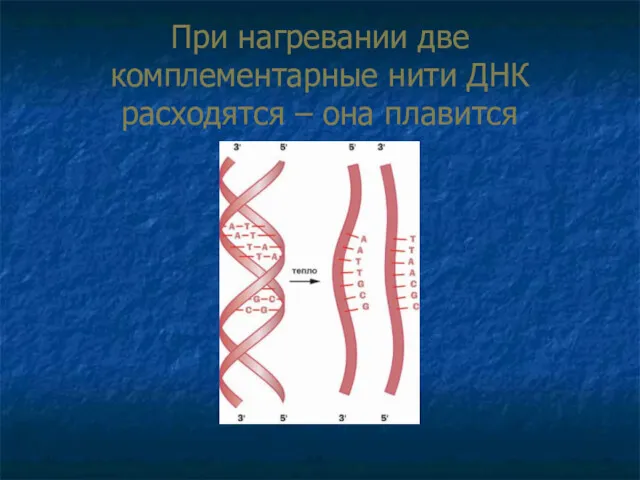
При нагревании две комплементарные нити ДНК расходятся – она плавится
При нагревании две комплементарные нити ДНК расходятся – она плавится

ПЦР
ПЦР

Рестрикционный анализ
Расщепление ДНК микроорганизмов на фрагменты при помощи рестриктаз (эндонуклеаз),
От бактерий
Рестрикционный анализ
Расщепление ДНК микроорганизмов на фрагменты при помощи рестриктаз (эндонуклеаз),
От бактерий
 Плісеневі гриби
Плісеневі гриби Класс насекомые
Класс насекомые Царство Животные. Общая характеристика. Классификация животных
Царство Животные. Общая характеристика. Классификация животных Витамины
Витамины Отряд Хищные
Отряд Хищные СРС на тему: “Везикулярный транспорт”
СРС на тему: “Везикулярный транспорт” Растворы. Качественный и количественный состав растворов
Растворы. Качественный и количественный состав растворов Our world
Our world Хвощи и плауны
Хвощи и плауны פירמידות המזון
פירמידות המזון Гетерозис ғылымының теориясы мен болжамы
Гетерозис ғылымының теориясы мен болжамы Яйцеклетки. Овогенез
Яйцеклетки. Овогенез Глюконеогенез. Регуляция гликолиза и глюконеогенеза. Цикл кори. Пентозофосфатный путь превращения глюкозы. Спиртовое брожение
Глюконеогенез. Регуляция гликолиза и глюконеогенеза. Цикл кори. Пентозофосфатный путь превращения глюкозы. Спиртовое брожение Викторина Анализаторы
Викторина Анализаторы Диво-дивное. Любопытные факты о животных
Диво-дивное. Любопытные факты о животных Механизмы и закономерности эволюции
Механизмы и закономерности эволюции Тип Членистоногие
Тип Членистоногие Какие функции выполняет корень?
Какие функции выполняет корень? Растительный мир Курской области
Растительный мир Курской области Среды жизни и места обитания животных. Взаимосвязь животных в природе
Среды жизни и места обитания животных. Взаимосвязь животных в природе Подцарство Одноклеточные. Часть 1. 5 - 7 класс
Подцарство Одноклеточные. Часть 1. 5 - 7 класс Клітинна радіобіологія. Теорія мішеней в клітинній радіобіології
Клітинна радіобіологія. Теорія мішеней в клітинній радіобіології Эпиграф
Эпиграф Викторина Эти забавные животные
Викторина Эти забавные животные презентация на тему:Витамины
презентация на тему:Витамины Роль биологических и социальных факторов в эволюции человека
Роль биологических и социальных факторов в эволюции человека Школьная клумба
Школьная клумба Увеличительные приборы для исследования в биологии
Увеличительные приборы для исследования в биологии