- Генетика. Хромосомная теория наследственности

Содержание
- 2. ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ - теория, согласно которой передача наследственной информации в ряду поколений связана с передачей
- 3. ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ. ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ: Носителями наследственности являются гены, находящиеся в хромосомах.
- 4. ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ. ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ: Носителями наследственности являются гены, находящиеся в хромосомах. Каждый ген имеет
- 5. ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ. ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ: Носителями наследственности являются гены, находящиеся в хромосомах. Каждый ген имеет
- 6. ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ. ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ: Носителями наследственности являются гены, находящиеся в хромосомах. Каждый ген имеет
- 7. ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ. ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ: Носителями наследственности являются гены, находящиеся в хромосомах. Каждый ген имеет
- 8. ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ. ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ: Носителями наследственности являются гены, находящиеся в хромосомах. Каждый ген имеет
- 9. ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ. ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ: Носителями наследственности являются гены, находящиеся в хромосомах. Каждый ген имеет
- 10. ГЕНЕТИЧЕСКИЙ АНАЛИЗ исследование генотипа отдельных особей, групп особей и генетической структуры популяций, в том числе линий,
- 11. ГЕНЕТИЧЕСКИЙ АНАЛИЗ исследование генотипа отдельных особей, групп особей и генетической структуры популяций, в том числе линий,
- 12. ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ. - способ изучения наследственных свойств организма путём скрещивания его с родственной формой и
- 13. ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ. – заключается в анализе родословных и позволяет определить тип наследования (доминантный, рецессивный, аутосомный
- 14. ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ. - заключается в анализе и сравнении изменчивости признаков в пределах различных групп близнецов,
- 15. ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ. Гибридологический метод Генеалогический Близнецовый метод Цитогенетические методы Мутационные методы Молекулярно-генетические биохимические и физико-химические
- 16. ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ. Гибридологический метод Генеалогический Близнецовый метод Цитогенетический метод Мутационные методы Молекулярно-генетические биохимические и физико-химические
- 17. ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ. Гибридологический метод Генеалогический Близнецовый метод Цитогенетический метод Мутационные методы Молекулярно-генетические биохимические и физико-химические
- 18. ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ. Гибридологический метод Генеалогический Близнецовый метод Цитогенетический метод Мутационные методы Молекулярно-генетические биохимические и физико-химические
- 19. ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ. Гибридологический метод Генеалогический Близнецовый метод Цитогенетический метод Мутационные методы Молекулярно-генетические биохимические и физико-химические
- 20. ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ. Гибридологический метод Генеалогический Близнецовый метод Цитогенетический метод Мутационные методы Молекулярно-генетические биохимические и физико-химические
- 21. ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ. Гибридологический метод Генеалогический Близнецовый метод Цитогенетический метод Мутационные методы Молекулярно-генетические биохимические и физико-химические
- 22. РАСЩЕПЛЕНИЕ ПРИ МОНОГИБРИДНОМ СКРЕЩИВАНИИ. Моногибридное скрещивание - скрещивание форм, отличающихся друг от друга по одной паре
- 23. РАСЩЕПЛЕНИЕ ПРИ МОНОГИБРИДНОМ СКРЕЩИВАНИИ. Моногибридное скрещивание - скрещивание форм, отличающихся друг от друга по одной паре
- 24. РАСЩЕПЛЕНИЕ ПРИ МОНОГИБРИДНОМ СКРЕЩИВАНИИ. Моногибридное скрещивание - скрещивание форм, отличающихся друг от друга по одной паре
- 25. РАСЩЕПЛЕНИЕ ПРИ МОНОГИБРИДНОМ СКРЕЩИВАНИИ. Анализирующее скрещивание — скрещивание гибридной особи с особью, гомозиготной по рецессивным аллелям,
- 26. РАСЩЕПЛЕНИЕ ПРИ МОНОГИБРИДНОМ СКРЕЩИВАНИИ. Чистая линия — группа организмов, имеющих некоторые признаки, которые полностью передаются потомству
- 27. РАСЩЕПЛЕНИЕ ПРИ ДИГИБРИДНОМ СКРЕЩИВАНИИ. Дигибридное скрещивание — скрещивание организмов, различающихся по двум парам альтернативных признаков. Закон
- 28. Дигибридное скрещивание — скрещивание организмов, различающихся по двум парам альтернативных признаков. Расщепления, характерныедля независимого наследования при
- 29. РАСЩЕПЛЕНИЕ ПРИ ПОЛИГИБРИДНОМ СКРЕЩИВАНИИ.
- 30. ВЗАИМОДЕЙСТВИЕ ГЕНОВ. АЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ полное доминирование неполное доминирование сверхдоминирование кодоминирование множественный аллелизм Полное доминирование – взаимодействие
- 31. ВЗАИМОДЕЙСТВИЕ ГЕНОВ. АЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ полное доминирование неполное доминирование сверхдоминирование кодоминирование множественный аллелизм Неполное доминирование – доминантный
- 32. ВЗАИМОДЕЙСТВИЕ ГЕНОВ. АЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ полное доминирование неполное доминирование сверхдоминирование кодоминирование множественный аллелизм Сверхдоминирование – когда доминантный
- 33. ВЗАИМОДЕЙСТВИЕ ГЕНОВ. АЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ полное доминирование неполное доминирование сверхдоминирование кодоминирование множественный аллелизм Кодоминирование – проявление у
- 34. ВЗАИМОДЕЙСТВИЕ ГЕНОВ. АЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ полное доминирование неполное доминирование сверхдоминирование кодоминирование множественный аллелизм Множественный аллелизм. Есть несколько
- 35. ВЗАИМОДЕЙСТВИЕ ГЕНОВ. НЕАЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ комплементарность эпистаз полимерия плейотропия Комплементарность – один доминантный ген дополняет действие другого
- 36. ВЗАИМОДЕЙСТВИЕ ГЕНОВ. НЕАЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ комплементарность эпистаз полимерия плейотропия – один ген подавляет действие другого неаллельного гена.
- 37. ВЗАИМОДЕЙСТВИЕ ГЕНОВ. НЕАЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ комплементарность эпистаз полимерия плейотропия – один ген подавляет действие другого неаллельного гена.
- 38. ВЗАИМОДЕЙСТВИЕ ГЕНОВ. НЕАЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ комплементарность эпистаз полимерия плейотропия Полимерия – взаимодействие множественных генов, одновременно влияющих на
- 39. ВЗАИМОДЕЙСТВИЕ ГЕНОВ. НЕАЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ комплементарность эпистаз полимерия плейотропия Плейотропия – способность одного гена влиять на несколько
- 40. ОТКЛОНЕНИЕ ОТ РАСЩЕПЛЕНИЯ. ПРИЧИНЫ. неодинаковая жизнеспособность зигот; случайное нерасхождение гамет; неравная вероятность образования всех типов гамет;
- 41. НАСЛЕДОВАНИЕ ПРИЗНАКОВ, СЦЕПЛЕННЫХ С ПОЛОМ. Диплоидный набор хромосом можно разделить на аутосомы и половые хромосомы. Половые
- 42. НАСЛЕДОВАНИЕ ПРИЗНАКОВ, СЦЕПЛЕННЫХ С ПОЛОМ. Особь женского пола может быть как гомо-, так и гетерозиготной по
- 43. НАСЛЕДОВАНИЕ ПРИЗНАКОВ, СЦЕПЛЕННЫХ С ПОЛОМ.
- 44. НЕЗАВИСИМОЕ И СЦЕПЛЕННОЕ НАСЛЕДОВАНИЕ. Наследование признаков, которые кодируются генами, локализованными в разных парах хромосом, осуществляется независимо
- 45. КРОССИНГОВЕР. Кроссинговер - процесс обмена участками гомологичных хромосом во время конъюгации в профазе I мейоза. Процент
- 46. ИНТЕРФЕРЕНЦИЯ. Кроссоверные события влияют друг на друга. Это влияние называется интерференцией. Величину интерференции можно выразить с
- 47. СОМАТИЧЕСКИЙ КРОССИНГОВЕР. Кроссинговер может происходить не только в мейозе, но и в соматических клетках в ходе
- 48. НЕРАВНЫЙ КРОССИНГОВЕР. Неравный кроссинговер — кроссинговер, в результате которого образуются сестринские кроссоверные хроматиды, различающиеся по количеству
- 49. КОНВЕРСИЯ. Разрешение структур Холлидея состоит в разрезании перекрещивающихся цепей и сшивании их таким образом, что перекрест
- 50. НАСЛЕДСТВЕННАЯ ИЗМЕНЧИВОСТЬ. Изменчивость - один из основных факторов эволюции, является источником для искусственного и естественного отборов.
- 51. НЕНАСЛЕДСТВЕННАЯ ИЗМЕНЧИВОСТЬ. Ненаследственная (модификационная изменчивость) — это результат не изменений генотипа, а его реакции на условия
- 52. НОРМА РЕАКЦИИ ПРИЗНАКА. Норма реакции — спектр возможных уровней экспрессии генов, из которого выбирается уровень экспрессии,
- 54. Скачать презентацию

ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ
- теория, согласно которой передача наследственной информации в
ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ
- теория, согласно которой передача наследственной информации в

ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ.
ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ:
Носителями наследственности являются гены, находящиеся в
ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ.
ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ:
Носителями наследственности являются гены, находящиеся в

ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ.
ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ:
Носителями наследственности являются гены, находящиеся в
ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ.
ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ:
Носителями наследственности являются гены, находящиеся в

ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ.
ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ:
Носителями наследственности являются гены, находящиеся в
ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ.
ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ:
Носителями наследственности являются гены, находящиеся в

ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ.
ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ:
Носителями наследственности являются гены, находящиеся в
ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ.
ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ:
Носителями наследственности являются гены, находящиеся в

ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ.
ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ:
Носителями наследственности являются гены, находящиеся в
ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ.
ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ:
Носителями наследственности являются гены, находящиеся в

ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ.
ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ:
Носителями наследственности являются гены, находящиеся в
ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ.
ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ:
Носителями наследственности являются гены, находящиеся в

ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ.
ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ:
Носителями наследственности являются гены, находящиеся в
ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ.
ОСНОВНЫЕ ПОЛОЖЕНИЯ ТЕОРИИ:
Носителями наследственности являются гены, находящиеся в

ГЕНЕТИЧЕСКИЙ АНАЛИЗ
исследование генотипа отдельных особей, групп особей и генетической структуры популяций,
ГЕНЕТИЧЕСКИЙ АНАЛИЗ
исследование генотипа отдельных особей, групп особей и генетической структуры популяций,

ГЕНЕТИЧЕСКИЙ АНАЛИЗ
исследование генотипа отдельных особей, групп особей и генетической структуры популяций,
ГЕНЕТИЧЕСКИЙ АНАЛИЗ
исследование генотипа отдельных особей, групп особей и генетической структуры популяций,

ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
- способ изучения наследственных свойств организма путём скрещивания его
ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
- способ изучения наследственных свойств организма путём скрещивания его

ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
– заключается в анализе родословных и позволяет определить тип
ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
– заключается в анализе родословных и позволяет определить тип

ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
- заключается в анализе и сравнении изменчивости признаков в
ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
- заключается в анализе и сравнении изменчивости признаков в

ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
Гибридологический метод
Генеалогический
Близнецовый метод
Цитогенетические методы
Мутационные методы
Молекулярно-генетические
биохимические и физико-химические
Популяционный
Онтогенетический метод
Математический
ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
Гибридологический метод
Генеалогический
Близнецовый метод
Цитогенетические методы
Мутационные методы
Молекулярно-генетические
биохимические и физико-химические
Популяционный
Онтогенетический метод
Математический

ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
Гибридологический метод
Генеалогический
Близнецовый метод
Цитогенетический метод
Мутационные методы
Молекулярно-генетические
биохимические и физико-химические
Популяционный
Онтогенетический метод
Математический
ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
Гибридологический метод
Генеалогический
Близнецовый метод
Цитогенетический метод
Мутационные методы
Молекулярно-генетические
биохимические и физико-химические
Популяционный
Онтогенетический метод
Математический

ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
Гибридологический метод
Генеалогический
Близнецовый метод
Цитогенетический метод
Мутационные методы
Молекулярно-генетические
биохимические и физико-химические
Популяционный
Онтогенетический метод
Математический
ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
Гибридологический метод
Генеалогический
Близнецовый метод
Цитогенетический метод
Мутационные методы
Молекулярно-генетические
биохимические и физико-химические
Популяционный
Онтогенетический метод
Математический

ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
Гибридологический метод
Генеалогический
Близнецовый метод
Цитогенетический метод
Мутационные методы
Молекулярно-генетические
биохимические и физико-химические
Популяционный
Онтогенетический метод
Математический
ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
Гибридологический метод
Генеалогический
Близнецовый метод
Цитогенетический метод
Мутационные методы
Молекулярно-генетические
биохимические и физико-химические
Популяционный
Онтогенетический метод
Математический

ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
Гибридологический метод
Генеалогический
Близнецовый метод
Цитогенетический метод
Мутационные методы
Молекулярно-генетические
биохимические и физико-химические
Популяционный
Онтогенетический метод
Математический
ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
Гибридологический метод
Генеалогический
Близнецовый метод
Цитогенетический метод
Мутационные методы
Молекулярно-генетические
биохимические и физико-химические
Популяционный
Онтогенетический метод
Математический

ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
Гибридологический метод
Генеалогический
Близнецовый метод
Цитогенетический метод
Мутационные методы
Молекулярно-генетические
биохимические и физико-химические
Популяционный
Онтогенетический метод
Математический
ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
Гибридологический метод
Генеалогический
Близнецовый метод
Цитогенетический метод
Мутационные методы
Молекулярно-генетические
биохимические и физико-химические
Популяционный
Онтогенетический метод
Математический

ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
Гибридологический метод
Генеалогический
Близнецовый метод
Цитогенетический метод
Мутационные методы
Молекулярно-генетические
биохимические и физико-химические
Популяционный
Онтогенетический метод
Математический
ГЕНЕТИЧЕСКИЙ АНАЛИЗ. МЕТОДЫ.
Гибридологический метод
Генеалогический
Близнецовый метод
Цитогенетический метод
Мутационные методы
Молекулярно-генетические
биохимические и физико-химические
Популяционный
Онтогенетический метод
Математический
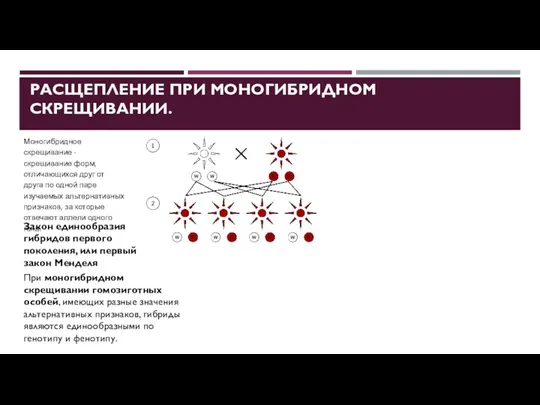
РАСЩЕПЛЕНИЕ ПРИ МОНОГИБРИДНОМ СКРЕЩИВАНИИ.
Моногибридное скрещивание - скрещивание форм, отличающихся друг
РАСЩЕПЛЕНИЕ ПРИ МОНОГИБРИДНОМ СКРЕЩИВАНИИ.
Моногибридное скрещивание - скрещивание форм, отличающихся друг

РАСЩЕПЛЕНИЕ ПРИ МОНОГИБРИДНОМ СКРЕЩИВАНИИ.
Моногибридное скрещивание - скрещивание форм, отличающихся друг
РАСЩЕПЛЕНИЕ ПРИ МОНОГИБРИДНОМ СКРЕЩИВАНИИ.
Моногибридное скрещивание - скрещивание форм, отличающихся друг
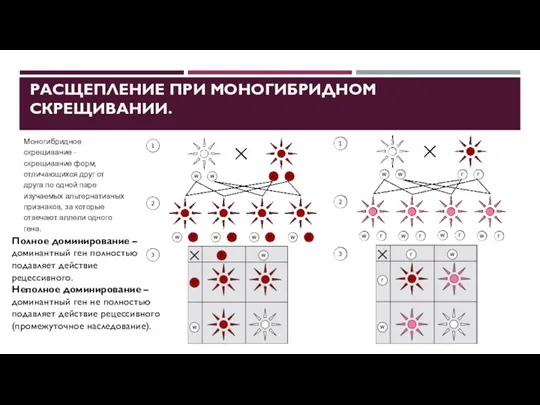
РАСЩЕПЛЕНИЕ ПРИ МОНОГИБРИДНОМ СКРЕЩИВАНИИ.
Моногибридное скрещивание - скрещивание форм, отличающихся друг
РАСЩЕПЛЕНИЕ ПРИ МОНОГИБРИДНОМ СКРЕЩИВАНИИ.
Моногибридное скрещивание - скрещивание форм, отличающихся друг
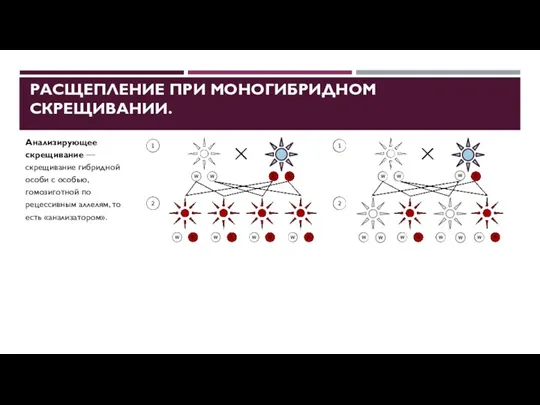
РАСЩЕПЛЕНИЕ ПРИ МОНОГИБРИДНОМ СКРЕЩИВАНИИ.
Анализирующее скрещивание — скрещивание гибридной особи с
РАСЩЕПЛЕНИЕ ПРИ МОНОГИБРИДНОМ СКРЕЩИВАНИИ.
Анализирующее скрещивание — скрещивание гибридной особи с

РАСЩЕПЛЕНИЕ ПРИ МОНОГИБРИДНОМ СКРЕЩИВАНИИ.
Чистая линия — группа организмов, имеющих некоторые
РАСЩЕПЛЕНИЕ ПРИ МОНОГИБРИДНОМ СКРЕЩИВАНИИ.
Чистая линия — группа организмов, имеющих некоторые

РАСЩЕПЛЕНИЕ ПРИ ДИГИБРИДНОМ СКРЕЩИВАНИИ.
Дигибридное скрещивание — скрещивание организмов, различающихся по
РАСЩЕПЛЕНИЕ ПРИ ДИГИБРИДНОМ СКРЕЩИВАНИИ.
Дигибридное скрещивание — скрещивание организмов, различающихся по

Дигибридное скрещивание — скрещивание организмов, различающихся по двум парам альтернативных признаков.
Расщепления,
Дигибридное скрещивание — скрещивание организмов, различающихся по двум парам альтернативных признаков.
Расщепления,

РАСЩЕПЛЕНИЕ ПРИ ПОЛИГИБРИДНОМ СКРЕЩИВАНИИ.
РАСЩЕПЛЕНИЕ ПРИ ПОЛИГИБРИДНОМ СКРЕЩИВАНИИ.

ВЗАИМОДЕЙСТВИЕ ГЕНОВ. АЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
полное доминирование
неполное доминирование
сверхдоминирование
кодоминирование
множественный аллелизм
Полное доминирование – взаимодействие двух
ВЗАИМОДЕЙСТВИЕ ГЕНОВ. АЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
полное доминирование
неполное доминирование
сверхдоминирование
кодоминирование
множественный аллелизм
Полное доминирование – взаимодействие двух

ВЗАИМОДЕЙСТВИЕ ГЕНОВ. АЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
полное доминирование
неполное доминирование
сверхдоминирование
кодоминирование
множественный аллелизм
Неполное доминирование – доминантный аллель
ВЗАИМОДЕЙСТВИЕ ГЕНОВ. АЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
полное доминирование
неполное доминирование
сверхдоминирование
кодоминирование
множественный аллелизм
Неполное доминирование – доминантный аллель

ВЗАИМОДЕЙСТВИЕ ГЕНОВ. АЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
полное доминирование
неполное доминирование
сверхдоминирование
кодоминирование
множественный аллелизм
Сверхдоминирование – когда доминантный ген
ВЗАИМОДЕЙСТВИЕ ГЕНОВ. АЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
полное доминирование
неполное доминирование
сверхдоминирование
кодоминирование
множественный аллелизм
Сверхдоминирование – когда доминантный ген

ВЗАИМОДЕЙСТВИЕ ГЕНОВ. АЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
полное доминирование
неполное доминирование
сверхдоминирование
кодоминирование
множественный аллелизм
Кодоминирование – проявление у гетерозигот
ВЗАИМОДЕЙСТВИЕ ГЕНОВ. АЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
полное доминирование
неполное доминирование
сверхдоминирование
кодоминирование
множественный аллелизм
Кодоминирование – проявление у гетерозигот

ВЗАИМОДЕЙСТВИЕ ГЕНОВ. АЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
полное доминирование
неполное доминирование
сверхдоминирование
кодоминирование
множественный аллелизм
Множественный аллелизм. Есть несколько аллелей.
ВЗАИМОДЕЙСТВИЕ ГЕНОВ. АЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
полное доминирование
неполное доминирование
сверхдоминирование
кодоминирование
множественный аллелизм
Множественный аллелизм. Есть несколько аллелей.

ВЗАИМОДЕЙСТВИЕ ГЕНОВ. НЕАЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
комплементарность
эпистаз
полимерия
плейотропия
Комплементарность – один доминантный ген дополняет действие другого
ВЗАИМОДЕЙСТВИЕ ГЕНОВ. НЕАЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
комплементарность
эпистаз
полимерия
плейотропия
Комплементарность – один доминантный ген дополняет действие другого

ВЗАИМОДЕЙСТВИЕ ГЕНОВ. НЕАЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
комплементарность
эпистаз
полимерия
плейотропия
– один ген подавляет действие другого неаллельного гена.
ВЗАИМОДЕЙСТВИЕ ГЕНОВ. НЕАЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
комплементарность
эпистаз
полимерия
плейотропия
– один ген подавляет действие другого неаллельного гена.

ВЗАИМОДЕЙСТВИЕ ГЕНОВ. НЕАЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
комплементарность
эпистаз
полимерия
плейотропия
– один ген подавляет действие другого неаллельного гена.
ВЗАИМОДЕЙСТВИЕ ГЕНОВ. НЕАЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
комплементарность
эпистаз
полимерия
плейотропия
– один ген подавляет действие другого неаллельного гена.

ВЗАИМОДЕЙСТВИЕ ГЕНОВ. НЕАЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
комплементарность
эпистаз
полимерия
плейотропия
Полимерия – взаимодействие множественных генов, одновременно влияющих на
ВЗАИМОДЕЙСТВИЕ ГЕНОВ. НЕАЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
комплементарность
эпистаз
полимерия
плейотропия
Полимерия – взаимодействие множественных генов, одновременно влияющих на

ВЗАИМОДЕЙСТВИЕ ГЕНОВ. НЕАЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
комплементарность
эпистаз
полимерия
плейотропия
Плейотропия – способность одного гена влиять на несколько
ВЗАИМОДЕЙСТВИЕ ГЕНОВ. НЕАЛЛЕЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
комплементарность
эпистаз
полимерия
плейотропия
Плейотропия – способность одного гена влиять на несколько

ОТКЛОНЕНИЕ ОТ РАСЩЕПЛЕНИЯ. ПРИЧИНЫ.
неодинаковая жизнеспособность зигот;
случайное нерасхождение гамет;
неравная вероятность образования всех
ОТКЛОНЕНИЕ ОТ РАСЩЕПЛЕНИЯ. ПРИЧИНЫ.
неодинаковая жизнеспособность зигот;
случайное нерасхождение гамет;
неравная вероятность образования всех

НАСЛЕДОВАНИЕ ПРИЗНАКОВ, СЦЕПЛЕННЫХ С ПОЛОМ.
Диплоидный набор хромосом можно разделить на
НАСЛЕДОВАНИЕ ПРИЗНАКОВ, СЦЕПЛЕННЫХ С ПОЛОМ.
Диплоидный набор хромосом можно разделить на
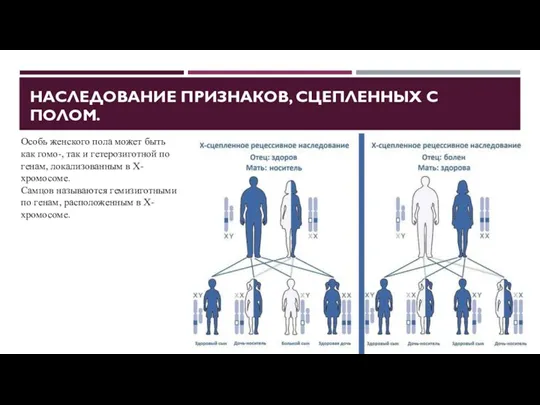
НАСЛЕДОВАНИЕ ПРИЗНАКОВ, СЦЕПЛЕННЫХ С ПОЛОМ.
Особь женского пола может быть как
НАСЛЕДОВАНИЕ ПРИЗНАКОВ, СЦЕПЛЕННЫХ С ПОЛОМ.
Особь женского пола может быть как
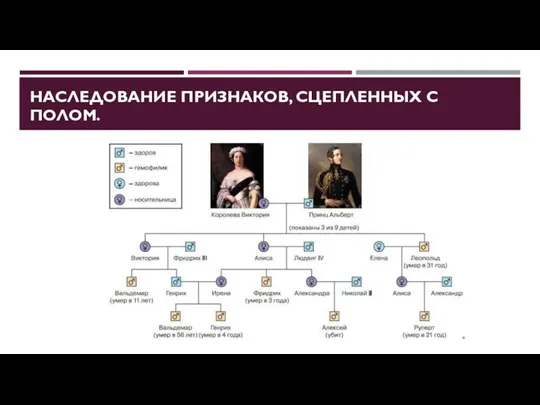
НАСЛЕДОВАНИЕ ПРИЗНАКОВ, СЦЕПЛЕННЫХ С ПОЛОМ.
НАСЛЕДОВАНИЕ ПРИЗНАКОВ, СЦЕПЛЕННЫХ С ПОЛОМ.

НЕЗАВИСИМОЕ И СЦЕПЛЕННОЕ НАСЛЕДОВАНИЕ.
Наследование признаков, которые кодируются генами, локализованными в
НЕЗАВИСИМОЕ И СЦЕПЛЕННОЕ НАСЛЕДОВАНИЕ.
Наследование признаков, которые кодируются генами, локализованными в

КРОССИНГОВЕР.
Кроссинговер - процесс обмена участками гомологичных хромосом во время конъюгации
КРОССИНГОВЕР.
Кроссинговер - процесс обмена участками гомологичных хромосом во время конъюгации

ИНТЕРФЕРЕНЦИЯ.
Кроссоверные события влияют друг на друга. Это влияние называется интерференцией.
ИНТЕРФЕРЕНЦИЯ.
Кроссоверные события влияют друг на друга. Это влияние называется интерференцией.

СОМАТИЧЕСКИЙ КРОССИНГОВЕР.
Кроссинговер может происходить не только в мейозе, но и
СОМАТИЧЕСКИЙ КРОССИНГОВЕР.
Кроссинговер может происходить не только в мейозе, но и

НЕРАВНЫЙ КРОССИНГОВЕР.
Неравный кроссинговер — кроссинговер, в результате которого образуются сестринские кроссоверные
НЕРАВНЫЙ КРОССИНГОВЕР.
Неравный кроссинговер — кроссинговер, в результате которого образуются сестринские кроссоверные
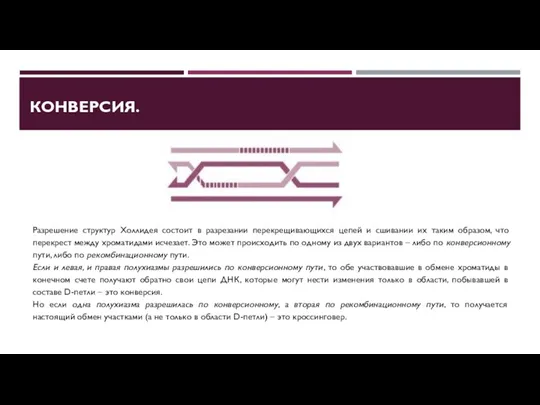
КОНВЕРСИЯ.
Разрешение структур Холлидея состоит в разрезании перекрещивающихся цепей и сшивании их
КОНВЕРСИЯ.
Разрешение структур Холлидея состоит в разрезании перекрещивающихся цепей и сшивании их

НАСЛЕДСТВЕННАЯ ИЗМЕНЧИВОСТЬ.
Изменчивость - один из основных факторов эволюции, является источником для
НАСЛЕДСТВЕННАЯ ИЗМЕНЧИВОСТЬ.
Изменчивость - один из основных факторов эволюции, является источником для

НЕНАСЛЕДСТВЕННАЯ ИЗМЕНЧИВОСТЬ.
Ненаследственная (модификационная изменчивость) — это результат не изменений генотипа, а
НЕНАСЛЕДСТВЕННАЯ ИЗМЕНЧИВОСТЬ.
Ненаследственная (модификационная изменчивость) — это результат не изменений генотипа, а

НОРМА РЕАКЦИИ ПРИЗНАКА.
Норма реакции — спектр возможных уровней экспрессии генов, из
НОРМА РЕАКЦИИ ПРИЗНАКА.
Норма реакции — спектр возможных уровней экспрессии генов, из
 Зеленые водоросли Chlorophyta
Зеленые водоросли Chlorophyta ЕГЭ Биология. Новый формат заданий. 2022
ЕГЭ Биология. Новый формат заданий. 2022 Антиоксидант астаксантин
Антиоксидант астаксантин Кожа. Наружный покров
Кожа. Наружный покров Генетический материал клетки. Генный уровень
Генетический материал клетки. Генный уровень Нуклеїнові кислоти. Класифікація. Будова. Біологічне значення. Нуклеопротеїни
Нуклеїнові кислоти. Класифікація. Будова. Біологічне значення. Нуклеопротеїни Vitamins. Classes of Vitamins
Vitamins. Classes of Vitamins Нуклеиновые кислоты
Нуклеиновые кислоты Урок обобщение по главе Класс птицы
Урок обобщение по главе Класс птицы Кожно-двигательный, зрительный, слуховой,вестибулярный, обонятельный и вкусовой анализаторы
Кожно-двигательный, зрительный, слуховой,вестибулярный, обонятельный и вкусовой анализаторы Птичьи рекорды
Птичьи рекорды Столовые сорта винограда (районированные и перспективные)
Столовые сорта винограда (районированные и перспективные) Охраняемые растения Крыма
Охраняемые растения Крыма Презентация для урока-обобщения по биологии в 5 классе
Презентация для урока-обобщения по биологии в 5 классе Культурні рослини
Культурні рослини Нужны ли минеральные соли животным и человеку
Нужны ли минеральные соли животным и человеку История развития эволюционных идей
История развития эволюционных идей Тестовыя работа по теме: Вирусы
Тестовыя работа по теме: Вирусы Molecular genetic factors controlling plant resistance to diseases
Molecular genetic factors controlling plant resistance to diseases Эпиграф
Эпиграф Биология полорогих: дзерена, сайгака, горала
Биология полорогих: дзерена, сайгака, горала Предмет и задачи токсикологической химии. Взаимосвязь с другими дисциплинами. (Лекция 1)
Предмет и задачи токсикологической химии. Взаимосвязь с другими дисциплинами. (Лекция 1) Становление систематики. 9 класс
Становление систематики. 9 класс Ищите доктора в природе
Ищите доктора в природе Откуда берутся шоколад, изюм и мед
Откуда берутся шоколад, изюм и мед Экзаменационные микропрепараты
Экзаменационные микропрепараты Отряд Пчёлы
Отряд Пчёлы Пищеварение в полости рта: состав и свойства слюны
Пищеварение в полости рта: состав и свойства слюны