Генетика клеточного цикла. Надклеточный контроль клеточного деления, роста и апоптоза. (Глава 5) презентация
- Генетика клеточного цикла. Надклеточный контроль клеточного деления, роста и апоптоза. (Глава 5)

Содержание
- 2. Надклеточный контроль клеточного деления, роста и апоптоза
- 3. В точке контроля G1 р53 блокирует G1-Cdk через белок р21 В точке контроля G2/М р53 блокирует
- 4. Белки GADD в ответе клетки на генотоксический стресс IR ионизирующая UV (MMS) радиация метилметан сульфонат P53
- 5. Надклеточный контроль клеточного деления, роста и апоптоза Размер организма и органа зависит от числа клеток и
- 6. Надклеточный контроль клеточного деления, роста и апоптоза 1. Для поддержания жизни нормальных клеток высших организмов необходимы
- 7. Апоптоз – программируемая гибель клеток Лапа эмбриона мыши с интервалом в сутки, окраска на апоптоз Апоптоз
- 8. Варианты гибели клеток Некроз клетки в клеточной культуре Гибель клетки апоптозом в клеточной культуре Фагоцитоз клетки,
- 9. Апоптоз в трофоцитах бабочки
- 10. Апоптоз в трофоцитах перепончатокрылого
- 11. Апоптоз в сперматоцитах жука, вероятно, вызванный активацией точки контроля
- 12. Апоптоз, морфологические изменения: Конденсация хроматина, фрагментация и разрушение ядра. Цитоскелет сжимается. Клеточная мембрана вспухает. Клетки фрагментируются,
- 13. Каспазы - протеазы, имеющие цистеин в активном сайте и разрезающие белки-мишени по аспарагиновой кислоте – caspases
- 14. Активация апоптоза с внешней стороны через Fas-путь Fas-путь. Лимфоцит-киллер активирует death –рецепторы на поверхности клетки: Fas
- 15. Апоптосома (700 кДа): Активация апоптоза изнутри клетки
- 16. Прокаспаза-3 Один из путей: митохондрии индуцируются к выбросу цитохрома с в цитозоль. Цитохром с присоединяется к
- 17. Митогены (более 50 белков) Фактор роста тромбоцитов PDGF –platelet-derived grows factor Клетки фибробластов в культуре делились
- 18. Один из путей стимуляции клеточных делений митогеном Small GTPasa Ras MAP-киназный каскад Активация транскрипции гена myc
- 19. Small GTPasa Ras MAP-киназный каскад Активация гена myc Гиперактивный Ras- продукт мутантного гена ras часто вызывает
- 20. Ras- мономерная GTF-аза, имеет пренильную группу. Ras-GEF- guanine nucleotide exchange factors Стимуляция митогеном сигнального пути Ras
- 21. Фактор роста тромбоцитов (вирус саркомы обезьян) Рецептор фактора роста эпидермиса (глиобластома, вирус эритробластоза птиц) Рецептор колониестимулирующего
- 22. Стимуляция митогеном сигнального пути Ras MAP- mitogen-activated protein kinase (MAP-киназа). Для активации необходимо фосфорилирование треонина и
- 23. Raf (рак лёгкого, слюнной железы, саркома кошки) MEK у млек. Гиперактивация гена myc – мелкоклеточный рак
- 24. В большинстве нормальных клеток гиперактивация Ras и Myc приводит к активации точки контроля. Нормальная клетка в
- 25. Другие способы регуляции митогенной активности Репликативное старение клеток, связанное с теломерами. У фибробластов через 25-50 делений
- 26. Факторы роста У одноклеточных для роста необходимо только питание. У многоклеточных – ростовой фактор. Увеличение скорости
- 27. Tyr740 Tyr751 Tyr771 Tyr1009 Tyr1021 Рецептор PDGR – фактора роста тромбоцитов, одна из цепей димера Тирозинкиназные
- 28. Многообразие белков, связывающихся с фосфотирозинами. Докинг-сайты для белков, имеющих домены SH2 (src homology region), SH3
- 29. Внеклеточные сигнальные белки могут действовать как ростовой фактор, фактор выживания и митоген одновременно: PDGF –фактор роста
- 30. Возможные превращения PI 3 под действием PI 3-киназы Фосфолипаза С Создание докинг-сайтов для внутриклеточных сигнальных белков
- 31. Если клетки лишены факторов выживания, они активируют программу апоптоза. Нервные клетки образуются в избытке и конкурируют
- 32. Фактор выживания супрессирует апоптоз у млекопитающих Протеинкиназа В Рецептор фактора выживания активирует протеинкиназы, в т.ч. PKB.
- 33. Survivin в апоптозе S.K. Knauer, W. Mann, R.H. Stauber, 2007 А. Survivin в составе комплекса белков
- 34. Некоторые сигнальные белки, действующие через тирозин-киназные рецепторы
- 35. Инсулиноподобный фактор роста IGF-1 Собаки крупных пород имели аллель гена IGF-1, ассоциированный с большей экспрессией, собаки
- 36. Конкуренция клеток за сигнальные белки Клетки в культуре. Явление контактного ингибирования Контактное ингибирование клеточных делений =
- 37. Способность к делению зависит от формы и прикрепления клеток Суспензия Слабая адгезия Хорошая адгезия Возможность вступления
- 38. Способность к делению зависит от формы и прикрепления клеток Фибробласты растут на субстрате, покрытом фибронектином (внеклеточный
- 39. Способность к делению зависит от формы и прикрепления клеток Флуоресцентная окраска: Зеленый- актин, красный- белки, содержащие
- 40. Цитокиновые рецепторы Jak-STAT – сигнальный путь: Интерфероновый рецептор Janus-киназы – цитоплазматические тирозин-киназы STAT – signal transducers
- 41. Цитокиновые рецепторы: Jak-STAT – сигнальный путь Alberts et al., 2002 Янус-киназы имеют 2 киназных домена-цитоплазматические тирозин-киназы,
- 42. Внеклеточные сигналы, ингибирующие рост TGF-β –большое семейство родственных белков. Растворимые димеры, действуют как гормоны или локальные
- 43. Белки семейства TGF β (Трансформирующий фактор роста β) Действуют как гормоны или локальные медиаторы Представитель суперсемейства
- 44. Рецепторы TGF-β : Тип II Тип I Семейство белков TGF-β:
- 45. Путь TGF-β - Smad Стратегия наиболее быстрой передачи сигнала в ядро Рецепторы TGF-β - серин-треонин киназы.
- 46. Участники пути TGF-β – Smad в онкогенезе Рецепторы TGF-β - серин-треонин киназы. Туморсупрессор Рецептор TGF-β II
- 47. Семейство TGFβ: BMP- bone morphogenetic protein Молочная железа мыши: А- железа в при нормальном высасывании; В
- 48. Myostatin– ингибирует пролиферацию миобластов. Семейство белков TGF-β Alberts et al., 2002
- 49. Сигнальный путь Delta – Notch – путь дифференцировки Alberts et al., 2002
- 50. Тирозин протеинкиназы Серин-треонин протеинкиназы Оновные протеинкиназы Alberts et al., 2002
- 51. Эволюционное древо некоторых протеин киназ TGF-β Семейство рецепторных сериновых кинах EGF-рецептор Src Lck
- 52. Овогенез, мейоз и оплодотворение у лягушки Рост овоцита Арест овоцита в профазе мейоза1 Прогестерон Оплодотворение M1,
- 53. Созревание ооцита запускается прогестероном. Он действует через неклассический рецептор на плазматической мембране и вызывает уменьшение уровня
- 54. Роль GPRx (G-protein coupled receptors) в поддержании мейотического ареста Уменьшение уровня cAMP запускает созревание ооцита. Проходит
- 56. Скачать презентацию

Надклеточный контроль клеточного деления, роста и апоптоза
Надклеточный контроль клеточного деления, роста и апоптоза
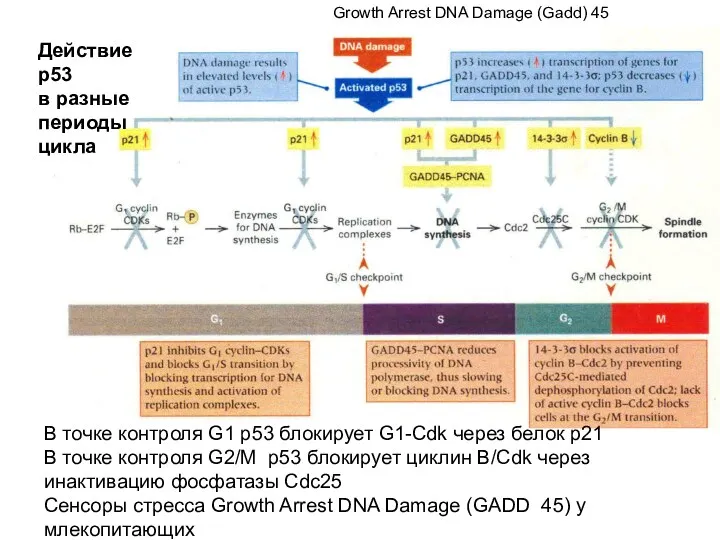
В точке контроля G1 р53 блокирует G1-Cdk через белок р21
В точке
В точке контроля G1 р53 блокирует G1-Cdk через белок р21
В точке
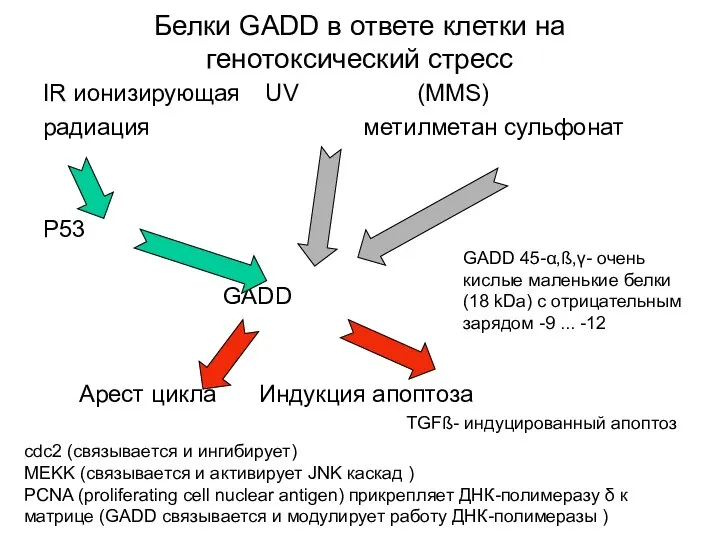
Белки GADD в ответе клетки на генотоксический стресс
IR ионизирующая UV
Белки GADD в ответе клетки на генотоксический стресс
IR ионизирующая UV

Надклеточный контроль клеточного деления, роста и апоптоза
Размер организма и органа зависит
Надклеточный контроль клеточного деления, роста и апоптоза
Размер организма и органа зависит

Надклеточный контроль клеточного деления, роста и апоптоза
1. Для поддержания жизни нормальных
Надклеточный контроль клеточного деления, роста и апоптоза
1. Для поддержания жизни нормальных
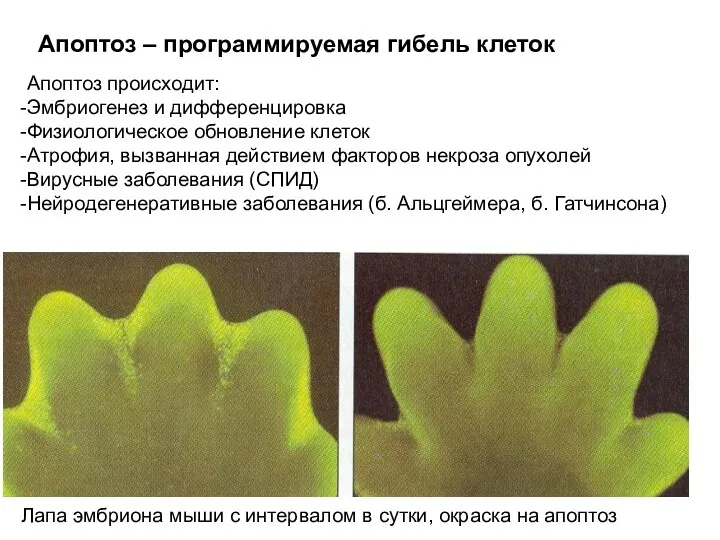
Апоптоз – программируемая гибель клеток
Лапа эмбриона мыши с интервалом в сутки,
Апоптоз – программируемая гибель клеток
Лапа эмбриона мыши с интервалом в сутки,
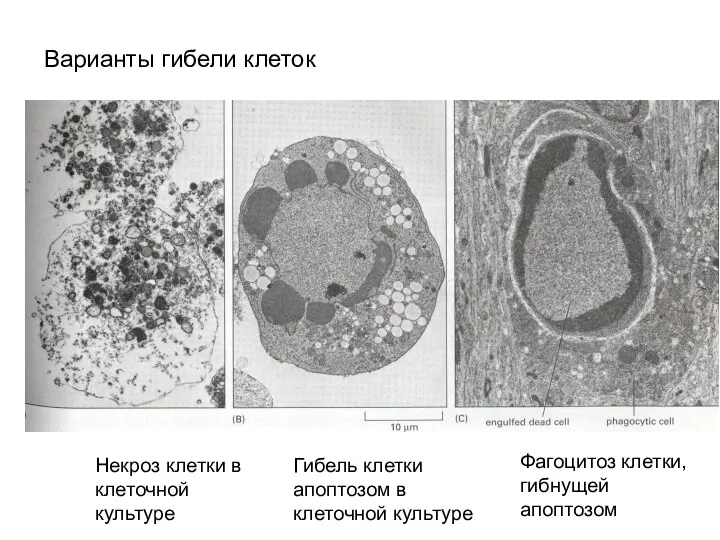
Варианты гибели клеток
Некроз клетки в клеточной культуре
Гибель клетки апоптозом в клеточной
Варианты гибели клеток
Некроз клетки в клеточной культуре
Гибель клетки апоптозом в клеточной

Апоптоз в трофоцитах бабочки
Апоптоз в трофоцитах бабочки
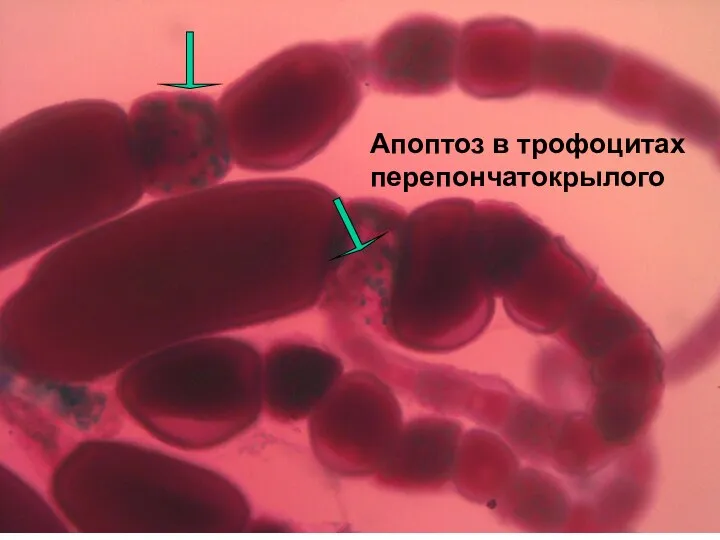
Апоптоз в трофоцитах перепончатокрылого
Апоптоз в трофоцитах перепончатокрылого

Апоптоз в сперматоцитах жука, вероятно, вызванный активацией точки контроля
Апоптоз в сперматоцитах жука, вероятно, вызванный активацией точки контроля

Апоптоз,
морфологические изменения:
Конденсация хроматина, фрагментация и разрушение ядра.
Цитоскелет сжимается.
Клеточная мембрана вспухает.
Клетки
Апоптоз,
морфологические изменения:
Конденсация хроматина, фрагментация и разрушение ядра.
Цитоскелет сжимается.
Клеточная мембрана вспухает.
Клетки
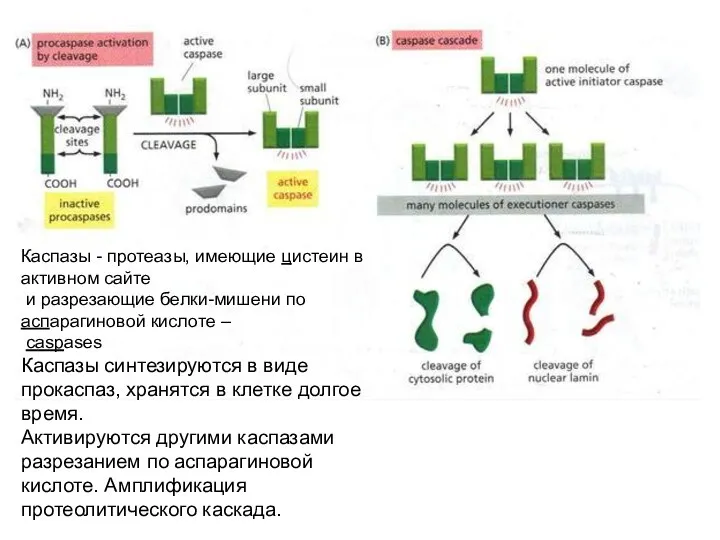
Каспазы - протеазы, имеющие цистеин в активном сайте
и разрезающие белки-мишени
Каспазы - протеазы, имеющие цистеин в активном сайте
и разрезающие белки-мишени
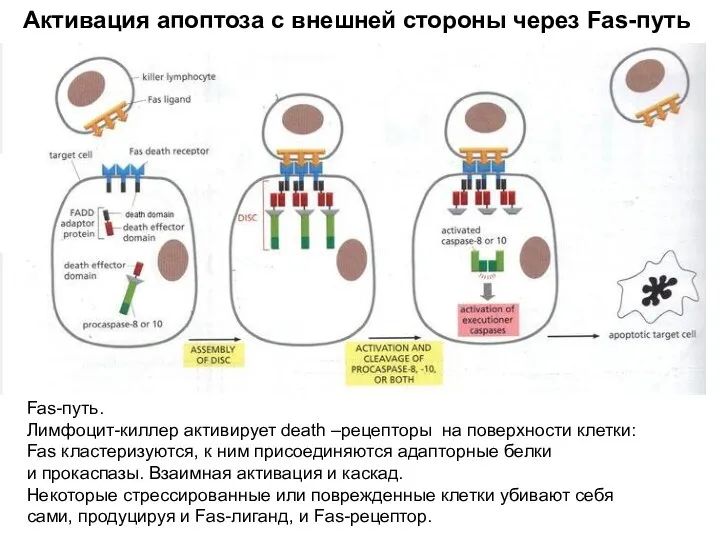
Активация апоптоза с внешней стороны через Fas-путь
Fas-путь.
Лимфоцит-киллер активирует death –рецепторы
Активация апоптоза с внешней стороны через Fas-путь
Fas-путь.
Лимфоцит-киллер активирует death –рецепторы
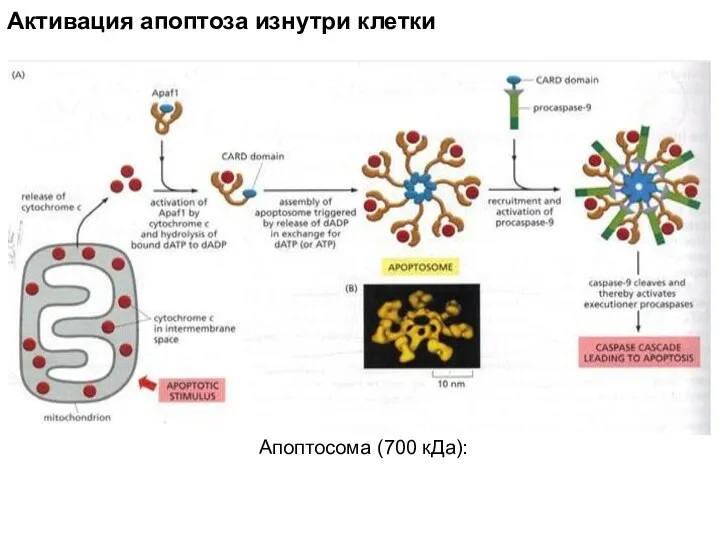
Апоптосома (700 кДа):
Активация апоптоза изнутри клетки
Апоптосома (700 кДа):
Активация апоптоза изнутри клетки
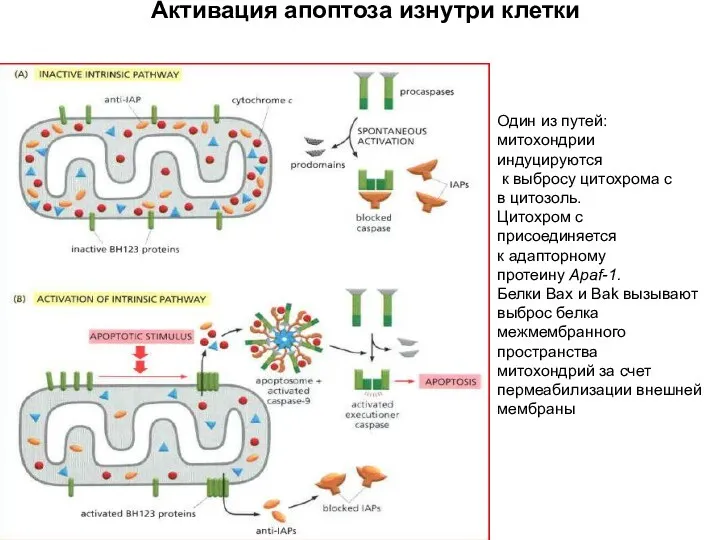
Прокаспаза-3
Один из путей:
митохондрии
индуцируются
к выбросу цитохрома с
в цитозоль.
Прокаспаза-3
Один из путей:
митохондрии
индуцируются
к выбросу цитохрома с
в цитозоль.

Митогены (более 50 белков)
Фактор роста тромбоцитов PDGF –platelet-derived grows factor
Клетки фибробластов
Митогены (более 50 белков)
Фактор роста тромбоцитов PDGF –platelet-derived grows factor
Клетки фибробластов
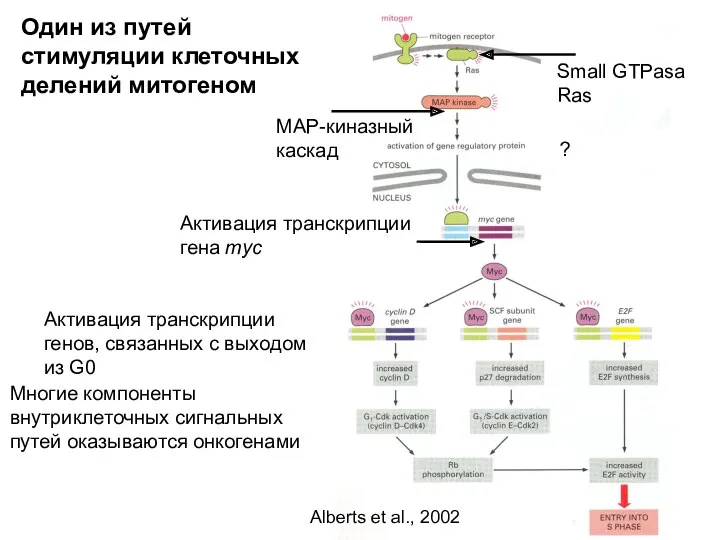
Один из путей стимуляции клеточных делений митогеном
Small GTPasa Ras
MAP-киназный каскад
Активация транскрипции
Один из путей стимуляции клеточных делений митогеном
Small GTPasa Ras
MAP-киназный каскад
Активация транскрипции
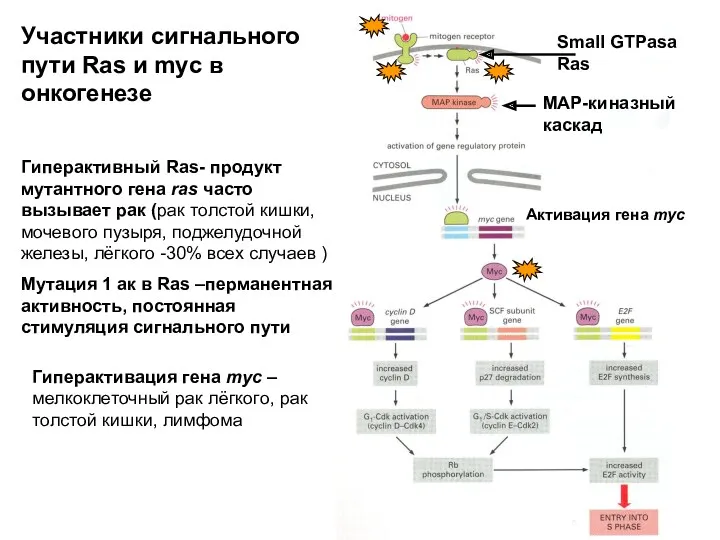
Small GTPasa Ras
MAP-киназный каскад
Активация гена myc
Гиперактивный Ras- продукт мутантного гена ras
Small GTPasa Ras
MAP-киназный каскад
Активация гена myc
Гиперактивный Ras- продукт мутантного гена ras
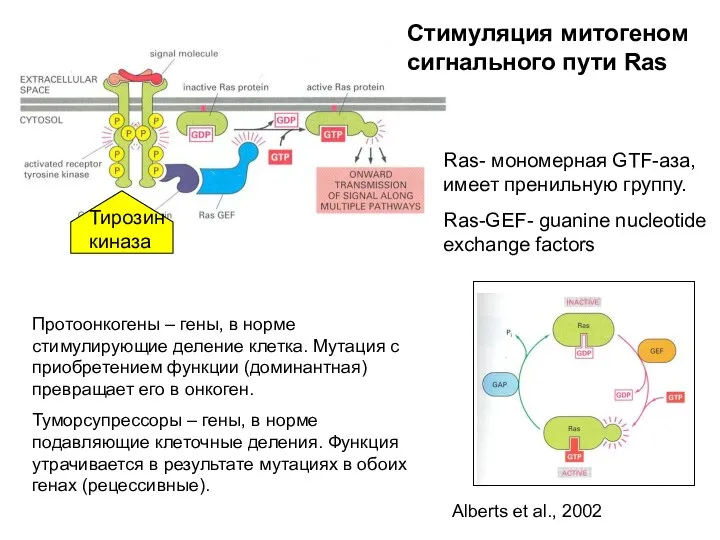
Ras- мономерная GTF-аза, имеет пренильную группу.
Ras-GEF- guanine nucleotide exchange factors
Стимуляция
Ras- мономерная GTF-аза, имеет пренильную группу.
Ras-GEF- guanine nucleotide exchange factors
Стимуляция
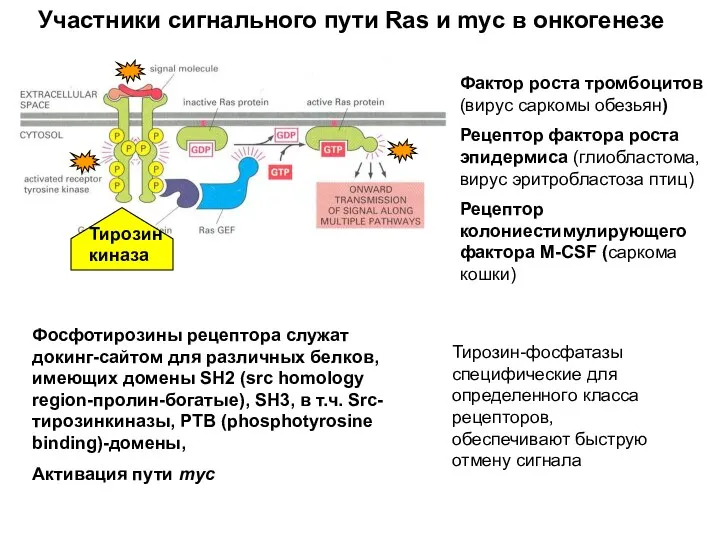
Фактор роста тромбоцитов (вирус саркомы обезьян)
Рецептор фактора роста эпидермиса (глиобластома, вирус
Фактор роста тромбоцитов (вирус саркомы обезьян)
Рецептор фактора роста эпидермиса (глиобластома, вирус
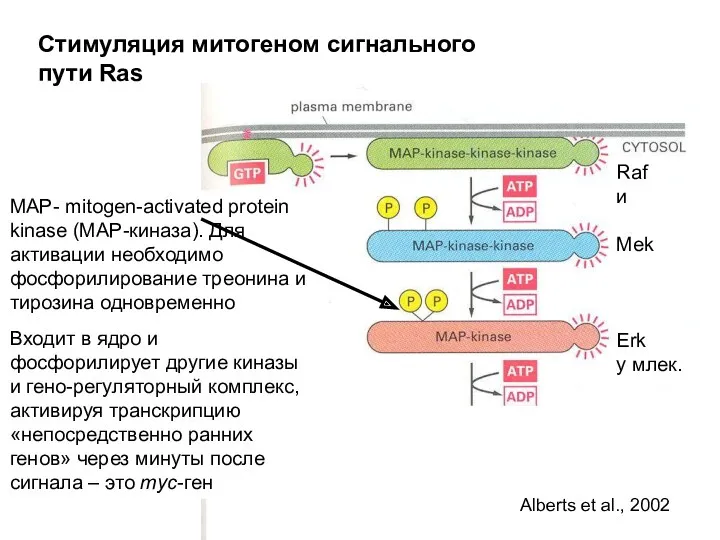
Стимуляция митогеном сигнального пути Ras
MAP- mitogen-activated protein kinase (MAP-киназа). Для активации
Стимуляция митогеном сигнального пути Ras
MAP- mitogen-activated protein kinase (MAP-киназа). Для активации
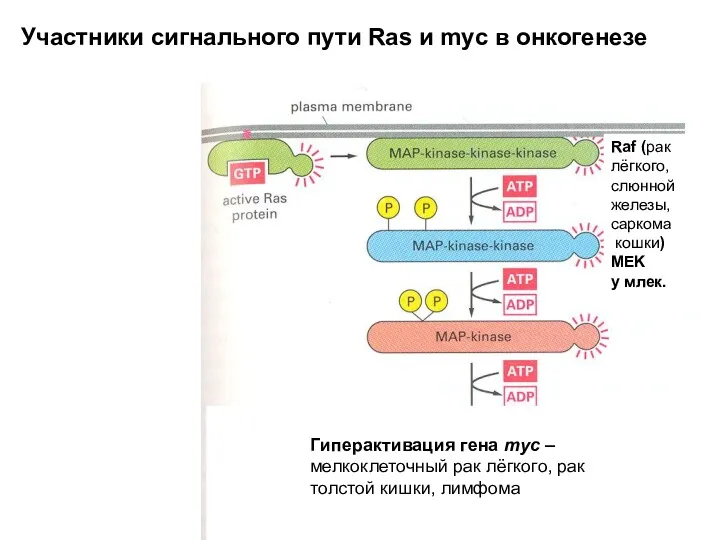
Raf (рак лёгкого, слюнной железы, саркома
кошки)
MEK
у млек.
Гиперактивация гена myc –
Raf (рак лёгкого, слюнной железы, саркома
кошки)
MEK
у млек.
Гиперактивация гена myc –
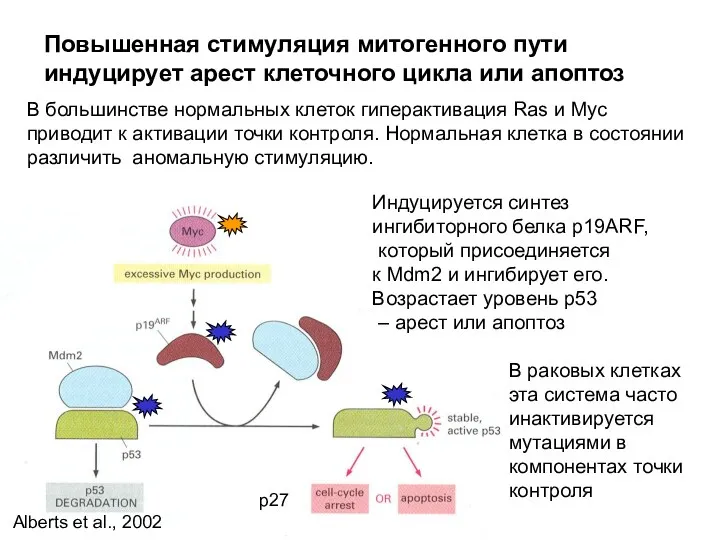
В большинстве нормальных клеток гиперактивация Ras и Myc приводит к активации
В большинстве нормальных клеток гиперактивация Ras и Myc приводит к активации

Другие способы регуляции митогенной активности
Репликативное старение клеток, связанное с теломерами. У
Другие способы регуляции митогенной активности
Репликативное старение клеток, связанное с теломерами. У
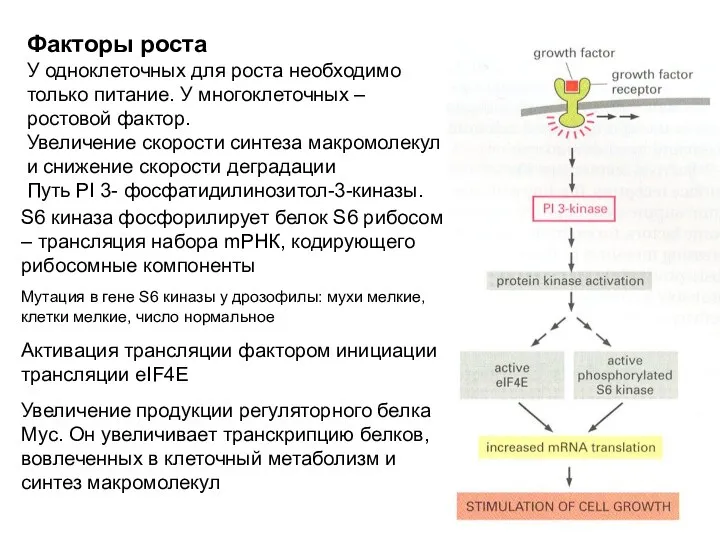
Факторы роста
У одноклеточных для роста необходимо только питание. У многоклеточных –
Факторы роста
У одноклеточных для роста необходимо только питание. У многоклеточных –
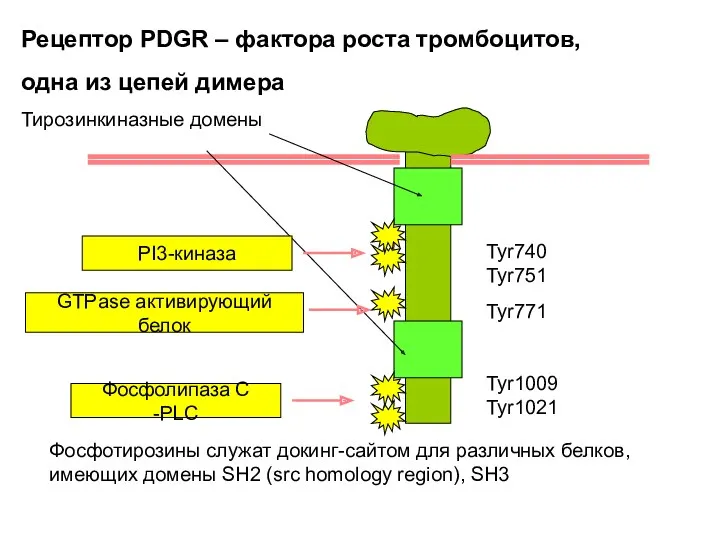
Tyr740 Tyr751
Tyr771
Tyr1009 Tyr1021
Рецептор PDGR – фактора роста тромбоцитов,
одна из цепей димера
Тирозинкиназные
Tyr740 Tyr751
Tyr771
Tyr1009 Tyr1021
Рецептор PDGR – фактора роста тромбоцитов,
одна из цепей димера
Тирозинкиназные
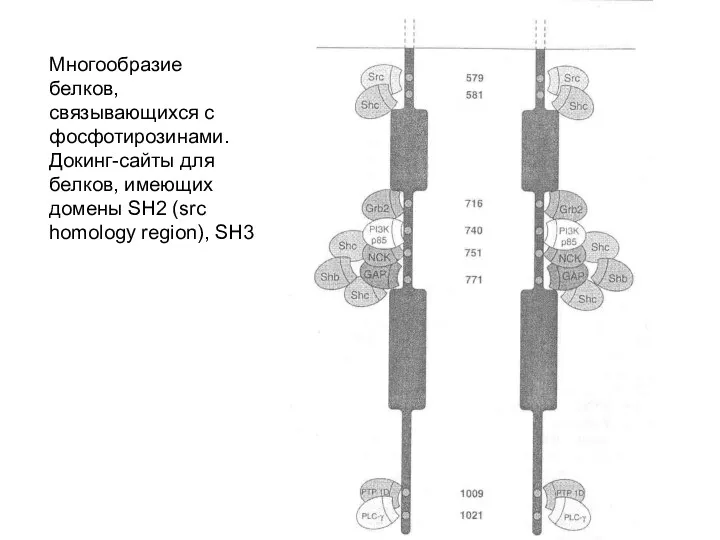
Многообразие белков, связывающихся с фосфотирозинами. Докинг-сайты для белков, имеющих домены SH2
Многообразие белков, связывающихся с фосфотирозинами. Докинг-сайты для белков, имеющих домены SH2
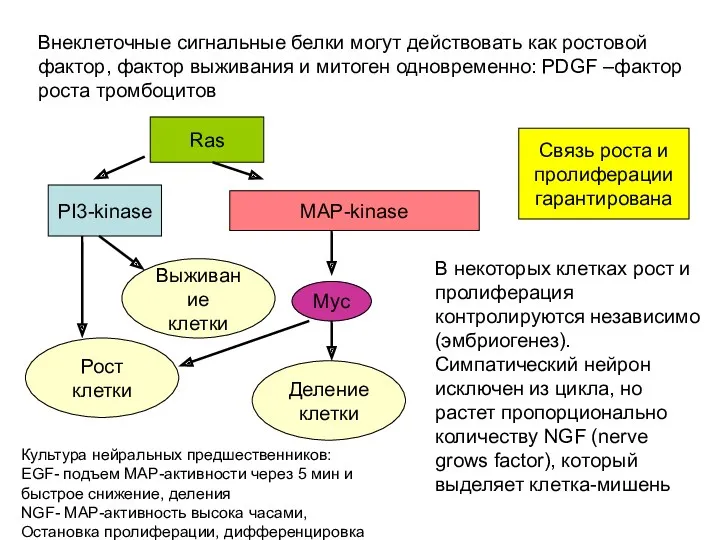
Внеклеточные сигнальные белки могут действовать как ростовой фактор, фактор выживания и
Внеклеточные сигнальные белки могут действовать как ростовой фактор, фактор выживания и
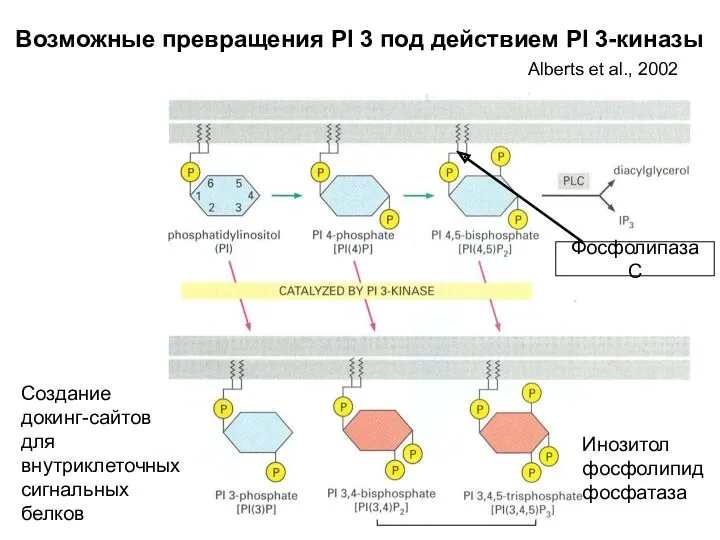
Возможные превращения PI 3 под действием PI 3-киназы
Фосфолипаза С
Создание докинг-сайтов для
Возможные превращения PI 3 под действием PI 3-киназы
Фосфолипаза С
Создание докинг-сайтов для
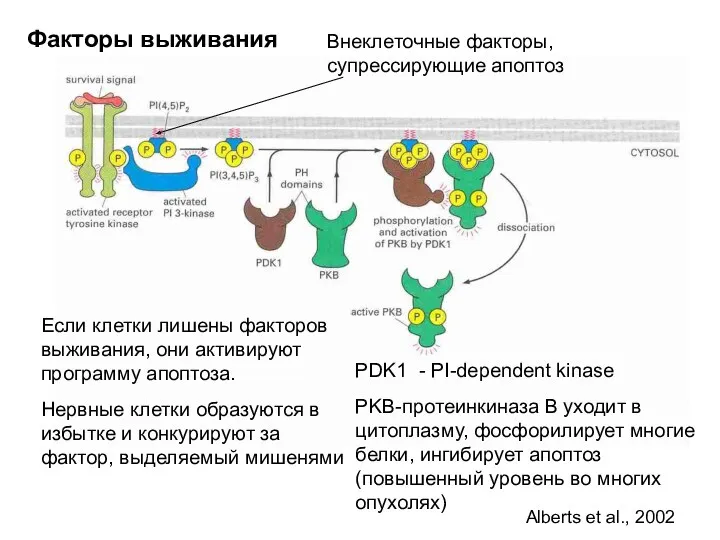
Если клетки лишены факторов выживания, они активируют программу апоптоза.
Нервные клетки образуются
Если клетки лишены факторов выживания, они активируют программу апоптоза.
Нервные клетки образуются
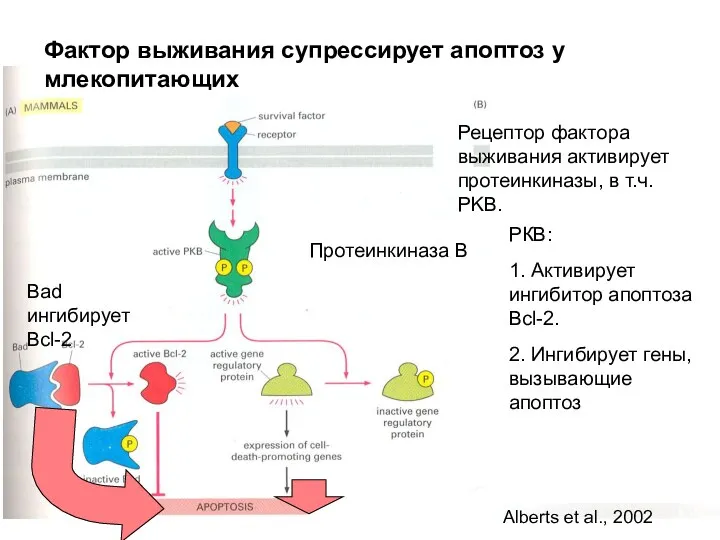
Фактор выживания супрессирует апоптоз у млекопитающих
Протеинкиназа В
Рецептор фактора выживания активирует протеинкиназы,
Фактор выживания супрессирует апоптоз у млекопитающих
Протеинкиназа В
Рецептор фактора выживания активирует протеинкиназы,
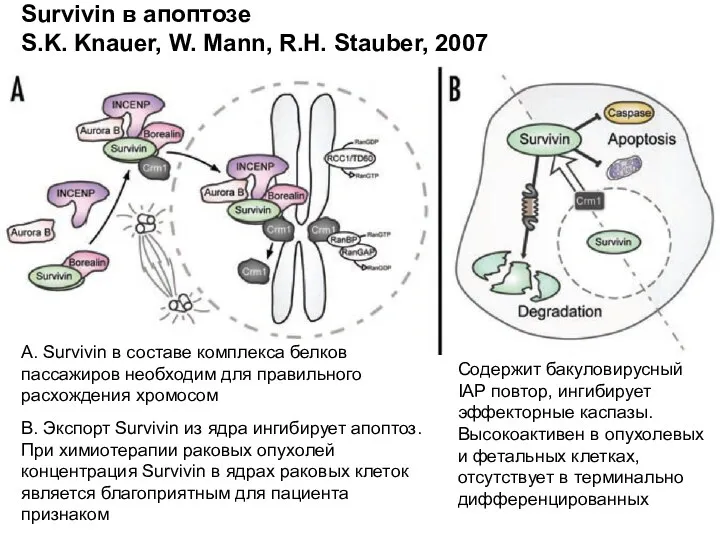
Survivin в апоптозе
S.K. Knauer, W. Mann, R.H. Stauber, 2007
А. Survivin
Survivin в апоптозе
S.K. Knauer, W. Mann, R.H. Stauber, 2007
А. Survivin
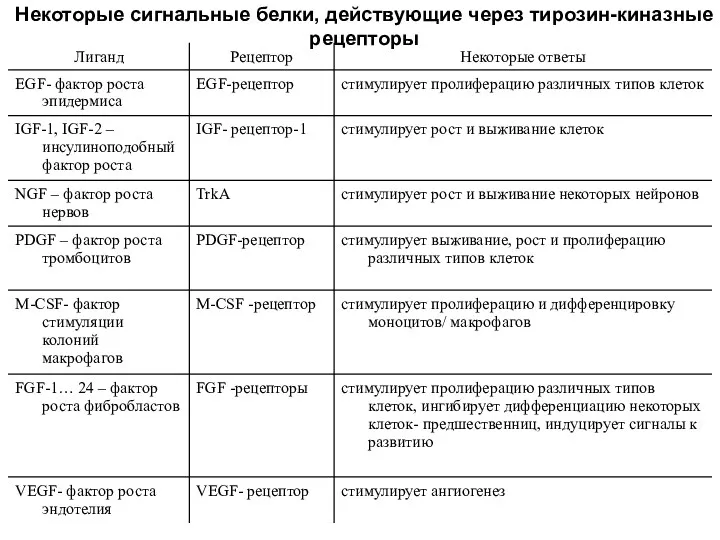
Некоторые сигнальные белки, действующие через тирозин-киназные рецепторы
Некоторые сигнальные белки, действующие через тирозин-киназные рецепторы

Инсулиноподобный фактор роста IGF-1
Собаки крупных пород имели аллель гена IGF-1, ассоциированный
Инсулиноподобный фактор роста IGF-1
Собаки крупных пород имели аллель гена IGF-1, ассоциированный
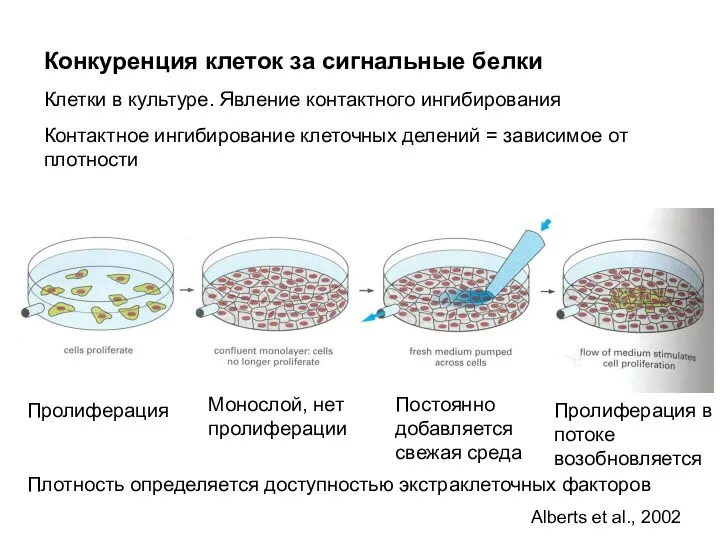
Конкуренция клеток за сигнальные белки
Клетки в культуре. Явление контактного ингибирования
Контактное
Конкуренция клеток за сигнальные белки
Клетки в культуре. Явление контактного ингибирования
Контактное
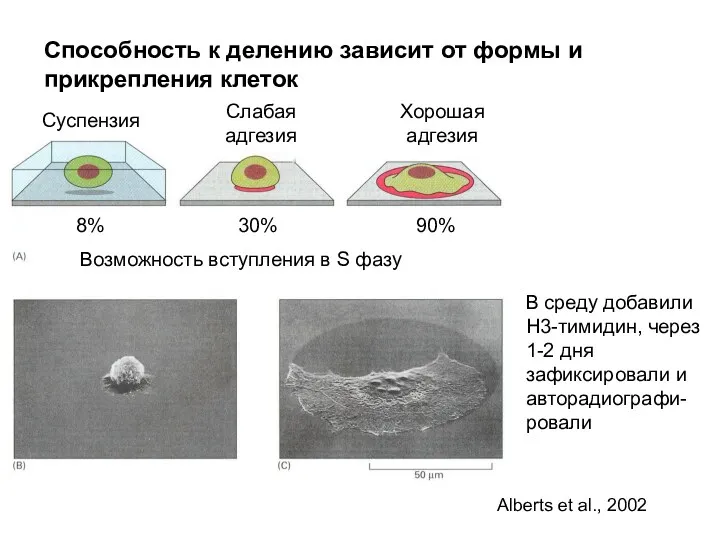
Способность к делению зависит от формы и прикрепления клеток
Суспензия
Слабая адгезия
Хорошая адгезия
Возможность
Способность к делению зависит от формы и прикрепления клеток
Суспензия
Слабая адгезия
Хорошая адгезия
Возможность
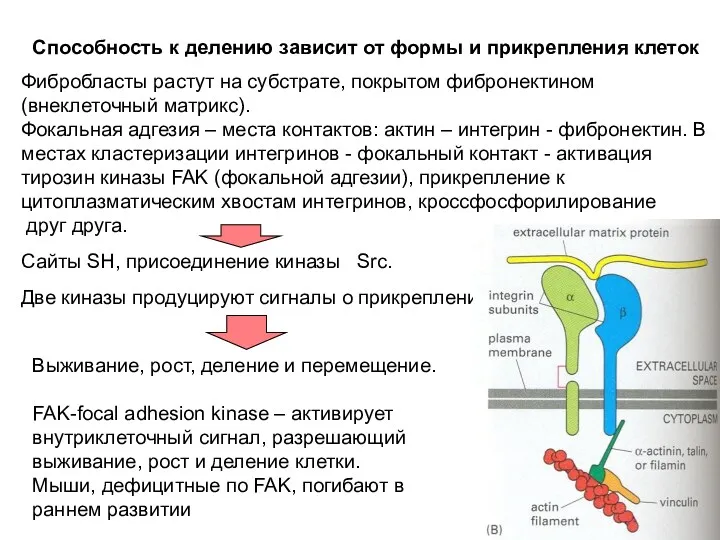
Способность к делению зависит от формы и прикрепления клеток
Фибробласты растут на
Способность к делению зависит от формы и прикрепления клеток
Фибробласты растут на
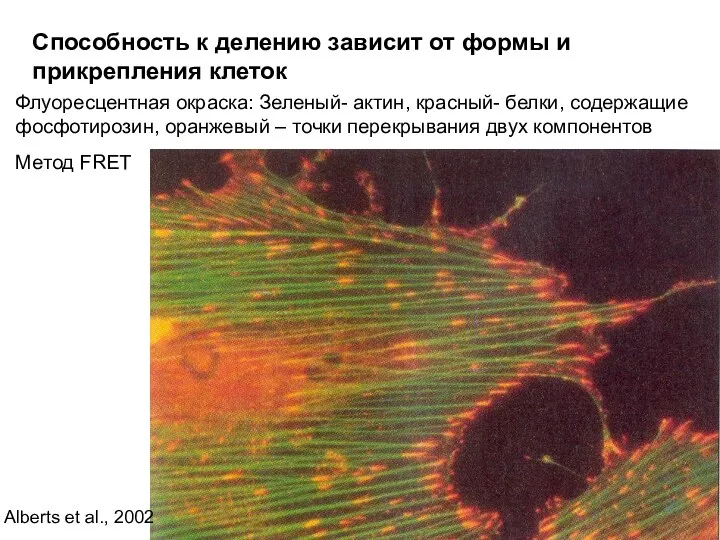
Способность к делению зависит от формы и прикрепления клеток
Флуоресцентная окраска: Зеленый-
Способность к делению зависит от формы и прикрепления клеток
Флуоресцентная окраска: Зеленый-

Цитокиновые рецепторы
Jak-STAT – сигнальный путь:
Интерфероновый рецептор
Janus-киназы – цитоплазматические тирозин-киназы
STAT – signal
Цитокиновые рецепторы
Jak-STAT – сигнальный путь:
Интерфероновый рецептор
Janus-киназы – цитоплазматические тирозин-киназы
STAT – signal
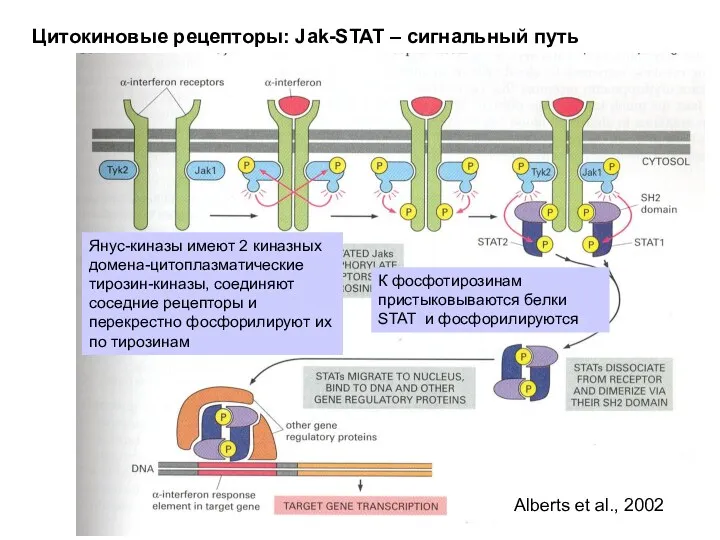
Цитокиновые рецепторы: Jak-STAT – сигнальный путь
Alberts et al., 2002
Янус-киназы имеют 2
Цитокиновые рецепторы: Jak-STAT – сигнальный путь
Alberts et al., 2002
Янус-киназы имеют 2

Внеклеточные сигналы, ингибирующие рост
TGF-β –большое семейство родственных белков. Растворимые димеры, действуют
Внеклеточные сигналы, ингибирующие рост
TGF-β –большое семейство родственных белков. Растворимые димеры, действуют

Белки семейства TGF β (Трансформирующий фактор роста β) Действуют как гормоны
Белки семейства TGF β (Трансформирующий фактор роста β) Действуют как гормоны

Рецепторы TGF-β :
Тип II
Тип I
Семейство белков TGF-β:
Рецепторы TGF-β :
Тип II
Тип I
Семейство белков TGF-β:
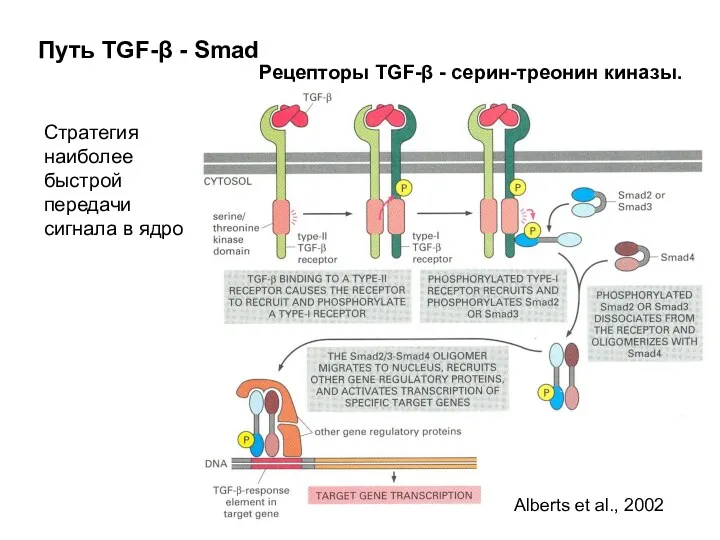
Путь TGF-β - Smad
Стратегия наиболее быстрой передачи сигнала в ядро
Рецепторы
Путь TGF-β - Smad
Стратегия наиболее быстрой передачи сигнала в ядро
Рецепторы
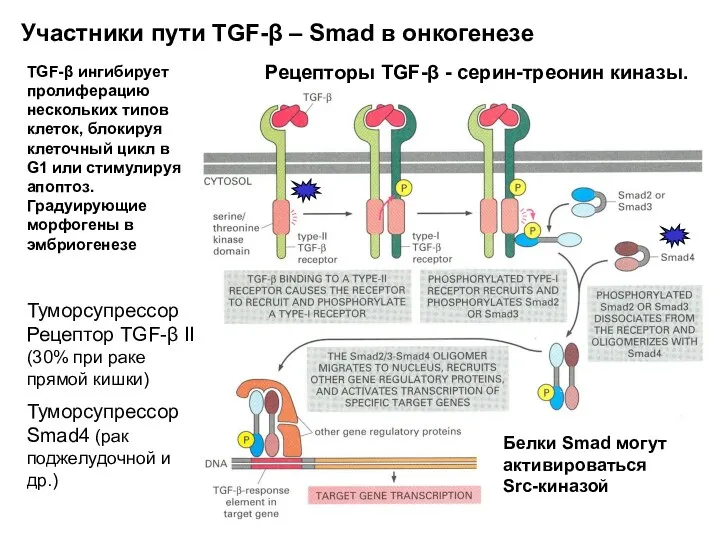
Участники пути TGF-β – Smad в онкогенезе
Рецепторы TGF-β - серин-треонин
Участники пути TGF-β – Smad в онкогенезе
Рецепторы TGF-β - серин-треонин
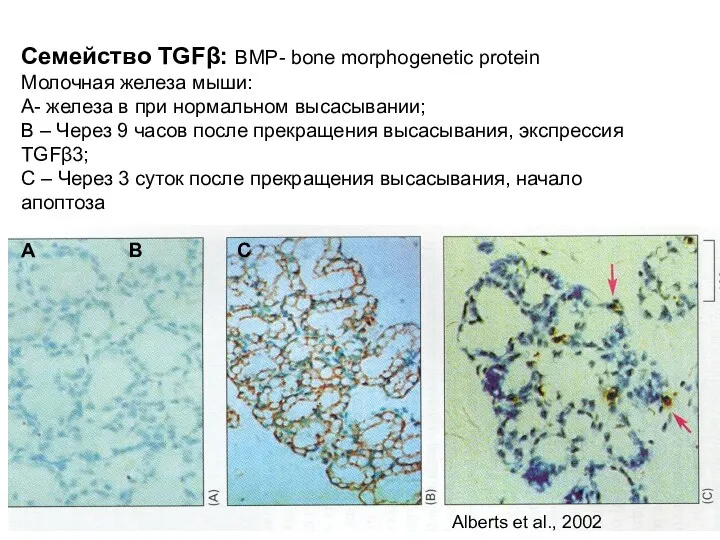
Семейство TGFβ: BMP- bone morphogenetic protein
Молочная железа мыши:
А- железа в
Семейство TGFβ: BMP- bone morphogenetic protein
Молочная железа мыши:
А- железа в
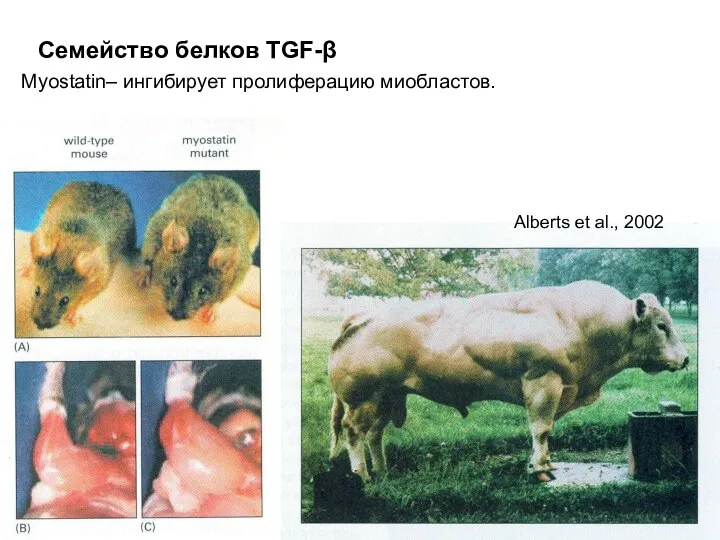
Myostatin– ингибирует пролиферацию миобластов.
Семейство белков TGF-β
Alberts et al., 2002
Myostatin– ингибирует пролиферацию миобластов.
Семейство белков TGF-β
Alberts et al., 2002
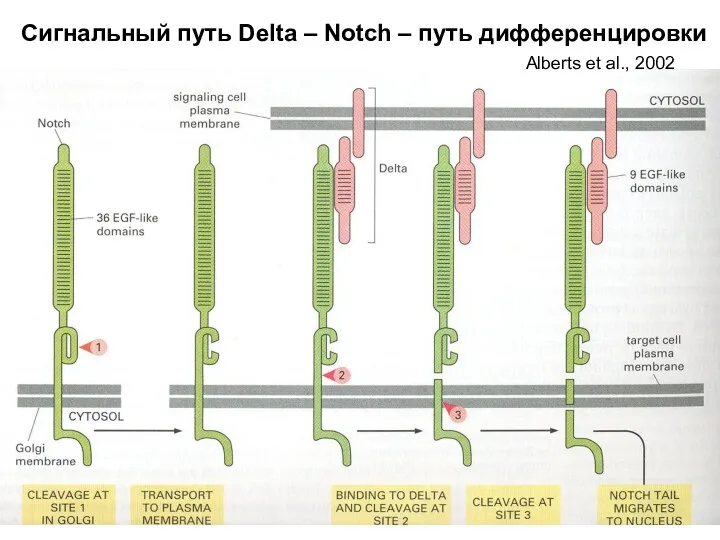
Сигнальный путь Delta – Notch – путь дифференцировки
Alberts et al., 2002
Сигнальный путь Delta – Notch – путь дифференцировки
Alberts et al., 2002
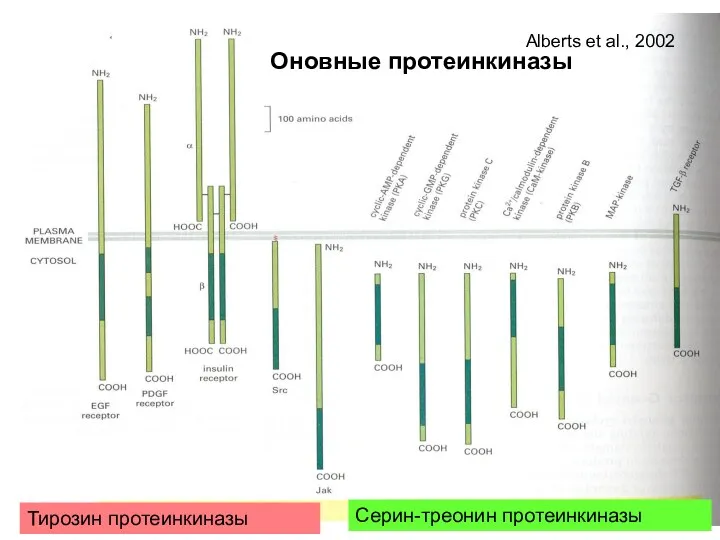
Тирозин протеинкиназы
Серин-треонин протеинкиназы
Оновные протеинкиназы
Alberts et al., 2002
Тирозин протеинкиназы
Серин-треонин протеинкиназы
Оновные протеинкиназы
Alberts et al., 2002
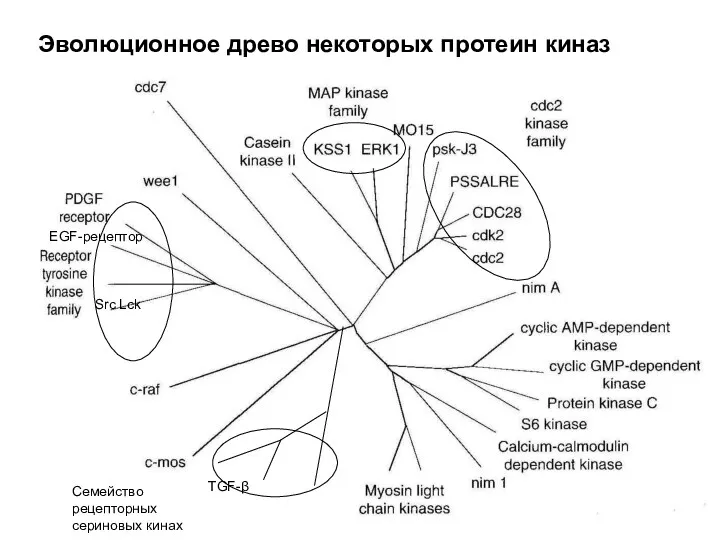
Эволюционное древо некоторых протеин киназ
TGF-β
Семейство рецепторных сериновых кинах
EGF-рецептор
Src Lck
Эволюционное древо некоторых протеин киназ
TGF-β
Семейство рецепторных сериновых кинах
EGF-рецептор
Src Lck
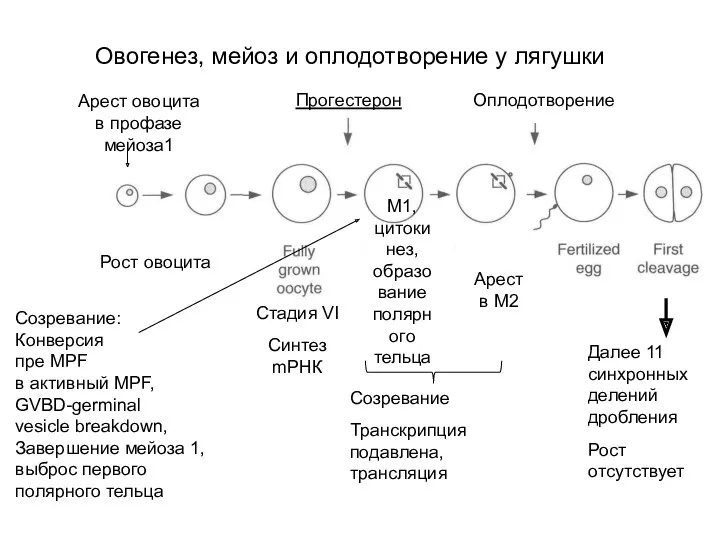
Овогенез, мейоз и оплодотворение у лягушки
Рост овоцита
Арест овоцита
в профазе мейоза1
Прогестерон
Оплодотворение
M1,
цитокинез,
образование
полярного
тельца
Арест
Овогенез, мейоз и оплодотворение у лягушки
Рост овоцита
Арест овоцита
в профазе мейоза1
Прогестерон
Оплодотворение
M1,
цитокинез,
образование
полярного
тельца
Арест

Созревание ооцита запускается прогестероном. Он действует через неклассический рецептор на плазматической
Созревание ооцита запускается прогестероном. Он действует через неклассический рецептор на плазматической
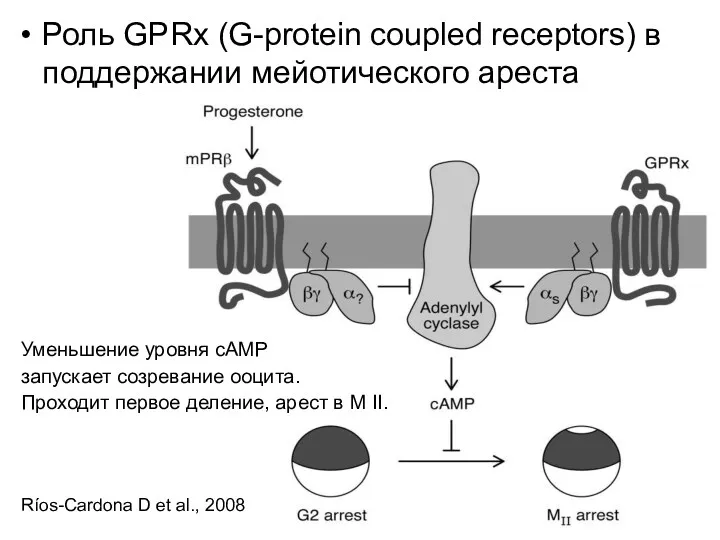
Роль GPRx (G-protein coupled receptors) в поддержании мейотического ареста
Уменьшение уровня cAMP
Роль GPRx (G-protein coupled receptors) в поддержании мейотического ареста
Уменьшение уровня cAMP
 Plant macrofossil
Plant macrofossil Нуклеиновые кислоты: состав, строение, функции. Биология. 10 класс
Нуклеиновые кислоты: состав, строение, функции. Биология. 10 класс Энергетический обмен, катаболизм
Энергетический обмен, катаболизм Птицы. Внешнее строение птиц. Систематические группы птиц
Птицы. Внешнее строение птиц. Систематические группы птиц Жыландар. Жылан шағуынан қорғану және жылан шаққанда көрсетілетін алғашқы көмек
Жыландар. Жылан шағуынан қорғану және жылан шаққанда көрсетілетін алғашқы көмек Protein structure and function
Protein structure and function Мир птиц
Мир птиц Видоизменения побегов
Видоизменения побегов Значение работ Менделя, Моргана и Кольцова для развития генетики
Значение работ Менделя, Моргана и Кольцова для развития генетики Рослини України, які походять з інших материків і частин Євразії
Рослини України, які походять з інших материків і частин Євразії Энзимопатия. Энзимотерапия. Энзимодиагностика
Энзимопатия. Энзимотерапия. Энзимодиагностика Водоросли. Особенности строения, питания, размножения
Водоросли. Особенности строения, питания, размножения Моногенне та полігенне успадкування ознак у людини.Позахромосомна (цитоплазматична) спадковість у людини
Моногенне та полігенне успадкування ознак у людини.Позахромосомна (цитоплазматична) спадковість у людини Взаимодействие аллельных и неаллельных генов
Взаимодействие аллельных и неаллельных генов Уход за морской свинкой
Уход за морской свинкой Деревья и кустарники
Деревья и кустарники Генетика микроорганизмов. Рекомбинации у бактерий
Генетика микроорганизмов. Рекомбинации у бактерий Рост и развитие волос
Рост и развитие волос Роль и функции в организме основных водорастворимых витаминов: витаминов В6 и В12
Роль и функции в организме основных водорастворимых витаминов: витаминов В6 и В12 Здоровый человек - здоровое общество
Здоровый человек - здоровое общество Презентация по биологии 7 класс Отряды насекомых
Презентация по биологии 7 класс Отряды насекомых Ячмінь. Господарське значення та поширення
Ячмінь. Господарське значення та поширення Ветеринарно-бактериологический надзор в животноводстве
Ветеринарно-бактериологический надзор в животноводстве Выращивание томатов в теплице
Выращивание томатов в теплице Царство Животные
Царство Животные Факты о генетике
Факты о генетике Углеводы, виды, значение в питании, гигиенические нормы, источники
Углеводы, виды, значение в питании, гигиенические нормы, источники Методы биологических исследований
Методы биологических исследований