- Генетика микроорганизмов. Биотехнология

Содержание
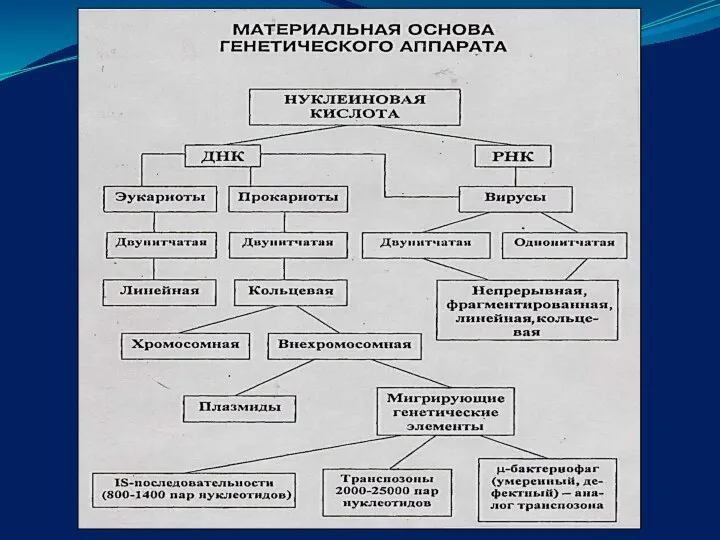
- 2. Генетика- наука о наследственности и изменчивости Ген- фрагмент молекулы ДНК, контролирующий один признак или пептид .
- 3. Особенности бактерий как генетического объекта микроорганизмы имеют малые размеры, высокую скорость размножения, легко культивируются в искусственных
- 5. Отличительные особенности организации генома прокариот Высокое абсолютное число генов. Относительно высокое (70%) содержание структурных генов на
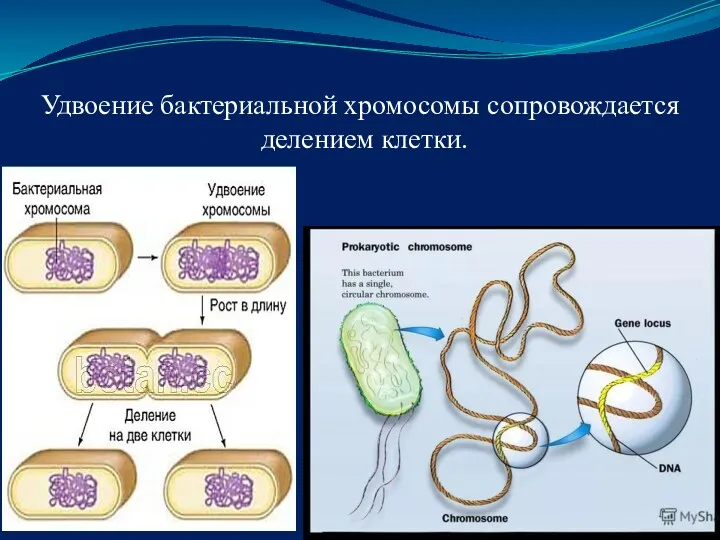
- 6. Удвоение бактериальной хромосомы сопровождается делением клетки.
- 7. Этапы репликации ДНК 3 этапа: инициация, элонгация и терминация. Репликация начинается в определенной точке ori (от
- 8. Одна из цепей достраивается последовательно. Другая - достраивается ступенчато, посегментно, фрагментами (Оказаки) по 1-2 тыс. нуклеотидов,
- 9. Синтез каждого фрагмента на отстающей цепи идет с участием ДНК-праймазы и затравочной РНК. Затравочная РНК -
- 10. Пространственная организация участка прикрепления ДНК в зоне роста цитоплазматической мембраны и клеточной стенки обеспечивают растаскивание двух
- 11. Структурно-функциональная единица ДНК Основной структурно-функциональной единицей ДНК является оперон. Оперон включает в себя группу структурных генов
- 12. Регуляторные элементы: Энхансер - генетический элемент, усиливающий транскрипцию оперона Аттенуатор - генетический элемент, ослабляющий работу оперона.
- 13. Организация работы оперона В обычных условиях ген-регулятор активен и в клетке наблюдается синтез белка-репрессора. Белок-репрессор имеет
- 15. Внехромосомные генетические элементы Представлены: плазмидами, транспозонами, вставочными элементами (инсерционные) умеренными или дефектными фагами. Подвижные генетические элементы
- 16. Плазмиды подвижные генетические элементы, представленные замкнутой ДНК. Могут быть несвязанными с бактериальной хромосомой (автономные) или встроенными
- 17. Функции плазмид регуляторная кодирующая Регуляторная функция состоит в компенсации нарушений метаболизма ДНК Кодирующая функция состоит во
- 18. Плазмиды широко распространены у микроорганизмов. Они могут передаваться внутри вида, между видами и даже между родами.
- 19. Классификация плазмид по свойствам, которыми они наделяет своих носителей
- 20. Транспозоны – нуклеотидные последовательности от 2000- до 20 000 пар нуклеотидов, несут дополнительную генетическую информацию, и
- 21. Транспозоны могут находиться в свободном состоянии в виде кольцевой молекулы, неспособной к самостоятельной репликации и быть
- 22. Транспозоны могут выполнять регуляторную кодирующую индуцируют мутации Основной механизм перемещения - вырезание/ встраивание. Имеют особые концевые
- 23. Вставочные элементы (IS-последовательности) представляют собой мигрирующие элементы величиной от 800-1400 пар оснований. IS- последовательности содержат информацию
- 24. Функции IS- последовательностей: координируют взаимодействие транспозонов, плазмид и умеренных фагов, как между собой, так и с
- 26. А. Внедрение IS-элемента в геном; Б. Распространение IS-элемента в геноме за счет транспозиции; В. Реципрокные транслокации;
- 27. Умеренные и дефектные фаги Встраиваясь в хромосому бактерии, фаги вызывают ее лизогенизацию, в результате чего бактерия
- 28. Изменчивость микроорганизмов Фенотипическая Генотипическая
- 29. Наследственная изменчивость связана с изменением последовательности нуклеотидов в ДНК, полной или частичной их утратой, структурной перестройкой
- 30. Спонтанные мутации проявляются в популяции в естественных условиях под влиянием невыясненных причин. К их появлению приводят
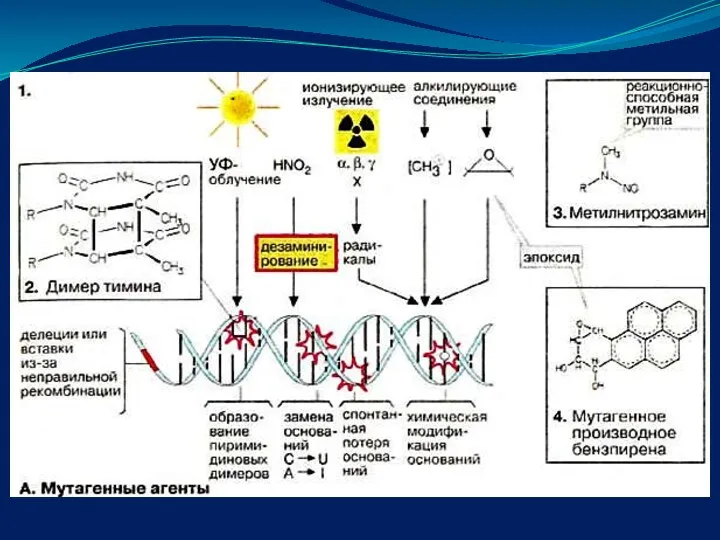
- 31. Индуцированные мутации возникают под влиянием конкретного события или воздействия. Мутагены - вещества вызвавшие мутацию. Мутагены могут
- 33. Типы мутаций: Мутация со сдвигом считывания, вставки или выпадения азотистых оснований. Хромосомные и генные мутации По
- 34. S – R – диссоциации Возникают после встраивания внехромосомных факторов наследования в бактериальную хромосому. Образуется две
- 35. Репарации -специальные системы, восстанавливающие повреждения генетического материала. Направления коррекции повреждений ДНК: Реверсия от поврежденной ДНК к
- 37. Генетические рекомбинации Обмен генетическим материалом между отдельными особями в популяции клеток В процессе генетического переноса участвуют
- 38. Рекомбинация Законная Требует наличия протяженных комплементарных участков ДНК в рекомбинируемых молекулах Происходит только между близкородственными видами
- 39. общая или гомологическая рекомбинация, когда в структуре взаимодействующей ДНК есть гомологические участки; Сайт – специфическая рекомбинация.
- 40. Механизмы генетических рекомбинаций Трансформация Трансдукция Коньюгация
- 41. Трансформация перенос генетического материала клетки донора, при котором реципиент захватывает из внешней среды фрагменты чужеродной ДНК.
- 42. Индуцированная трансформация происходит при добавлении к культуре бактерий очищенной ДНК, признаки которых стремятся передать исследуемой культуре.
- 43. Фазы трансформации: 1) адсорбция ДНК - донора на клетке реципиента; 2) проникновение ДНК внутрь клетки-реципиента; 3)
- 44. Трансдукция передача ДНК от бактерии - донора к бактерии – реципиенту при участии бактериофага. Различают специфическую,
- 45. Специфическая – перенос определенного фрагмента ДНК донора, только в определенные участки ДНК реципиента. Неспецифическая - случайный
- 46. Абортивная трансдукция, когда фрагмент ДНК, привнесенный фагом не вступает в рекомбинацию и не реплицируется, но с
- 47. Конъюгация перенос генетического материала из клетки в клетку при их непосредственном взаимодействии. Донорами являются клетки, несущие
- 48. При взаимодействии F “+” клетки с F “-” клеткой половой фактор передается независимо от хромосомы донора,
- 49. Первый этап конъюгации – прикреплении клетки-донора к реципиентной клетке с помощью половых ворсинок (sex pili). Образование
- 50. Генетика вирусов Характеристика вирусных популяций Высокая численность популяции увеличивает вероятность мутаций Быстрая смена поколений Гаплоидность и
- 51. Ненаследуемые изменения у вирусов связаны с особенностями клетки хозяина и проявляются изменением химического состава суперкапсида, в
- 52. Фенотипическое смешивание при смешанном заражении клеток несколькими вирусами, если часть потомства одного вируса приобретает свойства обоих
- 53. Мутации У вирусов возникают во время репликации их нуклеиновых кислот. Мутанты вирусов фенотипически различаются по строению
- 54. Вирусы способны и к генетическим рекомбинациям. При одновременном заражении двумя вирусами чувствительной клетки - хозяина их
- 55. Генетическая реактивация - перераспределение генов, когда у родственных вирусов инактивированы разные гены. При их взаимодействии могут
- 57. Комплементация когда белки кодируемые геном одного вируса, способствуют репродукции другого вируса. Функциональное взаимодействие двух дефектных вирусов.
- 58. Интерферирующие взаимодействия Состояние невосприимчивости к вторичному заражению клетки уже инфицированной вирусами Интерференция может быть: Гетерологической гомологической
- 59. Гетерологическая интерференция. Инфицирование одним вирусом полностью блокирует возможность репликации другого вируса в пределах одной клетки. Связан
- 60. Биотехнология Bios- жизнь, tecen- искусство, logos- наука Термин БИОТЕХHОЛОГИЯ впервые использовал К. Эреки в 1919 г.
- 61. H а п р а в л е н и я б и о т е
- 62. Медицинская биотехнология Иммунобиологическая биотехнология изучает способы и методы конструирования, биотехнологию получения, стандартизации и оценки свойств иммунобиологических
- 63. Иммуногенетическая биотехнология связана с производством вакцин нового поколения для профилактики инфекционных заболеваний человека и животных. Это
- 64. Объект исследования Биообъект - Целостный сохранивший жизнеспособность многоклеточный или одноклеточный организм, либо изолированные клетки и мультиферментные
- 65. Б и о о б ъ е к т ы: Микроорганизмы: бактерии, вирусы, дрожжи; одноклеточные организмы
- 66. Биообъект, осуществляющий полный синтез целевого продукта, называется продуцентом.
- 67. Бактерии - продуценты представители родов: Acetobaсter - превращают этанол в уксусную кислоту; Bacillus - получение ферментов
- 68. Клетки животных или растений. Из них извлекают широкий ассортимент сложной и ценной продукции (алкалоиды, противо-воспалительные вещества,
- 69. Типы биотехнологических производств Использование живой или инактивированной биомассы (закваски, получение пекарских дрожжей, эубиотиков и др.); Получение
- 70. Прогресс биотехнологии в дальнейшем будет определяться развитием генной, клеточной и эмбриогенетической инженерии.
- 71. Генная инженерия- совокупность приемов и методов связанных с целенаправленным конструированием in vitro новых комбинаций генетического материала,
- 72. Механизмы конструирования рекомбинантной ДНК Обработка ферментами- рестриктазами (более 200 ферментов) донорной и реципиентной ДHК (гидролиз) в
- 73. Трансгенезом называется встраивание чужих генов растениям и животным. генетически модифицированные источники пищи (трансгенные соя, рапс, кукуруза,
- 74. Трансгенные животные Животные в геном которых интегрируют чужеродные гены, называют трансгенными. В 1980 г. Дж.Гордон с
- 75. В клетки животных гены вводятся: при добавлении в среду для культивирования; В виде микроинъекций в оплодотворенную
- 76. Клеточная инженерия- метод конструирования клеток нового типа на основе их культивирования, гибридизации и реконструкции (гибридомная технология)
- 77. Гибридомная технология Важнейшим этапом в развитии биотехнологии стало создание гибридомы (Д.Келлер, Д.Мистейн – 1975 г. Нобелевская
- 78. Основана на получении гибридных клеток В- лимфоцитов, стимулированных конкретным антигеном Миелоидных (опухолевых клеток), способных к неограниченному
- 79. Моноклональная технология применяется: Для диагностики: - инфекционных заболеваний - аутоимунных заболеваний - опухолевых заболеваний - неинфекционных
- 81. стволовые клетки Открыты в 1981 г. М. Эвансом (у мышей). В 1998 г. Дж.Томпсон и Д.Беккер
- 82. Генетическая информация в стволовой клетке находится в «нулевой точке» отсчета. Клетка еще не имеет специализации и
- 83. Биопринтинг- технология трехмерной биопечати органов из аутологичных клеток.
- 84. Эмбриогенетическая инженерия Перестройка генома- реконструкция эмбрионов путем клонирования. генно-инженерные методы радикального лечения наследственных болезней.
- 85. Типы генотерапии ex vivo- пораженные клетки выделяют из организма человека, инкубируют с вектором, а затем вновь
- 86. Баллистическая трансфекция- основана на обстреле органов и тканей частицами тяжелых металлов (золото, вольфрам), покрытых плазмидной ДНК.
- 87. Избирательная инактивация гена «адресное» разрушение гена, («антисмысловая» блокировка гена или производимой им РНК), позволяющая вывести из
- 88. Биосенсоры, биочипы В 1975 г. Э.Саузерн- использовал меченную нуклеиновую кислоту, иммобилизированную на плотной основе. У нас
- 89. Иммобилизованные биообъекты Под иммобилизацией понимают связывание биообъекта с нерастворимым носителем при сохранении его функциональной активности фермента
- 90. Основной принцип работы биосенсоров- взаимодействие комплементарных цепей нуклеиновых кислот . Происходит взаимодействие ДНК-мишени с иммобилизированной пробой
- 91. Технология биочипов Применяется: Для выявления инфекционных агентов и их антибиотикоустойчивых форм Выявления полиморфизма по единичным нуклеотидам
- 94. Скачать презентацию

Генетика- наука о наследственности и изменчивости
Ген- фрагмент молекулы ДНК, контролирующий
Генетика- наука о наследственности и изменчивости
Ген- фрагмент молекулы ДНК, контролирующий

Особенности бактерий как генетического объекта
микроорганизмы имеют малые размеры, высокую скорость размножения,
Особенности бактерий как генетического объекта
микроорганизмы имеют малые размеры, высокую скорость размножения,

Отличительные особенности организации генома прокариот
Высокое абсолютное число генов.
Относительно высокое (70%) содержание
Отличительные особенности организации генома прокариот
Высокое абсолютное число генов.
Относительно высокое (70%) содержание
Удвоение бактериальной хромосомы сопровождается делением клетки.

Этапы репликации ДНК
3 этапа: инициация, элонгация и терминация.
Репликация начинается
Этапы репликации ДНК
3 этапа: инициация, элонгация и терминация.
Репликация начинается
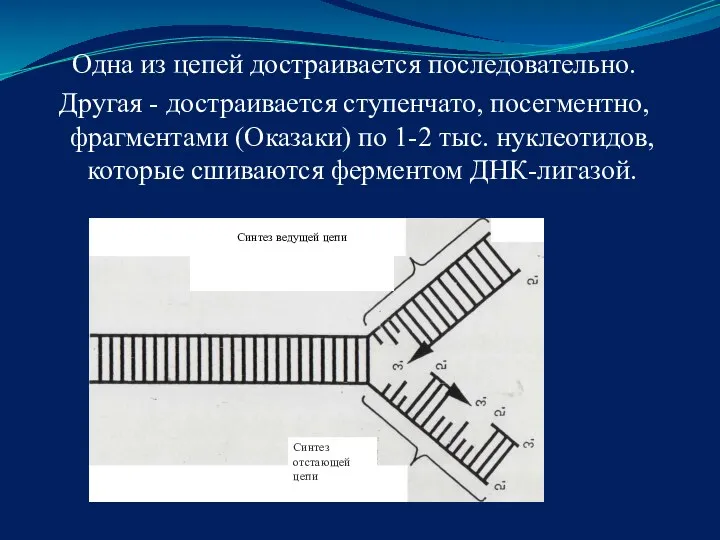
Одна из цепей достраивается последовательно.
Другая - достраивается ступенчато, посегментно, фрагментами
Одна из цепей достраивается последовательно.
Другая - достраивается ступенчато, посегментно, фрагментами
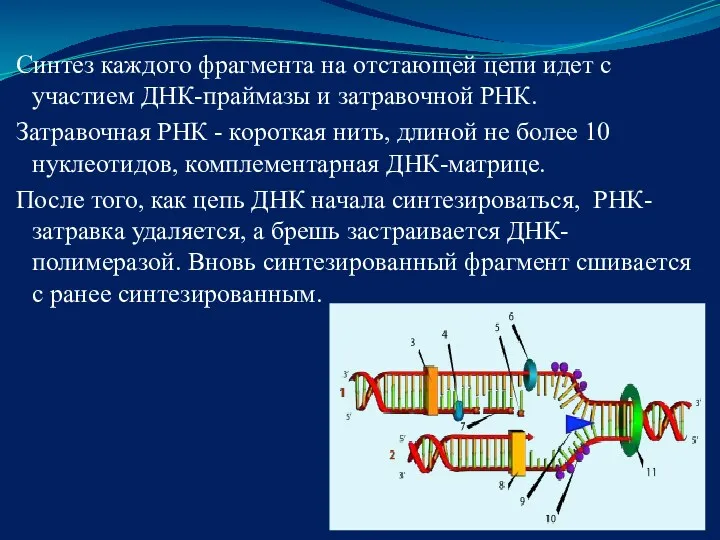
Синтез каждого фрагмента на отстающей цепи идет с участием ДНК-праймазы и
Синтез каждого фрагмента на отстающей цепи идет с участием ДНК-праймазы и
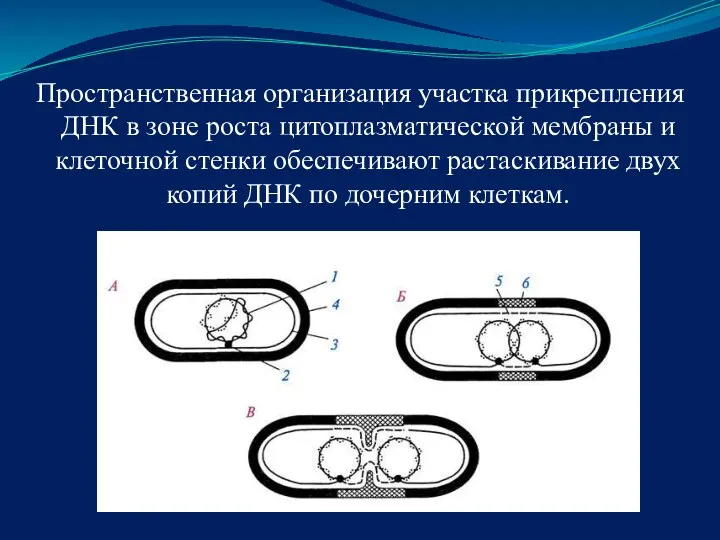
Пространственная организация участка прикрепления ДНК в зоне роста цитоплазматической мембраны и
Пространственная организация участка прикрепления ДНК в зоне роста цитоплазматической мембраны и

Структурно-функциональная единица ДНК
Основной структурно-функциональной единицей ДНК является оперон.
Оперон включает в себя
Структурно-функциональная единица ДНК
Основной структурно-функциональной единицей ДНК является оперон.
Оперон включает в себя

Регуляторные элементы:
Энхансер - генетический элемент, усиливающий транскрипцию оперона
Аттенуатор - генетический элемент,
Регуляторные элементы:
Энхансер - генетический элемент, усиливающий транскрипцию оперона
Аттенуатор - генетический элемент,

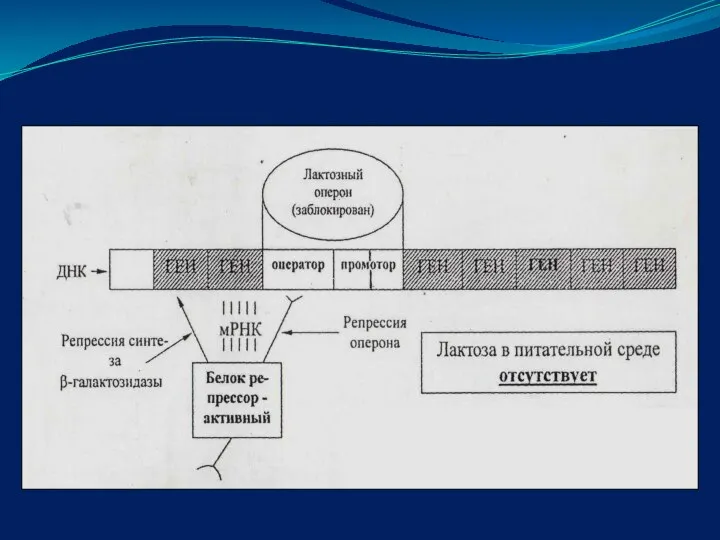
Организация работы оперона
В обычных условиях ген-регулятор активен и в клетке наблюдается
Организация работы оперона
В обычных условиях ген-регулятор активен и в клетке наблюдается

Внехромосомные генетические элементы
Представлены:
плазмидами,
транспозонами,
вставочными элементами (инсерционные)
умеренными или дефектными
Внехромосомные генетические элементы
Представлены:
плазмидами,
транспозонами,
вставочными элементами (инсерционные)
умеренными или дефектными

Плазмиды
подвижные генетические элементы, представленные замкнутой ДНК. Могут быть несвязанными с бактериальной
Плазмиды
подвижные генетические элементы, представленные замкнутой ДНК. Могут быть несвязанными с бактериальной
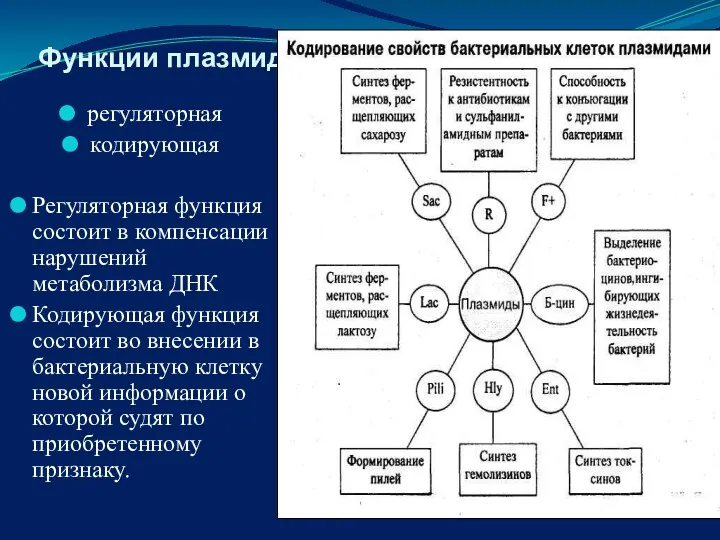
Функции плазмид
регуляторная
кодирующая
Регуляторная функция состоит в компенсации
Функции плазмид
регуляторная
кодирующая
Регуляторная функция состоит в компенсации

Плазмиды широко распространены у микроорганизмов. Они могут передаваться внутри вида,
Плазмиды широко распространены у микроорганизмов. Они могут передаваться внутри вида,

Классификация плазмид по свойствам,
которыми они наделяет своих носителей
Классификация плазмид по свойствам,
которыми они наделяет своих носителей

Транспозоны
– нуклеотидные последовательности от 2000- до 20 000 пар нуклеотидов, несут
Транспозоны
– нуклеотидные последовательности от 2000- до 20 000 пар нуклеотидов, несут

Транспозоны могут находиться в свободном состоянии в виде кольцевой молекулы, неспособной
Транспозоны могут находиться в свободном состоянии в виде кольцевой молекулы, неспособной
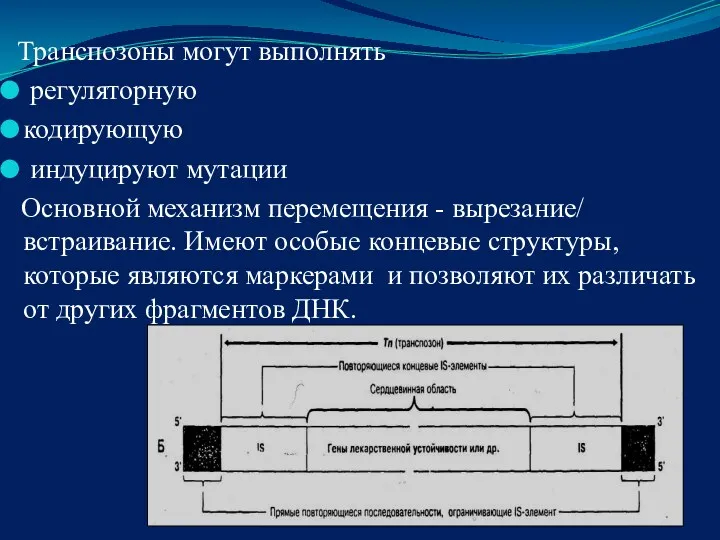
Транспозоны могут выполнять
регуляторную
кодирующую
индуцируют мутации
Основной механизм
Транспозоны могут выполнять
регуляторную
кодирующую
индуцируют мутации
Основной механизм

Вставочные элементы
(IS-последовательности) представляют собой мигрирующие элементы величиной от 800-1400 пар
Вставочные элементы
(IS-последовательности) представляют собой мигрирующие элементы величиной от 800-1400 пар

Функции IS- последовательностей:
координируют взаимодействие транспозонов, плазмид и умеренных фагов, как между
Функции IS- последовательностей:
координируют взаимодействие транспозонов, плазмид и умеренных фагов, как между
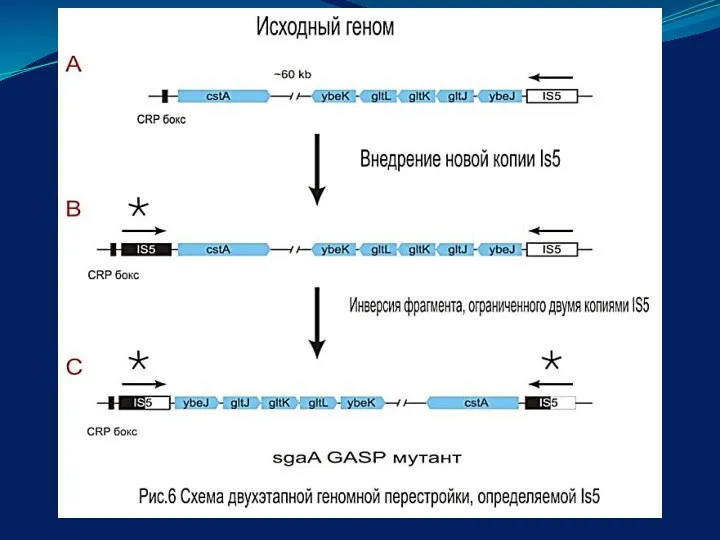

А. Внедрение IS-элемента в геном;
Б. Распространение IS-элемента в геноме за счет
А. Внедрение IS-элемента в геном; Б. Распространение IS-элемента в геноме за счет
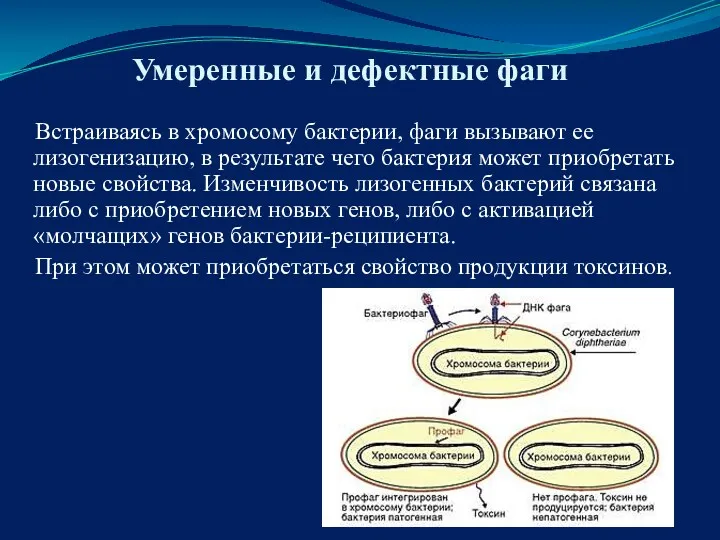
Умеренные и дефектные фаги
Встраиваясь в хромосому бактерии, фаги вызывают
Умеренные и дефектные фаги
Встраиваясь в хромосому бактерии, фаги вызывают

Изменчивость микроорганизмов
Фенотипическая
Генотипическая
Изменчивость микроорганизмов
Фенотипическая
Генотипическая

Наследственная изменчивость
связана с изменением последовательности нуклеотидов в ДНК, полной или
Наследственная изменчивость
связана с изменением последовательности нуклеотидов в ДНК, полной или

Спонтанные мутации
проявляются в популяции в естественных условиях под влиянием невыясненных
Спонтанные мутации
проявляются в популяции в естественных условиях под влиянием невыясненных

Индуцированные мутации возникают под влиянием конкретного события или воздействия.
Мутагены -
Индуцированные мутации возникают под влиянием конкретного события или воздействия.
Мутагены -
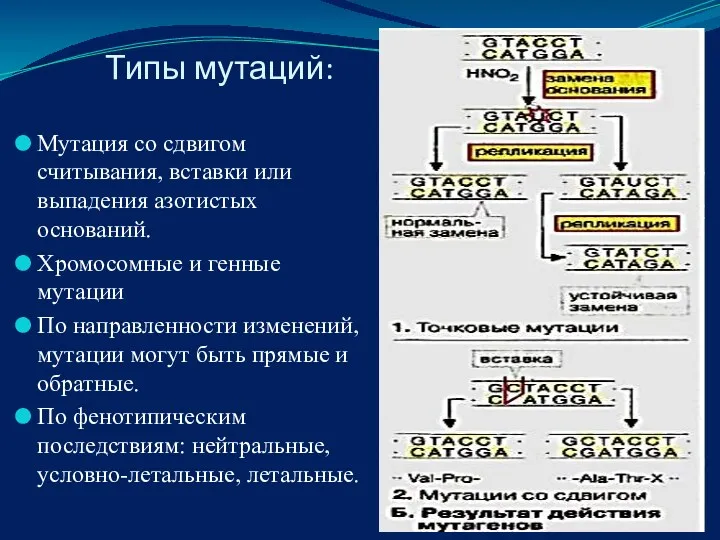
Типы мутаций:
Мутация со сдвигом считывания, вставки или выпадения азотистых оснований.
Типы мутаций:
Мутация со сдвигом считывания, вставки или выпадения азотистых оснований.

S – R – диссоциации
Возникают после встраивания внехромосомных факторов наследования
S – R – диссоциации
Возникают после встраивания внехромосомных факторов наследования
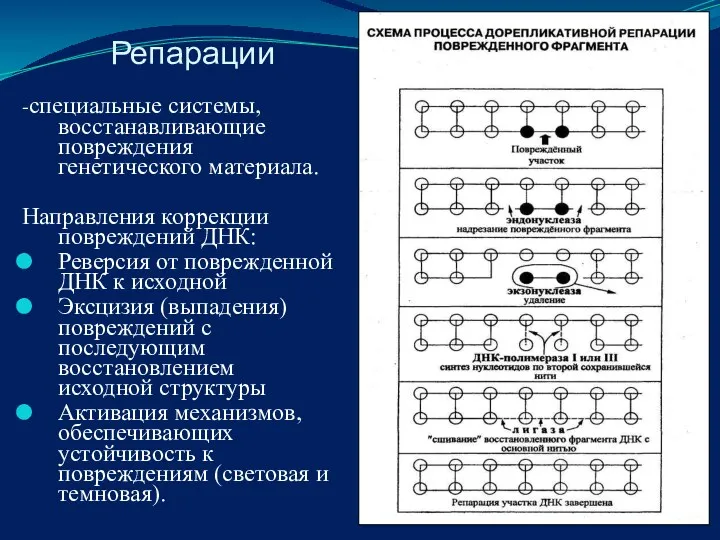
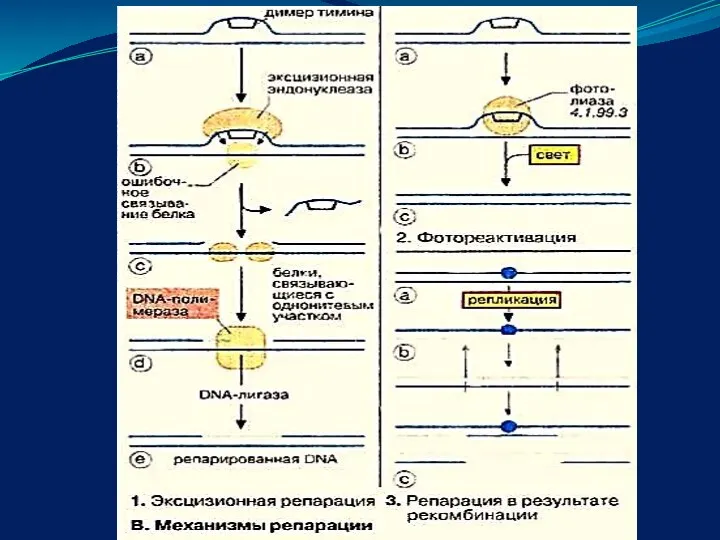
Репарации
-специальные системы, восстанавливающие повреждения генетического материала.
Направления коррекции повреждений ДНК:
Реверсия от поврежденной
Репарации
-специальные системы, восстанавливающие повреждения генетического материала.
Направления коррекции повреждений ДНК:
Реверсия от поврежденной

Генетические рекомбинации
Обмен генетическим материалом между отдельными особями в популяции клеток
В
Генетические рекомбинации
Обмен генетическим материалом между отдельными особями в популяции клеток
В
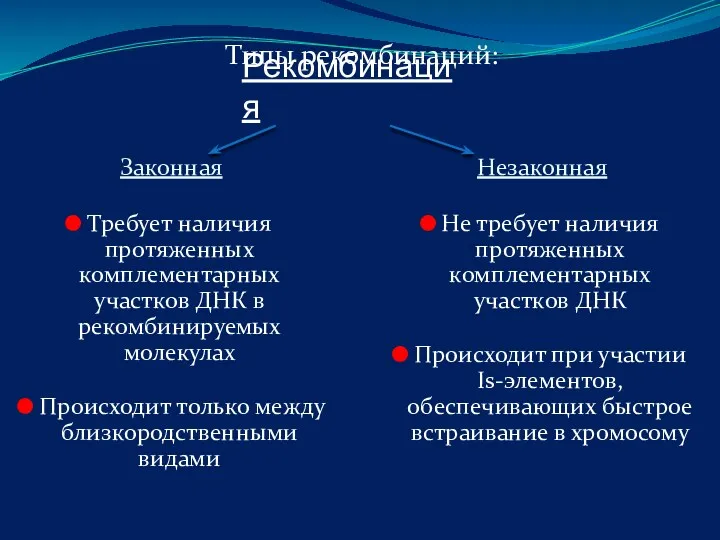
Рекомбинация
Законная
Требует наличия протяженных комплементарных участков ДНК в рекомбинируемых молекулах
Происходит только между
Рекомбинация
Законная
Требует наличия протяженных комплементарных участков ДНК в рекомбинируемых молекулах
Происходит только между
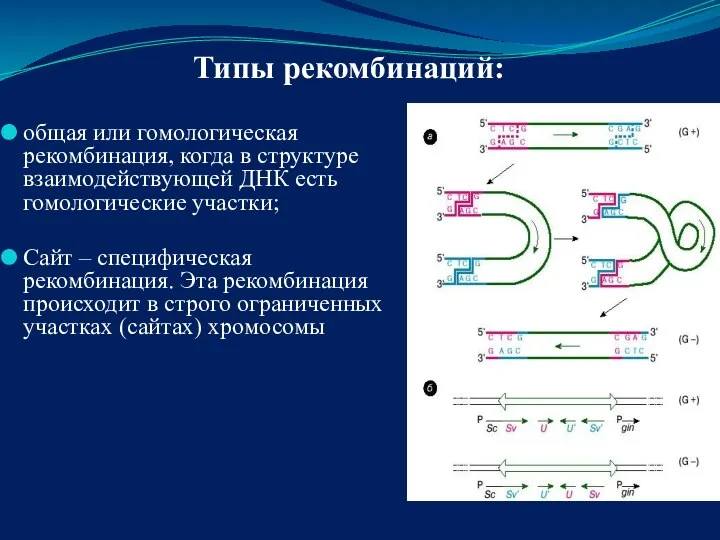
общая или гомологическая рекомбинация, когда в структуре взаимодействующей ДНК есть гомологические
общая или гомологическая рекомбинация, когда в структуре взаимодействующей ДНК есть гомологические
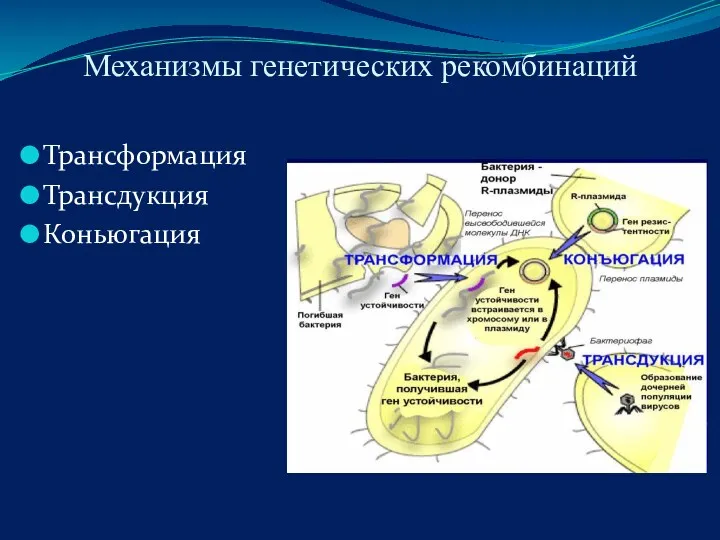
Механизмы генетических рекомбинаций
Трансформация
Трансдукция
Коньюгация
Механизмы генетических рекомбинаций
Трансформация
Трансдукция
Коньюгация

Трансформация
перенос генетического материала клетки донора, при котором реципиент захватывает из внешней
Трансформация
перенос генетического материала клетки донора, при котором реципиент захватывает из внешней
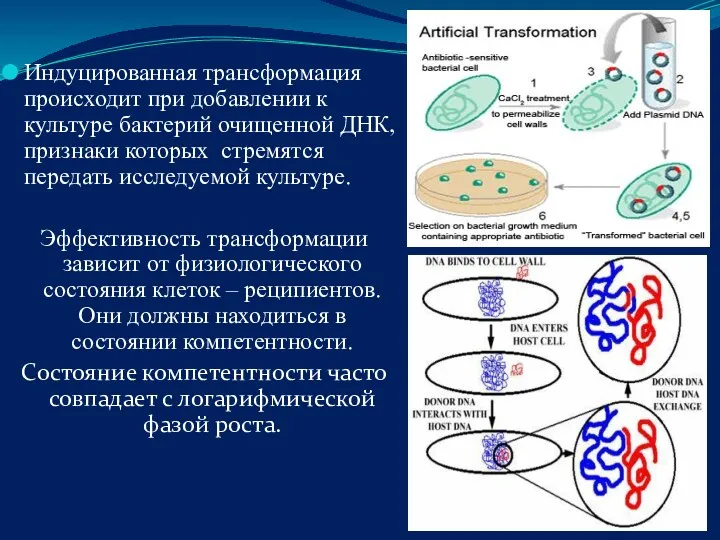
Индуцированная трансформация происходит при добавлении к культуре бактерий очищенной ДНК, признаки
Индуцированная трансформация происходит при добавлении к культуре бактерий очищенной ДНК, признаки
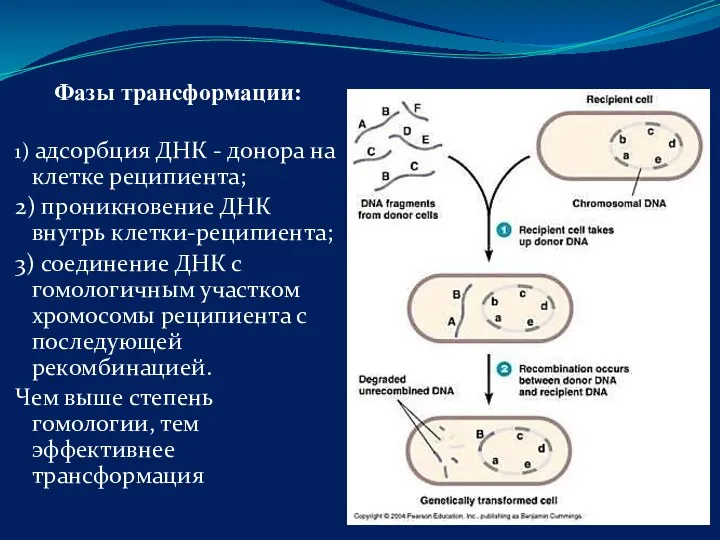
Фазы трансформации:
1) адсорбция ДНК - донора на клетке реципиента;
2) проникновение
Фазы трансформации:
1) адсорбция ДНК - донора на клетке реципиента;
2) проникновение
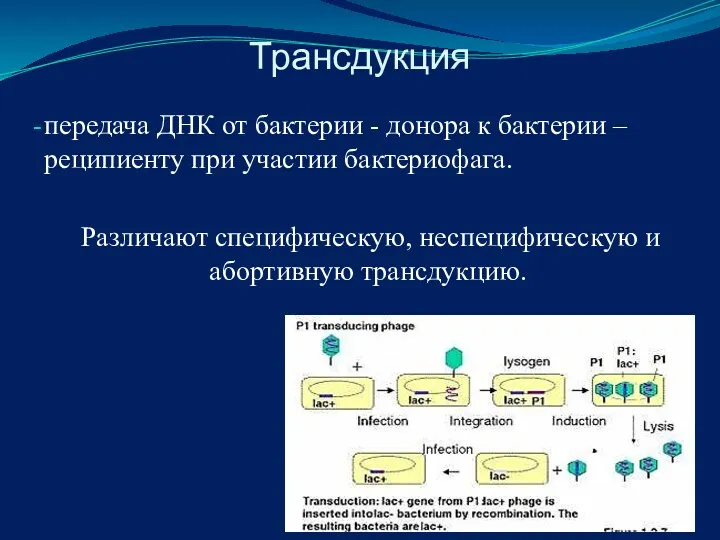
Трансдукция
передача ДНК от бактерии - донора к бактерии – реципиенту при
Трансдукция
передача ДНК от бактерии - донора к бактерии – реципиенту при
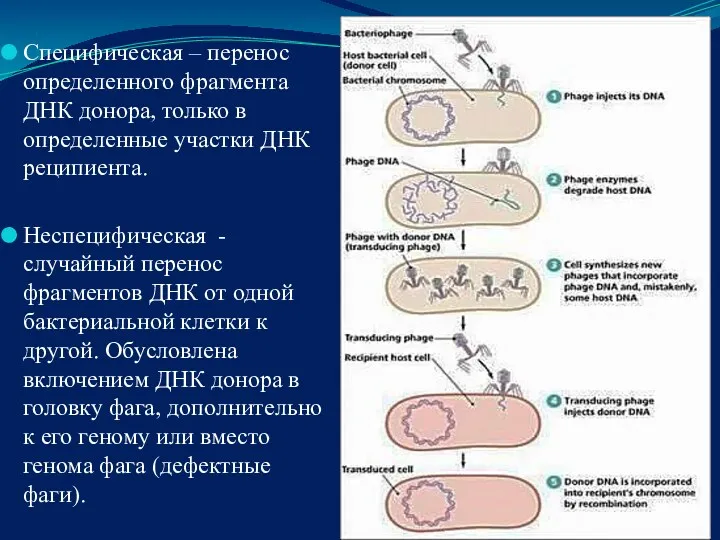
Специфическая – перенос определенного фрагмента ДНК донора, только в определенные участки
Специфическая – перенос определенного фрагмента ДНК донора, только в определенные участки
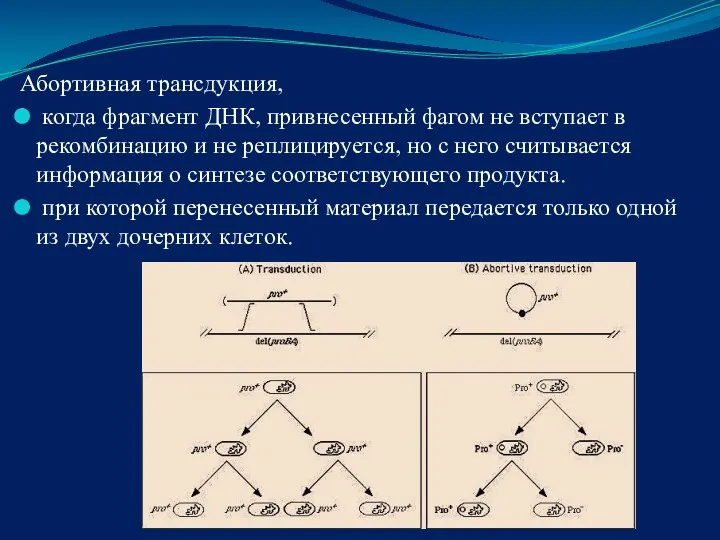
Абортивная трансдукция,
когда фрагмент ДНК, привнесенный фагом не вступает в рекомбинацию
Абортивная трансдукция,
когда фрагмент ДНК, привнесенный фагом не вступает в рекомбинацию

Конъюгация
перенос генетического материала из клетки в клетку при их непосредственном взаимодействии.
Донорами
Конъюгация
перенос генетического материала из клетки в клетку при их непосредственном взаимодействии.
Донорами

При взаимодействии F “+” клетки с F “-” клеткой половой фактор
При взаимодействии F “+” клетки с F “-” клеткой половой фактор

Первый этап конъюгации – прикреплении клетки-донора к реципиентной клетке с помощью
Первый этап конъюгации – прикреплении клетки-донора к реципиентной клетке с помощью

Генетика вирусов
Характеристика вирусных популяций
Высокая численность популяции увеличивает вероятность мутаций
Быстрая смена поколений
Генетика вирусов
Характеристика вирусных популяций
Высокая численность популяции увеличивает вероятность мутаций
Быстрая смена поколений

Ненаследуемые изменения у вирусов связаны с особенностями клетки хозяина и проявляются
Ненаследуемые изменения у вирусов связаны с особенностями клетки хозяина и проявляются
Фенотипическое смешивание
при смешанном заражении клеток несколькими вирусами, если часть потомства
Фенотипическое смешивание
при смешанном заражении клеток несколькими вирусами, если часть потомства

Мутации
У вирусов возникают во время репликации их нуклеиновых кислот.
Мутанты
Мутации
У вирусов возникают во время репликации их нуклеиновых кислот.
Мутанты

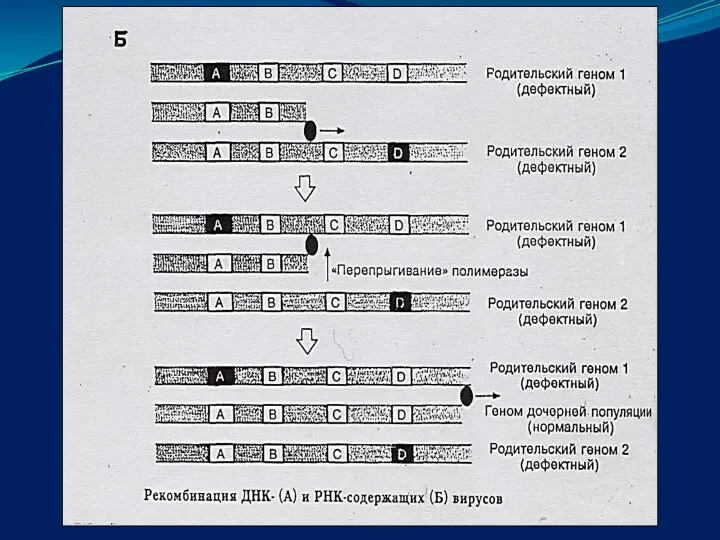
Вирусы способны и к генетическим рекомбинациям.
При одновременном заражении двумя вирусами
Вирусы способны и к генетическим рекомбинациям.
При одновременном заражении двумя вирусами

Генетическая реактивация - перераспределение генов, когда у родственных вирусов инактивированы разные
Генетическая реактивация - перераспределение генов, когда у родственных вирусов инактивированы разные
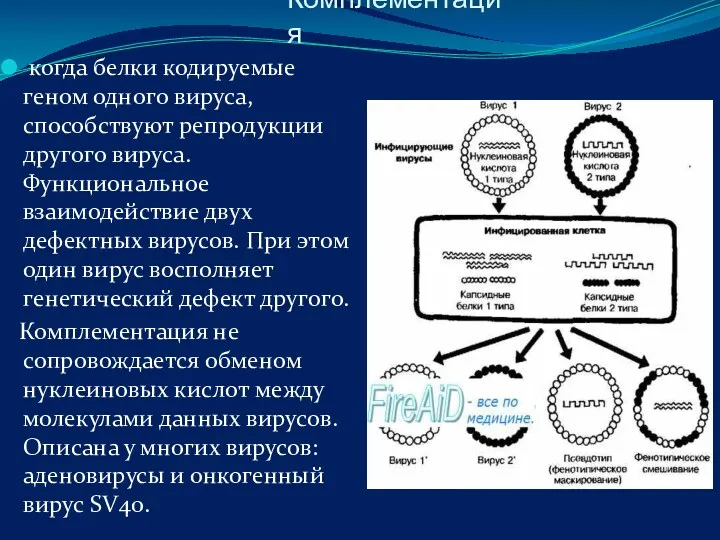
Комплементация
когда белки кодируемые геном одного вируса, способствуют репродукции другого
Комплементация
когда белки кодируемые геном одного вируса, способствуют репродукции другого

Интерферирующие взаимодействия
Состояние невосприимчивости к вторичному заражению клетки уже инфицированной вирусами
Интерференция может
Интерферирующие взаимодействия
Состояние невосприимчивости к вторичному заражению клетки уже инфицированной вирусами
Интерференция может

Гетерологическая интерференция. Инфицирование одним вирусом полностью блокирует возможность репликации другого вируса
Гетерологическая интерференция. Инфицирование одним вирусом полностью блокирует возможность репликации другого вируса

Биотехнология
Bios- жизнь, tecen- искусство, logos- наука
Термин БИОТЕХHОЛОГИЯ впервые использовал
К. Эреки
Биотехнология
Bios- жизнь, tecen- искусство, logos- наука
Термин БИОТЕХHОЛОГИЯ впервые использовал
К. Эреки

H а п р а в л е н и я
H а п р а в л е н и я

Медицинская биотехнология
Иммунобиологическая биотехнология изучает способы и методы конструирования, биотехнологию получения,
Медицинская биотехнология
Иммунобиологическая биотехнология изучает способы и методы конструирования, биотехнологию получения,
Иммуногенетическая биотехнология связана с производством вакцин нового поколения для профилактики
Иммуногенетическая биотехнология связана с производством вакцин нового поколения для профилактики

Объект исследования
Биообъект
- Целостный сохранивший жизнеспособность многоклеточный или одноклеточный организм, либо изолированные
Объект исследования
Биообъект
- Целостный сохранивший жизнеспособность многоклеточный или одноклеточный организм, либо изолированные

Б и о о б ъ е к т ы:
Микроорганизмы: бактерии,
вирусы,
Б и о о б ъ е к т ы:
Микроорганизмы: бактерии,
вирусы,

Биообъект, осуществляющий полный синтез целевого продукта, называется продуцентом.

Бактерии - продуценты
представители родов:
Acetobaсter - превращают этанол в
Бактерии - продуценты
представители родов:
Acetobaсter - превращают этанол в

Клетки животных или растений.
Из них извлекают широкий ассортимент сложной и
Клетки животных или растений.
Из них извлекают широкий ассортимент сложной и

Типы биотехнологических производств
Использование живой или инактивированной биомассы (закваски, получение пекарских дрожжей,
Типы биотехнологических производств
Использование живой или инактивированной биомассы (закваски, получение пекарских дрожжей,

Прогресс биотехнологии в дальнейшем будет определяться развитием генной, клеточной и эмбриогенетической
Генная инженерия- совокупность приемов и методов связанных с целенаправленным конструированием in
Генная инженерия- совокупность приемов и методов связанных с целенаправленным конструированием in
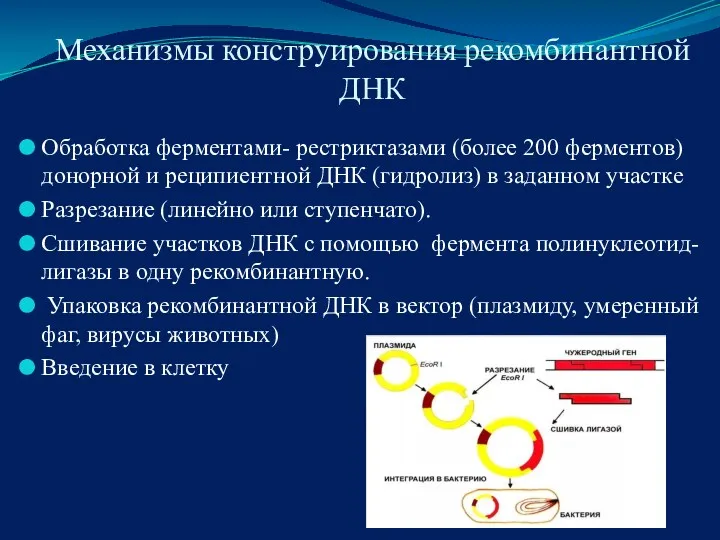
Механизмы конструирования рекомбинантной ДНК
Обработка ферментами- рестриктазами (более 200 ферментов) донорной и
Механизмы конструирования рекомбинантной ДНК
Обработка ферментами- рестриктазами (более 200 ферментов) донорной и

Трансгенезом называется встраивание чужих генов
растениям и животным.
генетически модифицированные источники
Трансгенезом называется встраивание чужих генов
растениям и животным.
генетически модифицированные источники
Трансгенные животные
Животные в геном которых интегрируют чужеродные гены, называют трансгенными.
В
Трансгенные животные
Животные в геном которых интегрируют чужеродные гены, называют трансгенными.
В

В клетки животных гены вводятся:
при добавлении в среду
для
В клетки животных гены вводятся:
при добавлении в среду
для

Клеточная инженерия- метод конструирования клеток нового типа на основе их культивирования,
Клеточная инженерия- метод конструирования клеток нового типа на основе их культивирования,
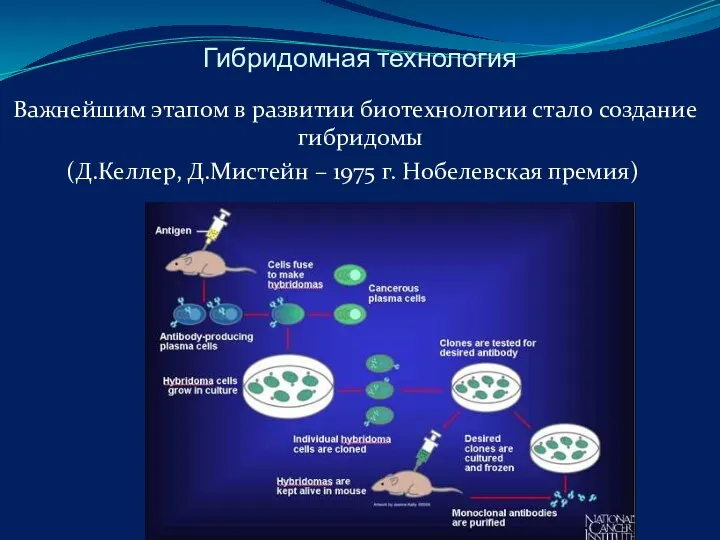
Гибридомная технология
Важнейшим этапом в развитии биотехнологии стало создание гибридомы
(Д.Келлер, Д.Мистейн –
Гибридомная технология
Важнейшим этапом в развитии биотехнологии стало создание гибридомы
(Д.Келлер, Д.Мистейн –
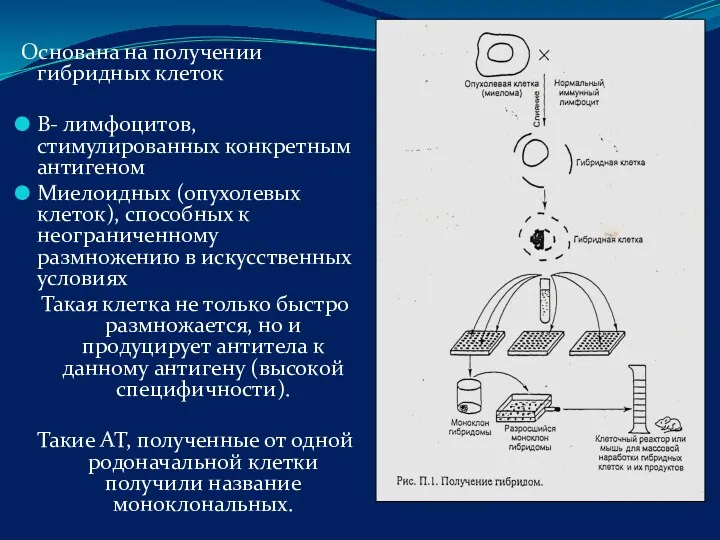
Основана на получении гибридных клеток
В- лимфоцитов, стимулированных конкретным антигеном
Миелоидных (опухолевых клеток),
Основана на получении гибридных клеток
В- лимфоцитов, стимулированных конкретным антигеном
Миелоидных (опухолевых клеток),

Моноклональная технология применяется:
Для диагностики:
- инфекционных заболеваний
- аутоимунных заболеваний
Моноклональная технология применяется:
Для диагностики:
- инфекционных заболеваний
- аутоимунных заболеваний

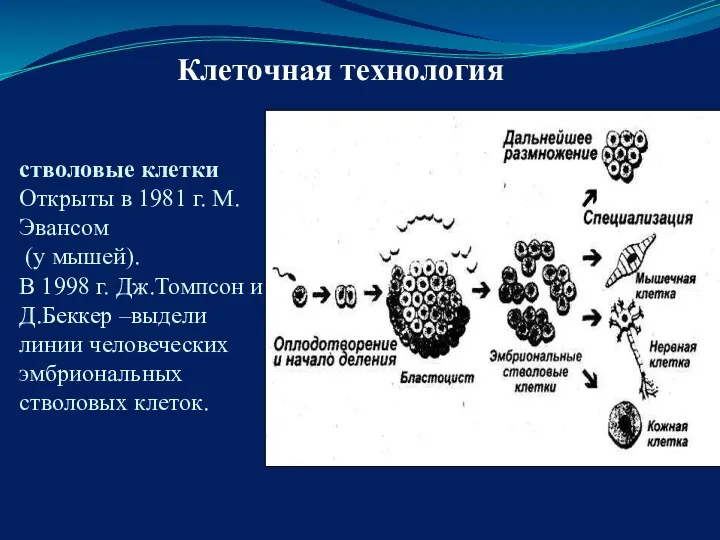
стволовые клетки
Открыты в 1981 г. М. Эвансом
(у мышей).
В 1998
стволовые клетки Открыты в 1981 г. М. Эвансом (у мышей). В 1998
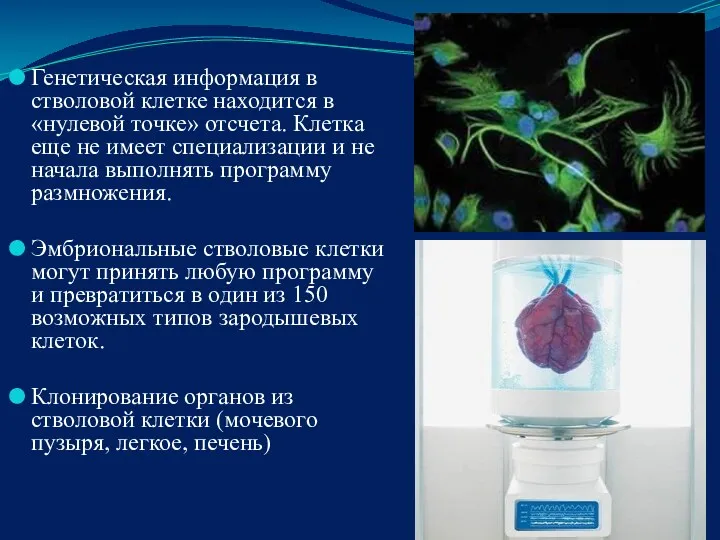
Генетическая информация в стволовой клетке находится в «нулевой точке» отсчета. Клетка
Генетическая информация в стволовой клетке находится в «нулевой точке» отсчета. Клетка
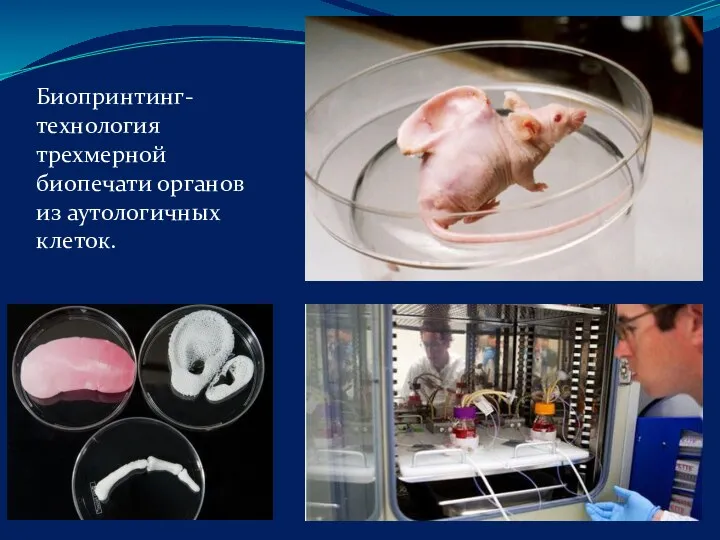
Биопринтинг- технология трехмерной биопечати органов из аутологичных клеток.
Биопринтинг- технология трехмерной биопечати органов из аутологичных клеток.

Эмбриогенетическая инженерия
Перестройка генома- реконструкция эмбрионов путем клонирования.
генно-инженерные методы радикального лечения наследственных
Эмбриогенетическая инженерия
Перестройка генома- реконструкция эмбрионов путем клонирования.
генно-инженерные методы радикального лечения наследственных
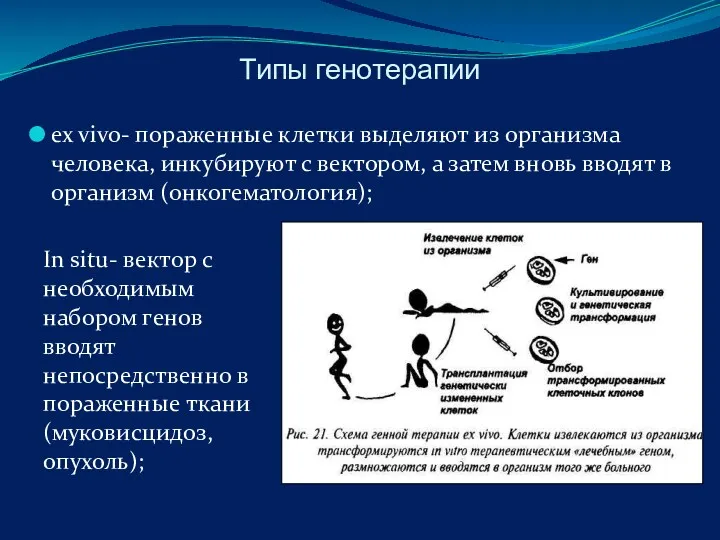
Типы генотерапии
ex vivo- пораженные клетки выделяют из организма человека, инкубируют с
Типы генотерапии
ex vivo- пораженные клетки выделяют из организма человека, инкубируют с

Баллистическая трансфекция- основана на обстреле органов и тканей частицами тяжелых металлов
Баллистическая трансфекция- основана на обстреле органов и тканей частицами тяжелых металлов

Избирательная инактивация гена
«адресное» разрушение гена, («антисмысловая» блокировка гена или производимой им
Избирательная инактивация гена
«адресное» разрушение гена, («антисмысловая» блокировка гена или производимой им

Биосенсоры, биочипы
В 1975 г. Э.Саузерн- использовал меченную нуклеиновую кислоту, иммобилизированную на
Биосенсоры, биочипы
В 1975 г. Э.Саузерн- использовал меченную нуклеиновую кислоту, иммобилизированную на

Иммобилизованные биообъекты
Под иммобилизацией понимают связывание биообъекта с нерастворимым носителем при сохранении
Иммобилизованные биообъекты
Под иммобилизацией понимают связывание биообъекта с нерастворимым носителем при сохранении

Основной принцип работы биосенсоров- взаимодействие комплементарных цепей нуклеиновых кислот .
Происходит взаимодействие
Основной принцип работы биосенсоров- взаимодействие комплементарных цепей нуклеиновых кислот .
Происходит взаимодействие

Технология биочипов
Применяется:
Для выявления инфекционных агентов и их антибиотикоустойчивых форм
Выявления полиморфизма по
Технология биочипов
Применяется:
Для выявления инфекционных агентов и их антибиотикоустойчивых форм
Выявления полиморфизма по

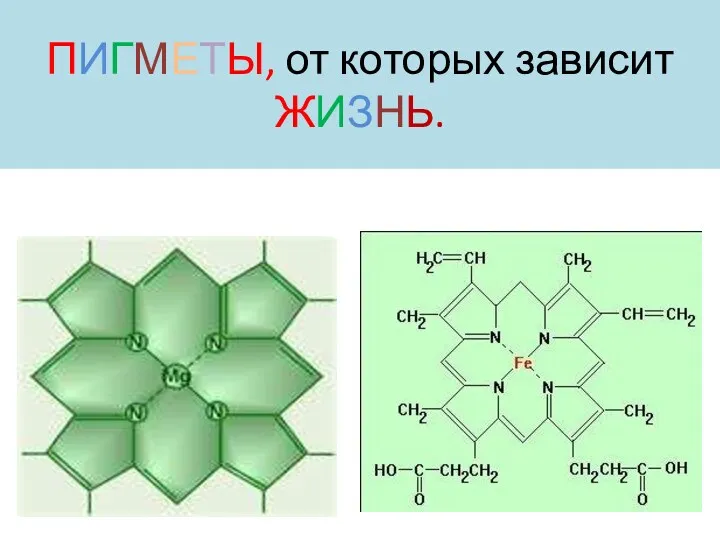 Пигменты, от которых зависит жизнь.
Пигменты, от которых зависит жизнь. Красная книга по исчезающим видам животных и растений Алькеевского района
Красная книга по исчезающим видам животных и растений Алькеевского района Пищеварительная система и процесс пищеварения у человека
Пищеварительная система и процесс пищеварения у человека Растения, вызывающие преимущественно поражение органов дыхания и пищеварительного тракта
Растения, вызывающие преимущественно поражение органов дыхания и пищеварительного тракта Черепные нервы IX-XII
Черепные нервы IX-XII 7 самых ядовитых растений России
7 самых ядовитых растений России Структурные компоненты нуклеиновых кислот. Уровни организации ДНК и РНК
Структурные компоненты нуклеиновых кислот. Уровни организации ДНК и РНК Популяция – структурная единица вида; основная единица эволюции
Популяция – структурная единица вида; основная единица эволюции Липиды. Классификация липидов
Липиды. Классификация липидов Белки, жиры и углеводы
Белки, жиры и углеводы АСҚ пайдаланылу жолдары
АСҚ пайдаланылу жолдары Течение и вязкость жидкостей
Течение и вязкость жидкостей Основные оложения клеточной теории. 10 кл.
Основные оложения клеточной теории. 10 кл. Дробление. Деление зиготы
Дробление. Деление зиготы Головной мозг. Черепные нервы
Головной мозг. Черепные нервы От микроскопа к клетке.
От микроскопа к клетке. Историческое и индивидуальное в развитии
Историческое и индивидуальное в развитии Нервная ткань
Нервная ткань Урок биологии 9 класс Звёздный час – Человек
Урок биологии 9 класс Звёздный час – Человек Антропология, расы, конституции
Антропология, расы, конституции Устройство увеличительных приборов
Устройство увеличительных приборов Глюконеогенез
Глюконеогенез Физиологический процесс дыхание
Физиологический процесс дыхание Обмен веществ. Метаболизм и его функции
Обмен веществ. Метаболизм и его функции Ткани растений
Ткани растений Акция Помоги птице зимой!
Акция Помоги птице зимой! Далалық және ауылшаруашылық дақылдары
Далалық және ауылшаруашылық дақылдары Внутрішня будова птахів
Внутрішня будова птахів