- Гормональная регуляция метаболизма: основы

Содержание
- 2. Метаболическая активность как отдельных клеток, так и органов, состоящих из диффе-ренцированных клеток, координируется и регулируется с
- 3. - Эндокринная система. Гормоны секретиру- ются специализированными клетками в кровоток, в результате чего гормоны могут транспортироваться
- 4. Все гормоны действуют через высокоспецифические рецепторы, которыми обладают клетки-мишени. Связывание гормона с рецептором происходит с вы-
- 5. Основы учения о вторичных мессенджерах были заложены открытием Э. Сазерленда явления передачи (трансдукции) регуляторного сигнала от
- 6. Внутриклеточные сигнальные пути могут содержать в своём составе несколько белковых молекул – посред-ников, связанных между собой
- 7. Первичные и вторичные мессенджеры Ядро Цитоплазма + Гормон (первичный мессенджер) Неактивированный рецептор Комплекс гормон-рецептор, активированный рецептор
- 8. Гормоны можно разделить на три группы – по тому, какой путь проходит их молекула от места
- 9. Классификация гомонов по химическому строению 1. Белково-пептидныне гормоны (инсулин, окистоцин, глюкагон); 2. Производные аминокислот (адреналин, тироксин);
- 10. Ни один из известных науке гормонов не является ферментом или коферментом – т.е. гормоны не син-тезируют
- 11. Все гормоны, по характеру их взаимодействия с клеточной мембраной, можно разделить на две группы: I группа:
- 12. II группа: Свободно проходят сквозь цитоплазматичес- кую мембрану в силу липофильности (непо- лярности) их молекул: стероидные
- 13. Важные дополнения: Наряду с перечисленными выше гормонами, суще- ствует масса различных регуляторных молекул – пер-вичных мессенджеров,
- 14. Отличие ПГ от истинных (классических) гормонов: - ПГ синтезируются во всех тканях организма человека, а не
- 15. Факторы роста. Белки, синтезируемые вне эндокрин- ных желёз, и специфически стимулирующие дифференцировку и деление клеток-мише- ней.
- 16. Общая характеристика рецепторов клеточной поверхности (мембранных рецепторов) Рецепторы представляют собой трансмембранный белок, состоящий из трёх доменов.
- 17. Внутриклеточный домен. Содержит С-концевой участок пептидной цепи. Отвечает за эффекторную функцию рецептора. Этот домен ретранслирует регу-
- 18. Т.о., система рецепторов на поверхности цито-плазматической мембране (система передачи регуля-торного сигнала) находится в постоянной готовно-сти. Её
- 19. На основании особенностей протекания пострецепторных событий, все мембранные рецепторы можно разделить на следующие классы: Гормон-чувствительные каналы.
- 20. 2. Рецепторы, обладающие собственной лиганд-зави-симой тирозинкиназной активностью (Рецептор-ассоциированные тирозиновые киназы или рецеп-торные тирозиновые киназы). Внеклеточный домен
- 21. К этому же классу рецепторов следует отнести рецеп- торные серин-треониновые киназы. Один из лигандов – трансформирующий
- 22. 4. Рецепторы цитокинов. После связывания цитокина с рецептором происхо-дит димеризация и активация последнего. Фермента-тивной активности нет,
- 23. STAT (signal transducers and activation transcription = проводники сигнала и активаторы транскрипции). Далее, Янус-киназа фосфорилирует связавшиеся
- 24. 5. Рецепторы, сопряженные с G-белком (G-protein coupled receptors = GPCR). Это наиболее многочисленная группа мембранных рецепторов
- 25. G-белки – состоят из 3-х субъединиц (с.е.): α, β и γ. В неактивной форме α- с.е.
- 26. Аденилатциклаза синтезирует вторичный мессенд- жер цАМФ: АТФ ? цАМФ цАМФ активирует цАМФ-зависимые протеинки-назы. Синтез цАМФ происходит
- 27. Фосфолипаза С – расщепляет мембранный фосфати- дилинозитол-бифосфат. В итоге образуется две мо- лекулы, два вторичных мессенджера
- 28. Общая характеристика внутриклеточных рецепторов Эта категория рецепторов служит для реализации регуляторных эффектов стериодных и тиреоидных гормонов,
- 29. Липофильные (неполярные) молекулы вышеука-занных гормонов легко проникают через цитоплаз-матическую мембрану путем диффузии. Рецепторы этих гормонов -
- 31. Скачать презентацию

Метаболическая активность как отдельных клеток, так и органов, состоящих из
Метаболическая активность как отдельных клеток, так и органов, состоящих из

- Эндокринная система. Гормоны секретиру-
ются специализированными клетками в кровоток, в
- Эндокринная система. Гормоны секретиру-
ются специализированными клетками в кровоток, в

Все гормоны действуют через высокоспецифические
рецепторы, которыми обладают клетки-мишени.
Связывание
Все гормоны действуют через высокоспецифические
рецепторы, которыми обладают клетки-мишени.
Связывание
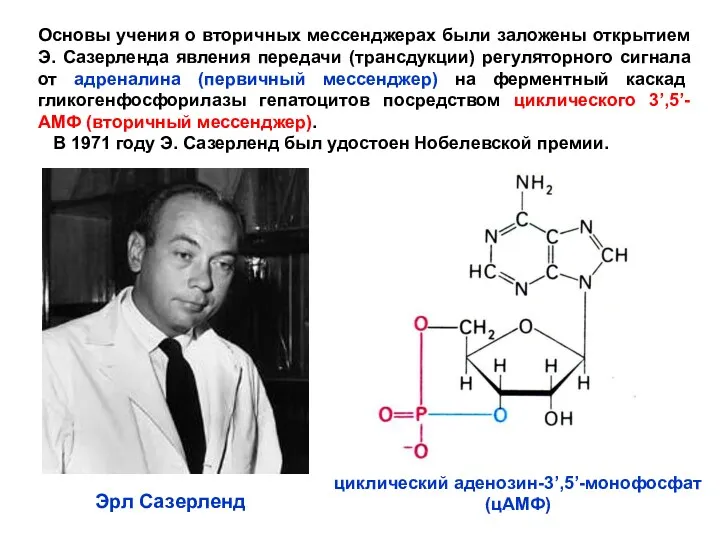
Основы учения о вторичных мессенджерах были заложены открытием Э. Сазерленда явления
Основы учения о вторичных мессенджерах были заложены открытием Э. Сазерленда явления

Внутриклеточные сигнальные пути могут содержать в своём составе несколько белковых молекул
Внутриклеточные сигнальные пути могут содержать в своём составе несколько белковых молекул
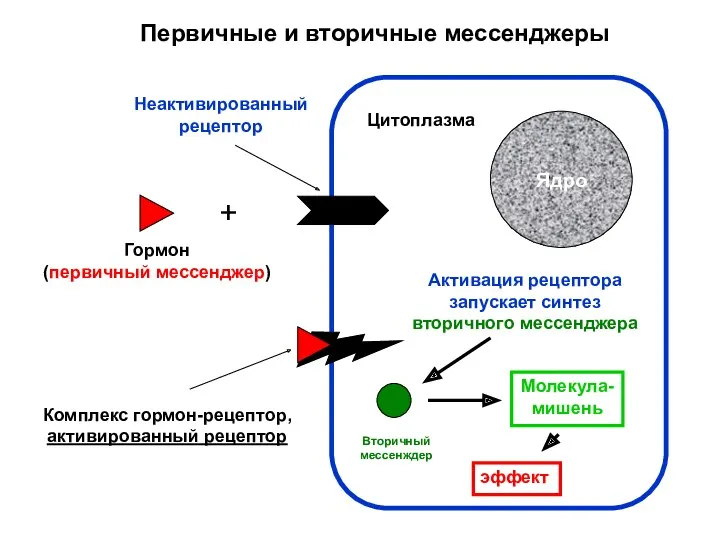
Первичные и вторичные мессенджеры
Ядро
Цитоплазма
+
Гормон
(первичный мессенджер)
Неактивированный
рецептор
Комплекс гормон-рецептор,
активированный рецептор
Активация рецептора
запускает синтез
вторичного
Первичные и вторичные мессенджеры
Ядро
Цитоплазма
+
Гормон
(первичный мессенджер)
Неактивированный
рецептор
Комплекс гормон-рецептор,
активированный рецептор
Активация рецептора
запускает синтез
вторичного

Гормоны можно разделить на три группы – по
тому, какой путь
Гормоны можно разделить на три группы – по
тому, какой путь

Классификация гомонов по химическому строению
1. Белково-пептидныне гормоны (инсулин, окистоцин,
глюкагон);
Классификация гомонов по химическому строению
1. Белково-пептидныне гормоны (инсулин, окистоцин,
глюкагон);

Ни один из известных науке гормонов не является
ферментом или
Ни один из известных науке гормонов не является
ферментом или

Все гормоны, по характеру их взаимодействия с
клеточной мембраной, можно
Все гормоны, по характеру их взаимодействия с
клеточной мембраной, можно

II группа: Свободно проходят сквозь цитоплазматичес-
кую мембрану в силу липофильности
II группа: Свободно проходят сквозь цитоплазматичес-
кую мембрану в силу липофильности

Важные дополнения:
Наряду с перечисленными выше гормонами, суще-
ствует масса различных
Важные дополнения:
Наряду с перечисленными выше гормонами, суще-
ствует масса различных

Отличие ПГ от истинных (классических) гормонов:
- ПГ синтезируются во всех
Отличие ПГ от истинных (классических) гормонов:
- ПГ синтезируются во всех

Факторы роста. Белки, синтезируемые вне эндокрин-
ных желёз, и специфически
Факторы роста. Белки, синтезируемые вне эндокрин-
ных желёз, и специфически

Общая характеристика рецепторов клеточной
поверхности (мембранных рецепторов)
Рецепторы представляют собой трансмембранный
Общая характеристика рецепторов клеточной
поверхности (мембранных рецепторов)
Рецепторы представляют собой трансмембранный

Внутриклеточный домен. Содержит С-концевой
участок пептидной цепи. Отвечает за эффекторную
функцию
Внутриклеточный домен. Содержит С-концевой
участок пептидной цепи. Отвечает за эффекторную
функцию

Т.о., система рецепторов на поверхности цито-плазматической мембране (система передачи регуля-торного
Т.о., система рецепторов на поверхности цито-плазматической мембране (система передачи регуля-торного

На основании особенностей протекания
пострецепторных событий, все мембранные
рецепторы можно разделить
На основании особенностей протекания
пострецепторных событий, все мембранные
рецепторы можно разделить

2. Рецепторы, обладающие собственной лиганд-зави-симой тирозинкиназной активностью (Рецептор-ассоциированные тирозиновые киназы или
2. Рецепторы, обладающие собственной лиганд-зави-симой тирозинкиназной активностью (Рецептор-ассоциированные тирозиновые киназы или

К этому же классу рецепторов следует отнести рецеп-
торные серин-треониновые киназы.
К этому же классу рецепторов следует отнести рецеп-
торные серин-треониновые киназы.

4. Рецепторы цитокинов.
После связывания цитокина с рецептором происхо-дит димеризация и активация
4. Рецепторы цитокинов.
После связывания цитокина с рецептором происхо-дит димеризация и активация

STAT (signal transducers and activation transcription =
проводники сигнала и
STAT (signal transducers and activation transcription =
проводники сигнала и

5. Рецепторы, сопряженные с G-белком (G-protein
coupled receptors = GPCR).
5. Рецепторы, сопряженные с G-белком (G-protein
coupled receptors = GPCR).

G-белки – состоят из 3-х субъединиц (с.е.): α, β и
G-белки – состоят из 3-х субъединиц (с.е.): α, β и

Аденилатциклаза синтезирует вторичный мессенд-
жер цАМФ:
АТФ ? цАМФ
цАМФ активирует
Аденилатциклаза синтезирует вторичный мессенд-
жер цАМФ:
АТФ ? цАМФ
цАМФ активирует

Фосфолипаза С – расщепляет мембранный фосфати-
дилинозитол-бифосфат. В итоге образуется две
Фосфолипаза С – расщепляет мембранный фосфати-
дилинозитол-бифосфат. В итоге образуется две

Общая характеристика внутриклеточных рецепторов
Эта категория рецепторов служит для реализации
регуляторных
Общая характеристика внутриклеточных рецепторов
Эта категория рецепторов служит для реализации
регуляторных

Липофильные (неполярные) молекулы вышеука-занных гормонов легко проникают через цитоплаз-матическую мембрану
Липофильные (неполярные) молекулы вышеука-занных гормонов легко проникают через цитоплаз-матическую мембрану
 Морфология и физиология вирусов
Морфология и физиология вирусов Отдел Красные Водоросли. Лекция 7
Отдел Красные Водоросли. Лекция 7 Молекулалық генетика негіздері. Ген негіздері. Гендердің интроиды, экзонды орналасуы. Прокориоттардың тұқым қуалауының
Молекулалық генетика негіздері. Ген негіздері. Гендердің интроиды, экзонды орналасуы. Прокориоттардың тұқым қуалауының Типология лесов и многолетних насаждений
Типология лесов и многолетних насаждений Паукообразные. Особенности строения и жизнедеятельности
Паукообразные. Особенности строения и жизнедеятельности Мышечные и нервная ткани
Мышечные и нервная ткани Урок биологии для 9 класса по теме Вредные привычки. Заболевания человека УМК Н.И.Сонина
Урок биологии для 9 класса по теме Вредные привычки. Заболевания человека УМК Н.И.Сонина Опорные системы и их значение в жизни организмов. Шаблон
Опорные системы и их значение в жизни организмов. Шаблон Набута поведінка тварин. 7 клас
Набута поведінка тварин. 7 клас Презентация 1
Презентация 1 Развитие производных эктодермы
Развитие производных эктодермы Антропогенез. Расы. Расизм. Часть 6
Антропогенез. Расы. Расизм. Часть 6 Генетические основы эволюции
Генетические основы эволюции Органы дыхания беспозвоночных и позвоночных животных
Органы дыхания беспозвоночных и позвоночных животных Презентация к уроку Транспорт веществ (6 класс)
Презентация к уроку Транспорт веществ (6 класс) Пчёлы. Продолжительность Жизни пчёл
Пчёлы. Продолжительность Жизни пчёл Молекулярная биология
Молекулярная биология Третий закон Грегора Менделя
Третий закон Грегора Менделя Растения и окружающая среда
Растения и окружающая среда Анатомия и физиология больших пищеварительных желёз: поджелудочная железа, большие слюнные железы. Лекция 39
Анатомия и физиология больших пищеварительных желёз: поджелудочная железа, большие слюнные железы. Лекция 39 Вендские жители Земли
Вендские жители Земли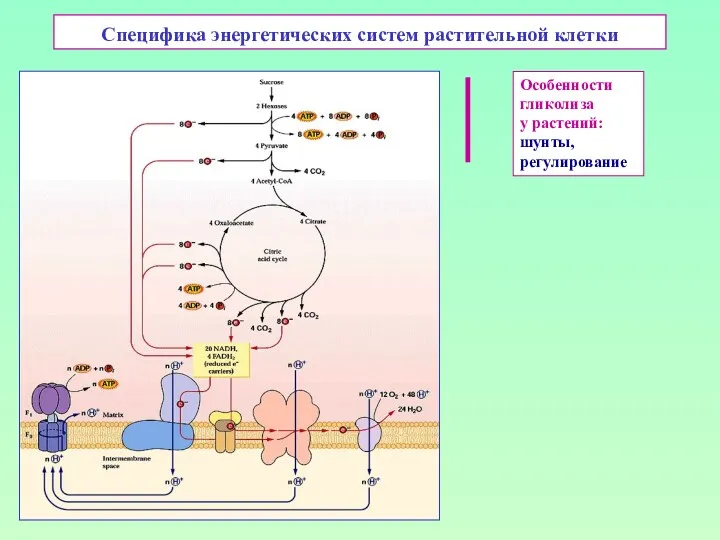 Специфика энергетических систем растительной клетки
Специфика энергетических систем растительной клетки Охорона біосфери
Охорона біосфери Какие бывают животные? Окружающий мир. 2 класс
Какие бывают животные? Окружающий мир. 2 класс Естественный отбор – главная движущая сила эволюции
Естественный отбор – главная движущая сила эволюции Тайны коры головного мозга
Тайны коры головного мозга Растения и животные пустынь
Растения и животные пустынь Белки
Белки