- Гормони

Содержание
- 2. Гормони – органічні речовини дистанційної дії, що виробляються в ендокринних залозах та здійснюють разом з нервовою
- 3. За допомогою крові гормони розносяться по всьому організму, але діють тільки на певні органи, тканини (які
- 4. За хімічною будовою розрізняють: Білки і пептиди Похідні окремих амінокислот Стероїди
- 7. Біологічна роль 1. Змінюють інтенсивність метаболізму 2. Активують (інгібують) ферменти 3. Прискорюють (сповільнюють) синтез і розпад
- 8. Основні загальні біологічні ознаки: Дистантність дії – біологічну дію проявляють на відстані від місця виділення, від
- 9. Схема взаємозв'язку регуляторних систем організму
- 10. Найбільш важливою є гіпоталамо-гіпофізарна система, яка контролюється центральною нервовою системою. Збудження передається по нервовим волокнам спочатку
- 11. Основні етапи гормональних сигналів
- 12. Початковий етап у дії гормону на клітину-мішень – взаємодія гормону з рецептором клітини. Концентрація гормонів у
- 14. Мембранний або аденілатциклазний механізм (для гормонів білкової і пептидної природи, а також похідних амінокислот катехоламінів -
- 15. Цитозольний механізм дії
- 16. Цитозольний (внутрішньоклітинний) Є характерним для стероїдних і основної маси тиреоїдних гормонів. Це так звані ліпофільні гормони.
- 17. Гіпоталамо – гіпофізарна система
- 19. Гормони гіпофізу Гормони передньої частки гіпофізу В передній частині гіпофізу синтезуються так звані тропні гормони, які
- 20. 1. Соматотропний гормон (СТГ, соматотропін, гормон росту)
- 21. Соматотропний гормон – (СТГ, гормон росту, соматотропін). Це поліпептид, який складається зі 191 амінокислотного залишку. Стимулюється
- 22. Біологічна дія гормону росту
- 23. Основна дія гормону росту направлена на регуляцію обміну білків і процесів, пов'язаних з ростом та розвитком
- 24. Гіпофункція гіпофізу по СТГ приводить у молодих організмів до карликовості – пропорційному зменшенню росту всіх органів.
- 28. Акромегалія
- 29. 2. Тиреотропний гормон – (ТТГ, тиреотропін). За хімічною природою це складний білок – глікопротеїн. Утворення ТТГ
- 30. 3. Адренокортикотропний гормон (АКТГ, кортикотропін). Це поліпептид, який містить 39 амінокислотних залишки. Синтез АКТГ стимулюється гормоном
- 31. 4. Фолікулостимулюючий гормон (ФСГ, фолітропін). За хімічною структурою це складний білок – глікопротеїн. ФСГ, в результаті
- 32. 5. Лютеїнізуючий гормон (ЛГ, лютропін). Є глікопротеїном. У самок ЛГ стимулює овуляцію, а також синтез в
- 33. 6. Пролактин (ПРЛ, лактотропін). Це білок, який складається зі 199 амінокислотних залишків, що утворюють один поліпептидний
- 34. Гормон середньої частки гіпофізу Меланоцит-стимулюючий гормон (МСГ) Поліпептид, у багатьох тварин складається з 18 а.к. залишків,
- 35. Гормони задньої частки гіпофізу Окситоцин і вазопресин Синтезуються в гіпоталамусі, а накопичуються в задній частині гіпофізу
- 36. Окситоцин стимулює скорочення гладкої мускулатури матки при пологах, а також відіграє важливу роль в стимуляції лактації.
- 37. Вазопресин часто називають антидіуретичним гормоном (АДГ), так як він контролює реабсорбцію води в ниркових канальцях. Стимулом,
- 38. Гормони периферичних ендокринних залоз Гормони периферичних ендокринних залоз (щитоподібної, прищитоподібних, підшлункової, надниркових, статевих) є регуляторами метаболізму.
- 39. Гормони щитоподібної залози 1). Тиреоїдні гормони – трийодтиронін та тетрайодтиронін (тироксин) – похідні амінокислоти тирозину: 2).
- 40. Стимуляція синтезу тиреоїдних гормонів Гіпоталамус (тиреоліберин) ↓ Передня частка гіпофізу (тиреотропний гормон, ТТГ) ↓ Щитоподібна залоза
- 41. Тиреоїдні гормони (йодтироніни) Діють по цитозольному механізму. Взаємодіючи з ДНК, підсилюють біосинтез багатьох білків, особливо білків-ферментів.
- 42. Гіперфункція (гіпертиреоз) Проявляється у вигляді тиреотоксикозу (токсичний зоб або Базедова хвороба). При цьому спостерігається: Різке схуднення
- 44. Гіпофункція (гіпотиреоз) У молодих організмів, що ростуть розвивається захворювання кретинізм, тобто відставання фізичного та розумового розвитку.
- 47. Тиреокальцитонін – це білок, який містить 32 амінокислотних залишки. Цей гормон разом з гормоном прищитоподібних залоз
- 48. Він інгібує вивільнення Са2+ з кісток, знижуючи активність остеокластів. Швидкість секреції кальцитоніну у жінок сильно залежить
- 49. Гормон прищитоподібних залоз Паратгормон – поліпептид, складається з 84 амінокислотних залишків. Основна дія направлена на підвищення
- 50. Гормони підшлункової залози Ендокринну активність виконує острівцева частина залози (острівці Лангерганса): α-клітини секретують глюкагон, β-клітини –
- 51. Інсулін Білок, що складається з двох поліпептидних ланцюгів, з'єднаних між собою двома дисульфідними містками. Ланцюг А
- 52. Хімічна будова інсуліну
- 53. Видова специфічність інсуліну
- 54. Біохімічні функції інсуліну Це головний анаболічний гормон. Підвищує проникність клітин печінки, нирок, жирової тканини та інших
- 55. Гіпоглікемічна дія інсуліну
- 56. Активує ключовий фермент обміну глюкози – глюкокіназу (гексокіназу): глюкоза + АТФ → глюкозо-6-фосфат + АДФ Глюкозо-6-фосфат
- 57. Дія інсуліну на обмін ліпідів і білків В печінці та жировій тканині стимулює синтез жирів та
- 58. Гіпофункція Нестача інсуліну призводить до розвитку цукрового діабету. Для цього захворювання характерним є: гіперглікемія; глюкозурія; кетонемія;
- 60. Глюкагон Синтезується в α-клітинах острівців Лангерганса. Це поліпептид, складається з 29 а.к. залишків. За характером дії
- 61. Біохімічна дія глюкагону Підвищує рівень глюкози в крові (гіперглікемічна дія). Стимулює глікогеноліз, тобто розпад глікогену в
- 62. Соматостатин Синтезується δ-клітинами острівців Лангерганса, а також в гіпоталамусі. Це поліпептид, складається з 14 а.к. залишків.
- 63. Гормони мозкової речовини надниркових залоз – адреналін і норадреналін.
- 64. За хімічною будовою ці гормони є похідними а.к. тирозину, так як вони містять кільце пірокатехіну, їх
- 65. Фізіологічна дія Адреналін активує фосфорилазу подібно глюкагону, прискорює глікогеноліз, переважно у м'язах, активує глюконеогенез в печінці,
- 66. Гормони кіркової речовини надниркових залоз Всі вони стероїдної природи, тому їх називають кортикостероїди. За характером дії
- 67. Діють вони за цитозольним механізмом, бо їх рецептори розташовані в цитозолі клітини або в ядрі. Регуляція
- 68. Глюкокортикоїди Найбільшу активність проявляє кортизол і кортикостерон.
- 69. Біохімічна дія глюкокортикоїдів Переважно діють на вуглеводний обмін. Підвищують концентрацію глюкози в крові (гіперглікемічна дія). Активують
- 70. Інгібують синтез нуклеїнових кислот у всіх тканинах, окрім печінки. У м'язах, лімфоїдній та жировій тканинах, шкірі
- 71. Гіпофункція кори надниркових залоз проявляється зменшенням вмісту кортикоїдних гормонів і носить назву Аддісонової хвороби (бронзової). Головними
- 74. Мінералокортикоїди Типовий представник цієї групи альдостерон.
- 75. Біохімічна дія мінералокортикоїдів Переважно діють на водно-сольовий обмін. Затримують в організмі іони натрію та хлору, підсилюючи
- 76. Фізіологічна дія кортикостероїдів Проявляють: антизапальну антиалергічну імунодепресивну дію. Тому кортикостероїди застосовуються в якості лікарських препаратів.
- 77. Гормони статевих залоз Синтезуються в статевих залозах або гонадах: - у чоловічих особин – в сім'яниках
- 78. Чоловічі статеві гормони - андрогени Синтезуються, в основному, в чоловічих статевих залозах – в інтерстиціальних клітинах
- 80. Фізіологічна дія андрогенів Уже в ембріональний період відповідають за формування вторинних статевих ознак, в тому числі
- 81. В репродуктивних тканинах відповідають за їх диференціровку та функціонування. Стимулюють статеве дозрівання і сперматогенез. Діють за
- 82. Біохімічна дія андрогенів Контролюють клітинний метаболізм як репродуктивних органів, так і багатьох інших органів і тканин.
- 83. Андрогени також сприяють синтезу фосфоліпідів, що забезпечує формування цитоплазматичних мембран і збільшення загальної кількості клітин в
- 84. Жіночі статеві гормони - естрогени Синтезуються в яєчниках Діляться на дві групи: 1. Власне естрогени (типовий
- 85. Регуляція секреції жіночих статевих гормонів.
- 86. Естрадіол Прогестерон
- 87. Дія естрогенів Діють як я андрогени за цитозольним механізмом. Стимулюють синтез специфічних білків у репродуктивних органах.
- 88. Прогестерон Утворюється в жовтому тілі яєчників, а також у фетоплацентарному комплексі в період вагітності. Діє за
- 89. Простагландини Вперше були виділені з передміхурової залози (prostate) – звідси їх назва. Слугують регуляторами дії гормонів.
- 90. До групи ейкозаноїдів відносять: - простагландини - лейкотрієни - простацикліни - тромбоксани Простагландини регулюють: - скорочувальну
- 91. Простацикліни є підвидом простагландинів (Pg I), але додатково мають особливу функцію - інгібують агрегацію тромбоцитів і
- 92. Синтез ейкозаноїдів
- 93. Гормони шлунково-кишкового тракту Сімейство гастрину: - гастрин - холецистокінін Сімейство секретину: - секретин - ендоглюкагон -
- 94. Окремі представники і їх дія ГАСТРИН Синтезується слизовою шлунка. Має 3 молекулярні форми з різною кількістю
- 95. СЕКРЕТИН Синтезується слизовою 12-типалої кишки. Пептид, складається з 27 а.к. залишків. Діє на клітини підшлункової залози
- 96. МОТИЛІН Синтезується в слизовій кишечника. Складається з 22 а.к. залишків. Стимулює секрецію пепсиногену в шлунку.
- 98. Скачать презентацию

Гормони – органічні речовини дистанційної дії, що виробляються в ендокринних залозах
Гормони – органічні речовини дистанційної дії, що виробляються в ендокринних залозах

За допомогою крові гормони розносяться по всьому організму, але діють тільки
За допомогою крові гормони розносяться по всьому організму, але діють тільки

За хімічною будовою розрізняють:
Білки і пептиди
Похідні окремих амінокислот
Стероїди
За хімічною будовою розрізняють:
Білки і пептиди
Похідні окремих амінокислот
Стероїди



Біологічна роль
1. Змінюють інтенсивність метаболізму
2. Активують (інгібують) ферменти
3. Прискорюють (сповільнюють) синтез
Біологічна роль
1. Змінюють інтенсивність метаболізму
2. Активують (інгібують) ферменти
3. Прискорюють (сповільнюють) синтез

Основні загальні біологічні ознаки:
Дистантність дії – біологічну дію проявляють на відстані
Основні загальні біологічні ознаки:
Дистантність дії – біологічну дію проявляють на відстані
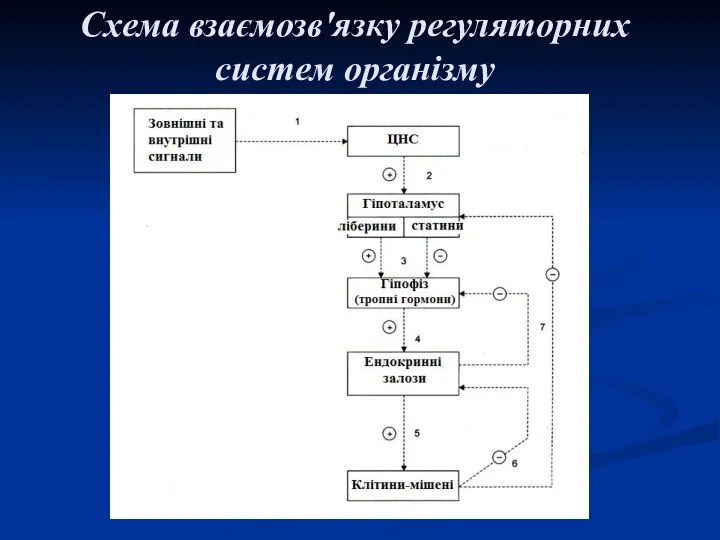
Схема взаємозв'язку регуляторних систем організму
Схема взаємозв'язку регуляторних систем організму

Найбільш важливою є гіпоталамо-гіпофізарна система, яка контролюється центральною нервовою системою. Збудження
Найбільш важливою є гіпоталамо-гіпофізарна система, яка контролюється центральною нервовою системою. Збудження
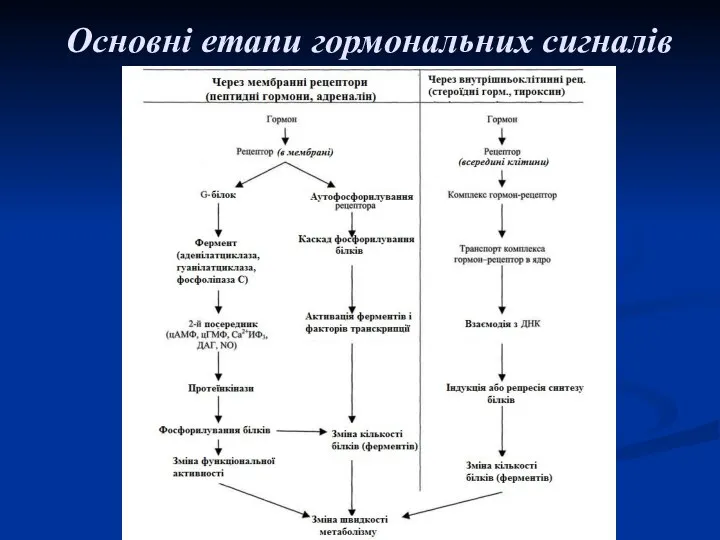
Основні етапи гормональних сигналів
Основні етапи гормональних сигналів

Початковий етап у дії гормону на клітину-мішень – взаємодія гормону з
Початковий етап у дії гормону на клітину-мішень – взаємодія гормону з
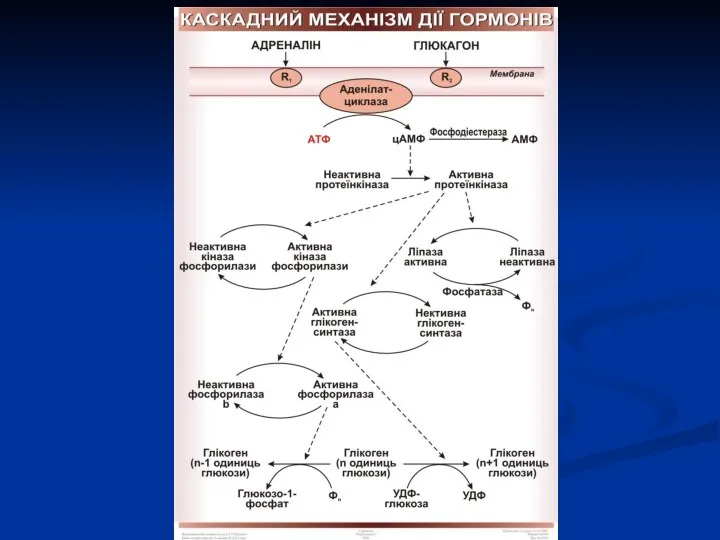

Мембранний або аденілатциклазний механізм (для гормонів білкової і пептидної природи, а
Мембранний або аденілатциклазний механізм (для гормонів білкової і пептидної природи, а
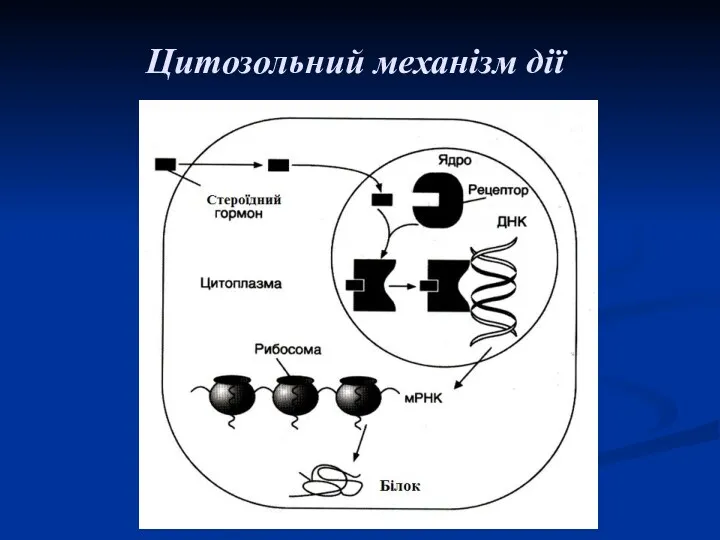
Цитозольний механізм дії
Цитозольний механізм дії

Цитозольний (внутрішньоклітинний)
Є характерним для стероїдних і основної маси тиреоїдних гормонів. Це
Цитозольний (внутрішньоклітинний)
Є характерним для стероїдних і основної маси тиреоїдних гормонів. Це
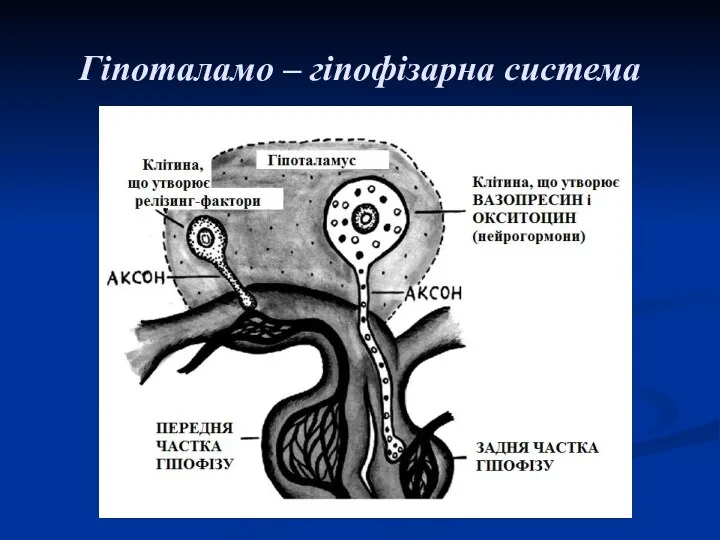
Гіпоталамо – гіпофізарна система
Гіпоталамо – гіпофізарна система


Гормони гіпофізу
Гормони передньої частки гіпофізу
В передній частині гіпофізу синтезуються так
Гормони гіпофізу
Гормони передньої частки гіпофізу
В передній частині гіпофізу синтезуються так
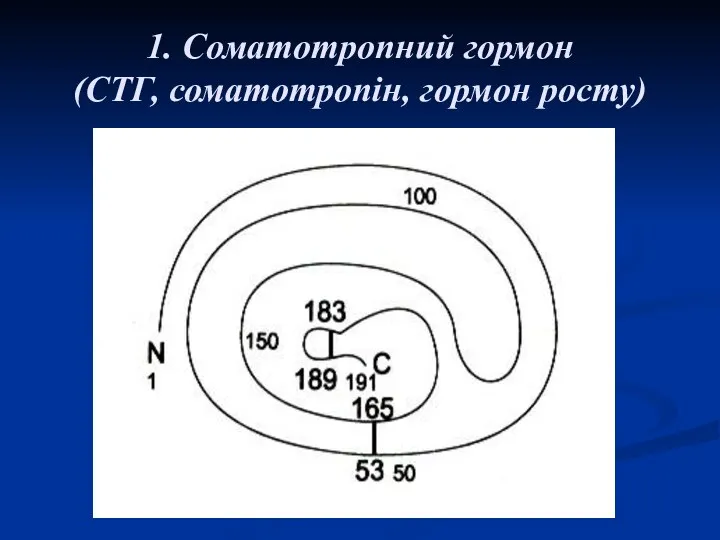
1. Соматотропний гормон
(СТГ, соматотропін, гормон росту)
1. Соматотропний гормон
(СТГ, соматотропін, гормон росту)

Соматотропний гормон –
(СТГ, гормон росту, соматотропін).
Це поліпептид, який складається зі
Соматотропний гормон –
(СТГ, гормон росту, соматотропін).
Це поліпептид, який складається зі
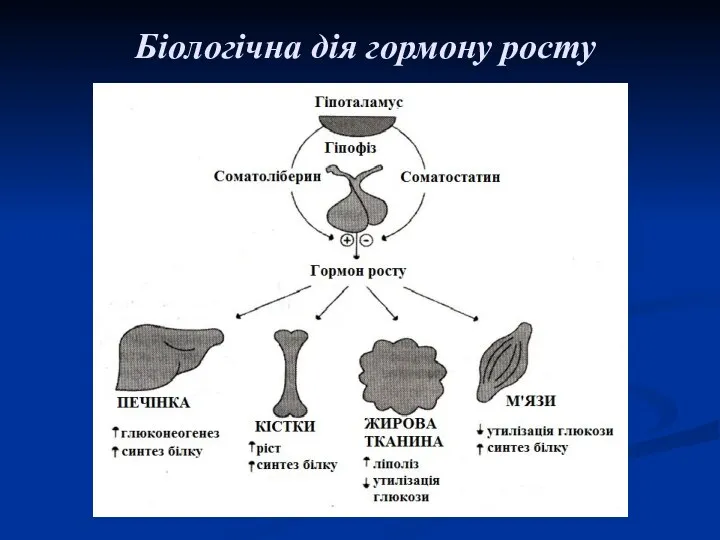
Біологічна дія гормону росту
Біологічна дія гормону росту

Основна дія гормону росту направлена на регуляцію обміну білків і
Основна дія гормону росту направлена на регуляцію обміну білків і

Гіпофункція гіпофізу по СТГ приводить у молодих організмів до карликовості –
Гіпофункція гіпофізу по СТГ приводить у молодих організмів до карликовості –




Акромегалія
Акромегалія

2. Тиреотропний гормон – (ТТГ, тиреотропін).
За хімічною природою це складний білок
2. Тиреотропний гормон – (ТТГ, тиреотропін).
За хімічною природою це складний білок

3. Адренокортикотропний гормон (АКТГ, кортикотропін).
Це поліпептид, який містить 39 амінокислотних залишки.
3. Адренокортикотропний гормон (АКТГ, кортикотропін).
Це поліпептид, який містить 39 амінокислотних залишки.

4. Фолікулостимулюючий гормон (ФСГ, фолітропін).
За хімічною структурою це складний білок –
4. Фолікулостимулюючий гормон (ФСГ, фолітропін).
За хімічною структурою це складний білок –

5. Лютеїнізуючий гормон (ЛГ, лютропін).
Є глікопротеїном.
У самок ЛГ стимулює овуляцію,
5. Лютеїнізуючий гормон (ЛГ, лютропін).
Є глікопротеїном.
У самок ЛГ стимулює овуляцію,

6. Пролактин (ПРЛ, лактотропін).
Це білок, який складається зі 199 амінокислотних залишків,
6. Пролактин (ПРЛ, лактотропін).
Це білок, який складається зі 199 амінокислотних залишків,

Гормон середньої частки гіпофізу
Меланоцит-стимулюючий гормон (МСГ)
Поліпептид, у багатьох тварин складається з
Гормон середньої частки гіпофізу
Меланоцит-стимулюючий гормон (МСГ)
Поліпептид, у багатьох тварин складається з

Гормони задньої частки гіпофізу
Окситоцин і вазопресин
Синтезуються в гіпоталамусі, а накопичуються в
Гормони задньої частки гіпофізу
Окситоцин і вазопресин
Синтезуються в гіпоталамусі, а накопичуються в

Окситоцин стимулює скорочення гладкої мускулатури матки при пологах, а також відіграє
Окситоцин стимулює скорочення гладкої мускулатури матки при пологах, а також відіграє

Вазопресин часто називають антидіуретичним гормоном (АДГ), так як він контролює реабсорбцію
Вазопресин часто називають антидіуретичним гормоном (АДГ), так як він контролює реабсорбцію

Гормони периферичних
ендокринних залоз
Гормони периферичних ендокринних залоз (щитоподібної, прищитоподібних, підшлункової, надниркових,
Гормони периферичних
ендокринних залоз
Гормони периферичних ендокринних залоз (щитоподібної, прищитоподібних, підшлункової, надниркових,
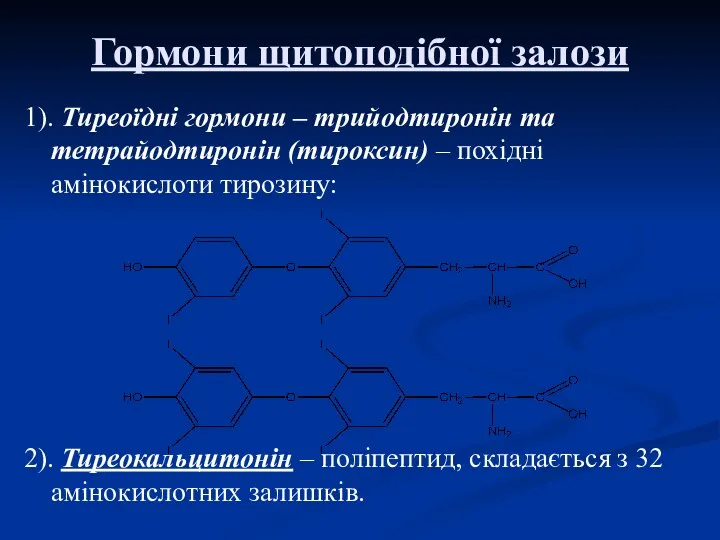
Гормони щитоподібної залози
1). Тиреоїдні гормони – трийодтиронін та тетрайодтиронін (тироксин) –
Гормони щитоподібної залози
1). Тиреоїдні гормони – трийодтиронін та тетрайодтиронін (тироксин) –

Стимуляція синтезу тиреоїдних гормонів
Гіпоталамус (тиреоліберин)
↓
Передня частка гіпофізу
(тиреотропний гормон, ТТГ)
↓
Щитоподібна залоза
Стимуляція синтезу тиреоїдних гормонів
Гіпоталамус (тиреоліберин)
↓
Передня частка гіпофізу
(тиреотропний гормон, ТТГ)
↓
Щитоподібна залоза

Тиреоїдні гормони
(йодтироніни)
Діють по цитозольному механізму.
Взаємодіючи з ДНК, підсилюють біосинтез багатьох
Тиреоїдні гормони
(йодтироніни)
Діють по цитозольному механізму.
Взаємодіючи з ДНК, підсилюють біосинтез багатьох

Гіперфункція (гіпертиреоз)
Проявляється у вигляді тиреотоксикозу (токсичний зоб або Базедова хвороба).
При
Гіперфункція (гіпертиреоз)
Проявляється у вигляді тиреотоксикозу (токсичний зоб або Базедова хвороба).
При
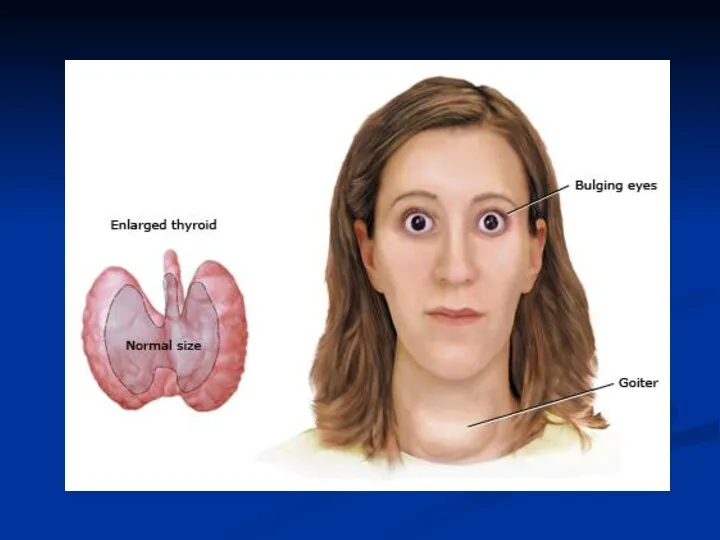

Гіпофункція (гіпотиреоз)
У молодих організмів, що ростуть розвивається захворювання кретинізм, тобто відставання
Гіпофункція (гіпотиреоз)
У молодих організмів, що ростуть розвивається захворювання кретинізм, тобто відставання

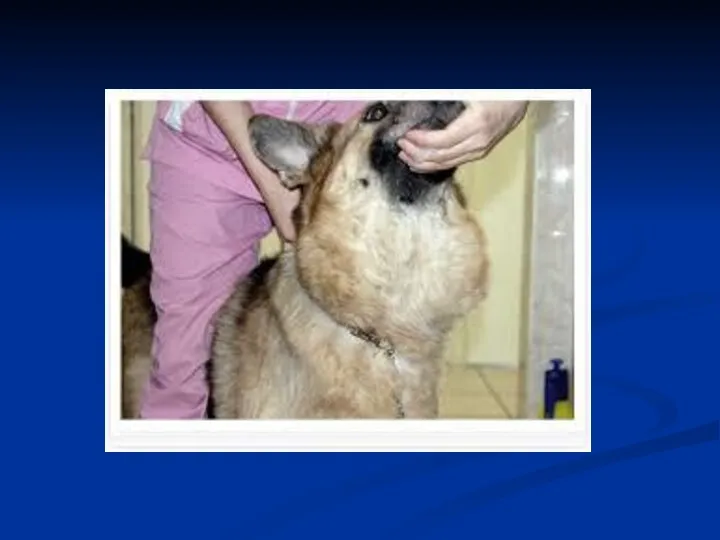

Тиреокальцитонін – це білок, який містить 32 амінокислотних залишки.
Цей гормон
Тиреокальцитонін – це білок, який містить 32 амінокислотних залишки.
Цей гормон

Він інгібує вивільнення Са2+ з кісток, знижуючи активність остеокластів.
Швидкість секреції кальцитоніну
Він інгібує вивільнення Са2+ з кісток, знижуючи активність остеокластів.
Швидкість секреції кальцитоніну

Гормон прищитоподібних залоз
Паратгормон – поліпептид, складається з 84 амінокислотних залишків.
Основна дія
Гормон прищитоподібних залоз
Паратгормон – поліпептид, складається з 84 амінокислотних залишків.
Основна дія

Гормони підшлункової залози
Ендокринну активність виконує острівцева частина залози (острівці Лангерганса):
α-клітини
Гормони підшлункової залози
Ендокринну активність виконує острівцева частина залози (острівці Лангерганса):
α-клітини

Інсулін
Білок, що складається з двох поліпептидних ланцюгів, з'єднаних між собою двома
Інсулін
Білок, що складається з двох поліпептидних ланцюгів, з'єднаних між собою двома
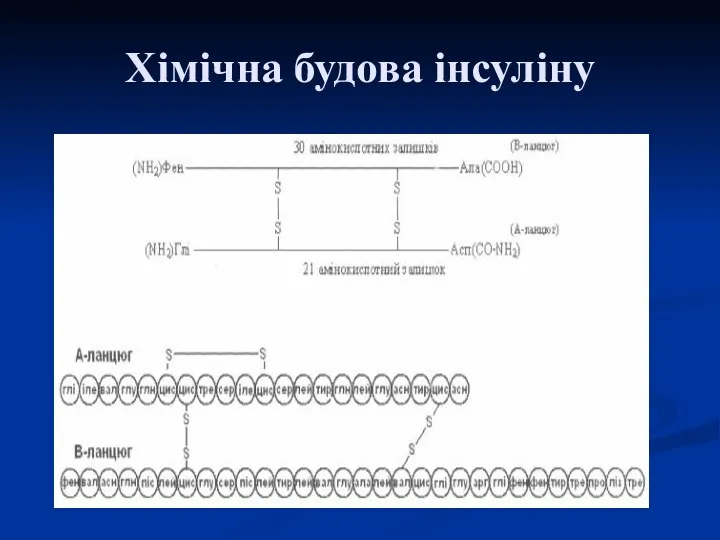
Хімічна будова інсуліну
Хімічна будова інсуліну

Видова специфічність інсуліну
Видова специфічність інсуліну

Біохімічні функції інсуліну
Це головний анаболічний гормон.
Підвищує проникність клітин печінки, нирок, жирової
Біохімічні функції інсуліну
Це головний анаболічний гормон.
Підвищує проникність клітин печінки, нирок, жирової
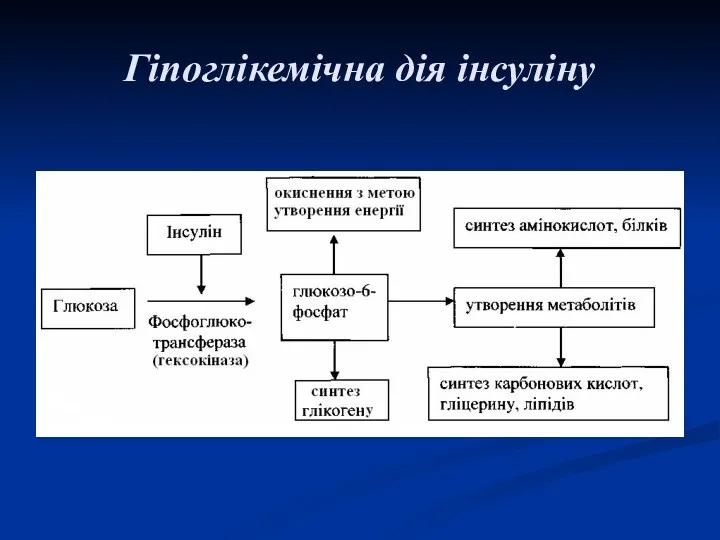
Гіпоглікемічна дія інсуліну
Гіпоглікемічна дія інсуліну

Активує ключовий фермент обміну глюкози – глюкокіназу (гексокіназу):
глюкоза + АТФ
Активує ключовий фермент обміну глюкози – глюкокіназу (гексокіназу):
глюкоза + АТФ

Дія інсуліну на обмін ліпідів і білків
В печінці та жировій тканині
Дія інсуліну на обмін ліпідів і білків
В печінці та жировій тканині

Гіпофункція
Нестача інсуліну призводить до розвитку
цукрового діабету.
Для цього захворювання характерним є:
гіперглікемія;
глюкозурія;
кетонемія;
кетонурія;
ацидоз;
азотемія;
азотурія;
поліурія;
полідипсія
Гіпофункція
Нестача інсуліну призводить до розвитку
цукрового діабету.
Для цього захворювання характерним є:
гіперглікемія;
глюкозурія;
кетонемія;
кетонурія;
ацидоз;
азотемія;
азотурія;
поліурія;
полідипсія


Глюкагон
Синтезується в α-клітинах острівців Лангерганса.
Це поліпептид, складається з 29 а.к. залишків.
За
Глюкагон
Синтезується в α-клітинах острівців Лангерганса.
Це поліпептид, складається з 29 а.к. залишків.
За

Біохімічна дія глюкагону
Підвищує рівень глюкози в крові (гіперглікемічна дія).
Стимулює глікогеноліз, тобто
Біохімічна дія глюкагону
Підвищує рівень глюкози в крові (гіперглікемічна дія).
Стимулює глікогеноліз, тобто

Соматостатин
Синтезується δ-клітинами острівців Лангерганса, а також в гіпоталамусі.
Це поліпептид, складається з
Соматостатин
Синтезується δ-клітинами острівців Лангерганса, а також в гіпоталамусі.
Це поліпептид, складається з
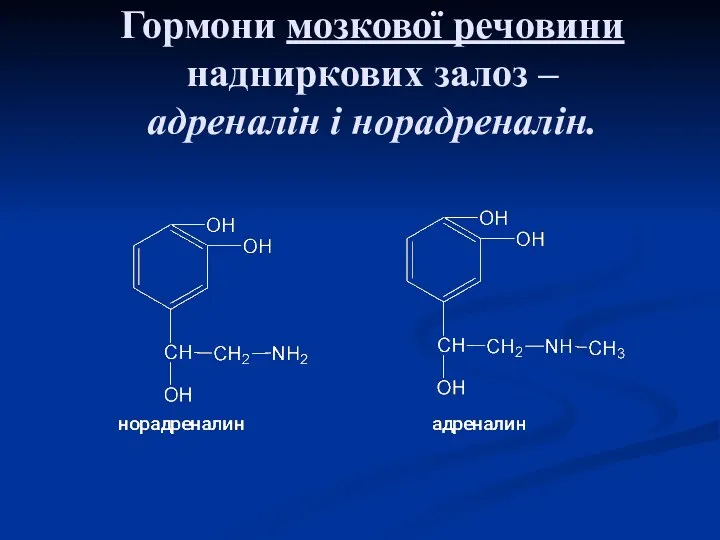
Гормони мозкової речовини надниркових залоз –
адреналін і норадреналін.
Гормони мозкової речовини надниркових залоз –
адреналін і норадреналін.

За хімічною будовою ці гормони є похідними а.к. тирозину, так як
За хімічною будовою ці гормони є похідними а.к. тирозину, так як

Фізіологічна дія
Адреналін активує фосфорилазу подібно глюкагону, прискорює глікогеноліз, переважно у
Фізіологічна дія
Адреналін активує фосфорилазу подібно глюкагону, прискорює глікогеноліз, переважно у

Гормони кіркової речовини надниркових залоз
Всі вони стероїдної природи, тому їх називають
Гормони кіркової речовини надниркових залоз
Всі вони стероїдної природи, тому їх називають

Діють вони за цитозольним механізмом, бо їх рецептори розташовані в цитозолі
Діють вони за цитозольним механізмом, бо їх рецептори розташовані в цитозолі
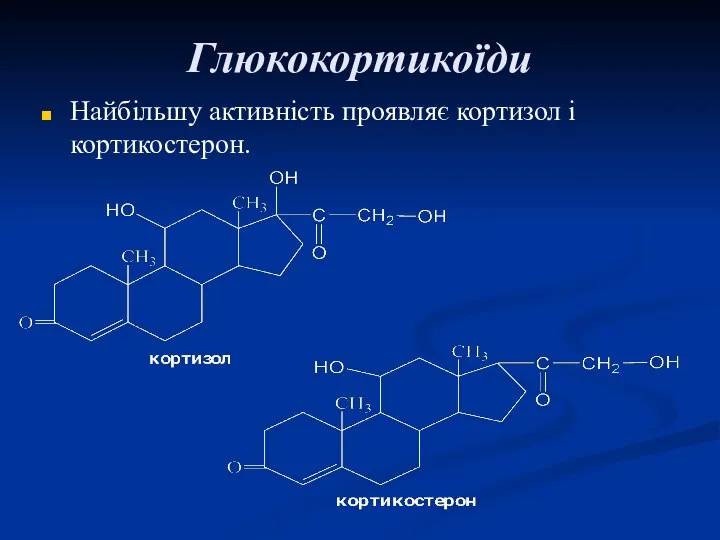
Глюкокортикоїди
Найбільшу активність проявляє кортизол і кортикостерон.
Глюкокортикоїди
Найбільшу активність проявляє кортизол і кортикостерон.

Біохімічна дія глюкокортикоїдів
Переважно діють на вуглеводний обмін.
Підвищують концентрацію глюкози в крові
Біохімічна дія глюкокортикоїдів
Переважно діють на вуглеводний обмін.
Підвищують концентрацію глюкози в крові

Інгібують синтез нуклеїнових кислот у всіх тканинах, окрім печінки.
У м'язах, лімфоїдній
Інгібують синтез нуклеїнових кислот у всіх тканинах, окрім печінки.
У м'язах, лімфоїдній

Гіпофункція кори надниркових залоз проявляється зменшенням вмісту кортикоїдних гормонів і носить
Гіпофункція кори надниркових залоз проявляється зменшенням вмісту кортикоїдних гормонів і носить

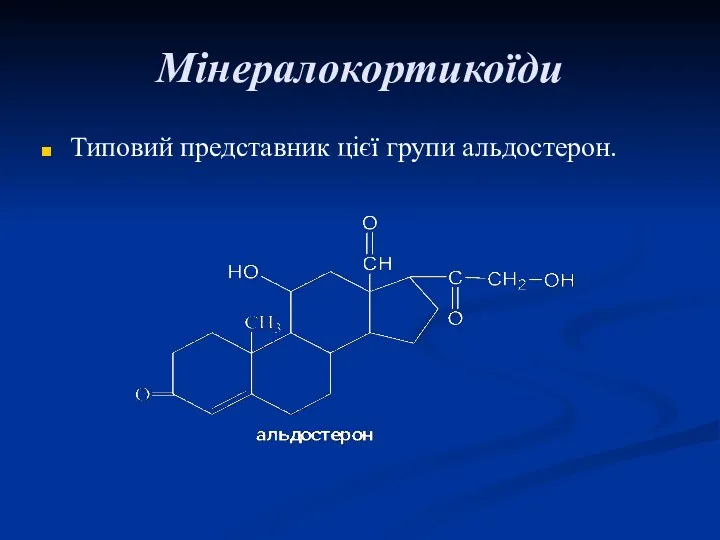
Мінералокортикоїди
Типовий представник цієї групи альдостерон.
Мінералокортикоїди
Типовий представник цієї групи альдостерон.

Біохімічна дія мінералокортикоїдів
Переважно діють на водно-сольовий обмін.
Затримують в організмі іони натрію
Біохімічна дія мінералокортикоїдів
Переважно діють на водно-сольовий обмін.
Затримують в організмі іони натрію

Фізіологічна дія кортикостероїдів
Проявляють:
антизапальну
антиалергічну
імунодепресивну дію.
Тому кортикостероїди застосовуються в якості лікарських препаратів.
Фізіологічна дія кортикостероїдів
Проявляють:
антизапальну
антиалергічну
імунодепресивну дію.
Тому кортикостероїди застосовуються в якості лікарських препаратів.

Гормони статевих залоз
Синтезуються в статевих залозах або гонадах:
- у чоловічих
Гормони статевих залоз
Синтезуються в статевих залозах або гонадах:
- у чоловічих

Чоловічі статеві гормони - андрогени
Синтезуються, в основному, в чоловічих статевих залозах
Чоловічі статеві гормони - андрогени
Синтезуються, в основному, в чоловічих статевих залозах
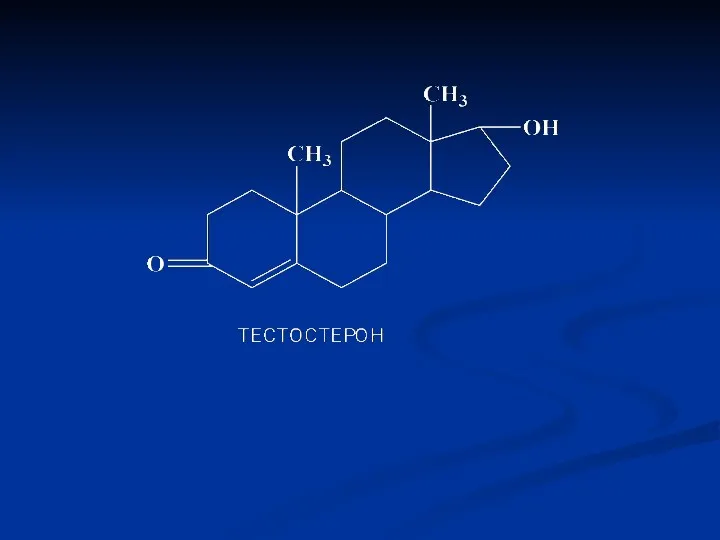

Фізіологічна дія андрогенів
Уже в ембріональний період відповідають за формування вторинних статевих
Фізіологічна дія андрогенів
Уже в ембріональний період відповідають за формування вторинних статевих

В репродуктивних тканинах відповідають за їх диференціровку та функціонування.
Стимулюють статеве дозрівання
В репродуктивних тканинах відповідають за їх диференціровку та функціонування.
Стимулюють статеве дозрівання

Біохімічна дія андрогенів
Контролюють клітинний метаболізм як репродуктивних органів, так і багатьох
Біохімічна дія андрогенів
Контролюють клітинний метаболізм як репродуктивних органів, так і багатьох

Андрогени також сприяють синтезу фосфоліпідів, що забезпечує формування цитоплазматичних мембран і
Андрогени також сприяють синтезу фосфоліпідів, що забезпечує формування цитоплазматичних мембран і

Жіночі статеві гормони - естрогени
Синтезуються в яєчниках
Діляться на дві групи:
1. Власне
Жіночі статеві гормони - естрогени
Синтезуються в яєчниках
Діляться на дві групи:
1. Власне
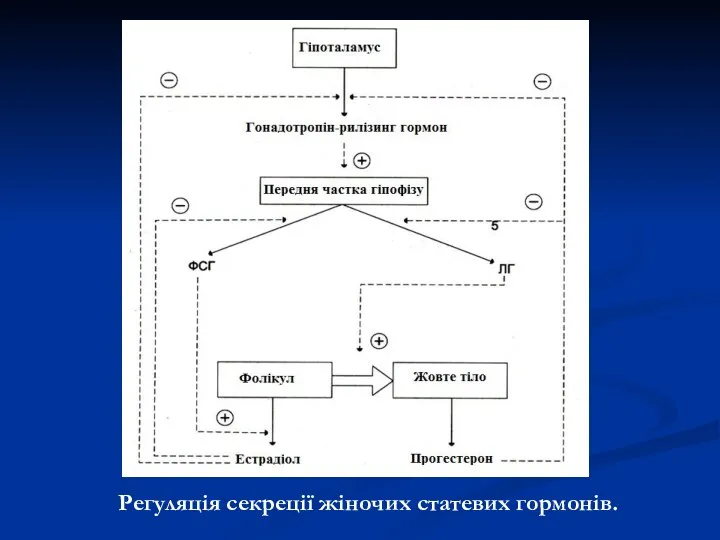
Регуляція секреції жіночих статевих гормонів.
Регуляція секреції жіночих статевих гормонів.
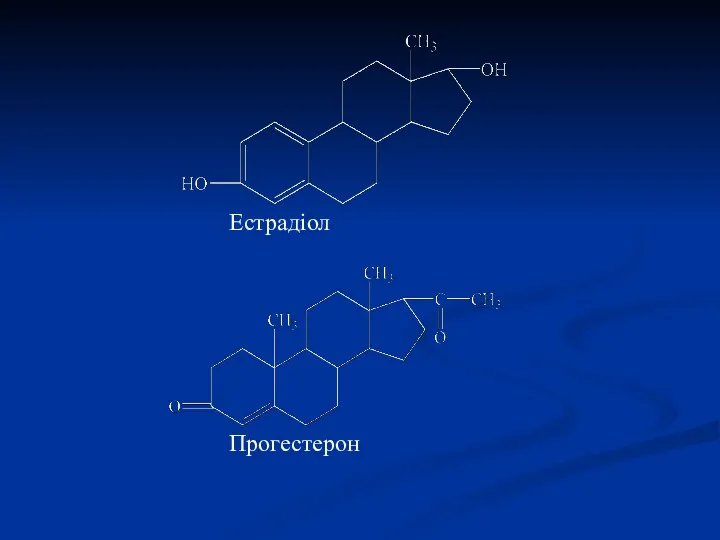
Естрадіол
Прогестерон
Естрадіол
Прогестерон

Дія естрогенів
Діють як я андрогени за цитозольним механізмом.
Стимулюють синтез специфічних білків
Дія естрогенів
Діють як я андрогени за цитозольним механізмом.
Стимулюють синтез специфічних білків

Прогестерон
Утворюється в жовтому тілі яєчників, а також у фетоплацентарному комплексі в
Прогестерон
Утворюється в жовтому тілі яєчників, а також у фетоплацентарному комплексі в

Простагландини
Вперше були виділені з передміхурової залози (prostate) – звідси їх назва.
Слугують
Простагландини
Вперше були виділені з передміхурової залози (prostate) – звідси їх назва.
Слугують

До групи ейкозаноїдів відносять:
- простагландини
- лейкотрієни
- простацикліни
- тромбоксани
Простагландини регулюють:
- скорочувальну функцію
До групи ейкозаноїдів відносять:
- простагландини
- лейкотрієни
- простацикліни
- тромбоксани
Простагландини регулюють:
- скорочувальну функцію

Простацикліни є підвидом простагландинів (Pg I), але додатково мають особливу функцію - інгібують
Простацикліни є підвидом простагландинів (Pg I), але додатково мають особливу функцію - інгібують
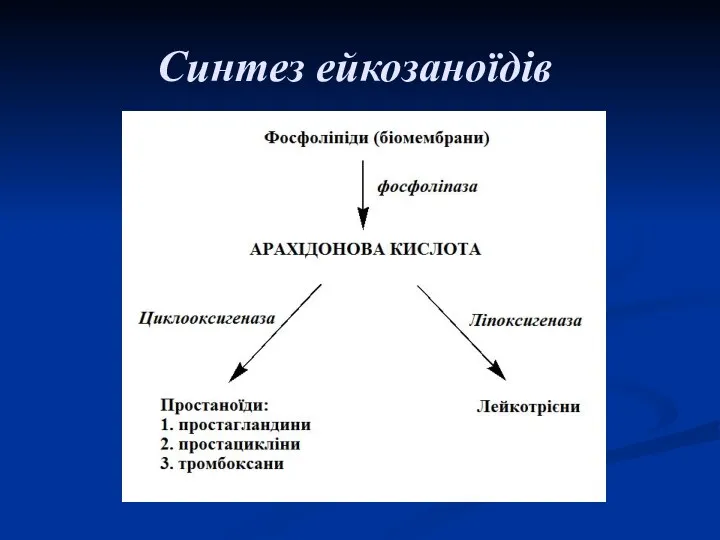
Синтез ейкозаноїдів
Синтез ейкозаноїдів

Гормони шлунково-кишкового тракту
Сімейство гастрину:
- гастрин
- холецистокінін
Сімейство секретину:
- секретин
- ендоглюкагон
- вазоактивний
Гормони шлунково-кишкового тракту
Сімейство гастрину:
- гастрин
- холецистокінін
Сімейство секретину:
- секретин
- ендоглюкагон
- вазоактивний

Окремі представники і їх дія
ГАСТРИН
Синтезується слизовою шлунка.
Має 3 молекулярні форми з
Окремі представники і їх дія
ГАСТРИН
Синтезується слизовою шлунка.
Має 3 молекулярні форми з

СЕКРЕТИН
Синтезується слизовою 12-типалої кишки.
Пептид, складається з 27 а.к. залишків.
Діє на клітини
СЕКРЕТИН
Синтезується слизовою 12-типалої кишки.
Пептид, складається з 27 а.к. залишків.
Діє на клітини

МОТИЛІН
Синтезується в слизовій кишечника.
Складається з 22 а.к. залишків.
Стимулює секрецію пепсиногену в
МОТИЛІН
Синтезується в слизовій кишечника.
Складається з 22 а.к. залишків.
Стимулює секрецію пепсиногену в
 Насекомые. Отгадайте загадки
Насекомые. Отгадайте загадки Игра-путешествие На лесной полянке
Игра-путешествие На лесной полянке Анатомия соединений костей туловища и головы
Анатомия соединений костей туловища и головы Жизненные циклы
Жизненные циклы Мотивация обучения
Мотивация обучения Мед и пчелы
Мед и пчелы Болезни птиц
Болезни птиц Развитие интереса на уроках биологии
Развитие интереса на уроках биологии Анализаторы. Органы чувств, их роль в организме. Строение и функции. Часть 2
Анализаторы. Органы чувств, их роль в организме. Строение и функции. Часть 2 Көру анализаторлары
Көру анализаторлары Класс Пресмыкающиеся
Класс Пресмыкающиеся Этология с основами зоопсихологии
Этология с основами зоопсихологии Общая и медицинская гельминтология
Общая и медицинская гельминтология Головоногие моллюски
Головоногие моллюски Тип Plathelminthes – плоские черви. Класс Cestoda – ленточные черви
Тип Plathelminthes – плоские черви. Класс Cestoda – ленточные черви Скелет туловища. Соединение позвонков
Скелет туловища. Соединение позвонков Evaluation of Tubocurarine ED50 on Electrically Stimulated Isolated Chick Biventer Cervicis Nerve-Muscle Preparation
Evaluation of Tubocurarine ED50 on Electrically Stimulated Isolated Chick Biventer Cervicis Nerve-Muscle Preparation презентация на тему:Удивительные животные
презентация на тему:Удивительные животные Красная книга Таштагольского района
Красная книга Таштагольского района Animals in danger. About tigers
Animals in danger. About tigers Шиншиллы и всё о них
Шиншиллы и всё о них Голонасінні. Кедр
Голонасінні. Кедр Экологиялық факторлар
Экологиялық факторлар Эмбриональное развитие человека
Эмбриональное развитие человека Онтогенез – индивидуальное развитие организма
Онтогенез – индивидуальное развитие организма Эволюция дыхательной системы
Эволюция дыхательной системы Геномика и транскриптомика
Геномика и транскриптомика Биологиялық объектілердің люминесценциясы
Биологиялық объектілердің люминесценциясы