- Характеристика нервной системы человека

Содержание
- 2. 1.Морфологическая и функциональная характеристика нервной системы человека. 2. Нервная ткань. 3.Строение, типы, функции нейронов. План
- 3. Функции нервной системы ▪ интегративно-координационная функция: обеспечивает функции различных органов и физиологических систем, согласует их деятельность
- 4. Основным принципом функционирования ЦНС является процесс регуляции, управления физиологическими функциями, которые направлены на поддержание постоянства свойств
- 5. Морфологическая и функциональная характеристика нервной системы человека Нервная система объединяет организм человека в единое целое, регулирует
- 6. В мозге человека происходят сложные процессы обработки поступившей в него информации. В результате из мозга также
- 7. Нервная система обеспечивает эффективное приспособление организма к изменениям окружающей среды, формирует целенаправленное поведение. Она представлена спинным,
- 8. Число нервных элементов, будучи очень ограниченным у примитивных организмов, в процессе эволюционного развития нервной системы достигает
- 9. Различают два основных вида регуляции: гуморальный и нервный Нервный процесс регуляции предусматривает управление изменения физиологических функций
- 10. Центральная нервная система (ЦНС) Периферическая нервная система Головной мозг Спинной мозг нервы Нервные узлы Нервные окончания
- 11. рвнанея система Регулирует работу скелетных мышц Регулирует работу внутренних органов Функциональное деление нервной системы Нервная система
- 12. Вегетативная нервная система Симпатическая Парасимпатическая Способствует восстановлению запасов энергии во время сна и отдыха Включается во
- 13. ИСТОЧНИК РАЗВИТИЯ – эктодерма (нейроэктодерма): нервная пластинка – нервные валики – нервный желобок – нервная трубка
- 14. НЕРВНАЯ ТРУБКА Вентрикулярная (эпендимная) зона ▪ делящиеся клетки цилиндрической формы. Промежуточная (плащевая, мантийная) зона ▪ нейробласты
- 15. ОБЩИЕ ПРИНЦИПЫ СТРОЕНИЯ 1. Большинство нервных клеток являются длинноотросчатыми. 2. Нейроны характеризуются полярностью. 3. Наличие межклеточных
- 16. НЕРВНЫЕ ТКАНИ (от греческого neuron – жила, струна, нейрон) – это система взаимосвязанных нервных клеток и
- 17. Нервная ткань Основу нервной ткани составляют нервные клетки – НЕЙРОНЫ
- 18. Основным структурным элементом нервной системы является нейрон. Через нейроны осуществляется передача информации от одного участка нервной
- 19. Функции нейронов 1) генерализация нервного импульса; 2) получение, хранение и передача информации; 3) способность суммировать возбуждающие
- 20. Структурно-функциональной единицей нервной системы является нейрон - специализированная клетка, способная принимать, обрабатывать, кодировать, передавать и хранить
- 21. Гибель (апоптоз) нейронов в физиологических условиях у взрослого человека сравнительно невелика и осуществляется механизмом апоптоза. Избыточной
- 22. Нейрон развивается из небольшой клетки - предшественницы, которая перестает делиться еще до того, как выпустит свои
- 23. Среди нейронов встречаются самые крупные клеточные элементы организма. Размеры их поперечника колеблются от 6-7 мк (мелкие
- 24. . В нейроне выделяют: Воспринимающую часть. Дендриты – основное воспринимающее поле нейрона. Мембрана дендрита способна реагировать
- 25. Строение нейрона дендриты тело нейрона аксон
- 27. МОРФОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА НЕЙРОНОВ
- 28. Строение перикариона (тела) нейроцита Плазмолемма (обеспечивает проведение нервного импульса) - ионные каналы Перикарион: - ядро -
- 29. Перикарион, или сома нейрона, имеет клеточную мембрану и содержит ядро, рибосомы, лизосомы, вещество Ниссля, аппарат Гольджи,
- 30. Ядро в большинстве нейронов расположено в центре тела клетки. Как правило, ядро крупное, сферической формы, светлое,
- 31. В нейронах хорошо развита гранулярная эндоплазматическая сеть, что говорит о высокой интенсивности синтетических процессов. Важной особенностью
- 32. У взрослых людей наиболее крупных размеров тельца Ниссля достигают в мотонейронах. Показано, что при длительном раздражении
- 33. Комплекс, или аппарат, Гольджи хорошо развит в нейронах (его первое описание было дано на основе исследования
- 34. Лизосомальный аппарат, или аппарат внутриклеточного переваривания, нейронов обладает высокой активностью и представлен эндосомами и многочисленными лизосомами
- 35. Пигменты (включения) нейронов представлены липофусцином и меланином. Липофусцин - это желто-коричневый пигмент, который находится в нейроне
- 36. Белки нейронов служат для пластических и информационных целей. В ядре нейрона содержится ДНК, в цитоплазме преобладает
- 37. Строение нейрона: Я - ядро. Яд. - ядрышко, М - митохондрии, Р - отдельные рибосомы и
- 38. Строение отростков Аксон у нейрона всегда один маловетвящийся (осевой) толстый отросток, чаще длинный, отсутствует тигроид, проводит
- 39. Рассмотрим морфологические особенности дендритов. Отростки, по которым к телу нервной клетки поступает возбуждение от рецепторов, называются
- 40. Дендриты проводят импульсы к телу нейрона, получая сигналы от других нейронов через многочисленные межнейронные контакты (аксо-дендритические
- 41. Аксон (от лат. аксис - ось), или нейрит, - это центральный, или осевой, отросток нейрона, по
- 42. Характеристика аксоплазмы аксона В крупных нейронах аксон может содержать до 99 % объема цитоплазмы нейрона. Центральная
- 43. Аксон может по своему ходу давать ответвления, или коллатерали (от лат. collateralis - боковой). Коллатеральные ветви,
- 44. На основании числа и расположения отростков нейроны делятся униполярные псевдоунипо лярные биполярные мультиполярные
- 45. Существует большое многообразие нейронов ЦНС. Поэтому предложены и различные варианты их классификации. Чаще всего эта классификация
- 46. Биполярные нейроны (синонимы - двухотросчатые, или двухполюсные, нейроны) имеют два отростка - аксон и дендрит, обычно
- 47. Мультиполярные нейроны имеют один аксон и много (2 и более) дендритов. Они наиболее распространены в нервной
- 48. Чувствительные нейроны проводят импульс от рецепторов – нервных окончаний в ЦНС Двигательные проводят импульс из ЦНС
- 49. Эфферентные нейроны связаны с передачей нисходящих влияний от вышележащих этажей нервной системы к нижележащим или из
- 50. Функциональная классификация нейронов разделяет их по характеру выполняемой ими функции (в соответствии с их местом в
- 51. Моносенсорные нейроны подразделяют функционально по их чувствительности к разным качествам одного раздражителя. Так, отдельные нейроны слуховой
- 52. Эфферентные нейроны (синонимы - двигательные, моторные, секреторные, центробежные, сердечные, сосудодвигательные и пр.) предназначены для передачи информации
- 53. Вставочные нейроны (синонимы - интернейроны, контактные, ассоциативные, коммуникативные, объединяющие, замыкательные, проводниковые, кондукторные) осуществляют передачу нервного импульса
- 54. Биохимическая классификация нейронов основана на химических особенностях нейромедиаторов, используемых нейронами в синаптической передаче нервных импульсов. Выделяют
- 55. Другие виды классификаций нейронов. Нервные клетки разных отделов нервной системы могут быть активными вне воздействия, т.е.
- 57. нейроглия Нейроглия - комплекс клеточных элементов, выполняющих в нервной ткани опорную, разграничительную, трофическую, секреторную и защитную
- 58. НЕЙРОГЛИЯ (греческое neuron – нерв, glia – клей) – термин, введенный для описания связующих элементов между
- 59. опорная трофическая разграничительная поддержание постоянства среды вокруг нейронов секреторная защитная ФУНКЦИИ НЕЙРОГЛИИ
- 60. Классификация нейроглии Микроглиальные клеткиМикроглиальные клетки, хоть и входят в понятие «глия», не являются собственно нервной тканью,
- 61. Виды астроцитов Различные виды нейроглии: а - плазматические астроциты; б - волокнистые астроциты; в - олигодендроглиоциты;
- 62. Морфология нейроглии Нейроглия - это обширная разнородная группа клеток (глиоцитов, или глиальных клеток) нервной ткани, обеспечивающая
- 63. Глиальные клетки по размерам в 3-4 раза меньше, чем нейроны. В мозге человека содержание глиоцитов в
- 64. На долю астроцитов (или звездчатых глиальных клеток) приходится около 40% от всех глиоцитов. Астроциты - это
- 65. Астроциты выполняют четыре основные функции - опорную, разграничительную (транспортную и барьерную), метаболическую (регуляторную) и защитную (иммунную
- 66. Разграничительная функция астроцитов, которая включает транспортную и барьерную функции, направлена на создание оптимального микроокружения нейронов. Эта
- 67. Метаболическая функция астроцитов, включающая в себя и регуляторную функцию - это одна из наиболее важных функций
- 68. Защитная (иммунная и репаративная) функция астроцитов заключается в формировании различных защитных реакций при повреждении нервной ткани.
- 69. Олигодендроциты - это обширная группа разнообразных мелких клеток с короткими немногочисленными отростками. Олигодендроцитов в коре больших
- 70. Олигодендроциты выполяют две основные функции - 1) образование миелина как компонента изолирующей оболочки у нервных волокон
- 71. Эпендимоциты - еще одна разновидность глиоцитов. Они образуют эпендимную глию, или эпендиму (от греч. ependyma -
- 72. Микроглия представляет собой совокупность мелких удлиненных звездчатых клеток (микроглиоцитов) с плотной цитоплазмой и сравнительно короткими немногочисленными
- 73. Основная функция микроглии - защитная (в том числе иммунная). Клетки микроглии - это, по сути, специализированные
- 74. Клетки-сателлиты (мантийные клетки) охватывают тела нейронов в спинальных, черепно-мозговых и вегетативных ганглиях. Они имеют уплощенную форму,
- 75. Нейропиль - это понятие морфофункциональное. Оно отражает пространство мозговой ткани, расположенное между телами нейронов, отростками нейронов,
- 76. Нервы – скопления отростков нейронов вне ЦНС, заключённые в общую оболочку и проводящие нервные импульсы Нервные
- 77. Нервом называется анатомическое образование, состоящее из нервных волокон, имеющих общую оболочку. Нервы образованы отростками нервных клеток,
- 78. Нервы бывают различной длины и толщины, более длинные нервы расположены в тканях конечностей, особенно нижних. Самым
- 79. нервы двигатель ные чувствительные Вегетатив ные смешанные
- 80. Двигательные нервы состоят из волокон клеток передних рогов спинного мозга или из ядер ствола мозга (глазодвигательный,
- 81. Аксон, заключенный в глиальную оболочку, называется нервным волокном
- 82. Совокупность нервных волокон образует нервные пучки, совокупность которых, в свою очередь, формирует нервный ствол, или нерв.
- 83. Все аксоны покрыты глиальной оболочкой, однако эта оболочка устроена по разному - в одних случаях она
- 84. Безмиелиновые нервные волокна у взрослого располагаются преимущественно в составе вегетативной нервной системы и характеризуются сравнительно низкой
- 85. Миелиновые нервные волокна - это большая часть всех нервных волокон. Они встречаются в ЦНС и в
- 86. Миелиновая оболочка на 80 % состоит из липидов, обладающих высоким омическим сопротивлением, и на 20 %
- 87. По длине волокна миелиновая оболочка имеет прерывистый ход, благодаря чему формируются узловые перехваты, или перехваты Ранвье.
- 88. Перехваты Ранвье играют важную роль в процессе проведения возбуждения по нервному волокну как в ЦНС, так
- 89. Критерием структурно-функциональной зрелости мякотных и безмякотных нервных волокон является увеличение их толщины и уменьшение проницаемости клеточной
- 90. В безмякотных волокнах распределение ионных каналов остается равномерным по всей длине волокна. К моменту рождения двигательные
- 91. Мембрана аксона разделяет жидкие среды, различающиеся по содержанию ионов натрия и калия (в жидкой среде, окружающей
- 92. Распространению нервного импульса вдоль аксона способствует локализованное поступление внутрь аксона ионов натрия с последующим выходом ионов
- 93. Возбудимость нервных волокон плода и новорожденного значительно ниже, чем у взрослого, но с 3-месячного возраста она
- 94. Фазовые изменения возбудимости во время возбуждения в раннем постнатальном онтогенезе также имеют особенности. В частности, длительность
- 95. Скорость распространения возбуждения по нервным волокнам у детей становится такой же, как у взрослых, к 5-9
- 96. Синапс - Место контакта нейронов друг с другом и с другими клетками Пузырьки с медиатором Синаптическая
- 98. Механизм работы синапса Из пресинаптического окончания вниз по аксону нейрона проходит электрический заряд от передающей клетки
- 99. Термин и понятие «синапс», «синаптическая передача» был введен в физиологию Ч. Шеррингтоном в 1897 году. Исследуя
- 100. 1. По локализации - центральные (головной и спинной мозг) и периферические (нервно-мышечный, нейросекреторный, синапс вегетативной нервной
- 101. а) по форме контакта - терминальные (колбообразные соединения) и проходящие (варикозные расширения аксона); б) по природе
- 102. Общие представления о строении и механизмах функционирования химического синапса Химические синапсы (далее по тексту - синапсы)
- 103. Пресинаптическая часть (или пресинаптический элемент, пресинпас) образуется аксоном по его ходу (проходящий синапс) или представляет собой
- 104. Постсинаптическая часть (или постсинаптический элемент) представлена постсинаптической мембраной, содержащей особые комплексы интегральных белков - синаптические рецепторы
- 105. В литературе существует большое разнообразие в названиях синапсов. Например, синаптическая бляшка - это синапс между нейронами,
- 106. Кроме того, в ряде синапсов имеются метаботропные рецепторы, при активации которых (за счет вторичных посредников и
- 107. Одновременно, выделившийся в синаптическую щель медиатор может взаимодействовать с рецепторами, расположенными на пресинаптической мембране. Таким способом
- 108. Рассмотрим, каким образом происходит активация постсинаптических рецепторов, а также вторичные посредники, принимающие участие в передаче нервного
- 109. При активации ионотропных рецепторов, содержащих внутри себя ионные каналы, происходит изменение проницаемости этих каналов, что в
- 110. Для различных синапсов существует своя последовательность передачи сигнала (медиатор, G-белок, вторичный посредник, протеинкиназа, белок-эффектор). Например, для
- 111. Схема строения синапса
- 112. Локализация медиаторов и соответствующих нейронов ЦНС В разных отделах ЦНС расположены нейроны, имеющие различные медиаторы. Ацетилхолин
- 113. Гистамин является медиатором нейронов сосцевидых телец гипоталамуса. Аксоны гистаминергических нейронов проецируются в кору мозга, таламус, базальные
- 114. Соматостат - гипоталамус и другие отделы мозга, желатинозная субстанция, сетчатка. Гонадолиберин - гипоталамус, хеморецептивные зоны желудочков
- 115. Синапсы на нейроне
- 116. Нервно-мышечный синапс Структурно-функциональное созревание нервно-мышечных синапсов охватывает почти весь период антенатального и раннего постнатального периодов развития.
- 117. В процессе развития усиливается синтез ацетилхолина в мотонейронах, возрастает количество активных зон в пресинаптическом окончании и
- 118. На постсинаптической мембране образуются складки, происходит концентрация холинорецепторов в синаптической зоне, в результате чего увеличивается амплитуда
- 119. Строение нейрона с нервно-мышечным синапсом (схема): а - строение нейрона: 1 - тело нейрона, 2 -
- 120. В основе работы нервной системы лежит рефлекс Рефлекс – ответ организма на раздражение, который осуществляется и
- 121. Деятельность коры головного мозга осуществляется по типу условных рефлексов, И.П. Павловым были сформулированы основные принципы, на
- 122. Основные законы проведения возбуждения по периферическим нервам 1. Возбуждение по нерву распространяется только при условии его
- 123. Особенности центральных отделов нервной системы 1.Возбуждение в пределах рефлекторной дуги распространяется в одном направлении. 2.Возбуждение в
- 124. Из мозга к рабочим органам (мышцам, железам и другим) нервные импульсы также следуют по цепям нейронов.
- 125. Виды рефлексов
- 127. Элементарная рефлекторная дуга безусловного рефлекса состоит из пяти основных звеньев. Она начинается рецепторами, которые трансформируют энергию
- 128. Простая рефлекторная дуга состоит из трех нейронов. Тело первого нейрона (чувствительного, приносящего) располагается в спинномозговом узле
- 129. Рефлекторная дуга - Путь, по которому проводятся нервные импульсы при осуществлении рефлекса Чувствительный нейрон Исполнительный нейрон
- 130. Схема. Распространение (направление показано стрелками) нервных импульсов по простой рефлекторной дуге. 1 - чувствительный (афферентный) нейрон;
- 132. Рефлекторная дуга состоит из пяти отделов: 1. Рецепторов, воспринимающих раздражение и отвечающих на него возбуждением. Рецепторами
- 134. Безусловные рефлексы— наследственно передаваемые (врожденные)реакции организма, присущие всему виду. Выполняют защитную функцию, а также функцию поддержания
- 135. Условные рефлексы— это приобретенные в течение индивидуальной жизни организма реакции, возникающие в определенных условиях на основе
- 136. Рефлекторное кольцо Представление о рефлексе как о целесообразной реакции организма заставляет дополнить рефлекторную дугу еще одним
- 137. Рецепторы В зависимости от локализации, если рассматривать организм снаружи внутрь, различают следующие виды рецепторов: 1) экстероцепторы
- 138. В зависимости от характера раздражения различают терморецепторы, механорецепторы, ноцирецепторы и др. Первые воспринимают изменение температуры, вторые
- 139. Другие нервные волокна, имеющие свободные окончания, проникают таким же образом в эпидермис и подходят к осязательным
- 140. Пластинчатые тельца (Фаттера - Паччини) - самые крупные из всех инкапсулированных нервных окончаний. Они имеют овальную
- 141. Мелкие (длиной 50-160 мкм, шириной около 60 мкм) овальные или цилиндрические осязательные тельца (Мейсснера) особенно многочисленны
- 142. Тельца Руффини веретенообразной формы расположены в коже пальцев кисти и стопы, в капсулах суставов и стенках
- 143. Барорецепторы представляют собой ветвящиеся свободные нервные окончания, залегающие в адвентициальном слое крупных артерий грудной полости и
- 144. Проприорецепторы воспринимают чувства сокращения мышц, натяжения сухожилий и суставных капсул, мышечной силы, необходимой для выполнения определенного
- 145. Хеморецепторы отвечают появлением нервного импульса в ответ на взаимодействие между рецепторным белком и определенной химической молекулой.
- 146. Каждым своим концом мышечное веретено прикрепляется к соединительнотканной оболочке (перимизию) пучка экстрафузальных волокон при помощи напоминающих
- 147. Наряду с сенсорной у интрафузальных мышечных волокон (как и у экстрафузальных) есть двигательная иннервация. Эти двигательные
- 148. Многообразие рецепторов, которыми располагает человек, обеспечивает ему объективную информацию об окружающем мире, о процессах, в нем
- 149. Нервный центр комплекс нейронов, сосредоточенных в одной структуре ЦНС (например, дыхательный центр продолговатого мозга), которые выполняют
- 150. Нейронные цепи соответствующим образом (последовательно) соединенные между собой нейроны, которые выполняют определенную задачу. Рефлекторная дуга является
- 151. Нейронные сети Объединение нейронов, которое содержит множество параллельно расположенных и связанных между собой последовательных цепей нейронов.
- 152. Типы нейронных сетей Иерархические сети характеризуются свойствами конвергенции (несколько нейронов одного уровня контактируют с меньшим числом
- 153. Типы нейронных сетей Локальные сети характеризуются тем, что в них поток информации удерживается в пределах одного
- 154. Типы нейронных сетей Дивергентные сети характеризуются наличием нейронов, которые, имея один вход, на выходе образуют контакты
- 155. В последние годы все более популярным в физиологии является представление о нейронных ансамблях, которые предложено рассматривать
- 156. Возрастные особенности структуры и функции нервных клеток Нервные клетки образуются из эктодермальных клеток первичной мозговой трубки
- 157. Растущий аксон на конце имеет колбу роста, в которой содержатся крупные пузырьки разной формы, но отсутствуют
- 158. Считают, что способность проводить возбуждение у дендрита появляется значительно позже, чем у аксона (аксон функционирует во
- 159. На основании величины ПКП (потенциала концевой пластинки) (около 4 мв) считают, что у плода в кванте
- 160. Механизм закручивания спирали точно неясен: большинство исследователей считает, что шванновская клетка вращается вокруг аксона и обволакивает
- 161. Схема, иллюстрирующая образование миелиновой оболочки: б-аксон (А) окружен отдельной шванновской клеткой, М-мезаксон, в - спиральное закручивание
- 162. Установлена зависимость возбудимости нервных волокон от миелинизации - по мере развития миелиновой оболочки возбудимость нервного волокна
- 163. Большинство смешанных и центростремительных нервов миелинизируются к 3-месячному возрасту, некоторые - к 3 годам. Миелинизация спинномозговых
- 164. Показана динамика миелинизации лицевого нерва. Его ветви, иннервирующие область губ миелинизируются в период от 21 до
- 165. Возрастные особенности нервной системы имеют некоторые отличия. Они различаются в детском, юном и пожилом возрасте. За
- 166. На этом возрастные особенности нервной системы детей не ограничиваются. Здесь перечислена лишь микроскопическая доля изменений в
- 167. К пяти годам некоторые дети умеют писать и читать. Малыш часами может проводить за красками и
- 168. Особенности нервной системы в период старения определяются общими изменениями организма в целом: изменение мышечного тонуса; умственная
- 169. При осмотре нейронов старого человека через микроскоп видны значительные изменения в виде увеличения нервных клеток и
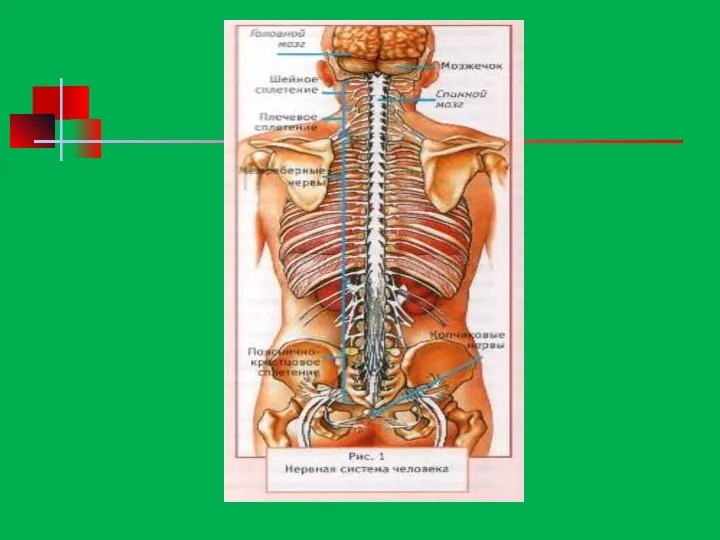
- 170. Центральная нервная система
- 171. Строение спинного мозга
- 172. Спинной мозг
- 174. Функции спинного мозга Рефлекторная – здесь находятся центры безусловных рефлексов Проводниковая функция – белое вещество спинного
- 175. Головной мозг
- 176. Строение головного мозга
- 177. Продолговатый мозг Мост Мозжечок Средний мозг Промежуточный мозг Большие полушария Мозолистое тело
- 178. Белое вещество Серое вещество Белое вещество составляет проводящие пути, связывающие головной мозг со спинным, а также
- 179. Продолговатый мозг и мост Регуляция: Дыхания Пищеварения (слюноотделение, жевание, глотание) Сердечно-сосудистой системы Защитные рефлексы: Чихание, моргание,
- 180. Мозжечок Координация произвольных движений Сохранение положения тела в пространстве Регуляция мышечного тонуса и равновесия
- 181. Средний мозг Ориентировочные рефлексы на зрительные и слуховые раздражители (поворот головы и тела в сторону световых
- 182. Промежуточный мозг Поддержание обмена веществ и энергии на оптимальном уровне Сбор и оценка поступающей информации от
- 183. Большие полушария Лобная доля Центральная борозда Боковая борозда Теменная доля Височная доля Затылочная доля
- 184. Затылочные доли – зрительная чувствительность Височные доли – слуховая, вкусовая, обонятельная чувствительность Лобные доли – произвольные
- 185. С большими полушариями мозга связаны: Память Речь Мышление Творческие процессы Личностные качества
- 189. Скачать презентацию

1.Морфологическая и функциональная характеристика нервной системы человека.
2. Нервная ткань.
3.Строение, типы,
1.Морфологическая и функциональная характеристика нервной системы человека.
2. Нервная ткань.
3.Строение, типы,

Функции нервной системы
▪ интегративно-координационная функция: обеспечивает функции различных органов и физиологических
Функции нервной системы
▪ интегративно-координационная функция: обеспечивает функции различных органов и физиологических

Основным принципом функционирования ЦНС является процесс регуляции, управления физиологическими функциями, которые
Основным принципом функционирования ЦНС является процесс регуляции, управления физиологическими функциями, которые

Морфологическая и функциональная характеристика нервной системы человека
Нервная система объединяет организм человека
Морфологическая и функциональная характеристика нервной системы человека
Нервная система объединяет организм человека

В мозге человека происходят сложные процессы обработки поступившей в него информации.
В мозге человека происходят сложные процессы обработки поступившей в него информации.

Нервная система обеспечивает эффективное приспособление организма к изменениям окружающей среды, формирует
Нервная система обеспечивает эффективное приспособление организма к изменениям окружающей среды, формирует

Число нервных элементов, будучи очень ограниченным у примитивных организмов, в процессе
Число нервных элементов, будучи очень ограниченным у примитивных организмов, в процессе

Различают два основных вида регуляции: гуморальный и нервный
Нервный процесс регуляции предусматривает
Различают два основных вида регуляции: гуморальный и нервный
Нервный процесс регуляции предусматривает
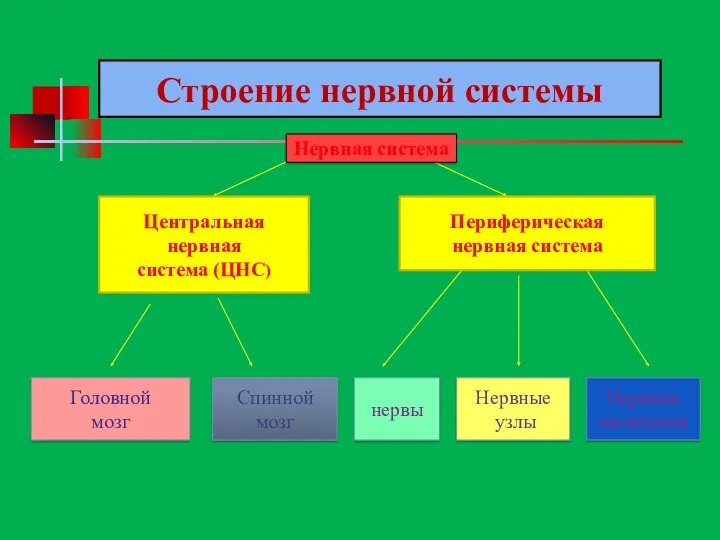
Центральная
нервная
система (ЦНС)
Периферическая
нервная система
Головной
мозг
Спинной
мозг
нервы
Нервные
узлы
Нервные
окончания
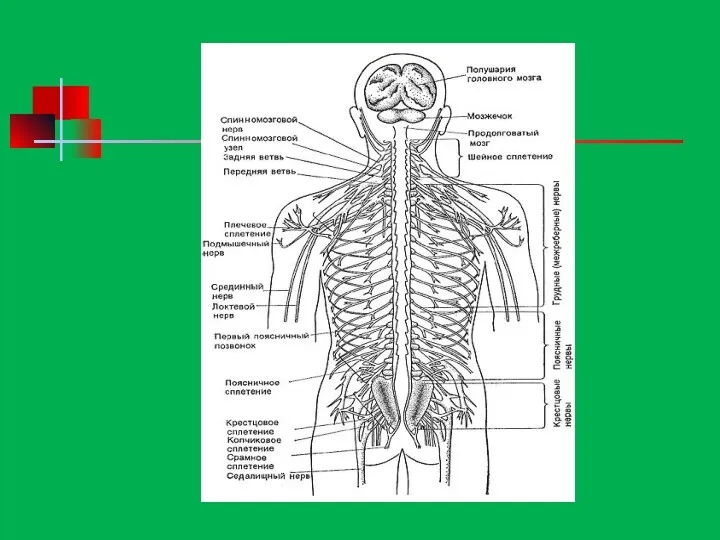
Строение нервной системы
Нервная система
Центральная
нервная
система (ЦНС)
Периферическая
нервная система
Головной
мозг
Спинной
мозг
нервы
Нервные
узлы
Нервные
окончания
Строение нервной системы
Нервная система
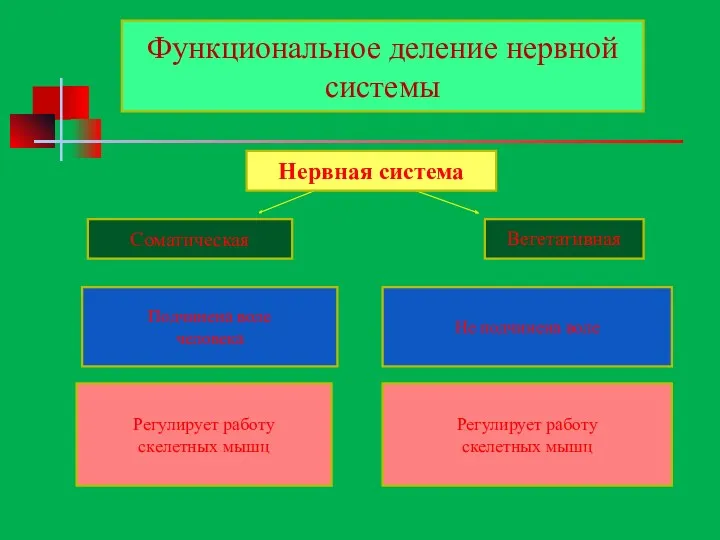
рвнанея система
Регулирует работу
скелетных мышц
Регулирует работу
внутренних
органов
Функциональное деление нервной системы
Нервная система
Соматическая
Вегетативная
Подчинена воле
человека
Не
рвнанея система
Регулирует работу
скелетных мышц
Регулирует работу
внутренних
органов
Функциональное деление нервной системы
Нервная система
Соматическая
Вегетативная
Подчинена воле
человека
Не
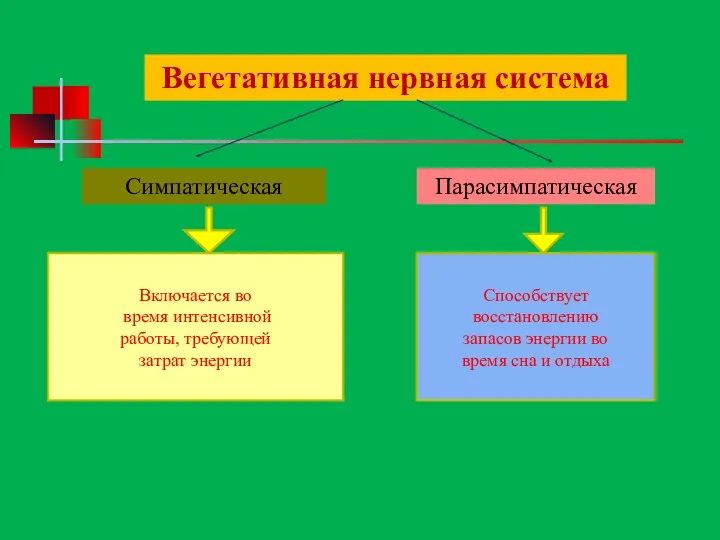
Вегетативная нервная система
Симпатическая
Парасимпатическая
Способствует
восстановлению
запасов энергии во
время сна и отдыха
Включается во
Вегетативная нервная система
Симпатическая
Парасимпатическая
Способствует
восстановлению
запасов энергии во
время сна и отдыха
Включается во

ИСТОЧНИК РАЗВИТИЯ – эктодерма (нейроэктодерма):
нервная пластинка – нервные валики –
нервная пластинка – нервные валики –

НЕРВНАЯ ТРУБКА
Вентрикулярная (эпендимная) зона
▪ делящиеся клетки цилиндрической формы.
Промежуточная (плащевая, мантийная)
НЕРВНАЯ ТРУБКА
Вентрикулярная (эпендимная) зона
▪ делящиеся клетки цилиндрической формы.
Промежуточная (плащевая, мантийная)

ОБЩИЕ ПРИНЦИПЫ СТРОЕНИЯ
1. Большинство нервных клеток являются длинноотросчатыми.
2. Нейроны характеризуются полярностью.
3.
ОБЩИЕ ПРИНЦИПЫ СТРОЕНИЯ
1. Большинство нервных клеток являются длинноотросчатыми.
2. Нейроны характеризуются полярностью.
3.

НЕРВНЫЕ ТКАНИ
(от греческого neuron – жила, струна, нейрон) – это
НЕРВНЫЕ ТКАНИ
(от греческого neuron – жила, струна, нейрон) – это

Нервная ткань
Основу нервной ткани составляют нервные клетки –
НЕЙРОНЫ
Нервная ткань
Основу нервной ткани составляют нервные клетки –
НЕЙРОНЫ
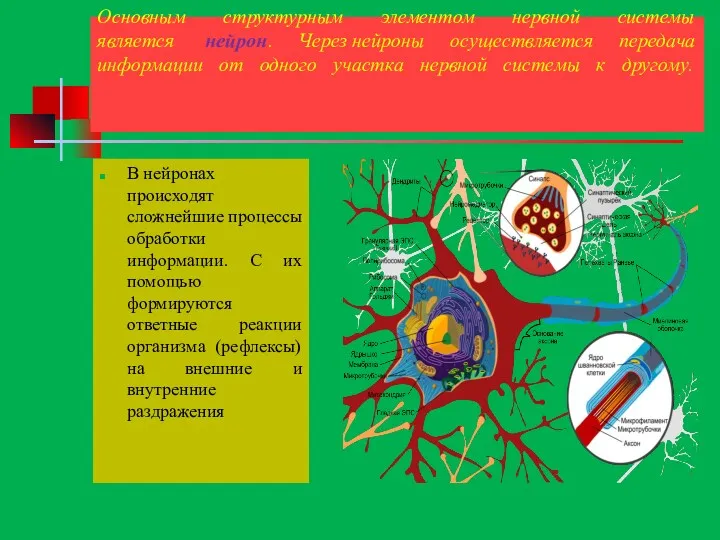
Основным структурным элементом нервной системы является нейрон. Через нейроны осуществляется передача информации
Основным структурным элементом нервной системы является нейрон. Через нейроны осуществляется передача информации

Функции нейронов
1) генерализация нервного импульса;
2) получение, хранение и передача информации;
3) способность суммировать возбуждающие и
Функции нейронов
1) генерализация нервного импульса;
2) получение, хранение и передача информации;
3) способность суммировать возбуждающие и

Структурно-функциональной единицей нервной системы является нейрон - специализированная клетка, способная принимать,
Структурно-функциональной единицей нервной системы является нейрон - специализированная клетка, способная принимать,

Гибель (апоптоз) нейронов в физиологических условиях у взрослого человека сравнительно невелика
Гибель (апоптоз) нейронов в физиологических условиях у взрослого человека сравнительно невелика

Нейрон развивается из небольшой клетки - предшественницы, которая перестает делиться еще
Нейрон развивается из небольшой клетки - предшественницы, которая перестает делиться еще

Среди нейронов встречаются самые крупные клеточные элементы организма. Размеры их поперечника
Среди нейронов встречаются самые крупные клеточные элементы организма. Размеры их поперечника

. В нейроне выделяют:
Воспринимающую часть.
Дендриты – основное воспринимающее поле нейрона. Мембрана дендрита
. В нейроне выделяют: Воспринимающую часть. Дендриты – основное воспринимающее поле нейрона. Мембрана дендрита
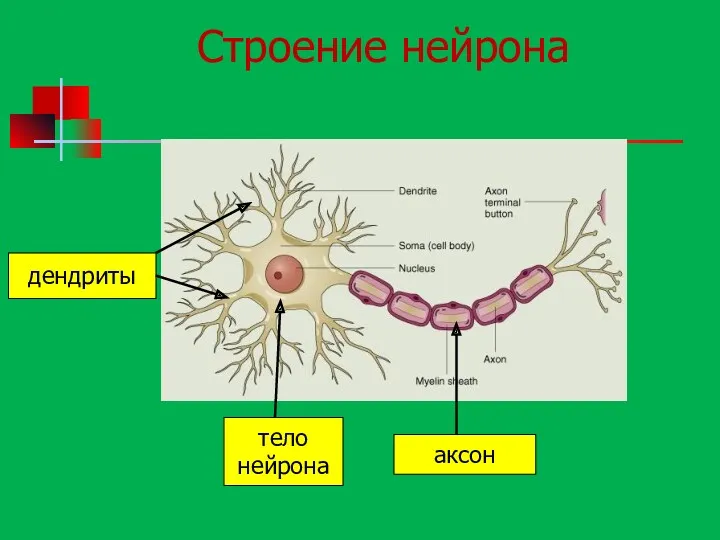
Строение нейрона
дендриты
тело
нейрона
аксон
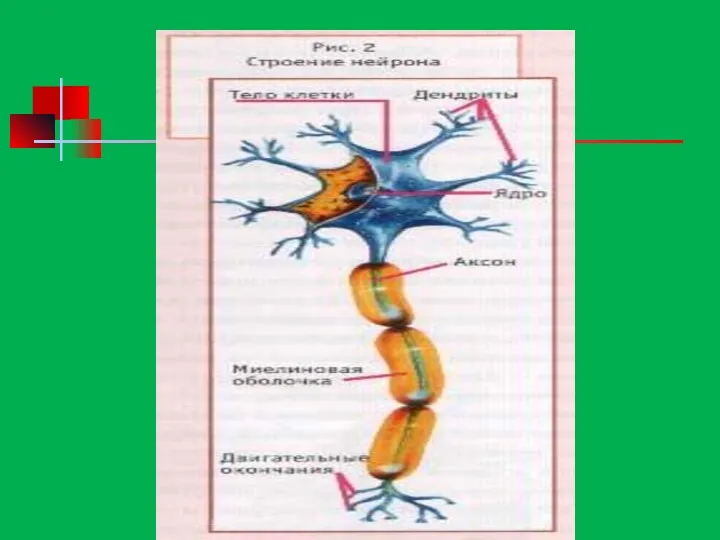
Строение нейрона
дендриты
тело
нейрона
аксон

МОРФОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА НЕЙРОНОВ
МОРФОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА НЕЙРОНОВ
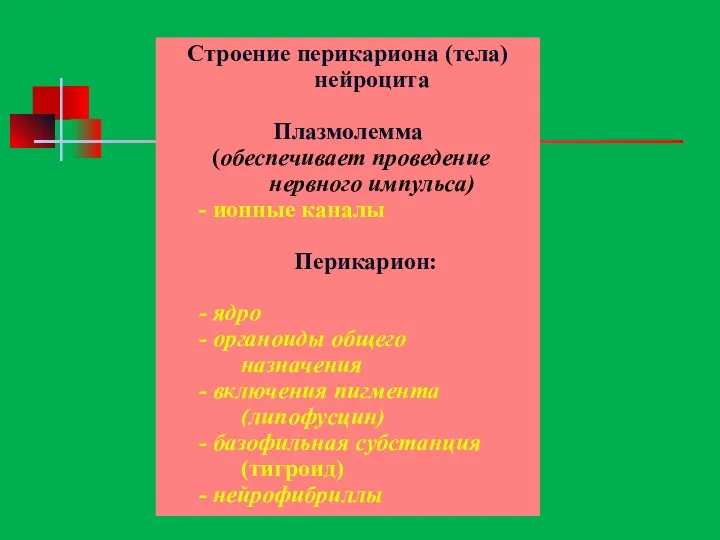
Строение перикариона (тела) нейроцита
Плазмолемма
(обеспечивает проведение нервного импульса)
- ионные каналы
Перикарион:
-
Строение перикариона (тела) нейроцита
Плазмолемма
(обеспечивает проведение нервного импульса)
- ионные каналы
Перикарион:
-

Перикарион, или сома нейрона, имеет клеточную мембрану и содержит ядро, рибосомы,
Перикарион, или сома нейрона, имеет клеточную мембрану и содержит ядро, рибосомы,

Ядро в большинстве нейронов расположено в центре тела клетки. Как правило,
Ядро в большинстве нейронов расположено в центре тела клетки. Как правило,

В нейронах хорошо развита гранулярная эндоплазматическая сеть, что говорит о высокой
В нейронах хорошо развита гранулярная эндоплазматическая сеть, что говорит о высокой

У взрослых людей наиболее крупных размеров тельца Ниссля достигают в мотонейронах.
У взрослых людей наиболее крупных размеров тельца Ниссля достигают в мотонейронах.

Комплекс, или аппарат, Гольджи хорошо развит в нейронах (его первое описание
Комплекс, или аппарат, Гольджи хорошо развит в нейронах (его первое описание

Лизосомальный аппарат, или аппарат внутриклеточного переваривания, нейронов обладает высокой активностью и
Лизосомальный аппарат, или аппарат внутриклеточного переваривания, нейронов обладает высокой активностью и

Пигменты (включения) нейронов представлены липофусцином и меланином. Липофусцин - это желто-коричневый
Пигменты (включения) нейронов представлены липофусцином и меланином. Липофусцин - это желто-коричневый

Белки нейронов служат для пластических и информационных целей. В ядре нейрона
Белки нейронов служат для пластических и информационных целей. В ядре нейрона
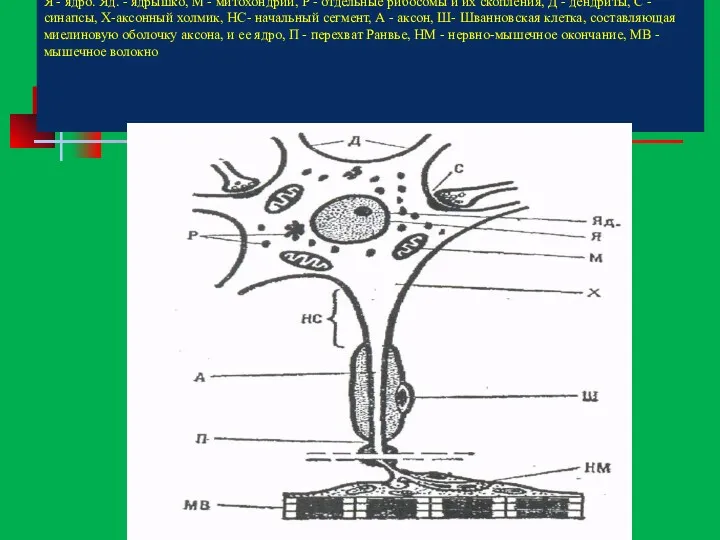
Строение нейрона:
Я - ядро. Яд. - ядрышко, М - митохондрии, Р
Строение нейрона: Я - ядро. Яд. - ядрышко, М - митохондрии, Р

Строение отростков
Аксон
у нейрона всегда один маловетвящийся (осевой)
толстый отросток, чаще длинный, отсутствует
тигроид,
Строение отростков
Аксон
у нейрона всегда один маловетвящийся (осевой)
толстый отросток, чаще длинный, отсутствует
тигроид,

Рассмотрим морфологические особенности дендритов. Отростки, по которым к телу нервной клетки
Рассмотрим морфологические особенности дендритов. Отростки, по которым к телу нервной клетки

Дендриты проводят импульсы к телу нейрона, получая сигналы от других нейронов
Дендриты проводят импульсы к телу нейрона, получая сигналы от других нейронов

Аксон (от лат. аксис - ось), или нейрит, - это центральный,
Аксон (от лат. аксис - ось), или нейрит, - это центральный,

Характеристика аксоплазмы аксона
В крупных нейронах аксон может содержать до 99 %
Характеристика аксоплазмы аксона
В крупных нейронах аксон может содержать до 99 %

Аксон может по своему ходу давать ответвления, или коллатерали (от лат.
Аксон может по своему ходу давать ответвления, или коллатерали (от лат.
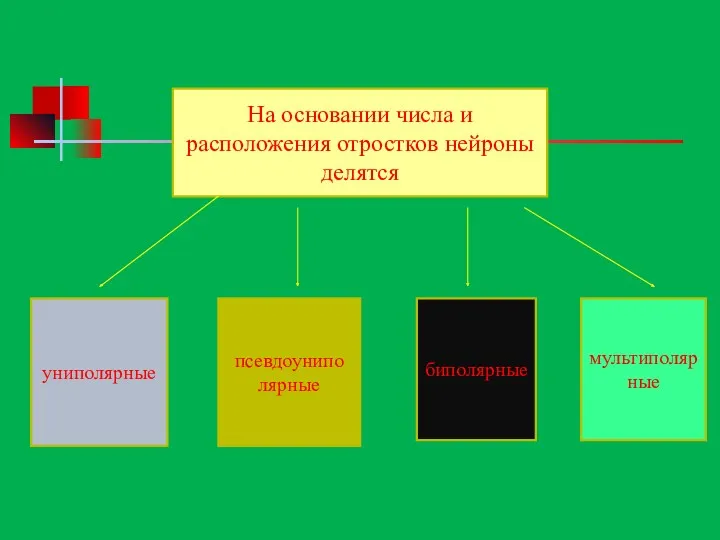
На основании числа и расположения отростков нейроны делятся
униполярные
псевдоунипо
лярные
биполярные
мультиполярные
На основании числа и расположения отростков нейроны делятся
униполярные
псевдоунипо
лярные
биполярные
мультиполярные

Существует большое многообразие нейронов ЦНС. Поэтому предложены и различные варианты их
Существует большое многообразие нейронов ЦНС. Поэтому предложены и различные варианты их

Биполярные нейроны (синонимы - двухотросчатые, или двухполюсные, нейроны) имеют два отростка
Биполярные нейроны (синонимы - двухотросчатые, или двухполюсные, нейроны) имеют два отростка

Мультиполярные нейроны имеют один аксон и много (2 и более) дендритов.
Мультиполярные нейроны имеют один аксон и много (2 и более) дендритов.
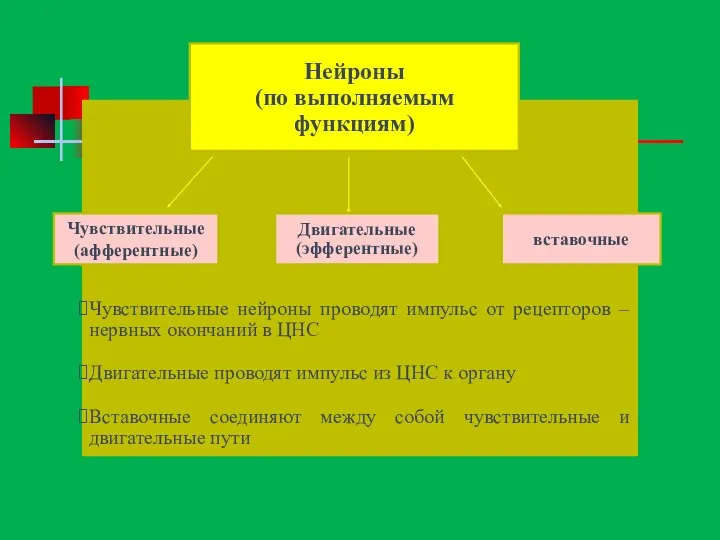
Чувствительные нейроны проводят импульс от рецепторов – нервных окончаний в ЦНС
Двигательные
Чувствительные нейроны проводят импульс от рецепторов – нервных окончаний в ЦНС
Двигательные

Эфферентные нейроны связаны с передачей нисходящих влияний от вышележащих этажей нервной
Эфферентные нейроны связаны с передачей нисходящих влияний от вышележащих этажей нервной

Функциональная классификация нейронов разделяет их по характеру выполняемой ими функции (в
Функциональная классификация нейронов разделяет их по характеру выполняемой ими функции (в

Моносенсорные нейроны подразделяют функционально по их чувствительности к разным качествам одного
Моносенсорные нейроны подразделяют функционально по их чувствительности к разным качествам одного

Эфферентные нейроны (синонимы - двигательные, моторные, секреторные, центробежные, сердечные, сосудодвигательные
Эфферентные нейроны (синонимы - двигательные, моторные, секреторные, центробежные, сердечные, сосудодвигательные

Вставочные нейроны (синонимы - интернейроны, контактные, ассоциативные, коммуникативные, объединяющие, замыкательные, проводниковые,
Вставочные нейроны (синонимы - интернейроны, контактные, ассоциативные, коммуникативные, объединяющие, замыкательные, проводниковые,

Биохимическая классификация нейронов основана на химических особенностях нейромедиаторов, используемых нейронами в
Биохимическая классификация нейронов основана на химических особенностях нейромедиаторов, используемых нейронами в

Другие виды классификаций нейронов. Нервные клетки разных отделов нервной системы могут
Другие виды классификаций нейронов. Нервные клетки разных отделов нервной системы могут
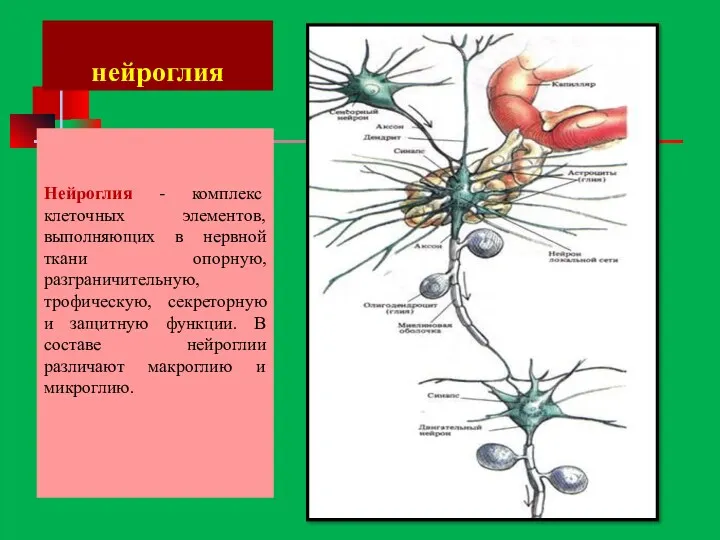
нейроглия
Нейроглия - комплекс клеточных элементов, выполняющих в нервной ткани опорную, разграничительную,
нейроглия
Нейроглия - комплекс клеточных элементов, выполняющих в нервной ткани опорную, разграничительную,

НЕЙРОГЛИЯ (греческое neuron – нерв, glia – клей) – термин, введенный
НЕЙРОГЛИЯ (греческое neuron – нерв, glia – клей) – термин, введенный
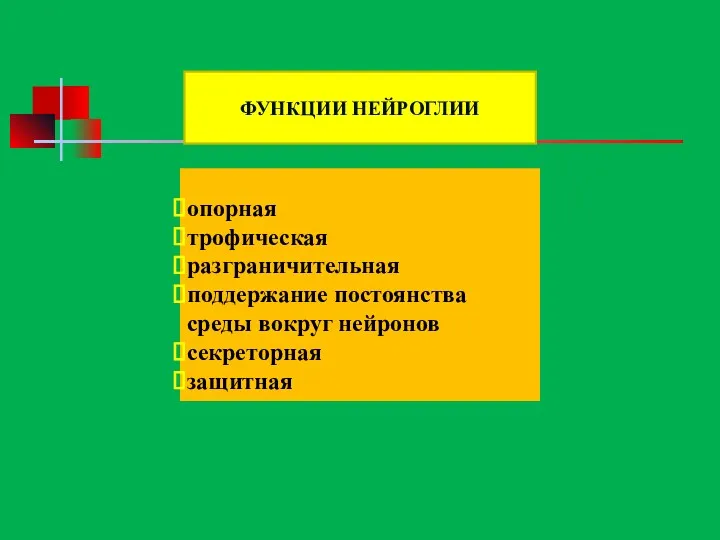
опорная
трофическая
разграничительная
поддержание постоянства среды вокруг нейронов
секреторная
защитная
ФУНКЦИИ НЕЙРОГЛИИ
трофическая
разграничительная
поддержание постоянства среды вокруг нейронов
секреторная
защитная
ФУНКЦИИ НЕЙРОГЛИИ

Классификация нейроглии
Микроглиальные клеткиМикроглиальные клетки, хоть и входят в понятие «глия», не
Классификация нейроглии
Микроглиальные клеткиМикроглиальные клетки, хоть и входят в понятие «глия», не
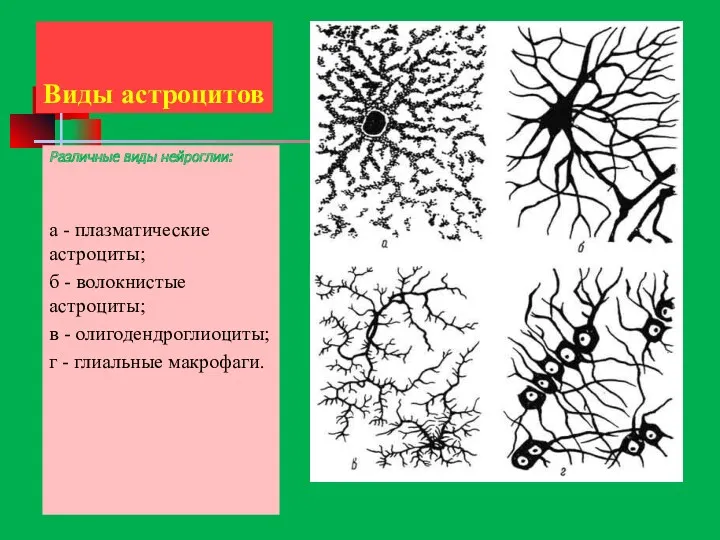
Виды астроцитов
Различные виды нейроглии:
а - плазматические астроциты;
б - волокнистые астроциты;
в
Виды астроцитов
Различные виды нейроглии:
а - плазматические астроциты;
б - волокнистые астроциты;
в

Морфология нейроглии
Нейроглия - это обширная разнородная группа клеток (глиоцитов, или глиальных
Морфология нейроглии
Нейроглия - это обширная разнородная группа клеток (глиоцитов, или глиальных

Глиальные клетки по размерам в 3-4 раза меньше, чем нейроны. В
Глиальные клетки по размерам в 3-4 раза меньше, чем нейроны. В

На долю астроцитов (или звездчатых глиальных клеток) приходится около 40% от
На долю астроцитов (или звездчатых глиальных клеток) приходится около 40% от

Астроциты выполняют четыре основные функции - опорную, разграничительную (транспортную и барьерную),
Астроциты выполняют четыре основные функции - опорную, разграничительную (транспортную и барьерную),

Разграничительная функция астроцитов, которая включает транспортную и барьерную функции, направлена на
Разграничительная функция астроцитов, которая включает транспортную и барьерную функции, направлена на

Метаболическая функция астроцитов, включающая в себя и регуляторную функцию - это
Метаболическая функция астроцитов, включающая в себя и регуляторную функцию - это

Защитная (иммунная и репаративная) функция астроцитов заключается в формировании различных защитных
Защитная (иммунная и репаративная) функция астроцитов заключается в формировании различных защитных

Олигодендроциты - это обширная группа разнообразных мелких клеток с короткими немногочисленными
Олигодендроциты - это обширная группа разнообразных мелких клеток с короткими немногочисленными

Олигодендроциты выполяют две основные функции - 1) образование миелина как компонента
Олигодендроциты выполяют две основные функции - 1) образование миелина как компонента

Эпендимоциты - еще одна разновидность глиоцитов. Они образуют эпендимную глию, или
Эпендимоциты - еще одна разновидность глиоцитов. Они образуют эпендимную глию, или

Микроглия представляет собой совокупность мелких удлиненных звездчатых клеток (микроглиоцитов) с плотной
Микроглия представляет собой совокупность мелких удлиненных звездчатых клеток (микроглиоцитов) с плотной

Основная функция микроглии - защитная (в том числе иммунная). Клетки микроглии
Основная функция микроглии - защитная (в том числе иммунная). Клетки микроглии

Клетки-сателлиты (мантийные клетки) охватывают тела нейронов в спинальных, черепно-мозговых и вегетативных
Клетки-сателлиты (мантийные клетки) охватывают тела нейронов в спинальных, черепно-мозговых и вегетативных

Нейропиль - это понятие морфофункциональное. Оно отражает пространство мозговой ткани, расположенное
Нейропиль - это понятие морфофункциональное. Оно отражает пространство мозговой ткани, расположенное

Нервы – скопления отростков нейронов вне ЦНС, заключённые в общую оболочку

Нервом называется анатомическое образование, состоящее из нервных волокон, имеющих общую оболочку.
Нервом называется анатомическое образование, состоящее из нервных волокон, имеющих общую оболочку.

Нервы бывают различной длины и толщины, более длинные нервы расположены в
Нервы бывают различной длины и толщины, более длинные нервы расположены в
нервы
двигатель
ные
чувствительные
Вегетатив
ные
смешанные
нервы
двигатель
ные
чувствительные
Вегетатив
ные
смешанные

Двигательные нервы состоят из волокон клеток передних рогов спинного мозга или
Двигательные нервы состоят из волокон клеток передних рогов спинного мозга или

Аксон, заключенный в глиальную оболочку, называется нервным волокном
Аксон, заключенный в глиальную оболочку, называется нервным волокном

Совокупность нервных волокон образует нервные пучки, совокупность которых, в свою очередь,
Совокупность нервных волокон образует нервные пучки, совокупность которых, в свою очередь,

Все аксоны покрыты глиальной оболочкой, однако эта оболочка устроена по разному
Все аксоны покрыты глиальной оболочкой, однако эта оболочка устроена по разному

Безмиелиновые нервные волокна у взрослого располагаются преимущественно в составе вегетативной нервной
Безмиелиновые нервные волокна у взрослого располагаются преимущественно в составе вегетативной нервной

Миелиновые нервные волокна - это большая часть всех нервных волокон. Они
Миелиновые нервные волокна - это большая часть всех нервных волокон. Они

Миелиновая оболочка на 80 % состоит из липидов, обладающих высоким омическим
Миелиновая оболочка на 80 % состоит из липидов, обладающих высоким омическим

По длине волокна миелиновая оболочка имеет прерывистый ход, благодаря чему формируются
По длине волокна миелиновая оболочка имеет прерывистый ход, благодаря чему формируются

Перехваты Ранвье играют важную роль в процессе проведения возбуждения по нервному
Перехваты Ранвье играют важную роль в процессе проведения возбуждения по нервному

Критерием структурно-функциональной зрелости мякотных и безмякотных нервных волокон является увеличение их
Критерием структурно-функциональной зрелости мякотных и безмякотных нервных волокон является увеличение их

В безмякотных волокнах распределение ионных каналов остается равномерным по всей длине
В безмякотных волокнах распределение ионных каналов остается равномерным по всей длине

Мембрана аксона разделяет жидкие среды, различающиеся по содержанию ионов натрия и
Мембрана аксона разделяет жидкие среды, различающиеся по содержанию ионов натрия и

Распространению нервного импульса вдоль аксона способствует локализованное поступление внутрь аксона ионов
Распространению нервного импульса вдоль аксона способствует локализованное поступление внутрь аксона ионов

Возбудимость нервных волокон плода и новорожденного значительно ниже, чем у взрослого,
Возбудимость нервных волокон плода и новорожденного значительно ниже, чем у взрослого,

Фазовые изменения возбудимости во время возбуждения в раннем постнатальном онтогенезе также
Фазовые изменения возбудимости во время возбуждения в раннем постнатальном онтогенезе также

Скорость распространения возбуждения по нервным волокнам у детей становится такой
Скорость распространения возбуждения по нервным волокнам у детей становится такой
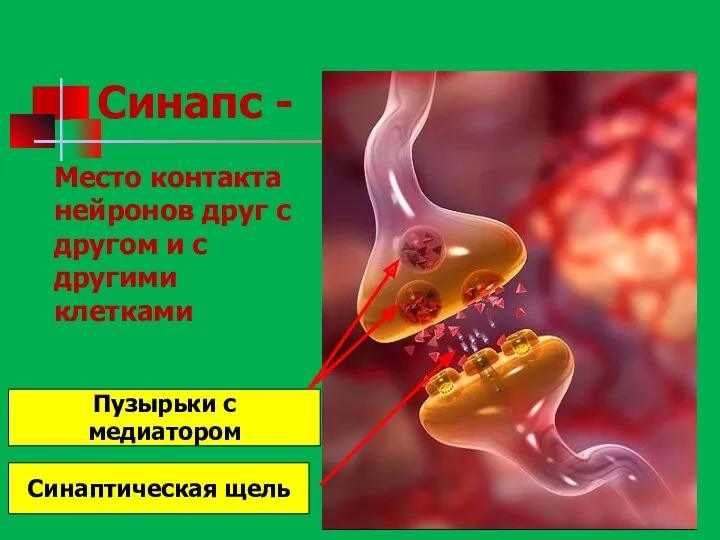
Синапс -
Место контакта нейронов друг с другом и с другими
Синапс -
Место контакта нейронов друг с другом и с другими
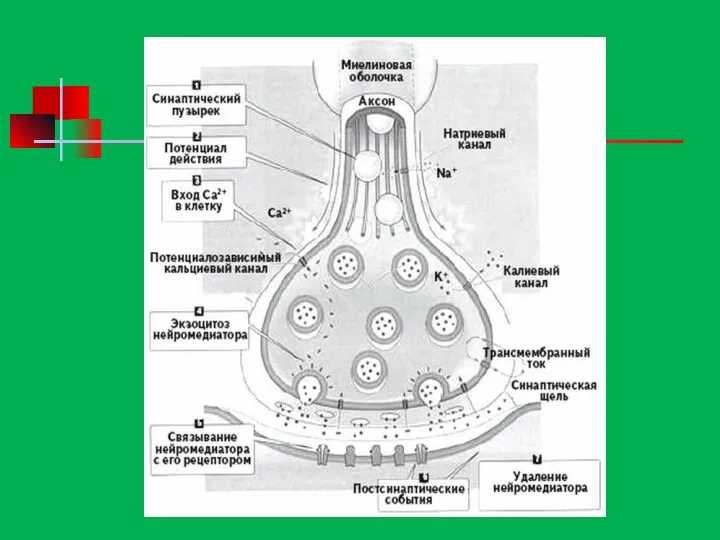
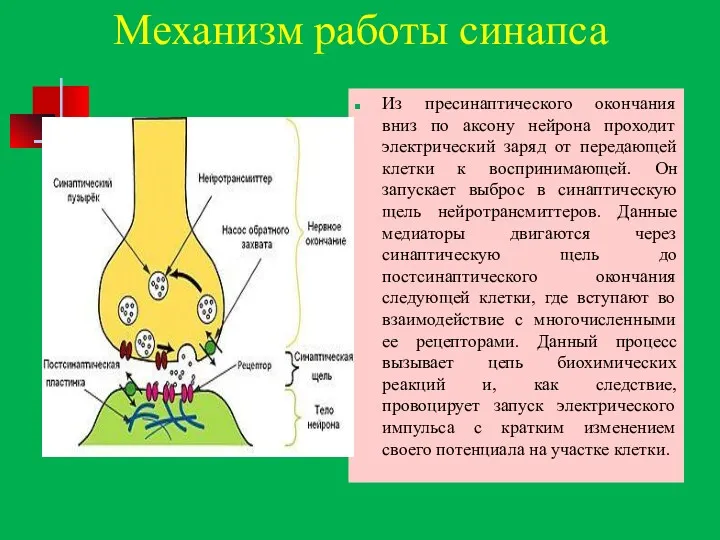
Механизм работы синапса
Из пресинаптического окончания вниз по аксону нейрона проходит электрический
Механизм работы синапса
Из пресинаптического окончания вниз по аксону нейрона проходит электрический

Термин и понятие «синапс», «синаптическая передача» был введен в физиологию Ч.
Термин и понятие «синапс», «синаптическая передача» был введен в физиологию Ч.

1. По локализации - центральные (головной и спинной мозг) и периферические

а) по форме контакта - терминальные (колбообразные соединения) и проходящие (варикозные

Общие представления о строении и механизмах функционирования химического синапса
Химические синапсы
Общие представления о строении и механизмах функционирования химического синапса
Химические синапсы

Пресинаптическая часть (или пресинаптический элемент, пресинпас) образуется аксоном по его ходу
Пресинаптическая часть (или пресинаптический элемент, пресинпас) образуется аксоном по его ходу

Постсинаптическая часть (или постсинаптический элемент) представлена постсинаптической мембраной, содержащей особые комплексы
Постсинаптическая часть (или постсинаптический элемент) представлена постсинаптической мембраной, содержащей особые комплексы

В литературе существует большое разнообразие в названиях синапсов. Например, синаптическая бляшка
В литературе существует большое разнообразие в названиях синапсов. Например, синаптическая бляшка

Кроме того, в ряде синапсов имеются метаботропные рецепторы, при активации которых
Кроме того, в ряде синапсов имеются метаботропные рецепторы, при активации которых

Одновременно, выделившийся в синаптическую щель медиатор может взаимодействовать с рецепторами, расположенными
Одновременно, выделившийся в синаптическую щель медиатор может взаимодействовать с рецепторами, расположенными

Рассмотрим, каким образом происходит активация постсинаптических рецепторов, а также вторичные посредники,
Рассмотрим, каким образом происходит активация постсинаптических рецепторов, а также вторичные посредники,

При активации ионотропных рецепторов, содержащих внутри себя ионные каналы, происходит изменение
При активации ионотропных рецепторов, содержащих внутри себя ионные каналы, происходит изменение

Для различных синапсов существует своя последовательность передачи сигнала (медиатор, G-белок, вторичный
Для различных синапсов существует своя последовательность передачи сигнала (медиатор, G-белок, вторичный
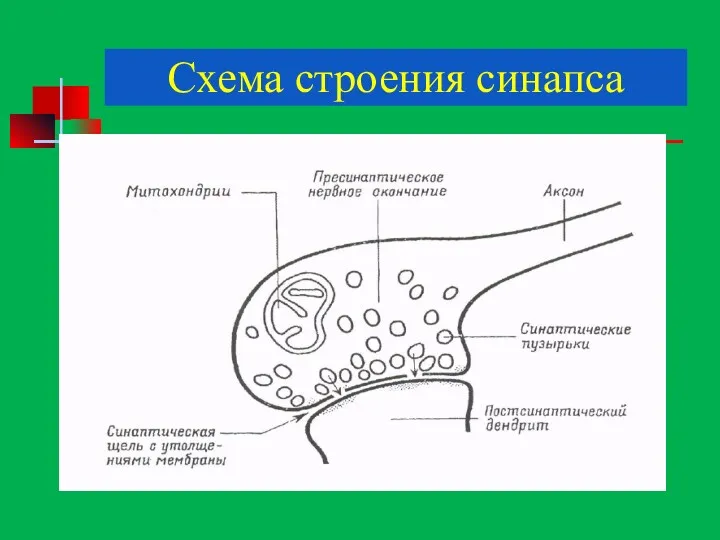
Схема строения синапса
Схема строения синапса

Локализация медиаторов и соответствующих нейронов ЦНС
В разных отделах ЦНС расположены
Локализация медиаторов и соответствующих нейронов ЦНС
В разных отделах ЦНС расположены

Гистамин является медиатором нейронов сосцевидых телец гипоталамуса. Аксоны гистаминергических нейронов проецируются
Гистамин является медиатором нейронов сосцевидых телец гипоталамуса. Аксоны гистаминергических нейронов проецируются

Соматостат - гипоталамус и другие отделы мозга, желатинозная субстанция, сетчатка. Гонадолиберин
Соматостат - гипоталамус и другие отделы мозга, желатинозная субстанция, сетчатка. Гонадолиберин
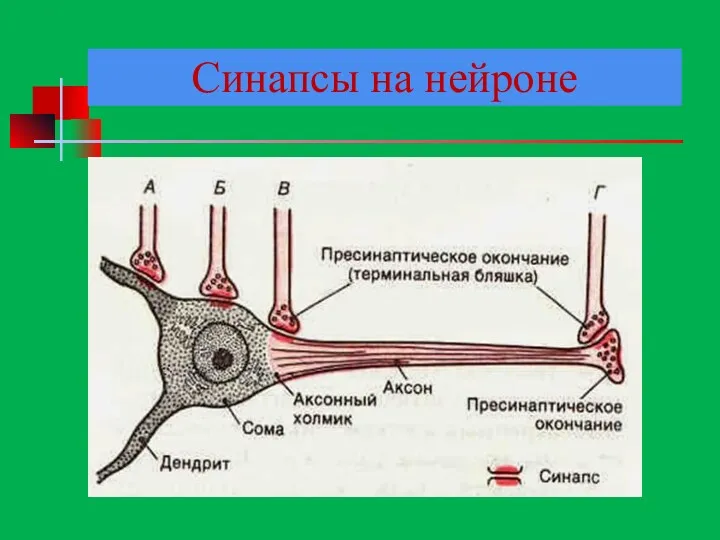
Синапсы на нейроне
Синапсы на нейроне

Нервно-мышечный синапс
Структурно-функциональное созревание нервно-мышечных синапсов охватывает почти весь период антенатального и
Нервно-мышечный синапс
Структурно-функциональное созревание нервно-мышечных синапсов охватывает почти весь период антенатального и

В процессе развития усиливается синтез ацетилхолина в мотонейронах, возрастает количество активных
В процессе развития усиливается синтез ацетилхолина в мотонейронах, возрастает количество активных

На постсинаптической мембране образуются складки, происходит концентрация холинорецепторов в синаптической зоне,
На постсинаптической мембране образуются складки, происходит концентрация холинорецепторов в синаптической зоне,
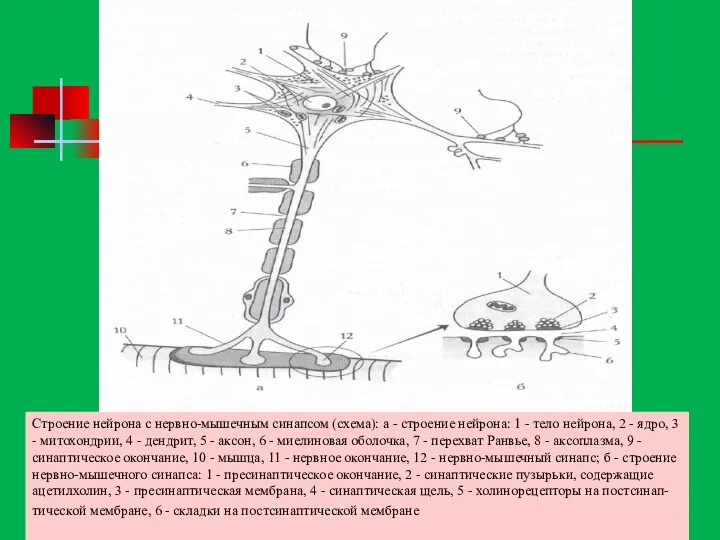
Строение нейрона с нервно-мышечным синапсом (схема): а - строение нейрона: 1
Строение нейрона с нервно-мышечным синапсом (схема): а - строение нейрона: 1

В основе работы нервной системы лежит рефлекс
Рефлекс – ответ организма на
В основе работы нервной системы лежит рефлекс
Рефлекс – ответ организма на

Деятельность коры головного мозга осуществляется по типу условных рефлексов, И.П. Павловым
Деятельность коры головного мозга осуществляется по типу условных рефлексов, И.П. Павловым

Основные законы проведения возбуждения по периферическим нервам
1. Возбуждение по нерву
Основные законы проведения возбуждения по периферическим нервам
1. Возбуждение по нерву

Особенности центральных отделов нервной системы
1.Возбуждение в пределах рефлекторной дуги распространяется в
Особенности центральных отделов нервной системы
1.Возбуждение в пределах рефлекторной дуги распространяется в

Из мозга к рабочим органам (мышцам, железам и другим) нервные импульсы
Из мозга к рабочим органам (мышцам, железам и другим) нервные импульсы

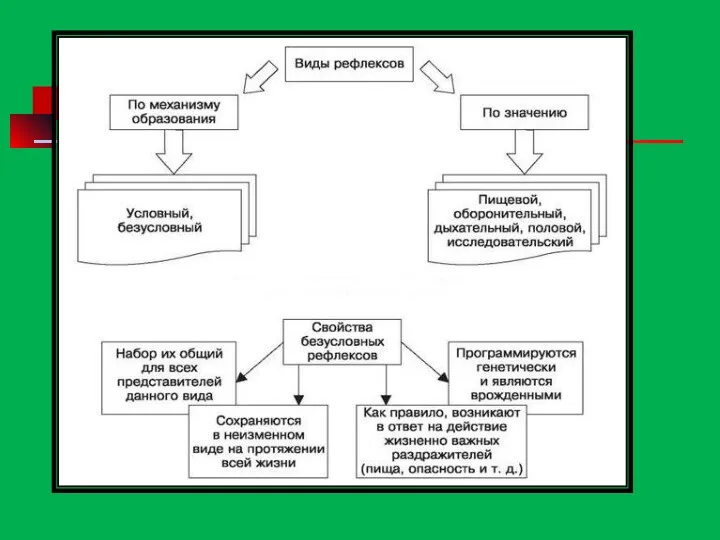
Виды рефлексов
Виды рефлексов

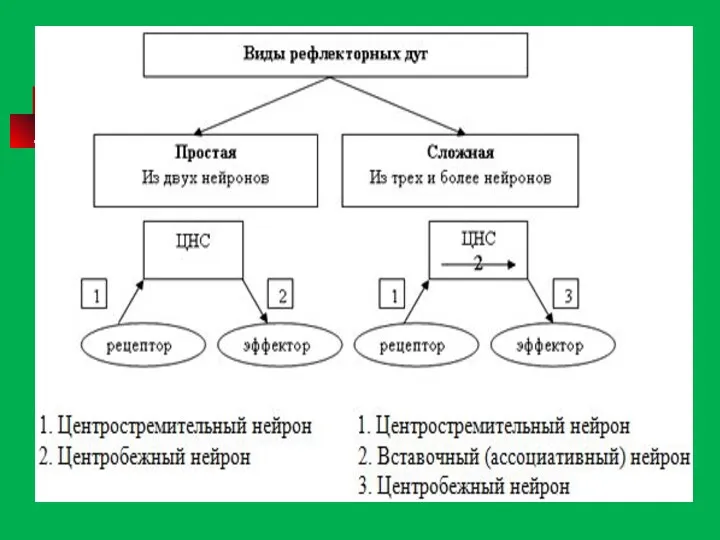
Элементарная рефлекторная дуга безусловного рефлекса состоит из пяти основных звеньев. Она
Элементарная рефлекторная дуга безусловного рефлекса состоит из пяти основных звеньев. Она

Простая рефлекторная дуга состоит из трех нейронов. Тело первого нейрона (чувствительного,
Простая рефлекторная дуга состоит из трех нейронов. Тело первого нейрона (чувствительного,
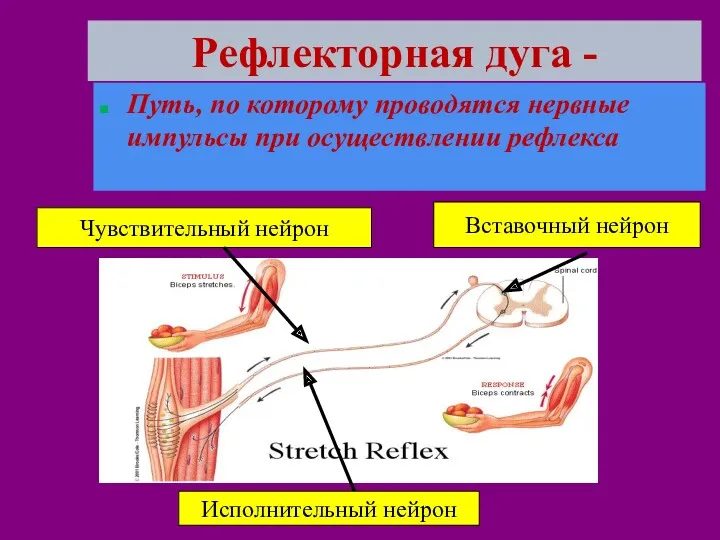
Рефлекторная дуга -
Путь, по которому проводятся нервные импульсы при осуществлении
Рефлекторная дуга -
Путь, по которому проводятся нервные импульсы при осуществлении
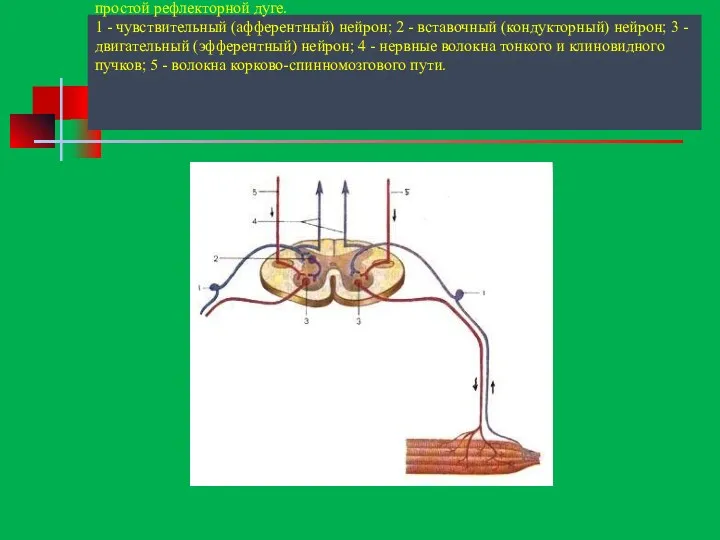
Схема. Распространение (направление показано стрелками) нервных импульсов по простой рефлекторной дуге.
1
Схема. Распространение (направление показано стрелками) нервных импульсов по простой рефлекторной дуге. 1
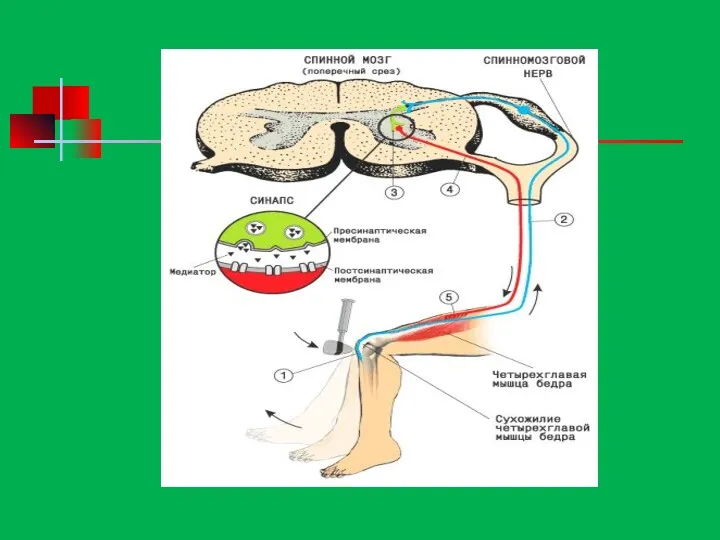

Рефлекторная дуга состоит из пяти отделов:
1. Рецепторов, воспринимающих раздражение и отвечающих
Рефлекторная дуга состоит из пяти отделов: 1. Рецепторов, воспринимающих раздражение и отвечающих
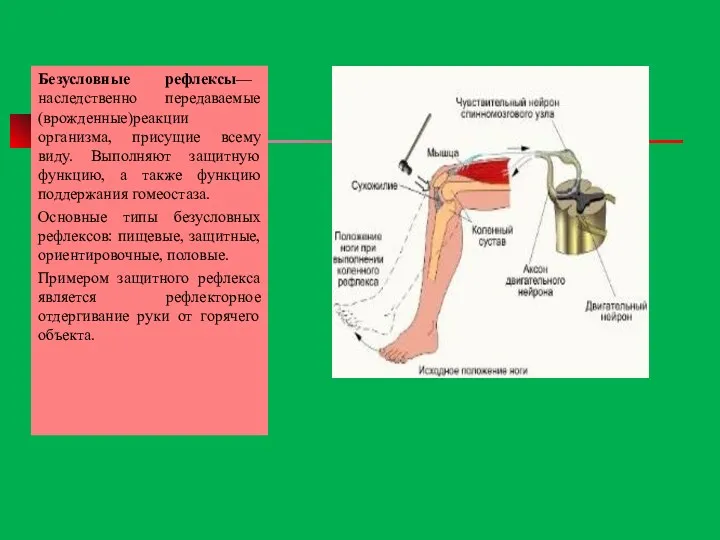
Безусловные рефлексы— наследственно передаваемые (врожденные)реакции организма, присущие всему виду. Выполняют защитную
Безусловные рефлексы— наследственно передаваемые (врожденные)реакции организма, присущие всему виду. Выполняют защитную

Условные рефлексы— это приобретенные в течение индивидуальной жизни организма реакции, возникающие
Условные рефлексы— это приобретенные в течение индивидуальной жизни организма реакции, возникающие

Рефлекторное кольцо
Представление о рефлексе как о целесообразной реакции организма заставляет
Рефлекторное кольцо
Представление о рефлексе как о целесообразной реакции организма заставляет

Рецепторы
В зависимости от локализации, если рассматривать организм снаружи внутрь, различают следующие
Рецепторы
В зависимости от локализации, если рассматривать организм снаружи внутрь, различают следующие

В зависимости от характера раздражения различают терморецепторы, механорецепторы, ноцирецепторы и др.
В зависимости от характера раздражения различают терморецепторы, механорецепторы, ноцирецепторы и др.

Другие нервные волокна, имеющие свободные окончания, проникают таким же образом в
Другие нервные волокна, имеющие свободные окончания, проникают таким же образом в

Пластинчатые тельца (Фаттера - Паччини) - самые крупные из всех
Пластинчатые тельца (Фаттера - Паччини) - самые крупные из всех

Мелкие (длиной 50-160 мкм, шириной около 60 мкм) овальные или цилиндрические
Мелкие (длиной 50-160 мкм, шириной около 60 мкм) овальные или цилиндрические

Тельца Руффини веретенообразной формы расположены в коже пальцев кисти и
Тельца Руффини веретенообразной формы расположены в коже пальцев кисти и

Барорецепторы представляют собой ветвящиеся свободные нервные окончания, залегающие в адвентициальном
Барорецепторы представляют собой ветвящиеся свободные нервные окончания, залегающие в адвентициальном

Проприорецепторы воспринимают чувства сокращения мышц, натяжения сухожилий и суставных капсул, мышечной
Проприорецепторы воспринимают чувства сокращения мышц, натяжения сухожилий и суставных капсул, мышечной

Хеморецепторы отвечают появлением нервного импульса в ответ на взаимодействие между
Хеморецепторы отвечают появлением нервного импульса в ответ на взаимодействие между

Каждым своим концом мышечное веретено прикрепляется к соединительнотканной оболочке (перимизию) пучка
Каждым своим концом мышечное веретено прикрепляется к соединительнотканной оболочке (перимизию) пучка

Наряду с сенсорной у интрафузальных мышечных волокон (как и у экстрафузальных)
Наряду с сенсорной у интрафузальных мышечных волокон (как и у экстрафузальных)

Многообразие рецепторов, которыми располагает человек, обеспечивает ему объективную информацию об окружающем
Многообразие рецепторов, которыми располагает человек, обеспечивает ему объективную информацию об окружающем

Нервный центр
комплекс нейронов, сосредоточенных в одной структуре ЦНС (например, дыхательный центр
Нервный центр
комплекс нейронов, сосредоточенных в одной структуре ЦНС (например, дыхательный центр

Нейронные цепи
соответствующим образом (последовательно) соединенные между собой нейроны, которые выполняют определенную
Нейронные цепи
соответствующим образом (последовательно) соединенные между собой нейроны, которые выполняют определенную

Нейронные сети
Объединение нейронов, которое содержит множество параллельно расположенных и связанных между
Нейронные сети
Объединение нейронов, которое содержит множество параллельно расположенных и связанных между

Типы нейронных сетей
Иерархические сети характеризуются свойствами конвергенции (несколько нейронов одного уровня
Типы нейронных сетей
Иерархические сети характеризуются свойствами конвергенции (несколько нейронов одного уровня

Типы нейронных сетей
Локальные сети характеризуются тем, что в них поток информации
Типы нейронных сетей
Локальные сети характеризуются тем, что в них поток информации

Типы нейронных сетей
Дивергентные сети характеризуются наличием нейронов, которые, имея один вход,
Типы нейронных сетей
Дивергентные сети характеризуются наличием нейронов, которые, имея один вход,

В последние годы все более популярным в физиологии является представление
В последние годы все более популярным в физиологии является представление

Возрастные особенности структуры и функции нервных клеток
Нервные клетки образуются из эктодермальных
Возрастные особенности структуры и функции нервных клеток
Нервные клетки образуются из эктодермальных

Растущий аксон на конце имеет колбу роста, в которой содержатся крупные
Растущий аксон на конце имеет колбу роста, в которой содержатся крупные

Считают, что способность проводить возбуждение у дендрита появляется значительно позже, чем
Считают, что способность проводить возбуждение у дендрита появляется значительно позже, чем

На основании величины ПКП (потенциала концевой пластинки) (около 4 мв) считают,
На основании величины ПКП (потенциала концевой пластинки) (около 4 мв) считают,

Механизм закручивания спирали точно неясен: большинство исследователей считает, что шванновская клетка
Механизм закручивания спирали точно неясен: большинство исследователей считает, что шванновская клетка
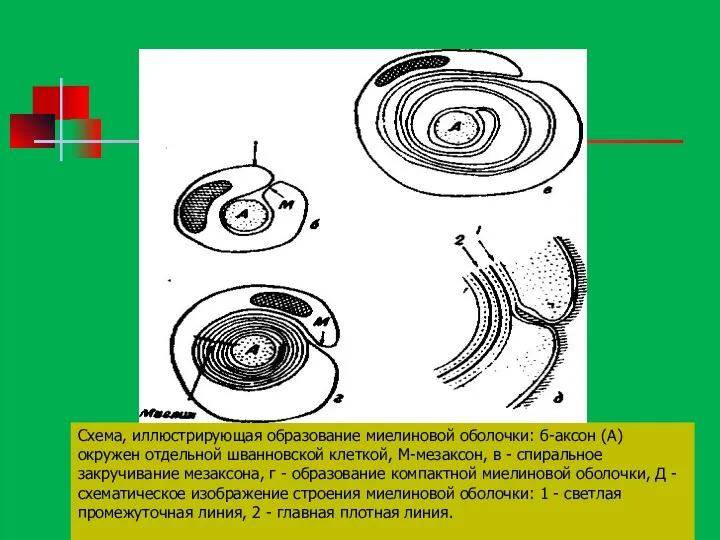
Схема, иллюстрирующая образование миелиновой оболочки: б-аксон (А) окружен отдельной шванновской клеткой,
Схема, иллюстрирующая образование миелиновой оболочки: б-аксон (А) окружен отдельной шванновской клеткой,

Установлена зависимость возбудимости нервных волокон от миелинизации - по мере развития
Установлена зависимость возбудимости нервных волокон от миелинизации - по мере развития

Большинство смешанных и центростремительных нервов миелинизируются к 3-месячному возрасту, некоторые -
Большинство смешанных и центростремительных нервов миелинизируются к 3-месячному возрасту, некоторые -

Показана динамика миелинизации лицевого нерва. Его ветви, иннервирующие область губ миелинизируются
Показана динамика миелинизации лицевого нерва. Его ветви, иннервирующие область губ миелинизируются

Возрастные особенности нервной системы имеют некоторые отличия. Они различаются в детском,
Возрастные особенности нервной системы имеют некоторые отличия. Они различаются в детском,

На этом возрастные особенности нервной системы детей не ограничиваются. Здесь перечислена
На этом возрастные особенности нервной системы детей не ограничиваются. Здесь перечислена

К пяти годам некоторые дети умеют писать и читать. Малыш часами
К пяти годам некоторые дети умеют писать и читать. Малыш часами

Особенности нервной системы в период старения определяются общими изменениями организма в
Особенности нервной системы в период старения определяются общими изменениями организма в

При осмотре нейронов старого человека через микроскоп видны значительные изменения в
При осмотре нейронов старого человека через микроскоп видны значительные изменения в

Центральная нервная система
Центральная нервная система
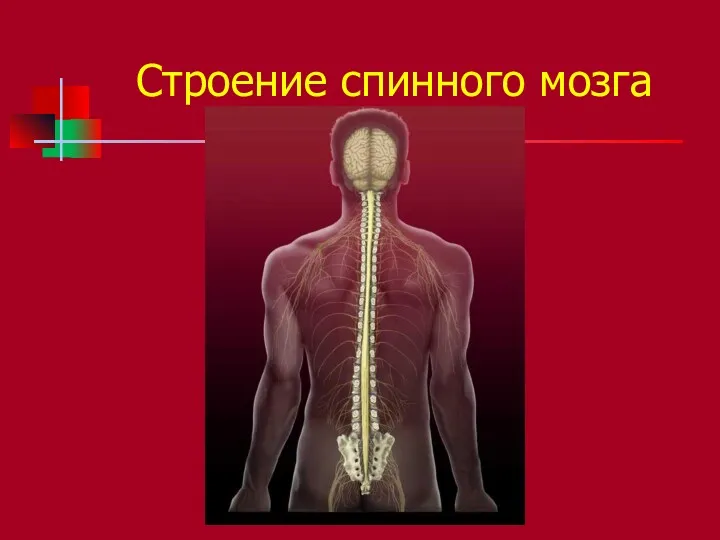
Строение спинного мозга
Строение спинного мозга
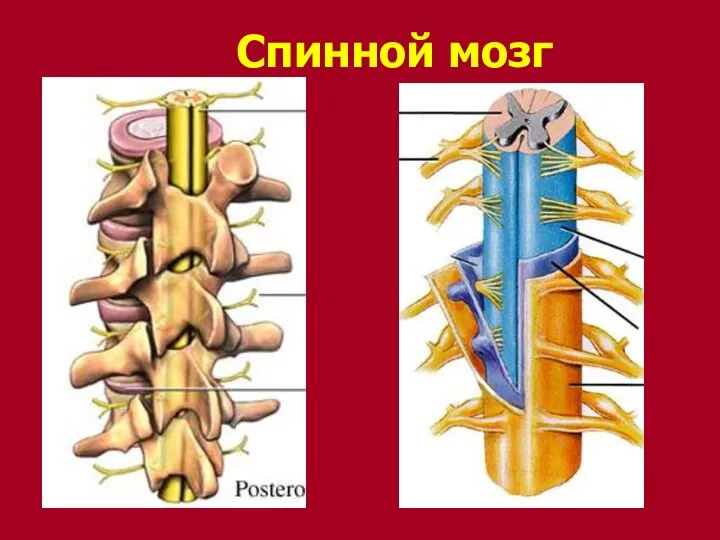
Спинной мозг
Спинной мозг
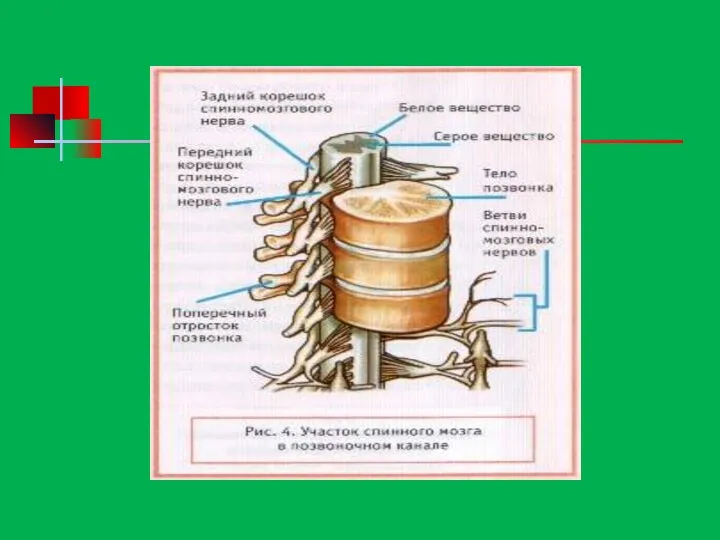

Функции спинного мозга
Рефлекторная – здесь находятся центры безусловных рефлексов
Проводниковая функция –
Функции спинного мозга
Рефлекторная – здесь находятся центры безусловных рефлексов
Проводниковая функция –
Головной мозг
Головной мозг
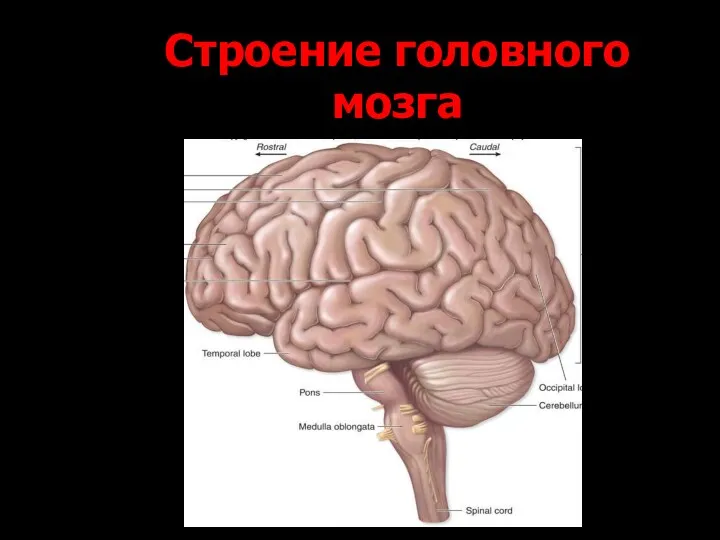
Строение головного мозга
Строение головного мозга
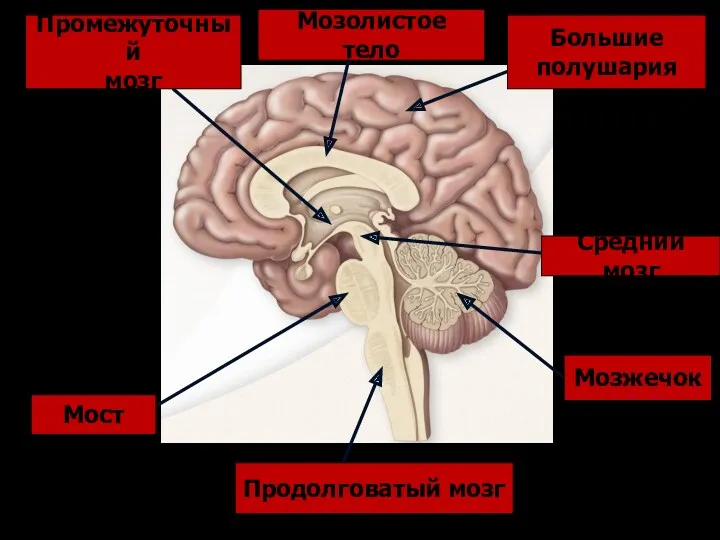
Продолговатый мозг
Мост
Мозжечок
Средний мозг
Промежуточный
мозг
Большие
полушария
Мозолистое тело
Продолговатый мозг
Мост
Мозжечок
Средний мозг
Промежуточный
мозг
Большие
полушария
Мозолистое тело
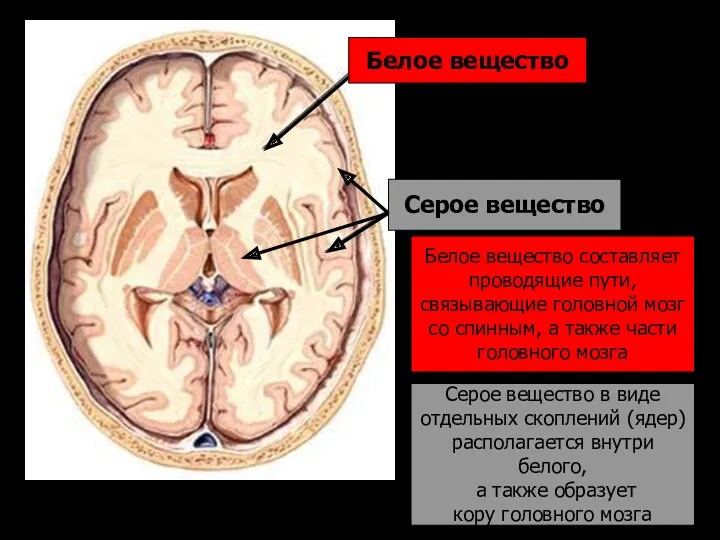
Белое вещество
Серое вещество
Белое вещество составляет
проводящие пути,
связывающие головной мозг
со
Белое вещество
Серое вещество
Белое вещество составляет
проводящие пути,
связывающие головной мозг
со
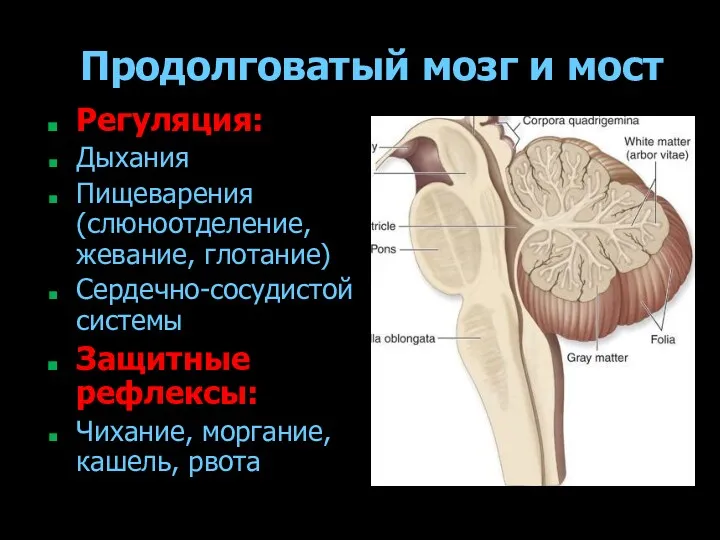
Продолговатый мозг и мост
Регуляция:
Дыхания
Пищеварения (слюноотделение, жевание, глотание)
Сердечно-сосудистой системы
Защитные рефлексы:
Чихание, моргание, кашель,
Продолговатый мозг и мост
Регуляция:
Дыхания
Пищеварения (слюноотделение, жевание, глотание)
Сердечно-сосудистой системы
Защитные рефлексы:
Чихание, моргание, кашель,
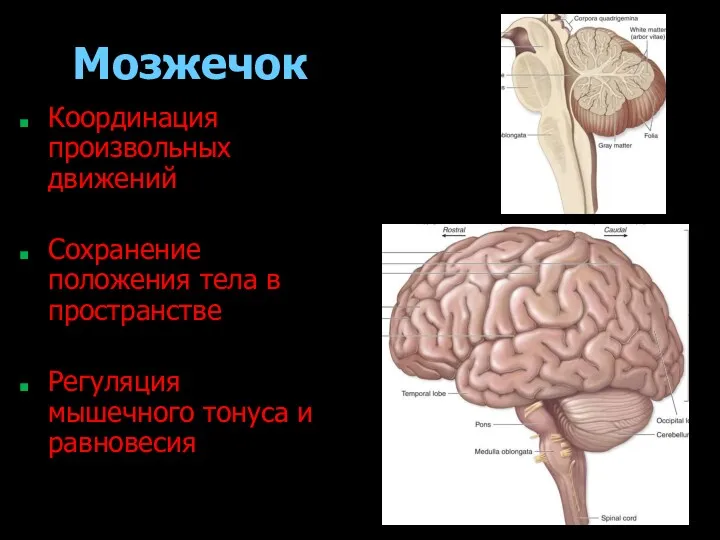
Мозжечок
Координация произвольных движений
Сохранение положения тела в пространстве
Регуляция мышечного тонуса и равновесия
Мозжечок
Координация произвольных движений
Сохранение положения тела в пространстве
Регуляция мышечного тонуса и равновесия
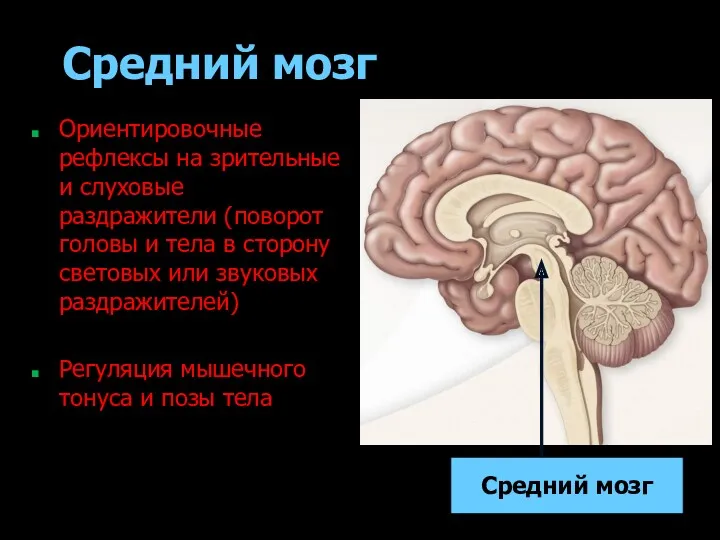
Средний мозг
Ориентировочные рефлексы на зрительные и слуховые раздражители (поворот головы и
Средний мозг
Ориентировочные рефлексы на зрительные и слуховые раздражители (поворот головы и
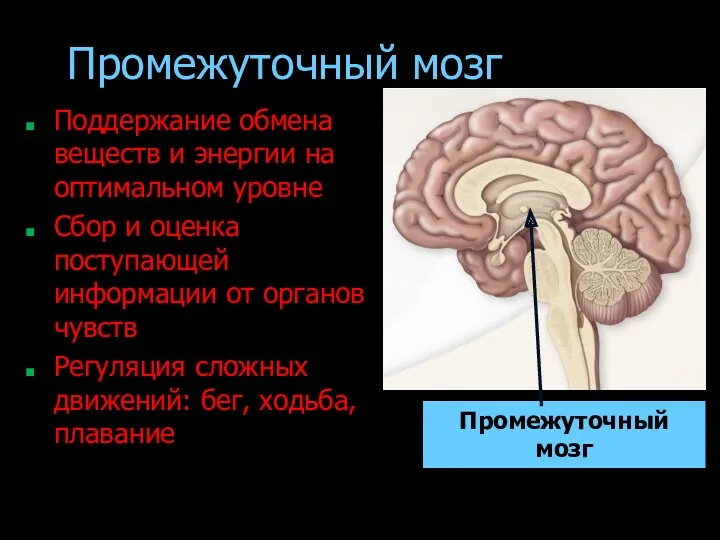
Промежуточный мозг
Поддержание обмена веществ и энергии на оптимальном уровне
Сбор и оценка
Промежуточный мозг
Поддержание обмена веществ и энергии на оптимальном уровне
Сбор и оценка
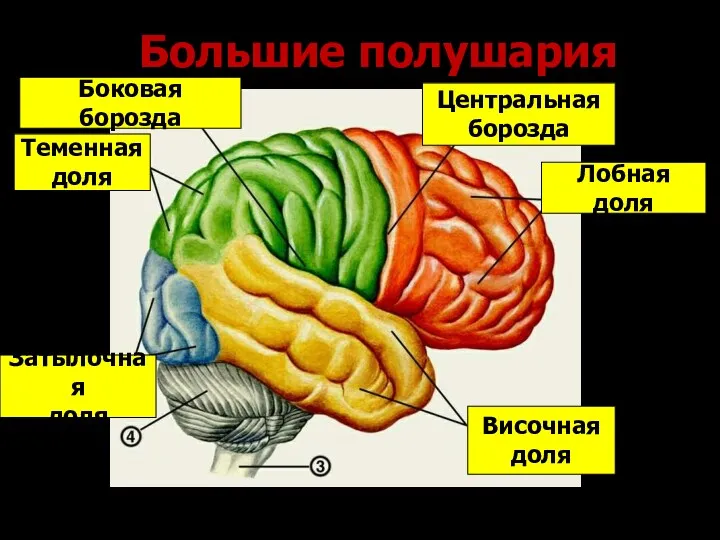
Большие полушария
Лобная доля
Центральная
борозда
Боковая борозда
Теменная
доля
Височная
доля
Затылочная
доля
Большие полушария
Лобная доля
Центральная
борозда
Боковая борозда
Теменная
доля
Височная
доля
Затылочная
доля

Затылочные доли – зрительная чувствительность
Височные доли – слуховая, вкусовая, обонятельная чувствительность
Лобные
Затылочные доли – зрительная чувствительность
Височные доли – слуховая, вкусовая, обонятельная чувствительность
Лобные

С большими полушариями мозга связаны:
Память
Речь
Мышление
Творческие процессы
Личностные качества
С большими полушариями мозга связаны:
Память
Речь
Мышление
Творческие процессы
Личностные качества
 Класс Млекопитающие (Звери)
Класс Млекопитающие (Звери) Нервная система
Нервная система Проект Геном человека - занятие по биохимии
Проект Геном человека - занятие по биохимии Отдел Мхи
Отдел Мхи Закономерности наследования признаков, установленные Г. Менделем. Моногибридное скрещивание. Первый и второй законы
Закономерности наследования признаков, установленные Г. Менделем. Моногибридное скрещивание. Первый и второй законы Поток энергии и цепи питания.
Поток энергии и цепи питания. Двигательные системы
Двигательные системы Экскурсия в науку (по кабинету биологии)
Экскурсия в науку (по кабинету биологии) Сущность жизни и свойства живого
Сущность жизни и свойства живого Қазақстандағы дәрілік өсімдіктер
Қазақстандағы дәрілік өсімдіктер Алгоритм решения задания №27 на антипараллельность
Алгоритм решения задания №27 на антипараллельность железы внутренней секреции
железы внутренней секреции Популяциядағы жүретін процестерді зерттеу
Популяциядағы жүретін процестерді зерттеу Різновиди хижих рослин
Різновиди хижих рослин Мейоз
Мейоз Как размножаются живые организмы
Как размножаются живые организмы Устройство пищевода
Устройство пищевода Физиология и экология диатомовых водорослей
Физиология и экология диатомовых водорослей Геоботаникаю лекция 2
Геоботаникаю лекция 2 Выступление на всероссийском семинаре Личностно- ориентированнный подход при создании проектных работ.
Выступление на всероссийском семинаре Личностно- ориентированнный подход при создании проектных работ. Домашние животные и птицы
Домашние животные и птицы Кальциевые каналы
Кальциевые каналы Сексована сперма
Сексована сперма Физиология сенсорных систем
Физиология сенсорных систем Наши домашние питомцы (1 класс)
Наши домашние питомцы (1 класс) Вода. Свойства воды. Тест
Вода. Свойства воды. Тест Сущность жизни и свойства живого
Сущность жизни и свойства живого Мезозойская эра
Мезозойская эра