История развития энзимологии. Номенклатура, классификация, структура и функции ферментов. Лекция 1 презентация
- История развития энзимологии. Номенклатура, классификация, структура и функции ферментов. Лекция 1

Содержание
- 2. Ферменты или энзимы (Е) – биокатализаторы, от латинского слова «fermentum» - закваска; от греческого «en zyme»
- 3. В 1783 г. итальянский ученый Спаланцани, изучая переваривание мяса у хищных птиц, показал, что в организме
- 4. В 1814 г. русский ученый, К. Кирхгоф впервые установил, что крахмал превращается в сахар под действием
- 5. В 1833 г. французские химики А. Пайен и Ж. Пирсо выделили вещество, расщепляющее крахмал, которое они
- 6. Луи Пастер высказал предположение, что процессы брожения могут вызывать микроорганизмы и, следовательно, связаны лишь с их
- 7. Ю. Либих и К. Бернар отстаивали химическую природу брожения, считая, что брожение вызывают особые вещества, подобные
- 8. В 1837 г. шведский химик Й. Берцелиус показал, что ферменты – это катализаторы, поставляемые живыми клетками.
- 9. В 1897 г. немецкие ученые Ганс и Эдвард Бухнеры показали, что дрожжевой бесклеточный сок способен Э.
- 10. После этого стало ясно, что в нем содержится смесь ферментов («зимаза»), которые функционируют как внутри, так
- 11. В 1926 г. американский биохимик Д. Самнер выделил из бобов канавалии фермент уреазу, катализирующий реакцию расщепления
- 12. В 1930 г., норвежец Д. Нортроп выделил фермент пепсин, а затем трипсин и химотрипсин
- 13. С этого периода стало общепринятым утверждение, что все ферменты являются белками
- 14. Большинство ферментов белки Но есть ферменты небелковой природы. Н-р, рибозимы, состоящие из РНК. Есть ферменты, которые
- 15. В 1961 г. в Москве на 5-ом биохимическом конгрессе Международная комиссия по ферментам предложила рекомендации по
- 16. Номенклатура ферментов Каждый фермент имеет два названия: 1 – рабочее (короткое), 2 – систематическое (более полное),
- 17. 1 тип названия находится в зависимости от субстрата, на который действует фермент с добавлением суффикса «аза»
- 18. Некоторые ферменты имеют специфическое название: например, пепсин, трипсин и т.д.
- 19. 2 тип включает в себя название субстрата + название реакции, катализируемой данным ферментом Например, лактатдегидрогеназа
- 20. Классификация ферментов Все ферменты разделены на 6 классов, каждый из которых имеет строго определенный номер. Классы
- 21. Для каждого фермента существует специальный шифр, состоящий из 4-х цифр: 1 – номер класса, 2 –
- 23. Например: 1.1.1.1. – алкогольдегидрогеназа (АДГ) 1.1.1.27 – лактатдегидрогеназа (ЛДГ)
- 24. Классы ферментов: I. Оксидоредуктазы II. Трансферазы III. Гидролазы IV. Лиазы V. Изомеразы VI. Лигазы (синтетазы)
- 25. I. Оксидоредуктазы Катализируют окислительно-восстановительные реакции с участием двух субстратов (перенос электронов или атомов водорода с одного
- 26. В классе 17 подклассов. Например: 1. Дегидрогеназы Сюда входят ферменты, катализирующие реакции дегидрирования (отщепления водорода). В
- 27. Пример реакции:
- 28. 2. Оксидазы Акцептором ē служит молекулярный кислород. Пример реакции: О2 + 4 Н+ + 4 е
- 29. 3. Оксигеназы (гидроксилазы) Атом кислорода из молекулы кислорода присоединяется к субстрату
- 30. Пример реакции:
- 31. II. Трансферазы Катализируют перенос функциональных групп от одного соединения к другому. В зависимости от переносимой группы:
- 32. Пример реакции:
- 33. III. Гидролазы Катализилируют реакции гидролиза (расщепление ковалентных связей с присоединением молекулы воды по месту разрыва). Например:
- 34. Например:
- 35. IV. Лиазы Катализируют отщепление от субстратов определенные группы (СО2, Н2О, NH2, SH2 и др.) без участия
- 36. Примеры реакций:
- 38. V. Изомеразы Катализируют р-ции изомеризации Например:
- 39. VI. Лигазы (синтетазы) Катализируют реакции синтеза различных веществ при участии АТФ
- 40. Например:
- 41. Структура ферментов Простые ферменты состоят только из белка (пепсин, трипсин, папаин, рибонуклеаза, фосфатаза, уреаза и др.)
- 42. Если константа диссоциации (Kd) комплекса «белок-кофактор» очень мала и обе части не разделяются при выделении и
- 43. Другие термины небелковой части: Кофермент (коэнзим) - часть некоторых ферментов, которая легко отделяется от белковой части
- 44. Коферменты: производные витаминов; гемы, входящие в состав цитохромов, каталазы, пероксидазы, гуанилатциклазы, NO-синтазы; нуклеотиды – доноры и
- 45. Убихинон (кофермент Q), участвующий в переносе ē и Н+ в дыхательной цепи; Фосфоаденозилфосфосульфат (ФАФС), участвующий в
- 46. Активный центр фермента (А) А – участок ответственный за присоединение субстрата (S) и его химическое превращение.
- 47. 1. Каталитический центр – участок, непосредственно вступающий в химическое взаимодействие с субстратом (S) 2. Связывающий центр
- 48. Простой фермент: 1-каталитический участок, 2 - контактный
- 49. Сложный фермент: 1-каталитический участок, 2 – контактный, 3 - кофермент
- 50. Аллостерический (регуляторный) фермент: R - аллостерический (регуляторный) центр
- 51. В активном центре обычно 12-16 аминокислотных остатков АМК А находятся в различных местах полипептидной цепи, нередко
- 55. У сложных ферментов главную роль контактных и каталитических центров активного центра играет кофермент
- 56. А может содержать различные функциональные группы: NH2 (лизина, гуанидиновых групп аргинина, концевых аминокислот), COOH (дикарбоновых и
- 57. имидазольные гистидина, тиоэфирные метионина, фенольные группы тирозина, гидрофобные цепи алифатических аминокислот, ароматическое кольцо фенилаланина
- 58. Взаимодействие субстрата с активным центром фермента
- 59. Свойства ферментов. Общность и отличия их от неорганических катализаторов
- 60. Сходство с неорганическими катализаторами : Повышают скорость химической реакции, но не являются их инициаторами и не
- 61. Отличия: Ферменты обладают намного большей активностью Неорганические катализаторы активны в очень жестких условиях (высокие t, давление,
- 62. Ферменты обладают специфичностью, т.е. способностью катализировать строго определенные реакции, в которые вовлечены только субстраты, взаимодействующие с
- 63. Виды специфичности Абсолютная (индивидуальная) – когда фермент действует только на единственный субстрат
- 64. Групповая (относительная) – когда фермент действует на группу субстратов или на один вид связи. Например, пепсин
- 65. Стереохимическая – когда фермент действует только на один из изомеров. Самая высокая специфичность
- 66. Выделяют: Стереоспецифичность к одному из оптических стереоизомеров: - к D-сахарам - к L-аминокислотам
- 67. Например:
- 68. Стереоспецифичность к одному из геометрических стереоизомеров: - к цис-транс-изомерам - к α- и β-гликозидным связям (например,
- 69. Пример стереоспецифичности к транс-изомеру
- 70. Различают две основные теории специфичности ферментов: «жесткого соответствия» «индуцированного соответствия»
- 71. 1-ая теория предложена Э. Фишером предусматривает наличие абсолютного совпадения А и S («ключ-замок»): S является как
- 74. Скачать презентацию

Ферменты или энзимы (Е) – биокатализаторы, от латинского слова «fermentum» -
Ферменты или энзимы (Е) – биокатализаторы, от латинского слова «fermentum» -

В 1783 г. итальянский ученый Спаланцани, изучая переваривание мяса у хищных
В 1783 г. итальянский ученый Спаланцани, изучая переваривание мяса у хищных

В 1814 г. русский ученый, К. Кирхгоф впервые установил, что крахмал
В 1814 г. русский ученый, К. Кирхгоф впервые установил, что крахмал

В 1833 г. французские химики А. Пайен и Ж. Пирсо выделили
В 1833 г. французские химики А. Пайен и Ж. Пирсо выделили

Луи Пастер высказал предположение, что процессы брожения могут вызывать микроорганизмы
Луи Пастер высказал предположение, что процессы брожения могут вызывать микроорганизмы

Ю. Либих и К. Бернар отстаивали химическую природу брожения, считая, что
Ю. Либих и К. Бернар отстаивали химическую природу брожения, считая, что

В 1837 г. шведский химик Й. Берцелиус показал, что ферменты
В 1837 г. шведский химик Й. Берцелиус показал, что ферменты

В 1897 г. немецкие ученые Ганс и Эдвард Бухнеры показали,
В 1897 г. немецкие ученые Ганс и Эдвард Бухнеры показали,

После этого стало ясно, что в нем содержится смесь ферментов («зимаза»),
После этого стало ясно, что в нем содержится смесь ферментов («зимаза»),

В 1926 г. американский биохимик Д. Самнер выделил из бобов
В 1926 г. американский биохимик Д. Самнер выделил из бобов

В 1930 г., норвежец
Д. Нортроп
выделил фермент
пепсин,
В 1930 г., норвежец
Д. Нортроп
выделил фермент
пепсин,

С этого периода стало общепринятым утверждение, что все ферменты являются белками
С этого периода стало общепринятым утверждение, что все ферменты являются белками

Большинство ферментов белки
Но есть ферменты небелковой природы.
Н-р, рибозимы, состоящие из
Большинство ферментов белки
Но есть ферменты небелковой природы.
Н-р, рибозимы, состоящие из

В 1961 г. в Москве на 5-ом биохимическом конгрессе Международная комиссия
В 1961 г. в Москве на 5-ом биохимическом конгрессе Международная комиссия

Номенклатура ферментов
Каждый фермент имеет два названия: 1 – рабочее (короткое), 2
Номенклатура ферментов
Каждый фермент имеет два названия: 1 – рабочее (короткое), 2

1 тип названия находится в зависимости от субстрата, на который действует
1 тип названия находится в зависимости от субстрата, на который действует

Некоторые ферменты имеют специфическое название: например, пепсин, трипсин и т.д.
Некоторые ферменты имеют специфическое название: например, пепсин, трипсин и т.д.

2 тип включает в себя название субстрата + название реакции, катализируемой
2 тип включает в себя название субстрата + название реакции, катализируемой

Классификация ферментов
Все ферменты разделены на 6 классов, каждый из которых имеет
Классификация ферментов
Все ферменты разделены на 6 классов, каждый из которых имеет

Для каждого фермента существует специальный шифр, состоящий из 4-х цифр: 1
Для каждого фермента существует специальный шифр, состоящий из 4-х цифр: 1
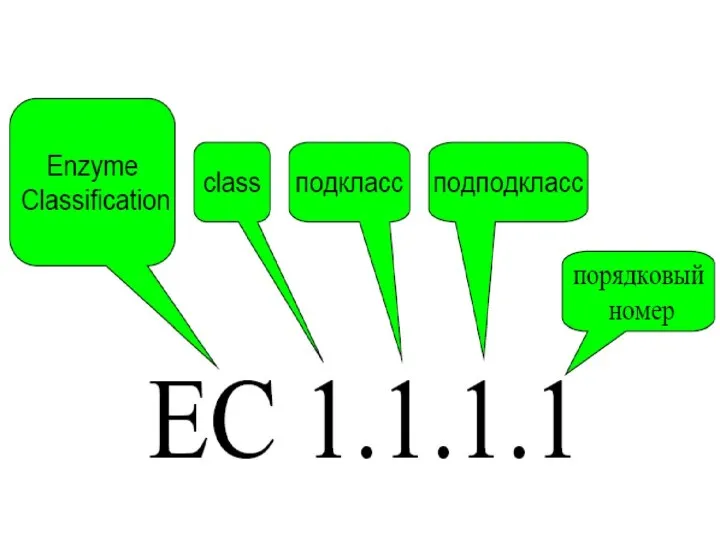
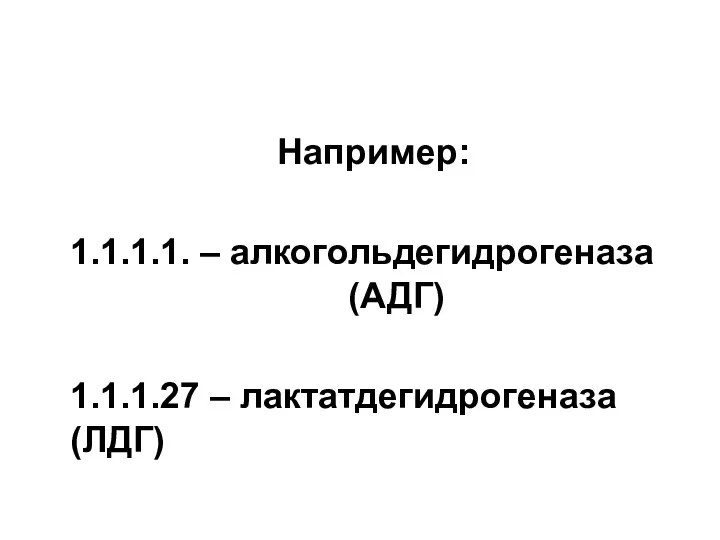
Например:
1.1.1.1. – алкогольдегидрогеназа (АДГ)
1.1.1.27 – лактатдегидрогеназа (ЛДГ)
Например:
1.1.1.1. – алкогольдегидрогеназа (АДГ)
1.1.1.27 – лактатдегидрогеназа (ЛДГ)

Классы ферментов:
I. Оксидоредуктазы
II. Трансферазы
III. Гидролазы
IV. Лиазы
V. Изомеразы
VI. Лигазы (синтетазы)
Классы ферментов:
I. Оксидоредуктазы
II. Трансферазы
III. Гидролазы
IV. Лиазы
V. Изомеразы
VI. Лигазы (синтетазы)

I. Оксидоредуктазы
Катализируют окислительно-восстановительные реакции с участием двух субстратов (перенос электронов или
I. Оксидоредуктазы
Катализируют окислительно-восстановительные реакции с участием двух субстратов (перенос электронов или

В классе 17 подклассов. Например:
1. Дегидрогеназы Сюда входят ферменты, катализирующие реакции
В классе 17 подклассов. Например:
1. Дегидрогеназы Сюда входят ферменты, катализирующие реакции
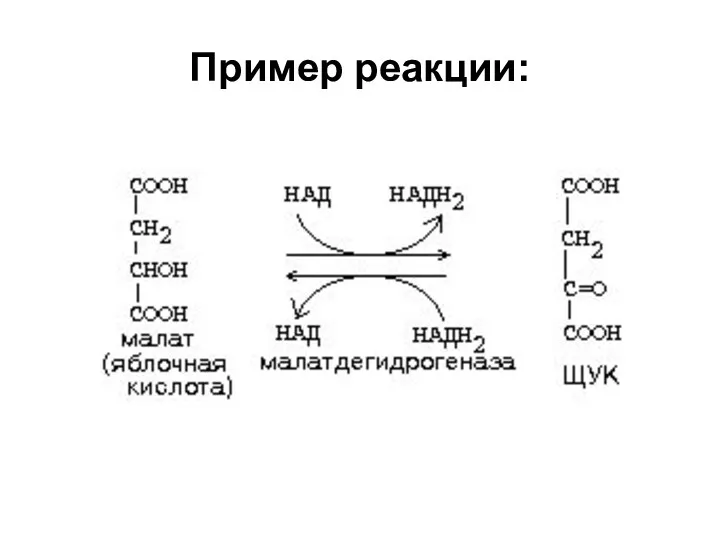
Пример реакции:
Пример реакции:

2. Оксидазы Акцептором ē служит молекулярный кислород.
Пример реакции:
О2 + 4 Н+
2. Оксидазы Акцептором ē служит молекулярный кислород.
Пример реакции:
О2 + 4 Н+

3. Оксигеназы (гидроксилазы)
Атом кислорода из молекулы кислорода присоединяется к субстрату
3. Оксигеназы (гидроксилазы)
Атом кислорода из молекулы кислорода присоединяется к субстрату
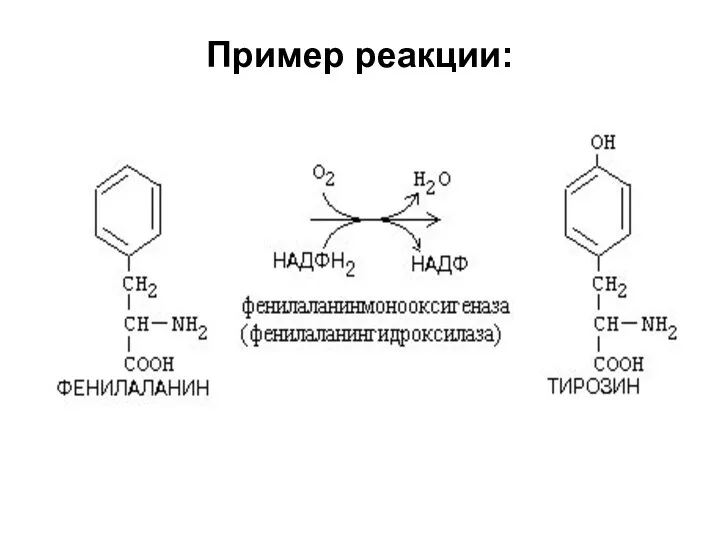
Пример реакции:
Пример реакции:

II. Трансферазы
Катализируют перенос функциональных групп от одного соединения к другому.
В
II. Трансферазы
Катализируют перенос функциональных групп от одного соединения к другому.
В
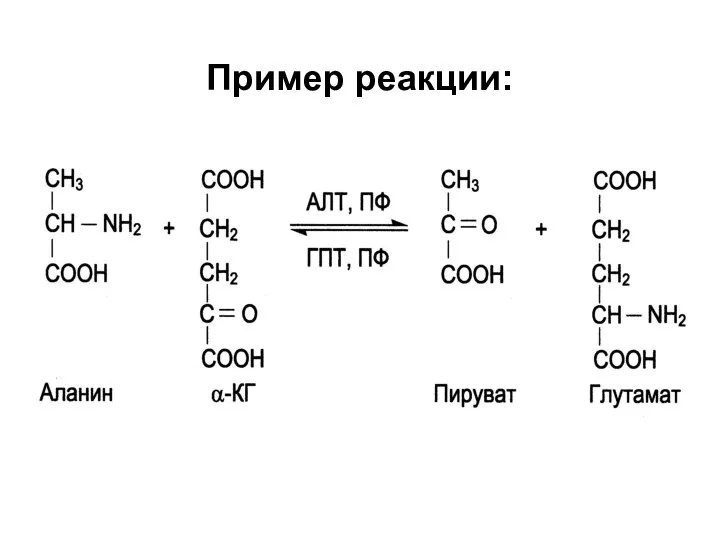
Пример реакции:
Пример реакции:

III. Гидролазы
Катализилируют реакции гидролиза (расщепление ковалентных связей с присоединением молекулы воды по
III. Гидролазы
Катализилируют реакции гидролиза (расщепление ковалентных связей с присоединением молекулы воды по
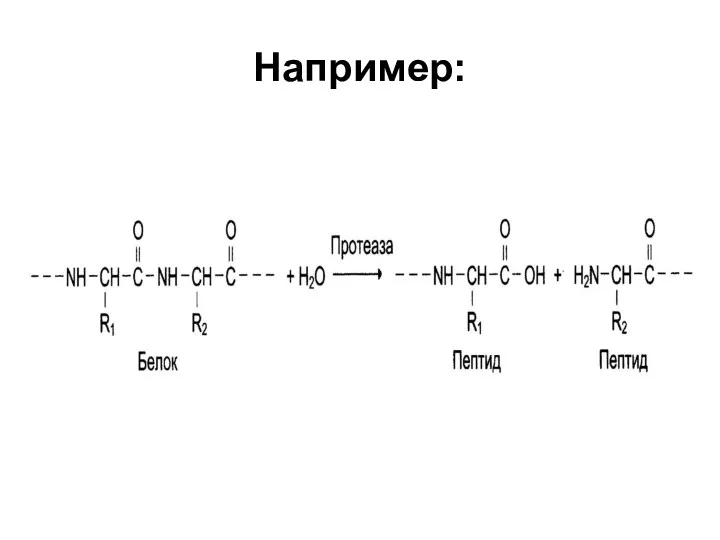
Например:
Например:

IV. Лиазы
Катализируют отщепление от субстратов определенные группы (СО2, Н2О, NH2, SH2 и
IV. Лиазы
Катализируют отщепление от субстратов определенные группы (СО2, Н2О, NH2, SH2 и
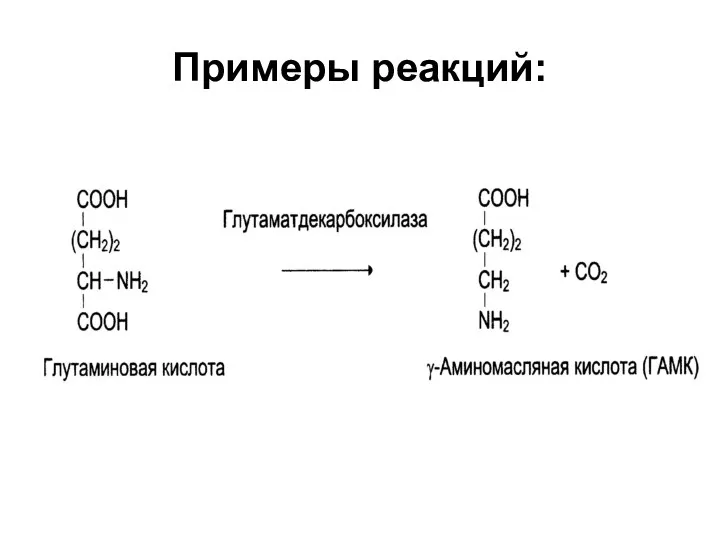
Примеры реакций:
Примеры реакций:
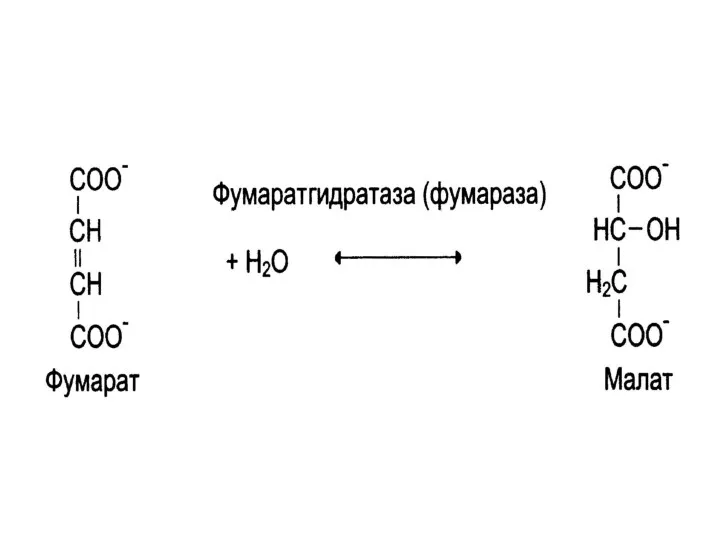
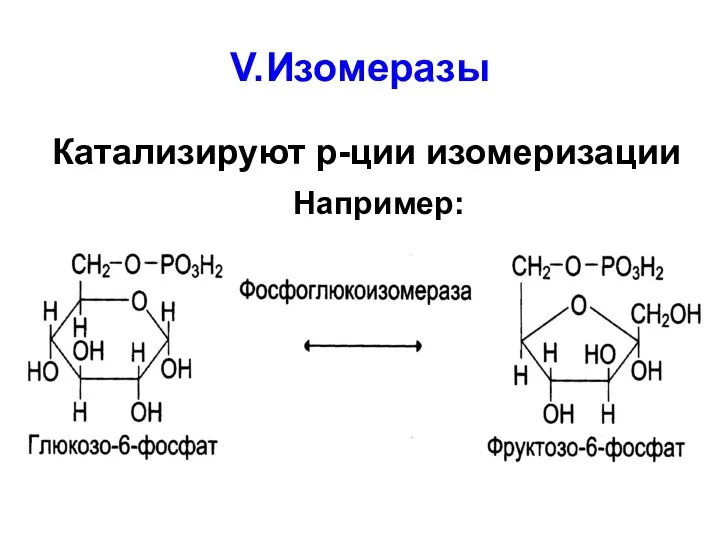
V. Изомеразы
Катализируют р-ции изомеризации
Например:
V. Изомеразы
Катализируют р-ции изомеризации
Например:

VI. Лигазы (синтетазы)
Катализируют реакции синтеза
различных веществ при участии
АТФ
VI. Лигазы (синтетазы)
Катализируют реакции синтеза
различных веществ при участии
АТФ
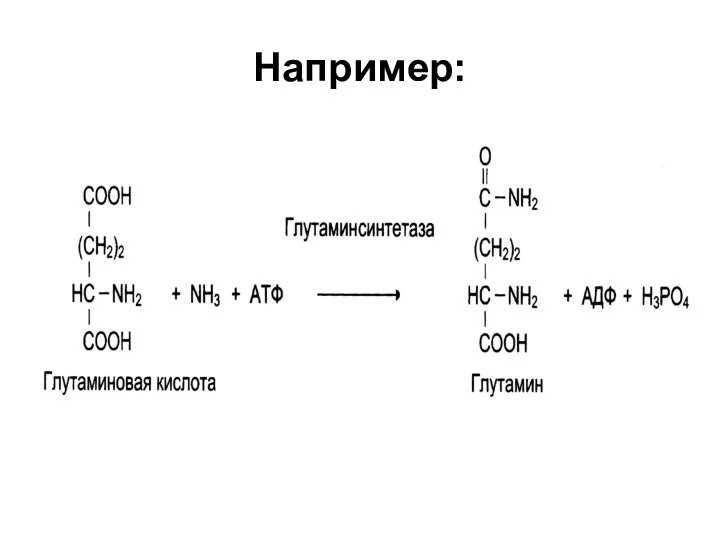
Например:
Например:

Структура ферментов
Простые ферменты состоят только из белка (пепсин, трипсин, папаин, рибонуклеаза,
Структура ферментов
Простые ферменты состоят только из белка (пепсин, трипсин, папаин, рибонуклеаза,

Если константа диссоциации (Kd) комплекса «белок-кофактор» очень мала и обе части
Если константа диссоциации (Kd) комплекса «белок-кофактор» очень мала и обе части

Другие термины небелковой части:
Кофермент (коэнзим) - часть некоторых ферментов, которая легко
Другие термины небелковой части:
Кофермент (коэнзим) - часть некоторых ферментов, которая легко

Коферменты:
производные витаминов;
гемы, входящие в состав цитохромов, каталазы, пероксидазы, гуанилатциклазы, NO-синтазы;
нуклеотиды –
Коферменты:
производные витаминов;
гемы, входящие в состав цитохромов, каталазы, пероксидазы, гуанилатциклазы, NO-синтазы;
нуклеотиды –

Убихинон (кофермент Q), участвующий в переносе ē и Н+ в дыхательной
Убихинон (кофермент Q), участвующий в переносе ē и Н+ в дыхательной

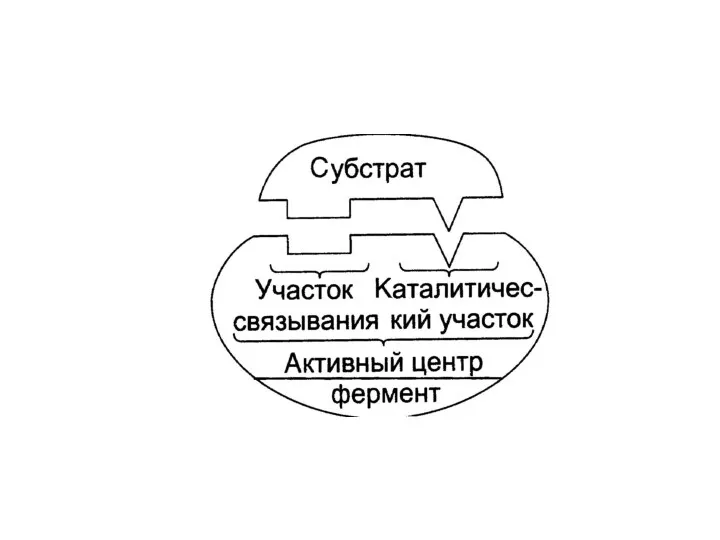
Активный центр фермента (А)
А – участок ответственный за присоединение субстрата (S)
Активный центр фермента (А)
А – участок ответственный за присоединение субстрата (S)

1. Каталитический центр – участок, непосредственно вступающий в химическое взаимодействие с
1. Каталитический центр – участок, непосредственно вступающий в химическое взаимодействие с
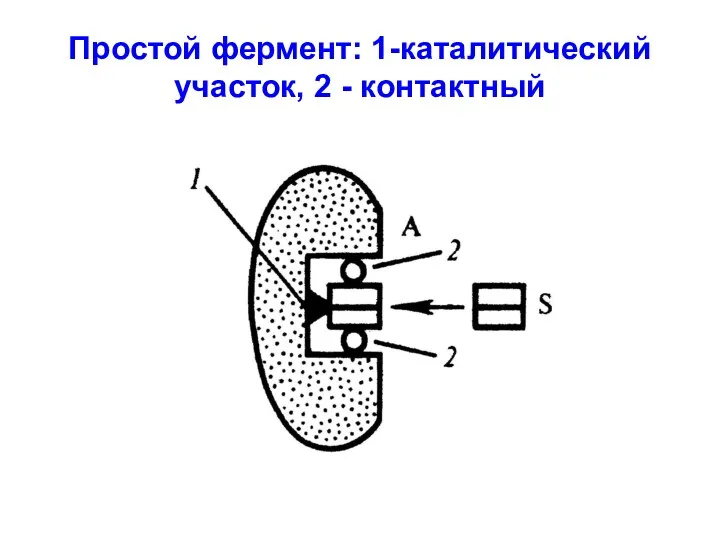
Простой фермент: 1-каталитический участок, 2 - контактный
Простой фермент: 1-каталитический участок, 2 - контактный
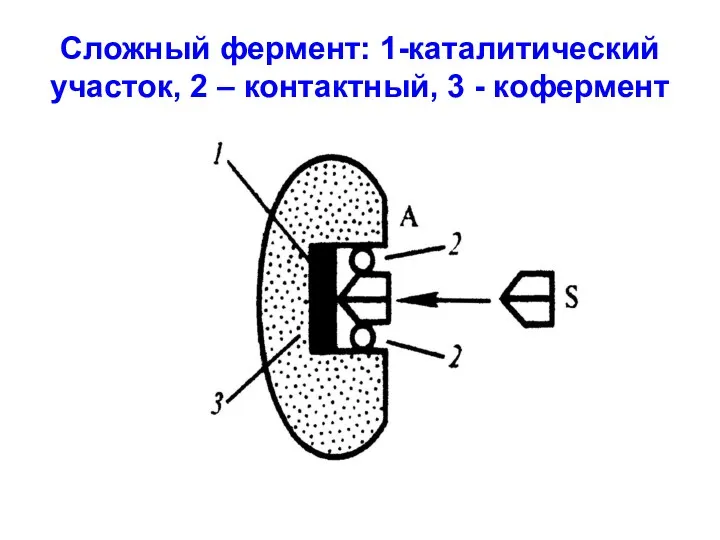
Сложный фермент: 1-каталитический участок, 2 – контактный, 3 - кофермент
Сложный фермент: 1-каталитический участок, 2 – контактный, 3 - кофермент
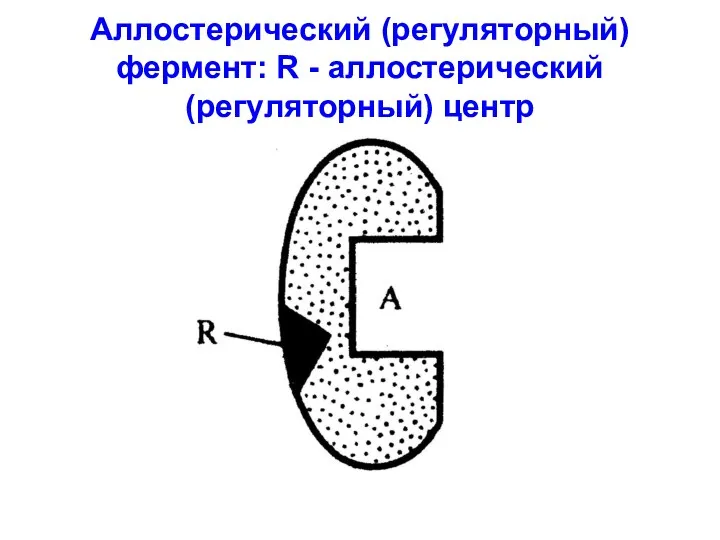
Аллостерический (регуляторный) фермент: R - аллостерический (регуляторный) центр
Аллостерический (регуляторный) фермент: R - аллостерический (регуляторный) центр

В активном центре обычно 12-16 аминокислотных остатков
АМК А находятся в различных
В активном центре обычно 12-16 аминокислотных остатков
АМК А находятся в различных
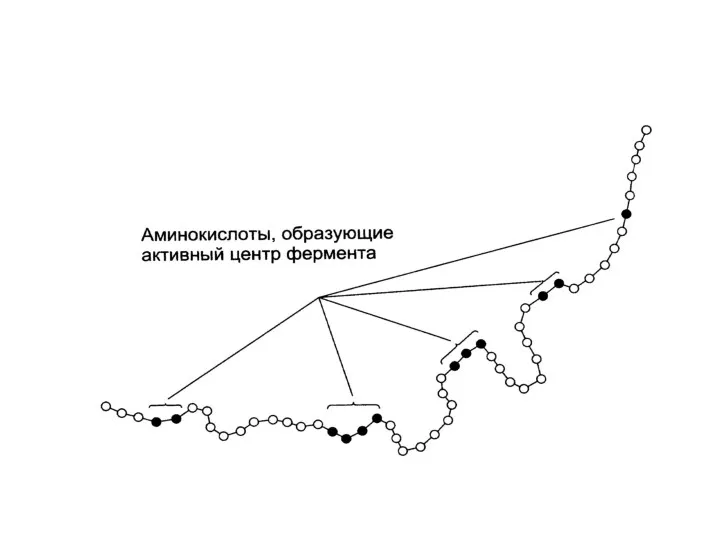
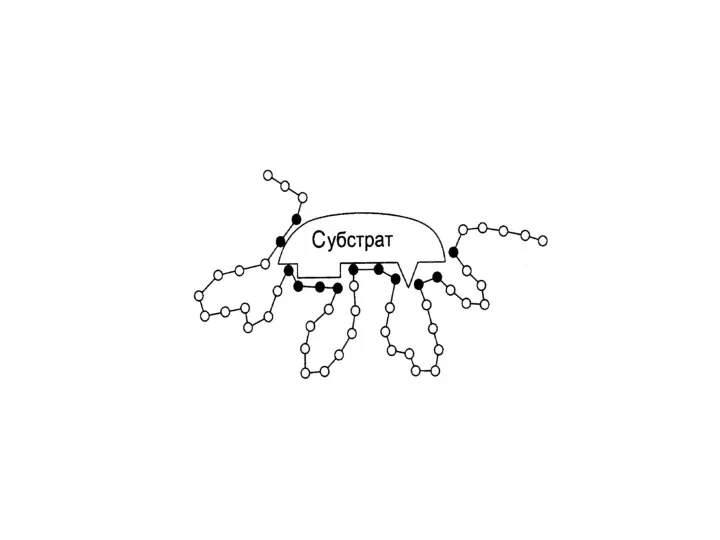

У сложных ферментов главную роль контактных и каталитических центров активного центра
У сложных ферментов главную роль контактных и каталитических центров активного центра

А может содержать различные функциональные группы:
NH2 (лизина, гуанидиновых групп аргинина,
А может содержать различные функциональные группы:
NH2 (лизина, гуанидиновых групп аргинина,

имидазольные гистидина,
тиоэфирные метионина,
фенольные группы тирозина,
гидрофобные цепи алифатических аминокислот,
ароматическое
имидазольные гистидина,
тиоэфирные метионина,
фенольные группы тирозина,
гидрофобные цепи алифатических аминокислот,
ароматическое
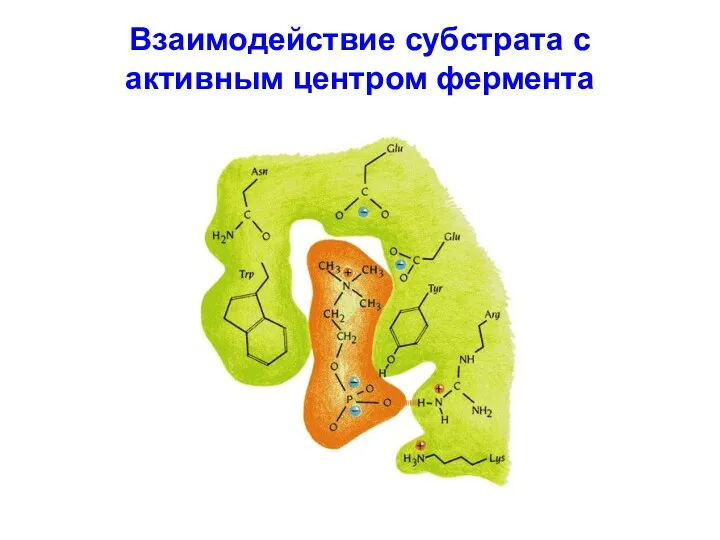
Взаимодействие субстрата с
активным центром фермента
Взаимодействие субстрата с
активным центром фермента

Свойства ферментов.
Общность и отличия их от
неорганических катализаторов
Свойства ферментов.
Общность и отличия их от
неорганических катализаторов

Сходство с неорганическими катализаторами :
Повышают скорость химической реакции, но не являются
Сходство с неорганическими катализаторами :
Повышают скорость химической реакции, но не являются

Отличия:
Ферменты обладают намного большей активностью
Неорганические катализаторы активны в очень жестких условиях
Отличия:
Ферменты обладают намного большей активностью
Неорганические катализаторы активны в очень жестких условиях

Ферменты обладают специфичностью, т.е. способностью катализировать строго определенные реакции, в которые
Ферменты обладают специфичностью, т.е. способностью катализировать строго определенные реакции, в которые
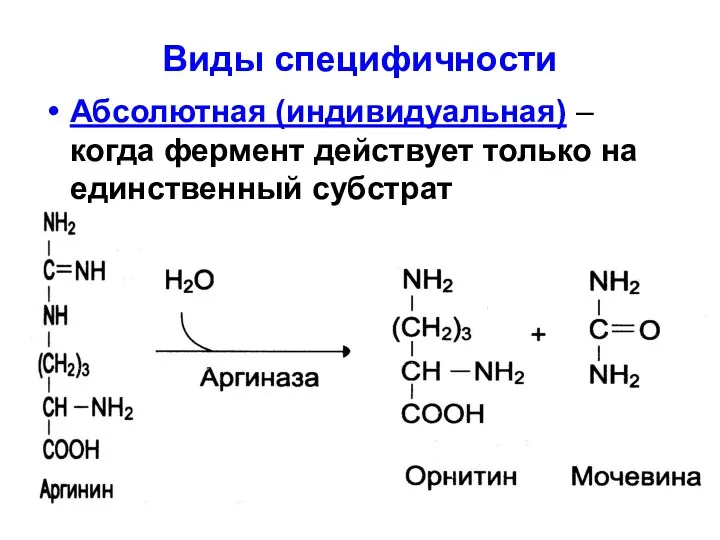
Виды специфичности
Абсолютная (индивидуальная) – когда фермент действует только на единственный
Виды специфичности
Абсолютная (индивидуальная) – когда фермент действует только на единственный

Групповая (относительная) – когда фермент действует на группу субстратов или на
Групповая (относительная) – когда фермент действует на группу субстратов или на

Стереохимическая – когда фермент действует только на один из изомеров.
Самая
Стереохимическая – когда фермент действует только на один из изомеров.
Самая

Выделяют:
Стереоспецифичность к одному из оптических стереоизомеров:
- к D-сахарам
- к
Выделяют:
Стереоспецифичность к одному из оптических стереоизомеров:
- к D-сахарам
- к
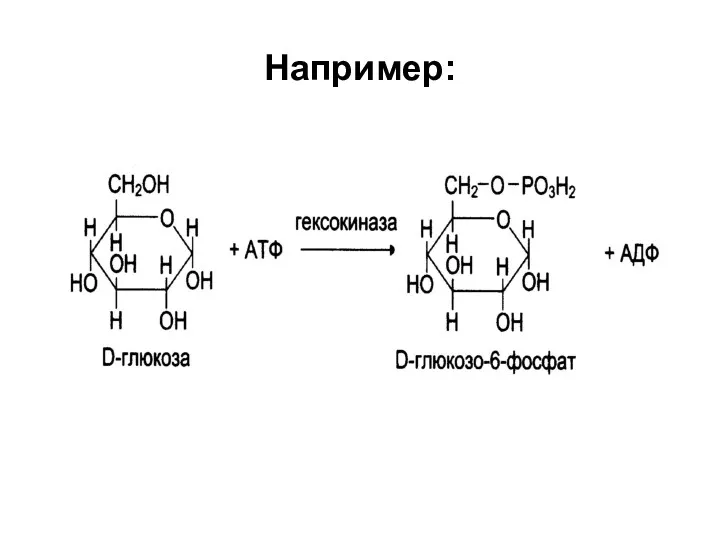
Например:
Например:

Стереоспецифичность к одному из геометрических стереоизомеров:
- к цис-транс-изомерам
- к α-
Стереоспецифичность к одному из геометрических стереоизомеров:
- к цис-транс-изомерам
- к α-
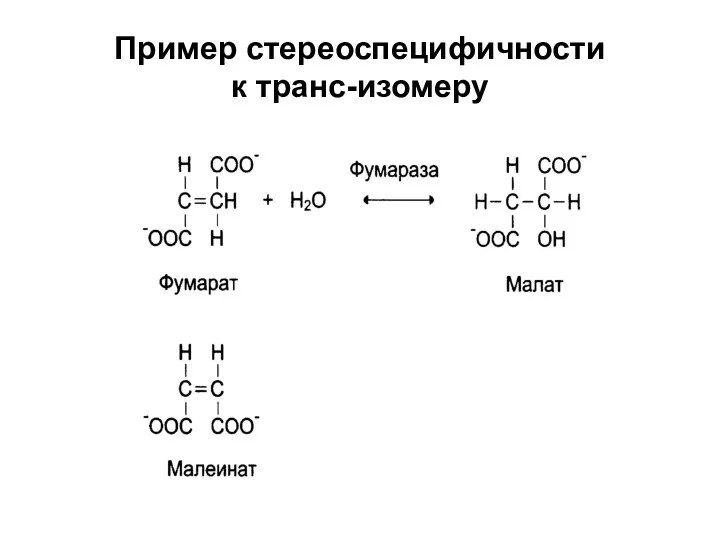
Пример стереоспецифичности к транс-изомеру
Пример стереоспецифичности к транс-изомеру

Различают две основные теории специфичности ферментов:
«жесткого соответствия»
«индуцированного соответствия»
Различают две основные теории специфичности ферментов:
«жесткого соответствия»
«индуцированного соответствия»

1-ая теория предложена Э. Фишером
предусматривает наличие абсолютного совпадения А и S
1-ая теория предложена Э. Фишером
предусматривает наличие абсолютного совпадения А и S

 Растительный мир Дальнего Востока
Растительный мир Дальнего Востока Постановка ИФА. Учет результатов реакций
Постановка ИФА. Учет результатов реакций Свойства живого
Свойства живого презентация Клетка
презентация Клетка Животные из красной книги
Животные из красной книги Картирование генов. Построение генетических и цитологических карт
Картирование генов. Построение генетических и цитологических карт 27 февраля – День белого медведя
27 февраля – День белого медведя Все о витамине С и его определение в лекарственных растениях и продуктах питания
Все о витамине С и его определение в лекарственных растениях и продуктах питания Периодизация и продолжительность жизни животных
Периодизация и продолжительность жизни животных Нормальная анатомия легких, плевры и средостения
Нормальная анатомия легких, плевры и средостения Причины разнообразия жизни на Земле
Причины разнообразия жизни на Земле Белки, жиры, углеводы
Белки, жиры, углеводы Биологическая безопасность в биотехнологии
Биологическая безопасность в биотехнологии Царство грибы
Царство грибы Қартаюдың молекулалық генетикалық механизмдері
Қартаюдың молекулалық генетикалық механизмдері Царство Животные Подцарство Простейшие (Одноклеточные)
Царство Животные Подцарство Простейшие (Одноклеточные) Органічні сполуки. Мікроелементи. Клітинна мембрана
Органічні сполуки. Мікроелементи. Клітинна мембрана Здоровьесберегающий подход к выбору туалетного мыла
Здоровьесберегающий подход к выбору туалетного мыла Изоляция
Изоляция Экологические группы растений
Экологические группы растений Система органов человека
Система органов человека Детям о насекомых
Детям о насекомых презентация Птицы нашего края
презентация Птицы нашего края 9 Обмен углеводов 1ч
9 Обмен углеводов 1ч Покровы тела. Сравнительная характеристика
Покровы тела. Сравнительная характеристика Естественный отбор и борьба за существование
Естественный отбор и борьба за существование Specimen Preparation for SEM investigation
Specimen Preparation for SEM investigation Пищеварительная система
Пищеварительная система