- Изучение роли генетических составляющих в этиологии и патогенезе заболеваний человека

Содержание
- 2. Современное представление «центральной догмы» молекулярной биологии
- 3. Уровни геномных исследований
- 4. Предрасположенность человека к тем или иным заболеваниям генетически детерминирована. Выделяют собственно наследственные болезни, включающие хромосомные и
- 5. Моногенные и полигенные заболевания Моногенные заболевания Мутации могут захватывать один или оба аллеля. Наследуются в соответствии
- 6. Примеры моногенных заболеваний сердечно-сосудистой системы
- 7. Распространенность полигенных заболеваний Частота наследственных заболеваний составляет 1,5%. На долю хромосомных болезней приходится 0,5%. На долю
- 8. Генетический полиморфизм: Генетический полиморфизм - это сосуществование в пределах популяции двух или нескольких различных наследственных форм,
- 9. Соотношение Харди — Вайнберга В популяции частоты генов и генотипов остаются постоянными от поколения к поколению.
- 10. Мутация или полиморфизм? Мутация - наследуемое изменение в нуклеотидной последовательности нуклеиновой кислоты, ответственной за хранение и
- 11. Причины генетического полиморфизма Генетический полиморфизм может быть обусловлен: заменой нуклеотидов, дупликацией, вставками, делециями, нуклетидными повторами. Большую
- 12. Влияние различных факторов на развитие полигенных заболеваний Наличие генов предрасположенности к заболеванию не означает, что данное
- 13. Участие факторов внешней среды в развитии полигенных заболеваний Факторы внешней среды делятся на: биологические нерациональное питание
- 14. Основные факторы риска сердечно-сосудистых заболеваний Немодифицируемые наследственность пол и возраст мужчины старше 45 лет, женщины старше
- 15. Стратегические этапы исследования генетической предрасположенности к полигенным заболеваниям Анализ ассоциации заболевания с полиморфными вариантами отдельных генов-кандидатов:
- 16. Полный геномный поиск ассоциации областей генома с заболеванием (GWAS) Группы более 1000 человек. Пациенты и контр.
- 17. синим обозначены маркеры генетической предрасположенности к инфаркту миокарда, оранжевым – к сердечной недостаточности (по O'Donnell CJ
- 18. Локусы, ассоциированные с ишемической болезнью сердца и с инфарктом миокарда Идентифицировано более 50 локусов, ассоциированные с
- 19. Проблема «потерянной» наследуемости» Проект «Геном человека» (The Human Genome Project, HGP) начался в 1990 году, под
- 20. Что такое наследуемость Наследуемость (heritability) - доля фенотипической изменчивости в популяции, обусловленная генетической изменчивостью (для определённого
- 21. Трудности в оценке наследуемости Для родственников часто общими являются не только гены, но и факторы окружающей
- 22. Гетерогенность полигенных заболеваний Генетическая и клиническая гетерогенность Полигенные заболевания часто генетически гетерогенны, причем разные генетические факторы
- 23. Совместное действие генов при полигенных заболеваниях Почти каждая болезнь зависит от многих генов. Почти каждый ген
- 24. Другие возможные источники «потерянной наследуемости» Фенотип - продукт взаимодействий множества генов не только друг с другом,
- 25. Этапы развития методик работы с ДНК
- 26. Методики работы с ДНК 1.Выделение 2.Электрофорез 3.Рестрикция 4.Лигирование 5.Гибридизация 6.Полимеразная цепная реакция 7.Секвенирование 8.ДНК-микрочипы
- 27. Из какого материала приходится выделять ДНК и РНК при проведении биомедицинских исследований? Клеточные«суспензии» Ликвор, плазма крови,смывы
- 28. Из какого материала приходится выделять ДНК и РНК при проведении биомедицинских исследований? Структурированная ткань Замороженные образцы
- 29. Выход ДНК из разных источников
- 30. Стабильность нуклеиновых кислот при выделении из клеток В целом, РНК менее стабильная молекула, чем ДНК. Наличие
- 31. Экспресс-методы выделения нуклеиновых кислот Образец биоматериала инкубируется с лизирующим буфером при высокой температуре (порядка 90-95°С), в
- 32. Метод сорбции нуклеиновых кислот на силиконовом носителе В основе лежит избирательная сорбция НК в присутствии хаотропной
- 33. Метод фенол-хлороформной экстракции Принцип метода основан на том, что НК является полярной молекулой и не растворяется
- 34. Оценка качества и количества ДНК •Электрофорез При этом для более точного определения количества нуклеиновой кислоты используют
- 35. Дополнительная обработка образцов нуклеиновых кислот ферментами нуклеинового обмена Если нужно получить ДНК без примеси РНК, то
- 36. Дополнительная пробоподготовка: бисульфитная конверсия Перед проведением метилспецифичной ПЦР или бисульфитного секвенирования необходимо провести обработку ДНК бисульфитом
- 37. Дополнительная пробоподготовка: обратная транскрипция Как правило, полимеразы, используемые в ПЦР, нуждаются в матрице ДНК. Поэтому для
- 38. Обратная транскрипция с олиго-dТ-праймерами
- 39. Метод ПЦР
- 40. ПЦР –это качественный или количественный анализ? Если оценивать по конечной точке - качественный Nn=N02n,где: n–номер цикла
- 41. ПЦР –это качественный или количественный анализ? Если измерять концентрации продуктов в log-фазе -количественный Nn=N0Еn,тогда: N1n/N2n=(N10/N20)(E1/E2) –
- 42. Программируемые термоциклеры Что важно учитывать при выборе: 1. Формат (стандартные 0,2 мл, или 0,1 мл, или
- 43. Мультиплексная ПЦР 1. При увеличении количества локусов в мультиплексной ПЦР, практически, экспоненциально растут затраты времени на
- 44. Аллель-специфичная ПЦР Основная проблема АС-ПЦР – это неспецифический синтез с частично комплементарного праймера. Решения: вводить неспаренные
- 45. Тетра-праймерная ПЦР (ARMS-PCR - amplification refractory mutation system polymerase chain reaction)
- 46. Способы визуализации ПЦР «по конечной точке» Детекция по конечной точке: 1.FLASH 2.Электрофорез в геле
- 47. Электрофорез в агарозном геле Гель формируется за счет нековалентных связей. Имеет относительно низкую разрешающую способность. Раньше
- 48. Электрофорез в полиакриламидном геле Большая разрешающая способность. Часто используют в молекулярно-генетической диагностике. Красить лучше нитратом серебра
- 49. Гетеродуплексный анализ Метод основан на том, что при ренатурации смеси нормальных и мутантных аллелей наряду с
- 50. SSCP-анализ SSCP–Single Strand Conformation Polymorphism. Метод основывается на различной подвижности однонитиевых ДНК-структур в неденатурирующем полиакриламидном геле,
- 51. Эндонуклеазы рестрикции класса II (рестриктазы) Узнают специфические последовательности – сайты рестрикции Активны в виде димеров в
- 52. Изомеры рестриктаз Изошизомеры Рестриктазы разных видов бактерий, узнающие одинаковые сайты рестрикции и одинаково их расщепляющие. Метилчувствительные
- 53. Формы разрывов ДНК, образующихся под действием рестриктаз класса II «Тупые» концы 5’-выступающие 3’-выступающие
- 54. ПЦР-ПДРФ и COBRA ПДРФ-анализ (RFLP -Restriction Fragment Length Polymorphism) Combined Bisulfite Restriction Analysis (or COBRA)
- 55. Эмульсионная ПЦР Фрагменты ДНК пришиваются к магнитным шарикам таким образом, чтобы на каждый шарик приходился только
- 56. Digital PCR Позволяет определять абсолютное количество копий ДНК-мишени в образце. Обладает высокой чувствительностью и специфичностью. Проводится
- 58. Скачать презентацию
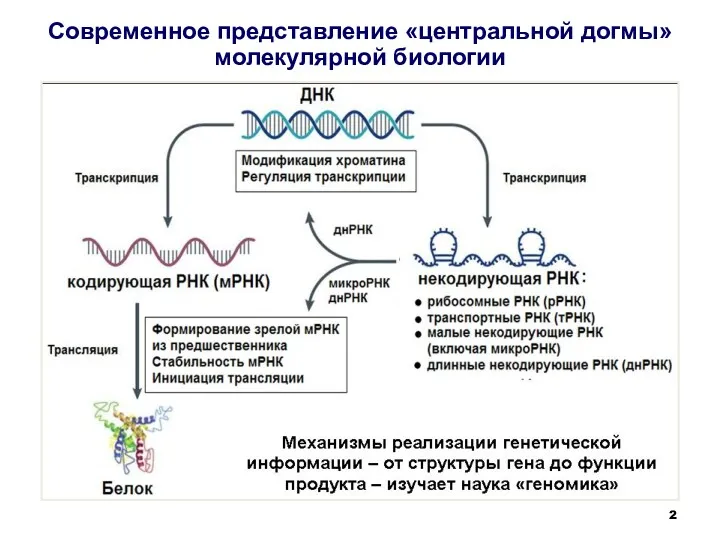
Современное представление «центральной догмы» молекулярной биологии
Современное представление «центральной догмы» молекулярной биологии
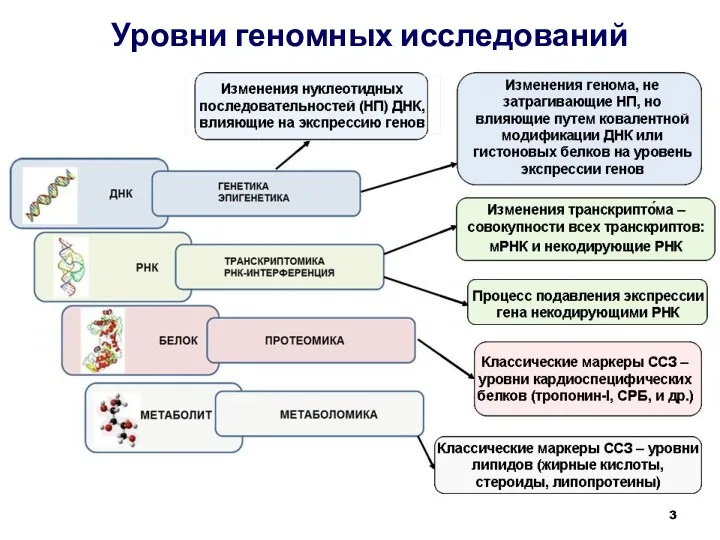
Уровни геномных исследований
Уровни геномных исследований

Предрасположенность человека к тем или иным заболеваниям генетически детерминирована.
Выделяют собственно наследственные
Предрасположенность человека к тем или иным заболеваниям генетически детерминирована.
Выделяют собственно наследственные

Моногенные и полигенные заболевания
Моногенные заболевания
Мутации могут захватывать один или оба
Моногенные и полигенные заболевания
Моногенные заболевания
Мутации могут захватывать один или оба
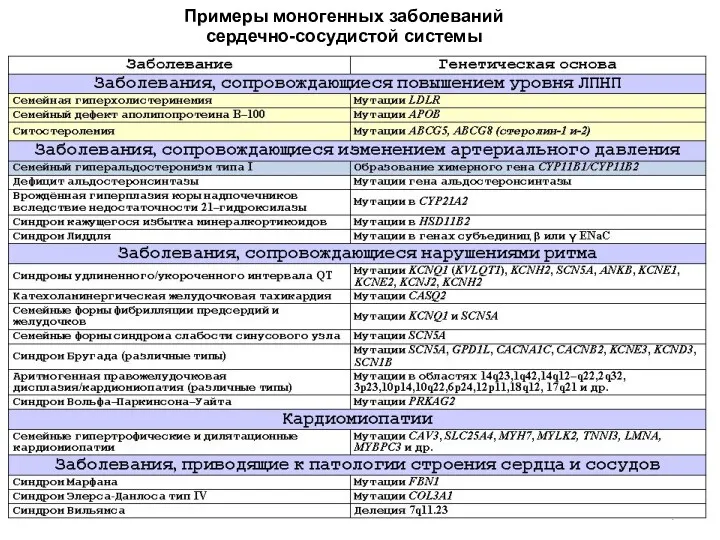
Примеры моногенных заболеваний
сердечно-сосудистой системы
Примеры моногенных заболеваний
сердечно-сосудистой системы

Распространенность полигенных заболеваний
Частота наследственных заболеваний составляет 1,5%.
На долю хромосомных болезней
Распространенность полигенных заболеваний
Частота наследственных заболеваний составляет 1,5%.
На долю хромосомных болезней

Генетический полиморфизм:
Генетический полиморфизм - это сосуществование в пределах популяции двух или
Генетический полиморфизм:
Генетический полиморфизм - это сосуществование в пределах популяции двух или

Соотношение Харди — Вайнберга
В популяции частоты генов и генотипов остаются постоянными от
Соотношение Харди — Вайнберга
В популяции частоты генов и генотипов остаются постоянными от

Мутация или полиморфизм?
Мутация - наследуемое изменение в нуклеотидной последовательности нуклеиновой кислоты,
Мутация или полиморфизм?
Мутация - наследуемое изменение в нуклеотидной последовательности нуклеиновой кислоты,

Причины генетического полиморфизма
Генетический полиморфизм может быть обусловлен:
заменой нуклеотидов,
дупликацией,
вставками,
Причины генетического полиморфизма
Генетический полиморфизм может быть обусловлен:
заменой нуклеотидов,
дупликацией,
вставками,
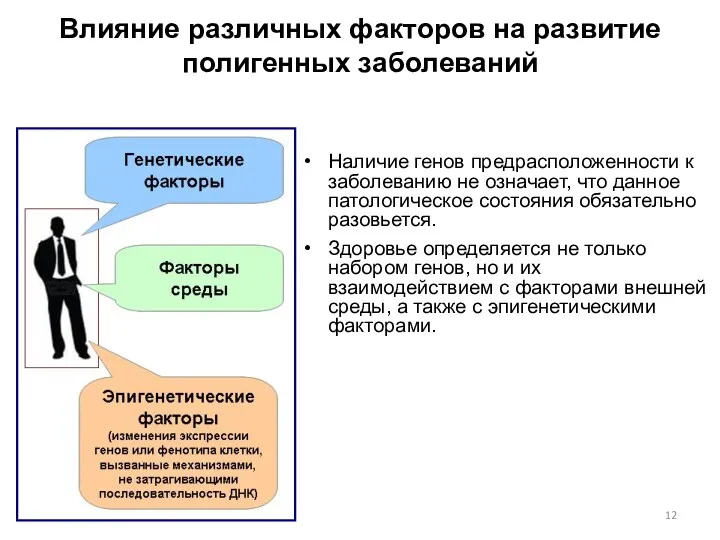
Влияние различных факторов на развитие полигенных заболеваний
Наличие генов предрасположенности к заболеванию
Влияние различных факторов на развитие полигенных заболеваний
Наличие генов предрасположенности к заболеванию

Участие факторов внешней среды в развитии полигенных заболеваний
Факторы внешней среды делятся
Участие факторов внешней среды в развитии полигенных заболеваний
Факторы внешней среды делятся

Основные факторы риска сердечно-сосудистых заболеваний
Немодифицируемые
наследственность
пол и возраст
мужчины старше 45 лет,
Основные факторы риска сердечно-сосудистых заболеваний
Немодифицируемые
наследственность
пол и возраст
мужчины старше 45 лет,

Стратегические этапы исследования генетической предрасположенности к полигенным заболеваниям
Анализ ассоциации заболевания
Стратегические этапы исследования генетической предрасположенности к полигенным заболеваниям
Анализ ассоциации заболевания
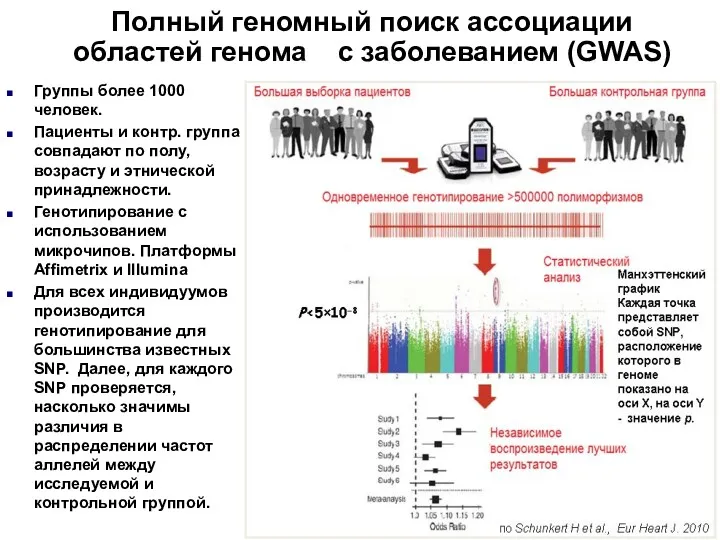
Полный геномный поиск ассоциации областей генома с заболеванием (GWAS)
Группы более 1000
Полный геномный поиск ассоциации областей генома с заболеванием (GWAS)
Группы более 1000

синим обозначены маркеры генетической предрасположенности к инфаркту миокарда,
оранжевым – к
синим обозначены маркеры генетической предрасположенности к инфаркту миокарда,
оранжевым – к

Локусы, ассоциированные с ишемической болезнью сердца и с инфарктом миокарда
Идентифицировано более
Локусы, ассоциированные с ишемической болезнью сердца и с инфарктом миокарда
Идентифицировано более

Проблема «потерянной» наследуемости»
Проект «Геном человека» (The Human Genome Project, HGP) начался в
Проблема «потерянной» наследуемости»
Проект «Геном человека» (The Human Genome Project, HGP) начался в
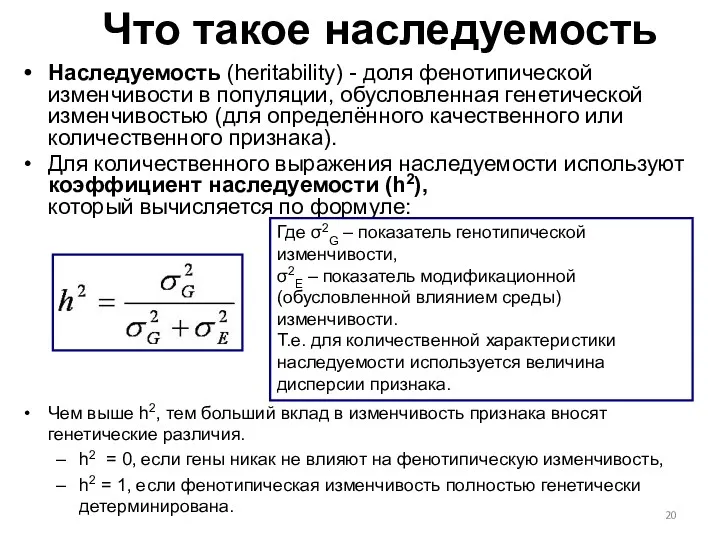
Что такое наследуемость
Наследуемость (heritability) - доля фенотипической изменчивости в популяции, обусловленная
Что такое наследуемость
Наследуемость (heritability) - доля фенотипической изменчивости в популяции, обусловленная

Трудности в оценке наследуемости
Для родственников часто общими являются не только гены,
Трудности в оценке наследуемости
Для родственников часто общими являются не только гены,

Гетерогенность полигенных заболеваний
Генетическая и клиническая гетерогенность
Полигенные заболевания часто генетически гетерогенны,
Гетерогенность полигенных заболеваний
Генетическая и клиническая гетерогенность
Полигенные заболевания часто генетически гетерогенны,

Совместное действие генов при полигенных заболеваниях
Почти каждая болезнь зависит от многих
Совместное действие генов при полигенных заболеваниях
Почти каждая болезнь зависит от многих

Другие возможные источники «потерянной наследуемости»
Фенотип - продукт взаимодействий множества генов не
Другие возможные источники «потерянной наследуемости»
Фенотип - продукт взаимодействий множества генов не

Этапы развития методик работы с ДНК
Этапы развития методик работы с ДНК

Методики работы с ДНК
1.Выделение
2.Электрофорез
3.Рестрикция
4.Лигирование
5.Гибридизация
6.Полимеразная цепная реакция
7.Секвенирование
8.ДНК-микрочипы
Методики работы с ДНК
1.Выделение
2.Электрофорез
3.Рестрикция
4.Лигирование
5.Гибридизация
6.Полимеразная цепная реакция
7.Секвенирование
8.ДНК-микрочипы

Из какого материала приходится выделять ДНК и РНК при проведении биомедицинских
Из какого материала приходится выделять ДНК и РНК при проведении биомедицинских

Из какого материала приходится выделять ДНК и РНК при проведении биомедицинских
Из какого материала приходится выделять ДНК и РНК при проведении биомедицинских

Выход ДНК из разных источников
Выход ДНК из разных источников
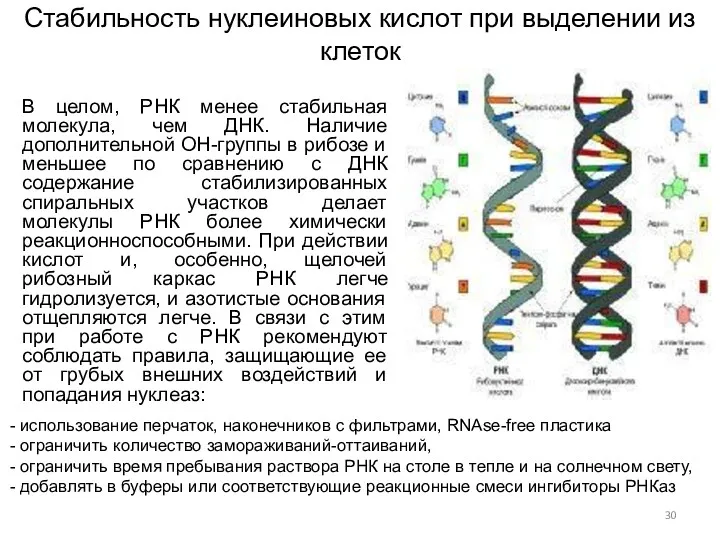
Стабильность нуклеиновых кислот при выделении из клеток
В целом, РНК менее стабильная
Стабильность нуклеиновых кислот при выделении из клеток
В целом, РНК менее стабильная

Экспресс-методы выделения нуклеиновых кислот
Образец биоматериала инкубируется с лизирующим буфером при высокой
Экспресс-методы выделения нуклеиновых кислот
Образец биоматериала инкубируется с лизирующим буфером при высокой

Метод сорбции нуклеиновых кислот на силиконовом носителе
В основе лежит избирательная сорбция
Метод сорбции нуклеиновых кислот на силиконовом носителе
В основе лежит избирательная сорбция
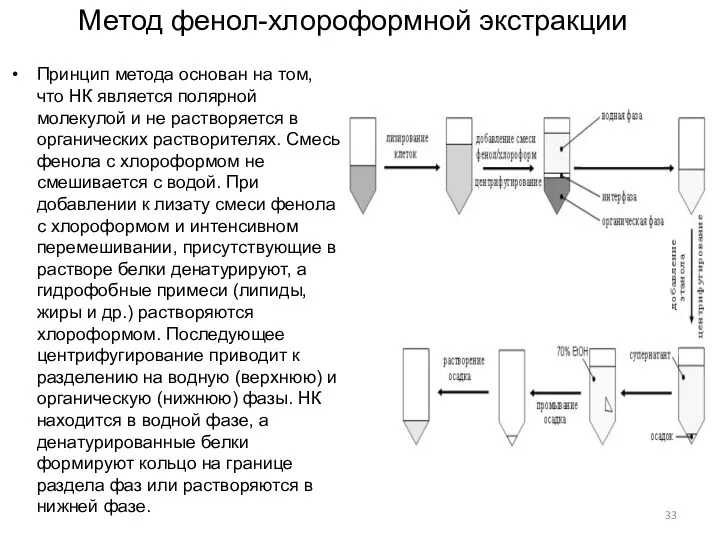
Метод фенол-хлороформной экстракции
Принцип метода основан на том, что НК является полярной
Метод фенол-хлороформной экстракции
Принцип метода основан на том, что НК является полярной
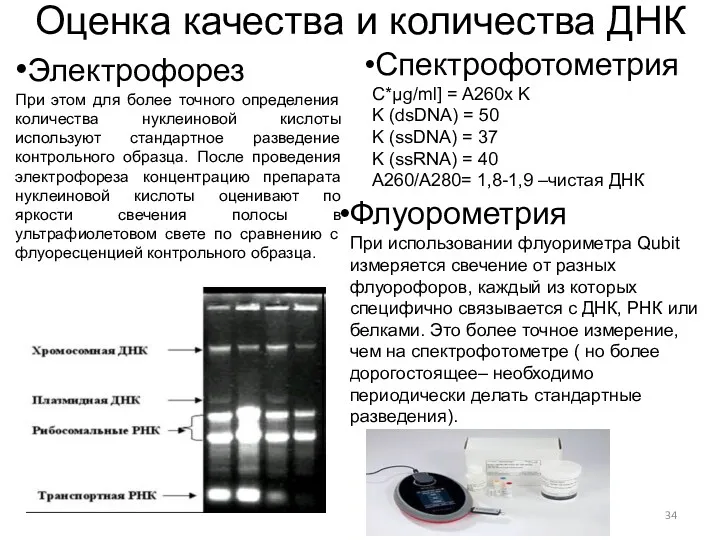
Оценка качества и количества ДНК
•Электрофорез
При этом для более точного определения количества
Оценка качества и количества ДНК
•Электрофорез
При этом для более точного определения количества

Дополнительная обработка образцов нуклеиновых кислот ферментами нуклеинового обмена
Если нужно получить ДНК
Дополнительная обработка образцов нуклеиновых кислот ферментами нуклеинового обмена
Если нужно получить ДНК
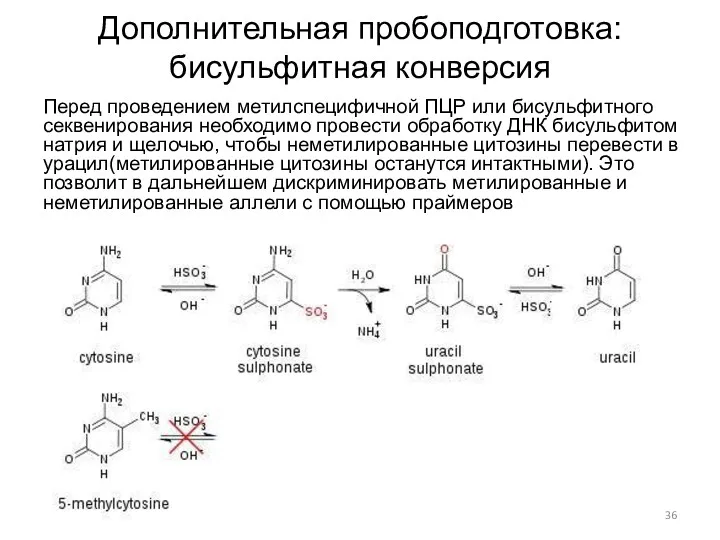
Дополнительная пробоподготовка: бисульфитная конверсия
Перед проведением метилспецифичной ПЦР или бисульфитного секвенирования необходимо
Дополнительная пробоподготовка: бисульфитная конверсия
Перед проведением метилспецифичной ПЦР или бисульфитного секвенирования необходимо
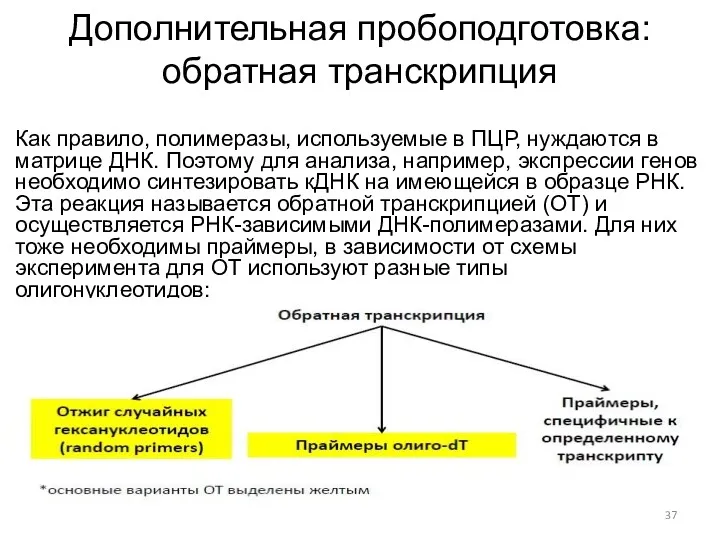
Дополнительная пробоподготовка: обратная транскрипция
Как правило, полимеразы, используемые в ПЦР, нуждаются в
Дополнительная пробоподготовка: обратная транскрипция
Как правило, полимеразы, используемые в ПЦР, нуждаются в
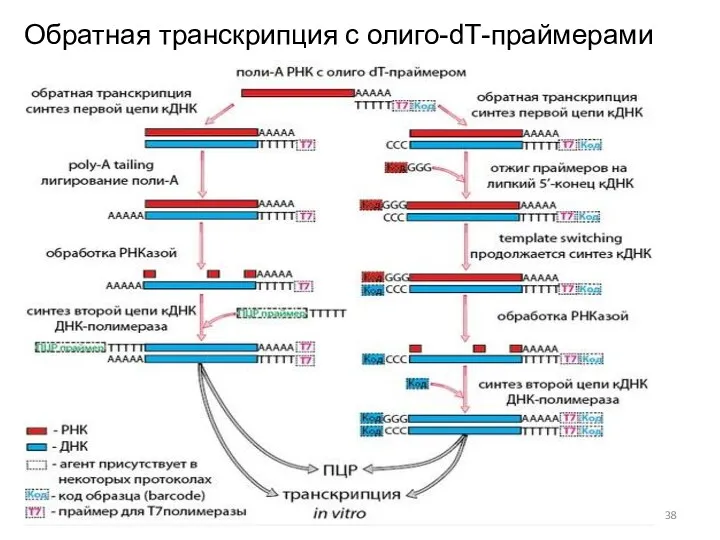
Обратная транскрипция с олиго-dТ-праймерами
Обратная транскрипция с олиго-dТ-праймерами
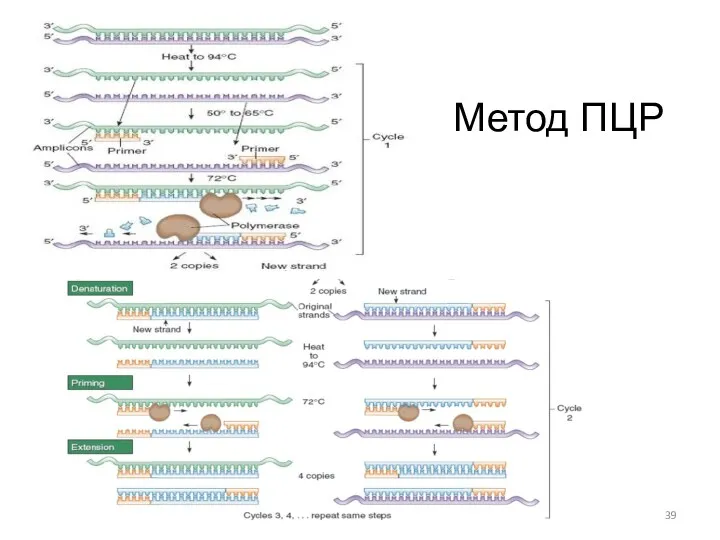
Метод ПЦР
Метод ПЦР
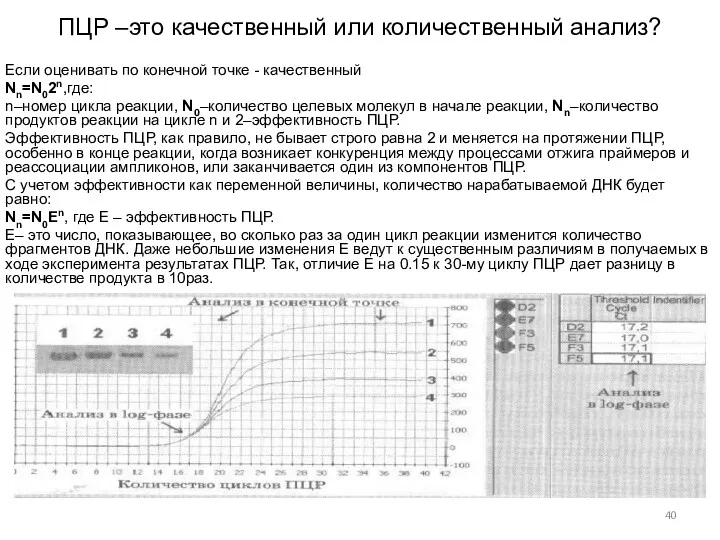
ПЦР –это качественный или количественный анализ?
Если оценивать по конечной точке -
ПЦР –это качественный или количественный анализ?
Если оценивать по конечной точке -
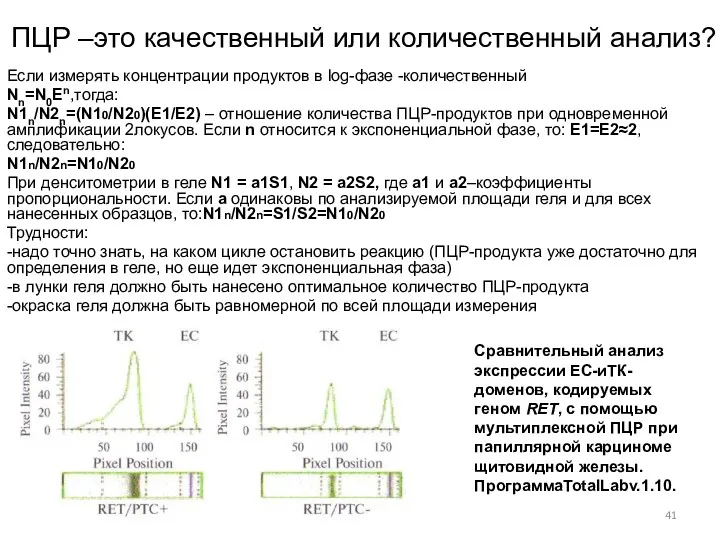
ПЦР –это качественный или количественный анализ?
Если измерять концентрации продуктов в log-фазе
ПЦР –это качественный или количественный анализ?
Если измерять концентрации продуктов в log-фазе

Программируемые термоциклеры
Что важно учитывать при выборе:
1. Формат (стандартные 0,2 мл, или
Программируемые термоциклеры
Что важно учитывать при выборе:
1. Формат (стандартные 0,2 мл, или

Мультиплексная ПЦР
1. При увеличении количества локусов в мультиплексной ПЦР, практически, экспоненциально
Мультиплексная ПЦР
1. При увеличении количества локусов в мультиплексной ПЦР, практически, экспоненциально
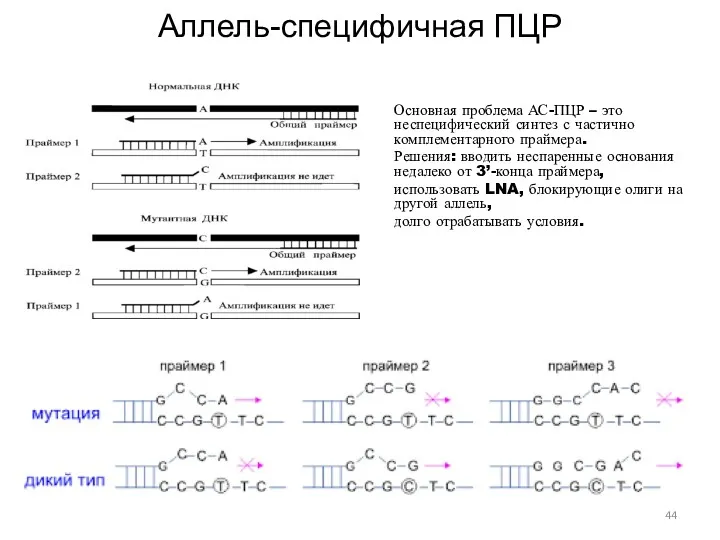
Аллель-специфичная ПЦР
Основная проблема АС-ПЦР – это неспецифический синтез с частично комплементарного
Аллель-специфичная ПЦР
Основная проблема АС-ПЦР – это неспецифический синтез с частично комплементарного
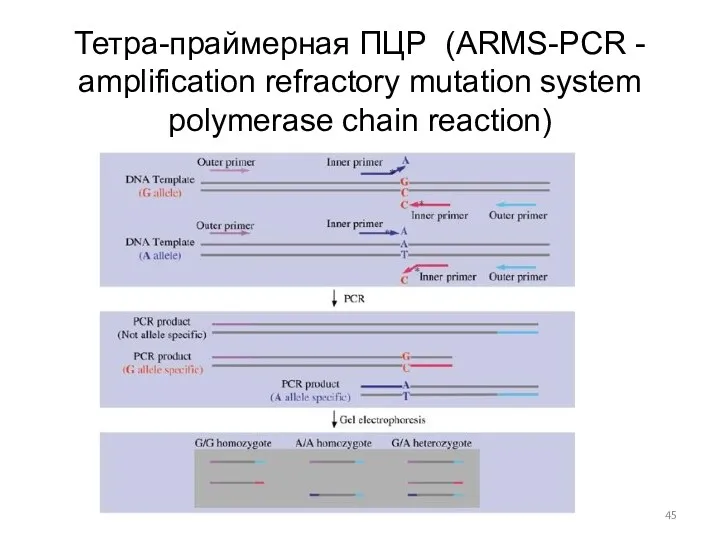
Тетра-праймерная ПЦР (ARMS-PCR - amplification refractory mutation system polymerase chain reaction)
Тетра-праймерная ПЦР (ARMS-PCR - amplification refractory mutation system polymerase chain reaction)

Способы визуализации ПЦР «по конечной точке»
Детекция по конечной точке:
1.FLASH
2.Электрофорез в геле
Способы визуализации ПЦР «по конечной точке»
Детекция по конечной точке:
1.FLASH
2.Электрофорез в геле

Электрофорез в агарозном геле
Гель формируется за счет нековалентных связей. Имеет относительно
Электрофорез в агарозном геле
Гель формируется за счет нековалентных связей. Имеет относительно
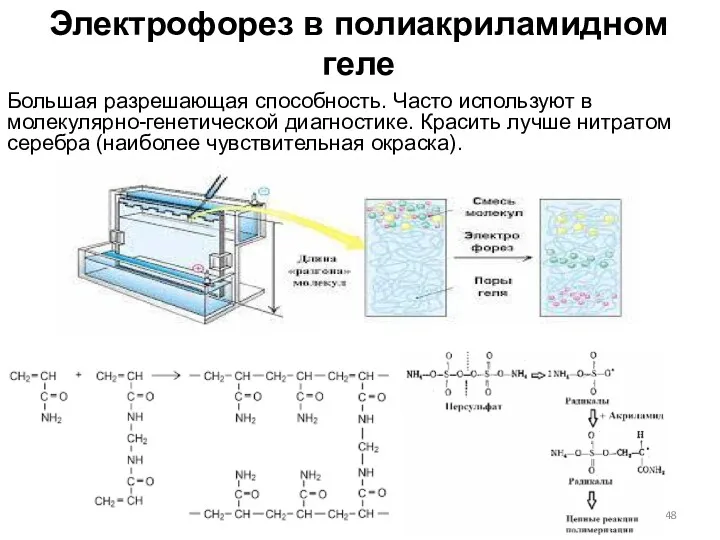
Электрофорез в полиакриламидном геле
Большая разрешающая способность. Часто используют в молекулярно-генетической диагностике.
Электрофорез в полиакриламидном геле
Большая разрешающая способность. Часто используют в молекулярно-генетической диагностике.

Гетеродуплексный анализ
Метод основан на том, что при ренатурации смеси нормальных и
Гетеродуплексный анализ
Метод основан на том, что при ренатурации смеси нормальных и

SSCP-анализ
SSCP–Single Strand Conformation Polymorphism.
Метод основывается на различной подвижности однонитиевых ДНК-структур
SSCP-анализ
SSCP–Single Strand Conformation Polymorphism.
Метод основывается на различной подвижности однонитиевых ДНК-структур

Эндонуклеазы рестрикции класса II (рестриктазы)
Узнают специфические последовательности – сайты рестрикции
Активны
Эндонуклеазы рестрикции класса II (рестриктазы)
Узнают специфические последовательности – сайты рестрикции
Активны

Изомеры рестриктаз
Изошизомеры
Рестриктазы разных видов бактерий, узнающие одинаковые сайты рестрикции и
Изомеры рестриктаз
Изошизомеры
Рестриктазы разных видов бактерий, узнающие одинаковые сайты рестрикции и
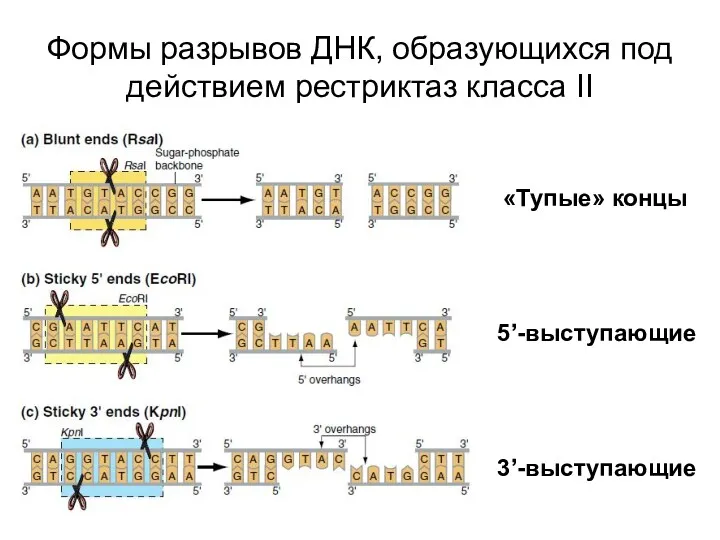
Формы разрывов ДНК, образующихся под действием рестриктаз класса II
«Тупые» концы
5’-выступающие
3’-выступающие
Формы разрывов ДНК, образующихся под действием рестриктаз класса II
«Тупые» концы
5’-выступающие
3’-выступающие
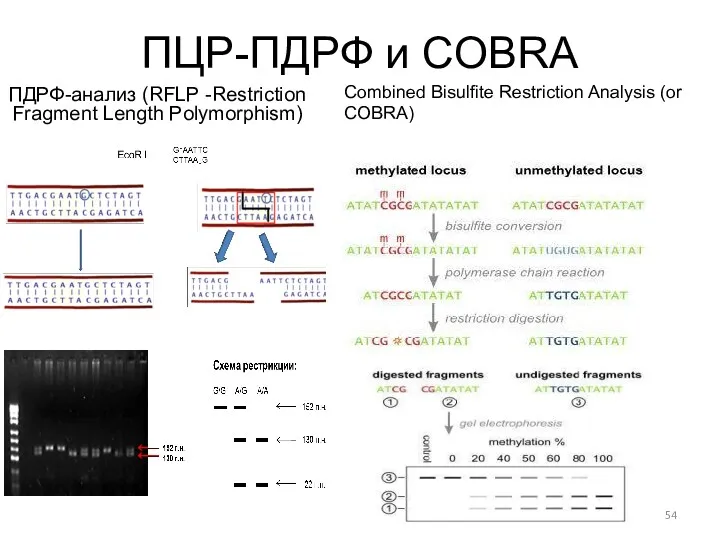
ПЦР-ПДРФ и COBRA
ПДРФ-анализ (RFLP -Restriction Fragment Length Polymorphism)
Combined Bisulfite Restriction Analysis
ПЦР-ПДРФ и COBRA
ПДРФ-анализ (RFLP -Restriction Fragment Length Polymorphism)
Combined Bisulfite Restriction Analysis
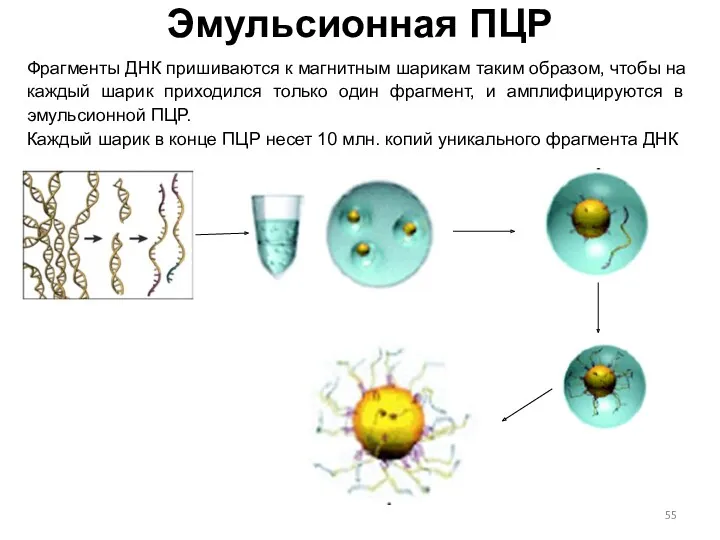
Эмульсионная ПЦР
Фрагменты ДНК пришиваются к магнитным шарикам таким образом, чтобы на
Эмульсионная ПЦР
Фрагменты ДНК пришиваются к магнитным шарикам таким образом, чтобы на
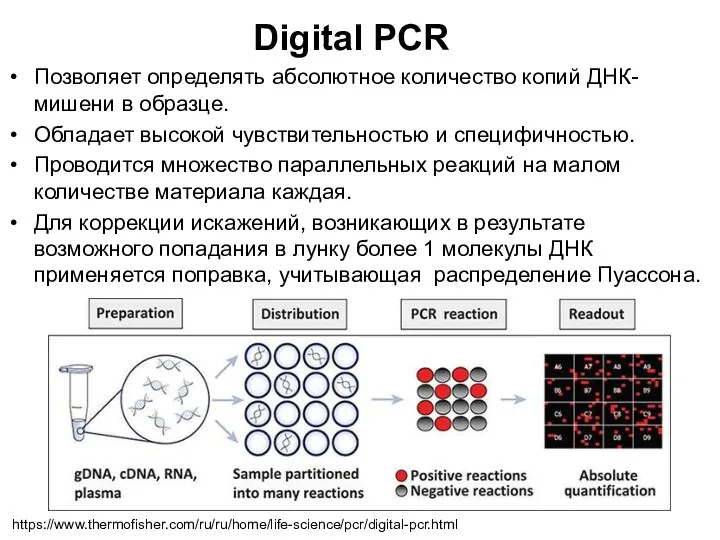
Digital PCR
Позволяет определять абсолютное количество копий ДНК-мишени в образце.
Обладает высокой чувствительностью
Digital PCR
Позволяет определять абсолютное количество копий ДНК-мишени в образце.
Обладает высокой чувствительностью
 Симпатический и парасимпатический подотделы вегетативной нервной системы, их строение и функции
Симпатический и парасимпатический подотделы вегетативной нервной системы, их строение и функции Пищеварение в кишечнике
Пищеварение в кишечнике Липидтер. Стероидтар. Терпеноидтар. Қорытынды. Пайдаланылған әдебиеттер
Липидтер. Стероидтар. Терпеноидтар. Қорытынды. Пайдаланылған әдебиеттер Коневодство. Чистокровная верховая порода
Коневодство. Чистокровная верховая порода Отряд черепахи
Отряд черепахи презентация на тему Плоды
презентация на тему Плоды Анализаторы и органы чувств. Органы осязания, обоняния и вкуса
Анализаторы и органы чувств. Органы осязания, обоняния и вкуса Почки растений
Почки растений Жизненный цикл клетки и его регуляция. Размножение организмов
Жизненный цикл клетки и его регуляция. Размножение организмов Экологическая экскурсия в Саблинские пещеры
Экологическая экскурсия в Саблинские пещеры Методическая разработка урока.
Методическая разработка урока. Происхождение и одомашнивание кошки
Происхождение и одомашнивание кошки История развития биологии
История развития биологии Растения в домашней аптечке
Растения в домашней аптечке Анатомия и физиология органов речи
Анатомия и физиология органов речи Спинной мозг
Спинной мозг Класс листостебельные мхи – Bryopsida, или Musci
Класс листостебельные мхи – Bryopsida, или Musci Строение слухового анализатора и его сенсорные расстройства
Строение слухового анализатора и его сенсорные расстройства Разработка и обоснование системы удобрения в севооборотах ООО Журавушка
Разработка и обоснование системы удобрения в севооборотах ООО Журавушка Лимбическая система
Лимбическая система Анализаторы - 1часть
Анализаторы - 1часть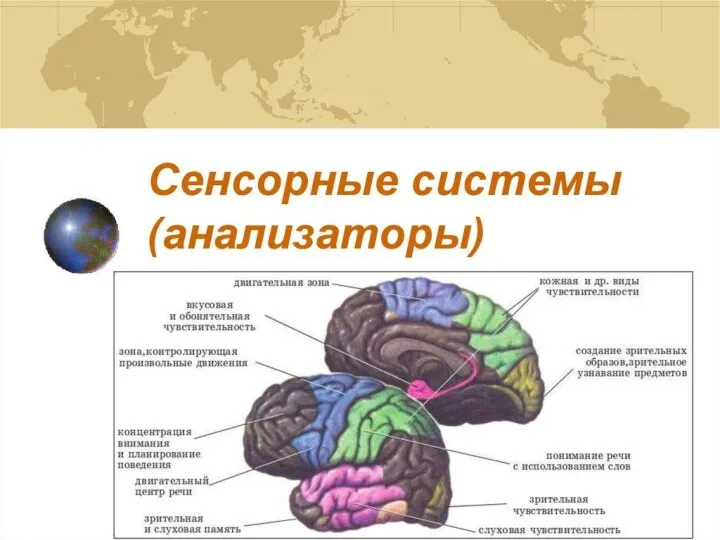 Сенсорные системы (анализаторы)
Сенсорные системы (анализаторы) Тип Членистоногие, класс Ракообразные
Тип Членистоногие, класс Ракообразные Теории эволюции жизни на Земле
Теории эволюции жизни на Земле Искусственное осеменение коров
Искусственное осеменение коров Хвоегрызущие и листогрызущие вредители. (Лекция 7)
Хвоегрызущие и листогрызущие вредители. (Лекция 7) Сон и сновидения
Сон и сновидения Возрастная физиология и психофизиология
Возрастная физиология и психофизиология