- Клетка как генетическая система

Содержание
- 2. Определение клетки Клетка – единая система, состоящая из множества закономерно связанных друг с другом элементов, представляющих
- 3. Определение клетки Клетки многоклеточных организмов тотипотентны, т.е. обладают генетическими потенциями всех клеток данного организма, равнозначны по
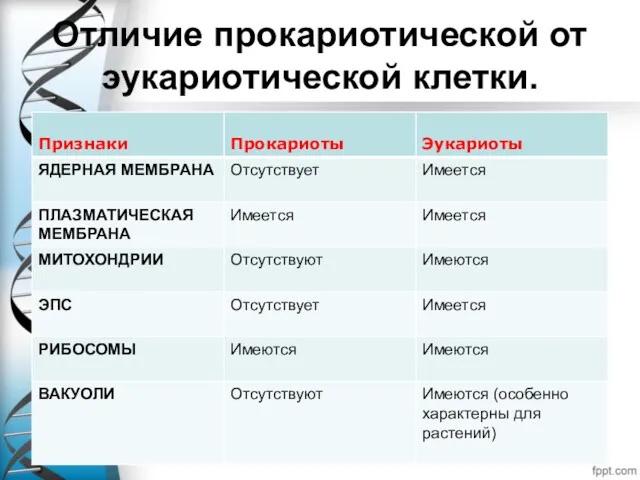
- 6. Отличие прокариотической от эукариотической клетки.
- 7. Отличие прокариотической от эукариотической клетки.
- 8. Цитоплазма Цитоплазма- это часть клетки, находящаяся между плазматической мембраной и ядром. В ней содержится от 60-90%
- 9. Ядро – важнейшая структура клетки. В нем сосредоточено 90% клеточной ДНК. Функция ядра-хранение наследственной информации. Кариоплазма
- 10. Ядро с ядрышком Ядро имеет форму шара с диаметром от 3-10 мкм. Оно окружено ядерной оболочкой,
- 11. Клеточный центр - Образован центриолями, центриоли расположены перпендикулярно друг другу, состоят из 9 триплетов микротрубочек (белок
- 12. Хромосомы -носители генетической информации Органоиды ядра эукариот, хромосома образована ДНК и молекулами белков – гистонов.
- 13. Плазматическая мембрана Липиды в мембране образуют двойной слой, а белки пронизывают всю ее толщу, погружены на
- 14. Функции мембраны: Отделяет клеточное содержимое от внешней среды. Регулирует обмен веществ между клеткой и средой. Протекают
- 16. Аппарат Гольджи – одномембранный органоид-10мкм,состоит сложенных стопкой, уплощенных, слегка изогнутых дискообразных цистерн. лизосомы мембрана лизосомы Функции
- 17. Митохондрии Имеют двумембранное строение. Внутренняя мембрана образует кристы. Митохондрия имеет свою ДНК, РНК. Функции митохондрий: Синтез
- 18. Пластиды-двумембранные органоиды растительных клеток. 1.хлоропласты 2.хромопласты 3.лейкопласты Структурные компоненты хлоропласта: 1-наружная мембрана 2- внутренняя мембрана 3-матрикс
- 19. Клеточный цикл Интерфаза. Деление ядра (митоз, мейоз).
- 20. Клеточный цикл Интерфа́за (англ. interphase) — период клеточного цикла, подразделяющаяся на G1-, S- и G2-фазы. Во
- 21. Клеточный цикл Мито́з (др.-греч. μίτος — нить) — непрямое деление клетки, наиболее распространённый способ репродукции эукариотических
- 22. Клеточный цикл Мейо́з (от др.-греч. μείωσις — уменьшение), или редукцио́нное деле́ние клетки — деление ядра эукариотической
- 23. Фазы митоза Фаза клеточного цикла, соответствующая делению клетки, называется М-фазой (от слова «митоз»). М-фазу условно подразделяют
- 24. Фазы митоза Длительность отдельных стадий различна и варьируется в зависимости от типа ткани, физиологического состояния организма,
- 25. Профаза
- 26. Профаза К основным событиям профазы относят конденсацию хромосом внутри ядра и образование веретена деления в цитоплазме
- 27. Профаза Конденсирующиеся сестринские хроматиды в ранней профазе остаются спаренными по всей своей длине с помощью белков-когезинов,
- 28. Профаза Наряду с процессами внутриядерной конденсации хромосом в цитоплазме начинает формироваться митотическое веретено — одна из
- 29. Профаза С началом формирования митотического веретена в профазе сопряжены разительные изменения динамических свойств микротрубочек. Время полужизни
- 30. Профаза Полимеризующиеся плюс-концы («+»-концы) являются «динамически нестабильными» и резко переходят от равномерного роста к быстрому укорочению,
- 31. Профаза Наряду с наблюдаемыми изменениями динамических свойств микротрубочек, слагающих нити веретена, в профазе закладываются полюса деления.
- 32. Профаза Параллельно конденсации хромосом и формированию митотического веретена, во время профазы происходит фрагментация эндоплазматического ретикулума, который
- 33. Метафаза
- 34. Метафаза В завершении прометафазы хромосомы располагаются в экваториальной плоскости веретена (а не всей клетки) примерно на
- 35. Метафаза Метафаза занимает значительную часть периода митоза, и отличается относительно стабильным состоянием. Все это время хромосомы
- 36. Метафаза В метафазе, также как и в течение других фаз митоза, продолжается активное обновление микротрубочек веретена
- 37. Анафаза
- 38. Анафаза Анафаза — самая короткая стадия митоза, которая начинается внезапным разделением и последующим расхождением сестринских хроматид
- 39. Анафаза Анафаза А характеризуется расхождением сестринских хроматид к противоположным полюсам деления клетки. За их движение при
- 40. Анафаза Вероятно, деполимеризация микротрубочек у кинетохоров либо в области полюсов деления является необходимым условием для перемещения
- 41. Анафаза Во время анафазы В расходятся сами полюса деления клетки, и, в отличие от анафазы А,
- 42. Анафаза Последовательность, продолжительность и относительный вклад каждого из двух процессов, слагающих анафазу, могут быть крайне различны.
- 43. Телофаза Телофаза (от греч. telos — конец) рассматривается как заключительная стадия митоза; за её начало принимается
- 44. Телофаза При этом отдельные пузырьки связываются с поверхностью хромосом и сливаются воедино. Постепенно восстанавливается наружная и
- 45. Телофаза Внутри заново сформировавшихся клеточных ядер хроматин переходит в дисперсное состояние, возобновляется синтез РНК, и становятся
- 46. Мейоз Профаза I — профаза первого деления очень сложная и состоит из 5 стадий: Метафаза I
- 47. Мейоз Профаза II — происходит конденсация хромосом, клеточный центр делится и продукты его деления расходятся к
- 49. Скачать презентацию

Определение клетки
Клетка – единая система, состоящая из множества закономерно связанных друг
Определение клетки
Клетка – единая система, состоящая из множества закономерно связанных друг

Определение клетки
Клетки многоклеточных организмов тотипотентны, т.е. обладают генетическими потенциями всех клеток
Определение клетки
Клетки многоклеточных организмов тотипотентны, т.е. обладают генетическими потенциями всех клеток
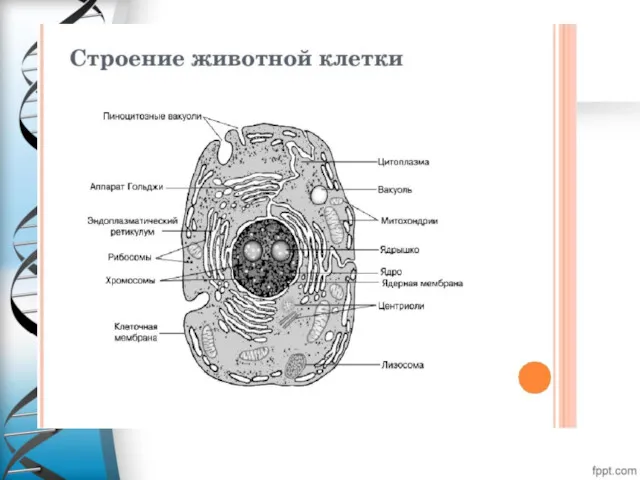
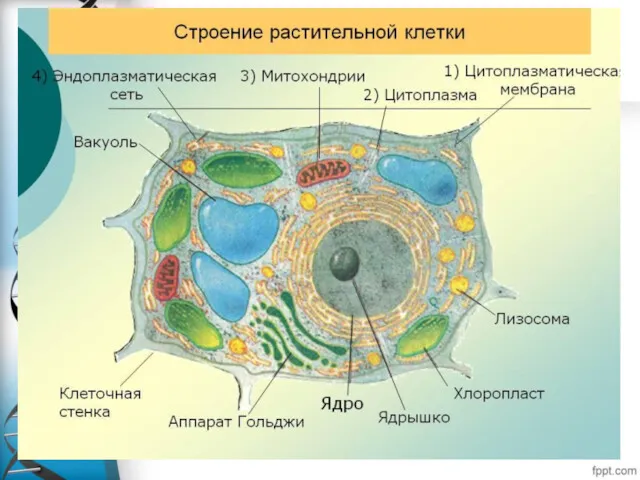
Отличие прокариотической от эукариотической клетки.
Отличие прокариотической от эукариотической клетки.
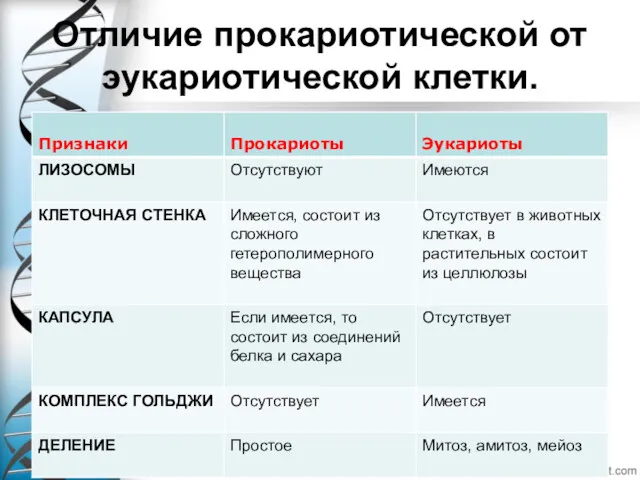
Отличие прокариотической от эукариотической клетки.
Отличие прокариотической от эукариотической клетки.

Цитоплазма
Цитоплазма- это часть клетки, находящаяся между плазматической мембраной и ядром.
Цитоплазма
Цитоплазма- это часть клетки, находящаяся между плазматической мембраной и ядром.
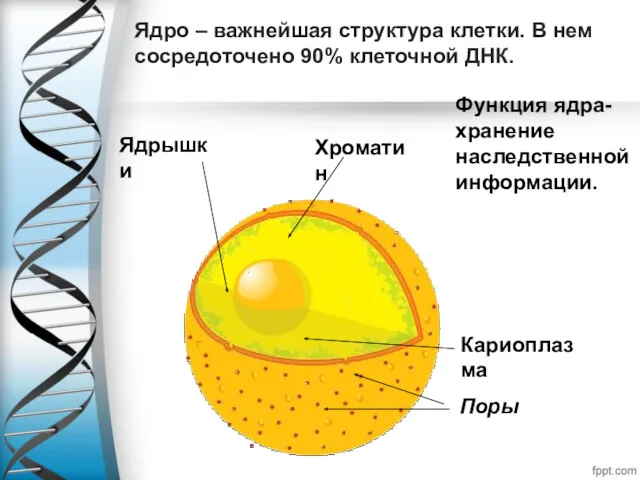
Ядро – важнейшая структура клетки. В нем сосредоточено 90% клеточной ДНК.
Функция
Ядро – важнейшая структура клетки. В нем сосредоточено 90% клеточной ДНК.
Функция

Ядро с ядрышком
Ядро имеет форму шара с диаметром от 3-10 мкм.
Ядро с ядрышком
Ядро имеет форму шара с диаметром от 3-10 мкм.
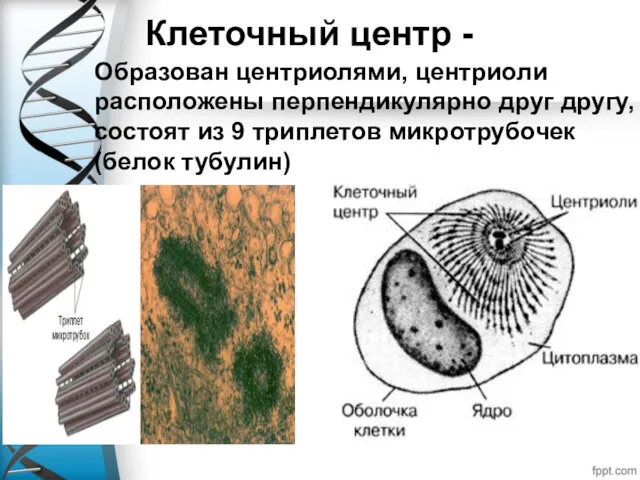
Клеточный центр -
Образован центриолями, центриоли расположены перпендикулярно друг другу, состоят
Клеточный центр -
Образован центриолями, центриоли расположены перпендикулярно друг другу, состоят
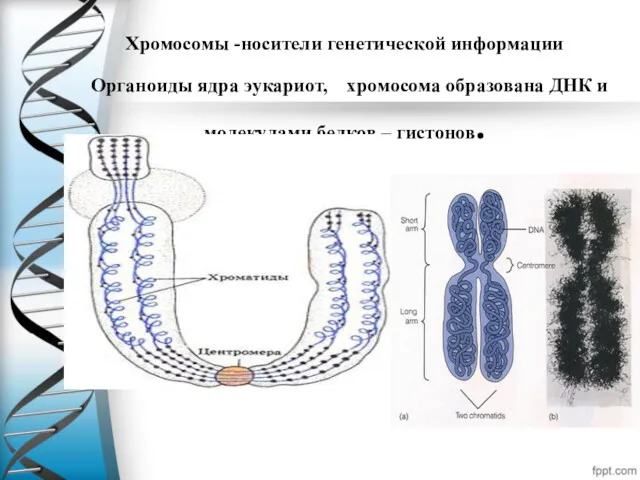
Хромосомы -носители генетической информации
Органоиды ядра эукариот, хромосома образована ДНК и
Хромосомы -носители генетической информации Органоиды ядра эукариот, хромосома образована ДНК и
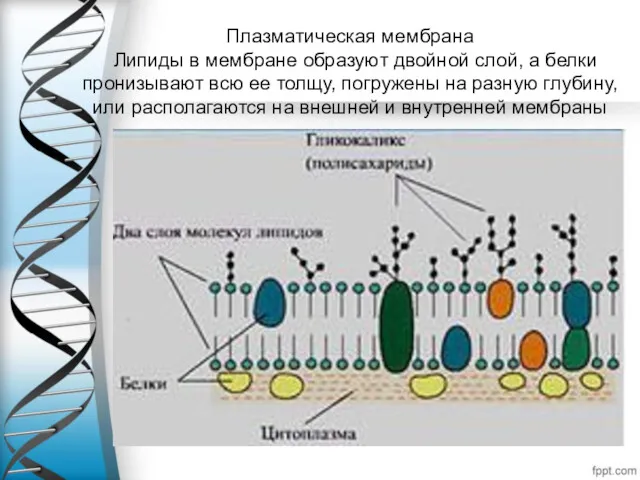
Плазматическая мембрана
Липиды в мембране образуют двойной слой, а белки пронизывают
Плазматическая мембрана Липиды в мембране образуют двойной слой, а белки пронизывают

Функции мембраны:
Отделяет клеточное содержимое от внешней среды.
Регулирует обмен веществ между клеткой
Функции мембраны:
Отделяет клеточное содержимое от внешней среды.
Регулирует обмен веществ между клеткой
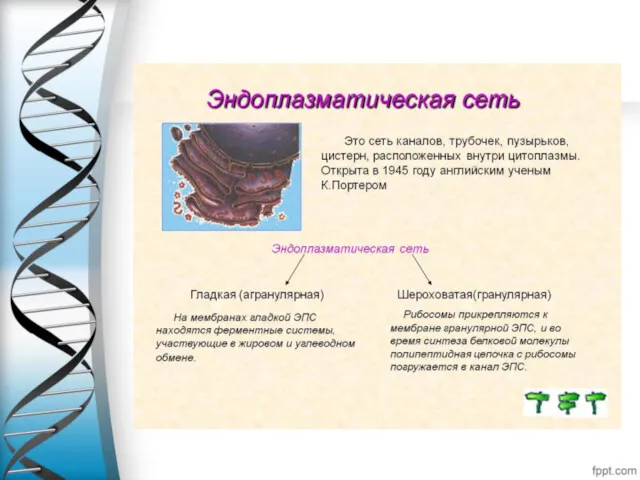
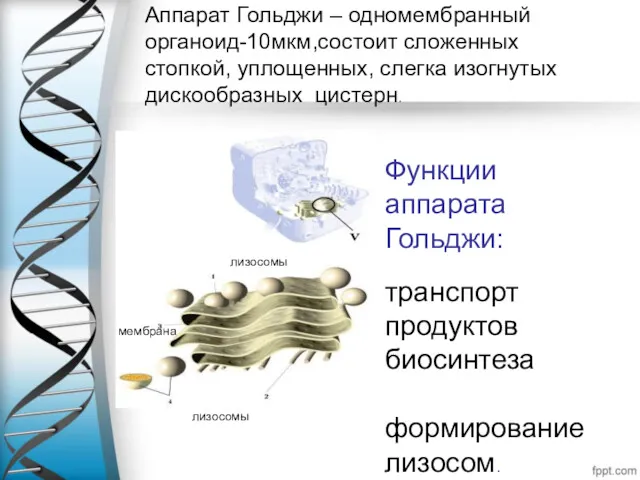
Аппарат Гольджи – одномембранный органоид-10мкм,состоит сложенных стопкой, уплощенных, слегка изогнутых дискообразных
Аппарат Гольджи – одномембранный органоид-10мкм,состоит сложенных стопкой, уплощенных, слегка изогнутых дискообразных
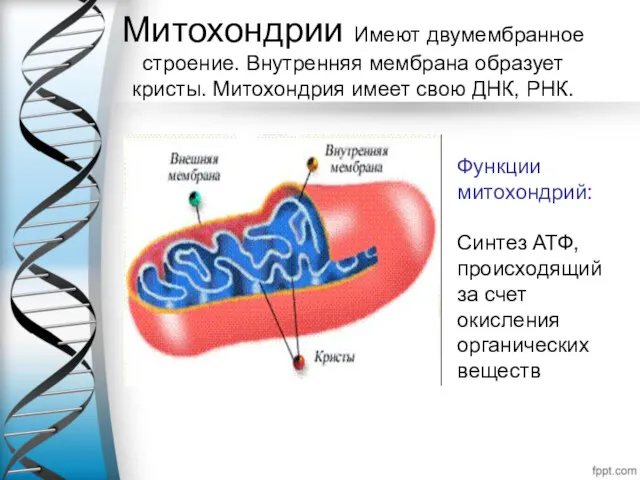
Митохондрии Имеют двумембранное строение. Внутренняя мембрана образует кристы. Митохондрия имеет свою
Митохондрии Имеют двумембранное строение. Внутренняя мембрана образует кристы. Митохондрия имеет свою

Пластиды-двумембранные органоиды растительных клеток.
1.хлоропласты
2.хромопласты
3.лейкопласты
Структурные компоненты
хлоропласта:
1-наружная мембрана
2- внутренняя мембрана
3-матрикс
4-граны
5-ламелла
6-тиллакойд
7-строма тиллакоида
Пластиды-двумембранные органоиды растительных клеток.
1.хлоропласты
2.хромопласты
3.лейкопласты
Структурные компоненты
хлоропласта:
1-наружная мембрана
2- внутренняя мембрана
3-матрикс
4-граны
5-ламелла
6-тиллакойд
7-строма тиллакоида

Клеточный цикл
Интерфаза.
Деление ядра (митоз, мейоз).
Клеточный цикл
Интерфаза.
Деление ядра (митоз, мейоз).

Клеточный цикл
Интерфа́за (англ. interphase) — период клеточного цикла, подразделяющаяся на G1-, S- и G2-фазы.
Клеточный цикл
Интерфа́за (англ. interphase) — период клеточного цикла, подразделяющаяся на G1-, S- и G2-фазы.

Клеточный цикл
Мито́з (др.-греч. μίτος — нить) — непрямое деление клетки, наиболее распространённый способ репродукции эукариотических клеток. Биологическое значение митоза
Клеточный цикл
Мито́з (др.-греч. μίτος — нить) — непрямое деление клетки, наиболее распространённый способ репродукции эукариотических клеток. Биологическое значение митоза

Клеточный цикл
Мейо́з (от др.-греч. μείωσις — уменьшение), или редукцио́нное деле́ние клетки — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит
Клеточный цикл
Мейо́з (от др.-греч. μείωσις — уменьшение), или редукцио́нное деле́ние клетки — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит

Фазы митоза
Фаза клеточного цикла, соответствующая делению клетки, называется М-фазой (от слова
Фазы митоза
Фаза клеточного цикла, соответствующая делению клетки, называется М-фазой (от слова

Фазы митоза
Длительность отдельных стадий различна и варьируется в зависимости от типа
Фазы митоза
Длительность отдельных стадий различна и варьируется в зависимости от типа
Профаза
Профаза

Профаза
К основным событиям профазы относят конденсацию хромосом внутри ядра и образование веретена
Профаза
К основным событиям профазы относят конденсацию хромосом внутри ядра и образование веретена

Профаза
Конденсирующиеся сестринские хроматиды в ранней профазе остаются спаренными по всей своей
Профаза
Конденсирующиеся сестринские хроматиды в ранней профазе остаются спаренными по всей своей

Профаза
Наряду с процессами внутриядерной конденсации хромосом в цитоплазме начинает формироваться митотическое
Профаза
Наряду с процессами внутриядерной конденсации хромосом в цитоплазме начинает формироваться митотическое

Профаза
С началом формирования митотического веретена в профазе сопряжены разительные изменения динамических
Профаза
С началом формирования митотического веретена в профазе сопряжены разительные изменения динамических

Профаза
Полимеризующиеся плюс-концы («+»-концы) являются «динамически нестабильными» и резко переходят от равномерного
Профаза
Полимеризующиеся плюс-концы («+»-концы) являются «динамически нестабильными» и резко переходят от равномерного

Профаза
Наряду с наблюдаемыми изменениями динамических свойств микротрубочек, слагающих нити веретена, в
Профаза
Наряду с наблюдаемыми изменениями динамических свойств микротрубочек, слагающих нити веретена, в

Профаза
Параллельно конденсации хромосом и формированию митотического веретена, во время профазы происходит
Профаза
Параллельно конденсации хромосом и формированию митотического веретена, во время профазы происходит
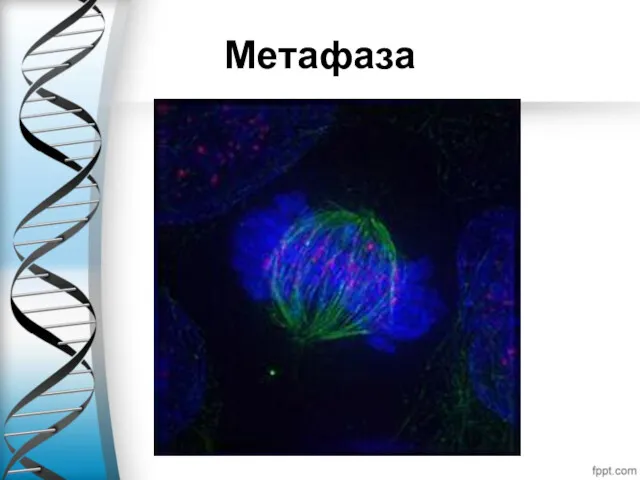
Метафаза
Метафаза

Метафаза
В завершении прометафазы хромосомы располагаются в экваториальной плоскости веретена (а не
Метафаза
В завершении прометафазы хромосомы располагаются в экваториальной плоскости веретена (а не

Метафаза
Метафаза занимает значительную часть периода митоза, и отличается относительно стабильным состоянием.
Метафаза
Метафаза занимает значительную часть периода митоза, и отличается относительно стабильным состоянием.

Метафаза
В метафазе, также как и в течение других фаз митоза, продолжается
Метафаза
В метафазе, также как и в течение других фаз митоза, продолжается
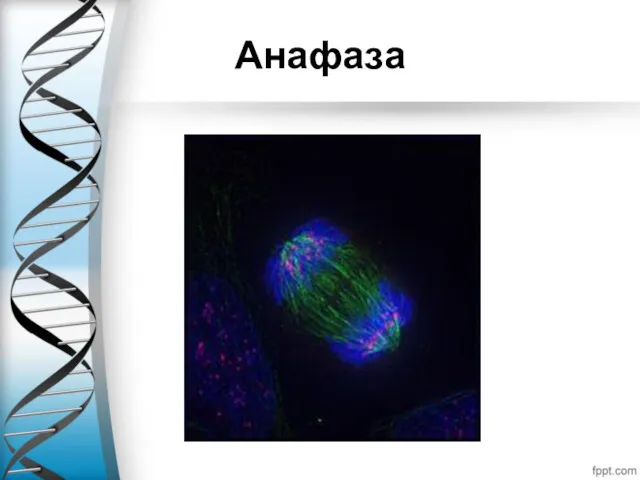
Анафаза
Анафаза

Анафаза
Анафаза — самая короткая стадия митоза, которая начинается внезапным разделением и последующим
Анафаза
Анафаза — самая короткая стадия митоза, которая начинается внезапным разделением и последующим

Анафаза
Анафаза А характеризуется расхождением сестринских хроматид к противоположным полюсам деления клетки. За
Анафаза
Анафаза А характеризуется расхождением сестринских хроматид к противоположным полюсам деления клетки. За

Анафаза
Вероятно, деполимеризация микротрубочек у кинетохоров либо в области полюсов деления является
Анафаза
Вероятно, деполимеризация микротрубочек у кинетохоров либо в области полюсов деления является

Анафаза
Во время анафазы В расходятся сами полюса деления клетки, и, в отличие
Анафаза
Во время анафазы В расходятся сами полюса деления клетки, и, в отличие

Анафаза
Последовательность, продолжительность и относительный вклад каждого из двух процессов, слагающих анафазу,
Анафаза
Последовательность, продолжительность и относительный вклад каждого из двух процессов, слагающих анафазу,

Телофаза
Телофаза (от греч. telos — конец) рассматривается как заключительная стадия митоза; за её
Телофаза
Телофаза (от греч. telos — конец) рассматривается как заключительная стадия митоза; за её

Телофаза
При этом отдельные пузырьки связываются с поверхностью хромосом и сливаются воедино.
Телофаза
При этом отдельные пузырьки связываются с поверхностью хромосом и сливаются воедино.

Телофаза
Внутри заново сформировавшихся клеточных ядер хроматин переходит в дисперсное состояние, возобновляется синтез РНК, и
Телофаза
Внутри заново сформировавшихся клеточных ядер хроматин переходит в дисперсное состояние, возобновляется синтез РНК, и

Мейоз
Профаза I — профаза первого деления очень сложная и состоит из 5
Мейоз
Профаза I — профаза первого деления очень сложная и состоит из 5

Мейоз
Профаза II — происходит конденсация хромосом, клеточный центр делится и продукты его
Мейоз
Профаза II — происходит конденсация хромосом, клеточный центр делится и продукты его
 Химизм фотосинтеза
Химизм фотосинтеза Ядовитые растения. Первая помощь, при отравлении ядовитыми растениями
Ядовитые растения. Первая помощь, при отравлении ядовитыми растениями Продление рода. Органы размножения. 7 класс
Продление рода. Органы размножения. 7 класс Органы и системы органов животных
Органы и системы органов животных Все самое интересное о кошках
Все самое интересное о кошках Вегетативное размножение растений
Вегетативное размножение растений Строение эукариотических клеток
Строение эукариотических клеток Воздушное питание растений. Фотосинтез
Воздушное питание растений. Фотосинтез Загальна будова та функції нервової системи
Загальна будова та функції нервової системи Фитоценоздағы өсімдіктердің ярустық жіктелуі
Фитоценоздағы өсімдіктердің ярустық жіктелуі Желтый Арбуз
Желтый Арбуз Цветы в легендах, поэзии, музыке
Цветы в легендах, поэзии, музыке Происхождение человека (теории происхождения человека)
Происхождение человека (теории происхождения человека) здоровьем дорожить умейте 3 часть
здоровьем дорожить умейте 3 часть Отряд пресмыкающиеся. Черепахи. Крокодилы
Отряд пресмыкающиеся. Черепахи. Крокодилы Обмен веществ и превращение энергии в клетке
Обмен веществ и превращение энергии в клетке Типи онтогенезу та його періодизація
Типи онтогенезу та його періодизація Размножение живых организмов
Размножение живых организмов Операции на голове
Операции на голове Дыхательная функция крови. Кислотно-основное состояние организма
Дыхательная функция крови. Кислотно-основное состояние организма Тип Гребневики (Ctenophora)
Тип Гребневики (Ctenophora) Занятие элективного курса Основы биологии для 11профильного класса.
Занятие элективного курса Основы биологии для 11профильного класса. Генна інженерія
Генна інженерія Репарация ДНК. Спонтанные мутации
Репарация ДНК. Спонтанные мутации Редуценты и их экологическая роль
Редуценты и их экологическая роль Вид и видообразование. (лекция 9-10)
Вид и видообразование. (лекция 9-10) Рысь. Внешний вид
Рысь. Внешний вид Викторина Эти забавные животные
Викторина Эти забавные животные