- Клеточный цикл и клеточная гибель

Содержание
- 2. Клеточный цикл - это промежуток времени от одного деления клетки до другого.
- 3. Фазы клеточного цикла Пресинтетическая(G1) Синтетическая(S) Постсинтетическая(G2) Митоз(M) Фаза покоя(G0)
- 4. Полиплоидия это клетки, в ядрах которых число хромосом кратно больше 2n. Клетки, содержащие наборы хромосом кратные
- 5. Политенные хромосомы -увеличенные в длину и ширину хромосомы в клетках некоторых тканей. Состоит такая хромосома из
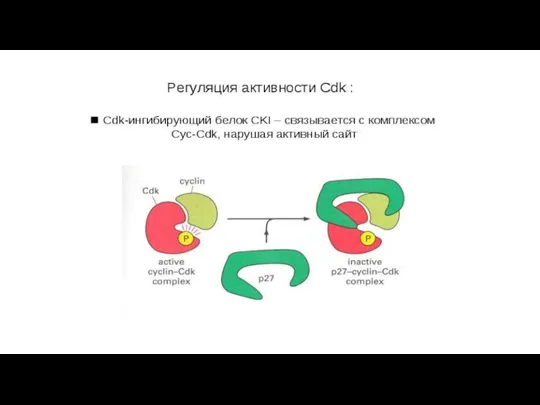
- 6. Регуляция клеточного цикла Осуществляется посредством обратимого фосфорилирования/дефосфорилирования регуляторных белков Ключевым белком, регулирующим вступление клетки в митоз,является
- 7. Пути регуляции клеточного цикла Экзогенные: 1.Ростовые факторы (стимулируют возвращение клетки из G0 обратно в цикл) Например,
- 9. Факторы роста Аутокринная стимуляция Паракринная стимуляция Гормональная стимуляция
- 10. Способы изучения клеточного цикла На крупных ооцитах шпорцевой лягушки(проводятся микроинъекции белков/химических препаратов) На почкующихся дрожжах Saccharomyces
- 11. Виды клеточной гибели Апоптотическая гибель- регулируемый процесс программируемой клеточной гибели. Некротическая гибель-это патологический процесс. Аутофагия- это
- 12. Апоптотическая гибель Одной из основных функций апоптоза является уничтожение дефектных клеток В многоклеточных организмах апоптоз к
- 13. Некротическая гибель Некроз проявляется в набухании, денатурации и коагуляции цитоплазматических белков, разрушении клеточных органелл и, наконец,
- 14. Аутофагия, ее типы Микроаутофагия- макромолекулы и обломки клеточных мембран просто захватываются лизосомой. Клетка может переваривать белки
- 15. Митотическая катастрофа Митотическая катастрофа принципиально отличается от апоптоза одноядерных клеток и аутофагической гибели тем, что нарушение
- 16. Механизмы запуска апоптоза Каспазный Некаспазный Рецепторый Митохондриальный Внутриклеточный Внешний
- 17. Каспазы Каспазы- цистеиновые протеазы, которые расщепляют белки по аспарагиновой кислоте - индуцирующие(активируют эффекторные каспазы) - эффекторные(активируют
- 18. Каспазный механизм Некоторые из прокаспаз, задействованных в апоптозе, запускают протеолитический каскад и называются инициаторными прокаспазами; Будучи
- 19. Некаспазный механизм Происходит выход из митохондрий и миграция в ядро флавопротеина AIF и эндонуклеазы G Как
- 20. Внешний и внутриклеточный механизмы Внешний механизм опосредован взаимодействием проапоптотических сигнальных молекул с рецепторами на плазматической мембране
- 21. Рецепторный механизм
- 22. Рецепторный механизм Fas-лиганд на поверхности лимфоцита-киллера активирует Fas-рецепторы смерти на поверхности клетки-мишени Цитоплазматическая часть Fas и
- 23. Митохондриальный механизм
- 24. Митохондриальный механизм Apaf-1 связывается с цитохромом с Apaf1 гидролизует связанный с ним dATP до dADP Замена
- 25. Более редкие механизмы Транскрипционный(активация гена p53, соответствующий белок р53 активируется при повреждении ДНК. Он останавливает клеточный
- 26. Ключевые белки В связи с какими-либо факторами, десфосфорилируется белок Bad, в следствии чего он связывается с
- 27. Проапоптотические белки BH123(основные- Bax и Bak), структурно подобные Bcl2, но без домена BH4 Белки, включающие только
- 28. Признаки апоптоза и некроза: их отличия
- 29. Значение программируемой клеточной гибели(на примере апоптоза) Апоптоз играет ключевую роль в ряде процессов развития организма, его
- 30. Использованная литература «Цитология» Ю.С.Ченцов 2010 «Молекулярная биология клетки» Брюс Альбертс 2013 «Клетки» Бенджамин Льюин 2011 http://www.rosoncoweb.ru/library/congress/ru/09/20.php
- 32. Скачать презентацию

Клеточный цикл - это промежуток времени от одного деления клетки до
Клеточный цикл - это промежуток времени от одного деления клетки до

Фазы клеточного цикла
Пресинтетическая(G1)
Синтетическая(S)
Постсинтетическая(G2)
Митоз(M)
Фаза покоя(G0)
Фазы клеточного цикла
Пресинтетическая(G1)
Синтетическая(S)
Постсинтетическая(G2)
Митоз(M)
Фаза покоя(G0)

Полиплоидия
это клетки, в ядрах которых число хромосом кратно больше 2n.
Клетки,
Полиплоидия
это клетки, в ядрах которых число хромосом кратно больше 2n. Клетки,

Политенные хромосомы
-увеличенные в длину и ширину хромосомы в клетках некоторых тканей.
Политенные хромосомы
-увеличенные в длину и ширину хромосомы в клетках некоторых тканей.

Регуляция клеточного цикла
Осуществляется посредством обратимого фосфорилирования/дефосфорилирования регуляторных белков
Ключевым белком, регулирующим
Регуляция клеточного цикла
Осуществляется посредством обратимого фосфорилирования/дефосфорилирования регуляторных белков
Ключевым белком, регулирующим

Пути регуляции клеточного цикла
Экзогенные:
1.Ростовые факторы (стимулируют возвращение клетки из G0
Пути регуляции клеточного цикла
Экзогенные:
1.Ростовые факторы (стимулируют возвращение клетки из G0

Факторы роста
Аутокринная стимуляция
Паракринная стимуляция
Гормональная стимуляция
Факторы роста
Аутокринная стимуляция
Паракринная стимуляция
Гормональная стимуляция

Способы изучения клеточного цикла
На крупных ооцитах шпорцевой лягушки(проводятся микроинъекции белков/химических препаратов)
На
Способы изучения клеточного цикла
На крупных ооцитах шпорцевой лягушки(проводятся микроинъекции белков/химических препаратов)
На

Виды клеточной гибели
Апоптотическая гибель- регулируемый процесс программируемой клеточной гибели.
Некротическая гибель-это патологический
Виды клеточной гибели
Апоптотическая гибель- регулируемый процесс программируемой клеточной гибели.
Некротическая гибель-это патологический

Апоптотическая гибель
Одной из основных функций апоптоза является уничтожение дефектных клеток
В
Апоптотическая гибель
Одной из основных функций апоптоза является уничтожение дефектных клеток
В

Некротическая гибель
Некроз проявляется в набухании, денатурации и коагуляции цитоплазматических белков, разрушении клеточных
Некротическая гибель
Некроз проявляется в набухании, денатурации и коагуляции цитоплазматических белков, разрушении клеточных

Аутофагия, ее типы
Микроаутофагия- макромолекулы и обломки клеточных мембран просто захватываются лизосомой.
Аутофагия, ее типы
Микроаутофагия- макромолекулы и обломки клеточных мембран просто захватываются лизосомой.

Митотическая катастрофа
Митотическая катастрофа принципиально отличается от апоптоза одноядерных клеток и аутофагической
Митотическая катастрофа
Митотическая катастрофа принципиально отличается от апоптоза одноядерных клеток и аутофагической

Механизмы запуска апоптоза
Каспазный
Некаспазный
Рецепторый
Митохондриальный
Внутриклеточный
Внешний
Механизмы запуска апоптоза
Каспазный
Некаспазный
Рецепторый
Митохондриальный
Внутриклеточный
Внешний

Каспазы
Каспазы- цистеиновые протеазы, которые расщепляют белки по аспарагиновой кислоте
- индуцирующие(активируют
Каспазы
Каспазы- цистеиновые протеазы, которые расщепляют белки по аспарагиновой кислоте
- индуцирующие(активируют

Каспазный механизм
Некоторые из прокаспаз, задействованных в апоптозе, запускают протеолитический каскад и
Каспазный механизм
Некоторые из прокаспаз, задействованных в апоптозе, запускают протеолитический каскад и

Некаспазный механизм
Происходит выход из митохондрий и миграция в ядро флавопротеина AIF
Некаспазный механизм
Происходит выход из митохондрий и миграция в ядро флавопротеина AIF

Внешний и внутриклеточный механизмы
Внешний механизм опосредован взаимодействием проапоптотических сигнальных молекул с
Внешний и внутриклеточный механизмы
Внешний механизм опосредован взаимодействием проапоптотических сигнальных молекул с
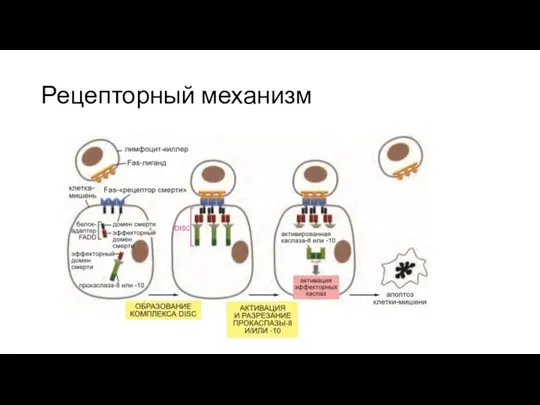
Рецепторный механизм
Рецепторный механизм

Рецепторный механизм
Fas-лиганд на поверхности лимфоцита-киллера активирует Fas-рецепторы смерти на поверхности клетки-мишени
Цитоплазматическая
Рецепторный механизм
Fas-лиганд на поверхности лимфоцита-киллера активирует Fas-рецепторы смерти на поверхности клетки-мишени
Цитоплазматическая
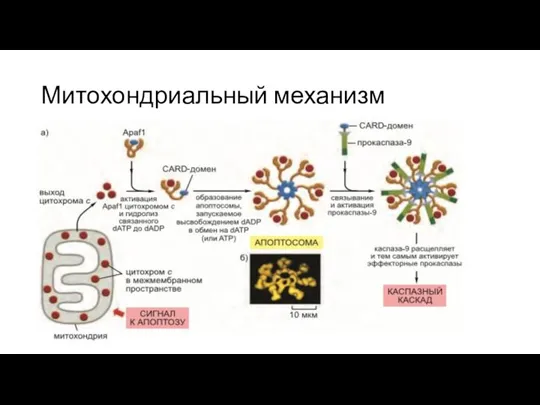
Митохондриальный механизм
Митохондриальный механизм

Митохондриальный механизм
Apaf-1 связывается с цитохромом с
Apaf1 гидролизует связанный с ним
Митохондриальный механизм
Apaf-1 связывается с цитохромом с
Apaf1 гидролизует связанный с ним

Более редкие механизмы
Транскрипционный(активация гена p53, соответствующий белок р53 активируется при повреждении
Более редкие механизмы
Транскрипционный(активация гена p53, соответствующий белок р53 активируется при повреждении

Ключевые белки
В связи с какими-либо факторами, десфосфорилируется белок Bad, в следствии
Ключевые белки
В связи с какими-либо факторами, десфосфорилируется белок Bad, в следствии

Проапоптотические белки
BH123(основные- Bax и Bak), структурно подобные Bcl2, но без домена
Проапоптотические белки
BH123(основные- Bax и Bak), структурно подобные Bcl2, но без домена
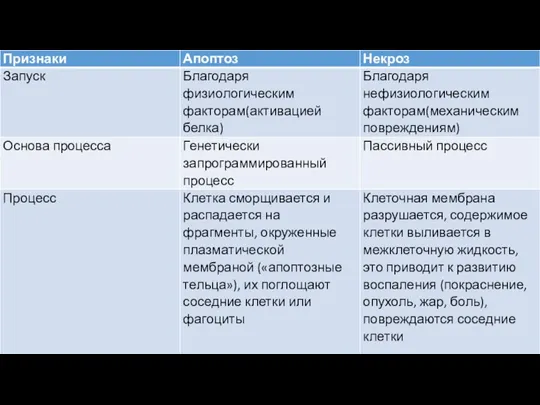
Признаки апоптоза и некроза: их отличия
Признаки апоптоза и некроза: их отличия

Значение программируемой клеточной гибели(на примере апоптоза)
Апоптоз играет ключевую роль в ряде
Значение программируемой клеточной гибели(на примере апоптоза)
Апоптоз играет ключевую роль в ряде

Использованная литература
«Цитология» Ю.С.Ченцов 2010
«Молекулярная биология клетки» Брюс Альбертс 2013
«Клетки» Бенджамин Льюин 2011
http://www.rosoncoweb.ru/library/congress/ru/09/20.php
Использованная литература
«Цитология» Ю.С.Ченцов 2010
«Молекулярная биология клетки» Брюс Альбертс 2013
«Клетки» Бенджамин Льюин 2011
http://www.rosoncoweb.ru/library/congress/ru/09/20.php
 Семейства Бобовые, Пасленовые, Сложноцветные
Семейства Бобовые, Пасленовые, Сложноцветные Микрофлора тела человека
Микрофлора тела человека Клетка
Клетка Женские половые органы
Женские половые органы Рослини-мандрівники. Тюльпан
Рослини-мандрівники. Тюльпан Общие вопросы анатомии и физиологии сердечно-сосудистой системы
Общие вопросы анатомии и физиологии сердечно-сосудистой системы Породы кошек. 1 класс
Породы кошек. 1 класс Домашние животные. (ДОУ)
Домашние животные. (ДОУ) Антропогенез. Этапы эволюции человека
Антропогенез. Этапы эволюции человека Водоросли (5 класс)
Водоросли (5 класс) Откуда берутся шоколад, изюм и мед
Откуда берутся шоколад, изюм и мед Чарлз Дарвин. Путешествие на Бигле
Чарлз Дарвин. Путешествие на Бигле Презентация по теме Анализаторы
Презентация по теме Анализаторы Уникальность человека. Биологическое и социальное в человеке
Уникальность человека. Биологическое и социальное в человеке Hereditary blood diseases and hereditary disease of endocrine system
Hereditary blood diseases and hereditary disease of endocrine system Божья коровка
Божья коровка Решение задач по генетике. Сцепленное с полом наследование. Половые хромосомы Х и У
Решение задач по генетике. Сцепленное с полом наследование. Половые хромосомы Х и У презентация к уроку анатомии
презентация к уроку анатомии Биохимические сдвиги в организме при мышечной работе
Биохимические сдвиги в организме при мышечной работе Дыхание. Значение дыхания в живых организмах
Дыхание. Значение дыхания в живых организмах Физические упражнения как средство профилактики болезней органов зрения
Физические упражнения как средство профилактики болезней органов зрения Кроссворд Газообмен
Кроссворд Газообмен Виды чеснока. Способы выращивания
Виды чеснока. Способы выращивания Регуляция пищеварения. Гигиена питания
Регуляция пищеварения. Гигиена питания Безусловные рефлексы
Безусловные рефлексы Один из методов решения генетических задач
Один из методов решения генетических задач Классификация животных. Основные систематические группы. Влияние человека на животных
Классификация животных. Основные систематические группы. Влияние человека на животных Физиология мышц и синапсов
Физиология мышц и синапсов