- Конечные продукты азотистого обмена. Биосинтез мочевины

Содержание
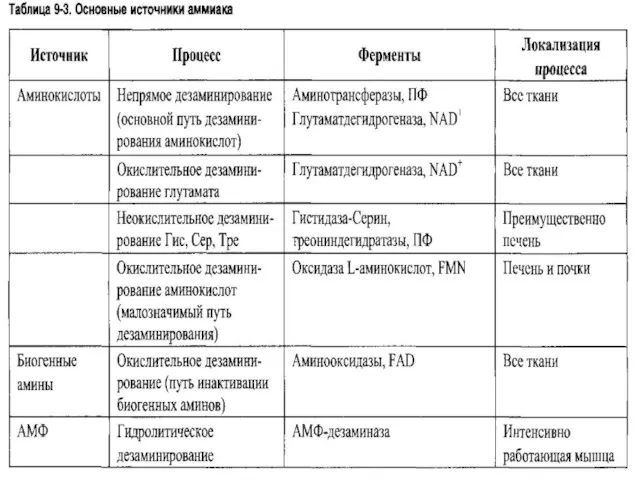
- 2. КОНЕЧНЫЕ ПРОДУКТЫ : АММИАК Деградация аминокислот происходит преимущественно в печени. При этом освобождается аммиак. Значительные количества
- 4. ТОКСИЧНОСТЬ АММИАКА Аммиак - NH3 является клеточным ядом. При высоких концентрациях он повреждает главным образом нервные
- 5. Токсичность аммиака В опытах на кроликах концентрация NH3 3 ммоль/л вызывала смерть! Причины токсичности: 1. при
- 6. Нейтр. мол. своб. NH3 легко проходят эти мембраны. При рН 7,4 только 1% NH3 от общего
- 7. Причины токсичности 2. NH3 + а-КГ + НАДФН2 -? Глу + НАДФ+ Н2О Отток альфа- КГ
- 8. Токсичность аммиака Аммиак настолько токсичен, что должен быть немедленно удален посредством экскреторного механизма, либо путем включения
- 9. Механизмы детоксикации аммиака 1. Синтез глутамина: Глн, аспарагина: Асн. 2. Синтез мочевины. 3. Аминирование а-КГ -->
- 10. Механизмы детоксикации аммиака 5.Синтез пурин. и пиримид. структур. 6. Нейтрализация в почках кислотами и выделение с
- 11. Восстановительное аминирование Большинство организмов обладает способностью реутилизировать аммиак за счет реакции, катализируемой глутаматдегидрогеназой. А-Кетоглутарат + NH3
- 12. Восстановительное аминирование
- 13. Биосинтез глутамина
- 14. Глутаминаза
- 17. Синтез аспарагина
- 18. МОЧЕВИНА У человека инактивация NH3 осуществляется за счет синтеза мочевины, часть NH3 выводится почками.
- 19. Синтез мочевины Мочевина - это нейтральное и нетоксичное соединение. Молекула мочевины может проходить через мембраны, из-за
- 20. СТАДИИ СИНТЕЗА МОЧЕВИНЫ Мочевина образуется в результате циклической последовательности реакций, протекающих в печени. Оба атома азота
- 21. Первая реакция На первой стадии из гидрокарбоната (НСО3-) и аммиака с потреблением 2 молекул АТФ образуется
- 22. Синтез карбамоилфосфата
- 23. Вторая стадия Карбамоильный остаток переносится на орнитин с образованием цитруллина. Вновь необходима энергия в форме АТФ,
- 24. Синтез цитруллина
- 25. Третья стадия Вторая аминогруппа молекулы мочевины поставляется за счет реакции аспартата с цитруллином.
- 26. Синтез аргининосукцината
- 27. Для обеспечения необратимости реакции дифосфат гидролизуется полностью. Отщепление фумарата от аргининосукцината приводит к аргинину,
- 28. Четвертая стадия
- 29. Пятая стадия из которого в результате гидролиза образуется мочевина. Остающийся орнитин вновь включается в цикл мочевины.
- 30. Пятая стадия
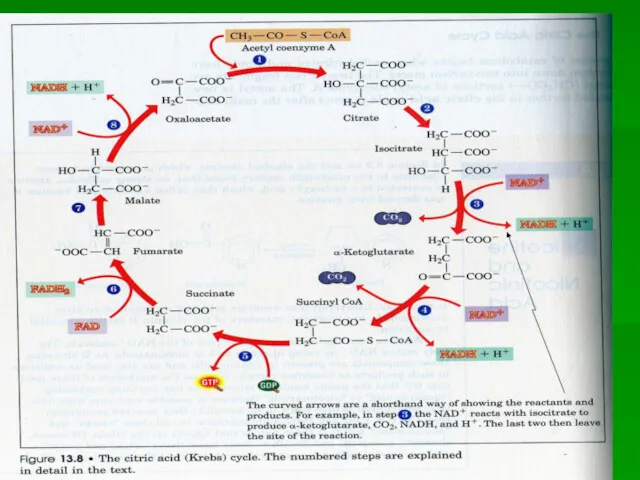
- 32. ВЕЛОСИПЕД КРЕБСА Фумарат, образующийся в цикле мочевины, может в результате двух стадий цитратного цикла [6, 7]
- 33. Регенерация аспартата
- 35. Взаимосвязь цикла мочевинообразования и ЦТК
- 36. ЭНЕРГОЗАВИСИМЫЙ ПРОЦЕСС Биосинтез мочевины требует больших затрат энергии. Энергия поставляется за счет расщепления четырех высокоэнергетических связей:
- 37. КОМПАРТМЕНТАЛИЗАЦИЯ Цикл мочевины протекает исключительно в печени. Он разделен на два компартмента: митохондрии и цитоплазму. Прохождение
- 38. АЛЛОСТЕРИЧЕСКАЯ РЕГУЛЯЦИЯ СИНТЕЗА МОЧЕВИНЫ Скорость синтеза мочевины определяется первой реакцией цикла. Карбамоилфосфатсинтаза активна только в присутствии
- 39. Наследственные нарушения орнитинового цикла и их симптомы
- 41. Включение безазотистого остатка АК в ЦТК
- 42. Пути биосинтеза заменимых АК
- 43. Глюкозо-аланиновый цикл
- 45. Скачать презентацию

КОНЕЧНЫЕ ПРОДУКТЫ : АММИАК
Деградация аминокислот происходит преимущественно в печени.
КОНЕЧНЫЕ ПРОДУКТЫ : АММИАК
Деградация аминокислот происходит преимущественно в печени.

ТОКСИЧНОСТЬ АММИАКА
Аммиак - NH3 является клеточным ядом. При высоких концентрациях он
ТОКСИЧНОСТЬ АММИАКА
Аммиак - NH3 является клеточным ядом. При высоких концентрациях он

Токсичность аммиака
В опытах на кроликах концентрация
NH3 3 ммоль/л вызывала
Токсичность аммиака
В опытах на кроликах концентрация
NH3 3 ммоль/л вызывала

Нейтр. мол. своб. NH3 легко проходят эти мембраны. При
Нейтр. мол. своб. NH3 легко проходят эти мембраны. При

Причины токсичности
2. NH3 + а-КГ + НАДФН2 -?
Глу + НАДФ+
Причины токсичности
2. NH3 + а-КГ + НАДФН2 -?
Глу + НАДФ+

Токсичность аммиака
Аммиак настолько токсичен, что должен быть немедленно удален посредством экскреторного
Токсичность аммиака
Аммиак настолько токсичен, что должен быть немедленно удален посредством экскреторного

Механизмы детоксикации аммиака
1. Синтез глутамина: Глн, аспарагина: Асн.
2. Синтез мочевины.
3. Аминирование
Механизмы детоксикации аммиака
1. Синтез глутамина: Глн, аспарагина: Асн.
2. Синтез мочевины.
3. Аминирование

Механизмы детоксикации аммиака
5.Синтез пурин. и пиримид. структур.
6. Нейтрализация в почках кислотами
Механизмы детоксикации аммиака
5.Синтез пурин. и пиримид. структур.
6. Нейтрализация в почках кислотами

Восстановительное аминирование
Большинство организмов обладает способностью реутилизировать аммиак за счет реакции, катализируемой
Восстановительное аминирование
Большинство организмов обладает способностью реутилизировать аммиак за счет реакции, катализируемой
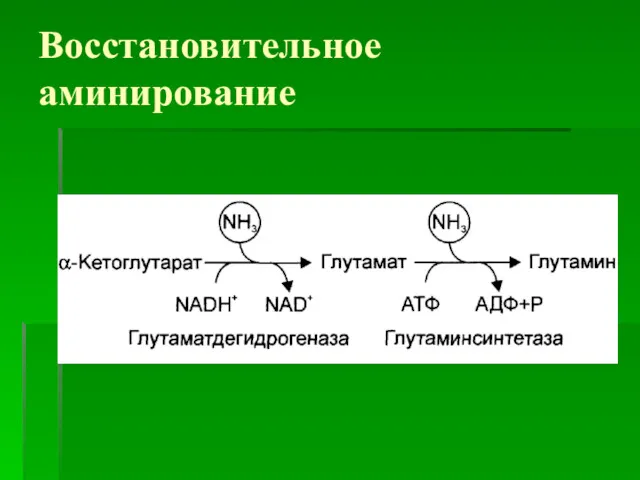
Восстановительное аминирование
Восстановительное аминирование
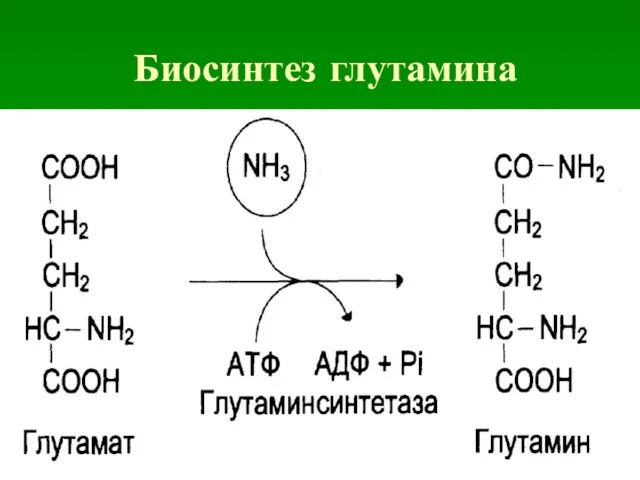
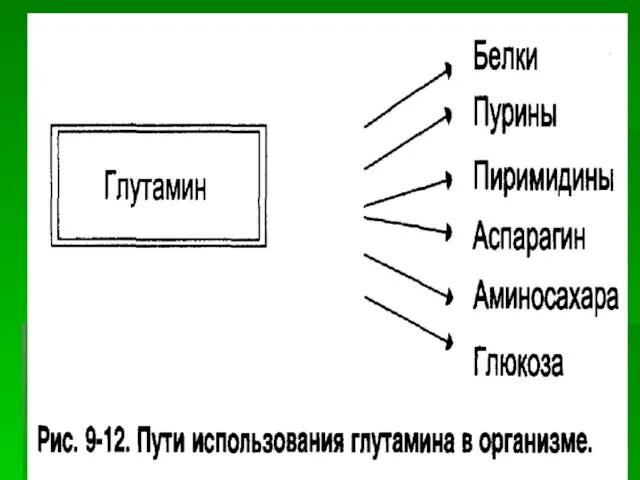
Биосинтез глутамина
Биосинтез глутамина
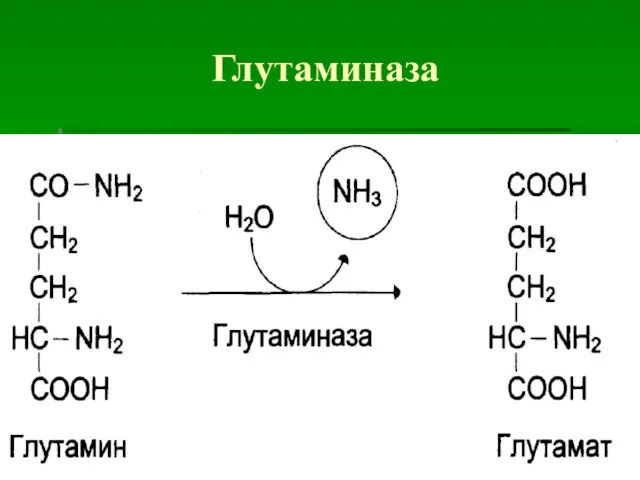
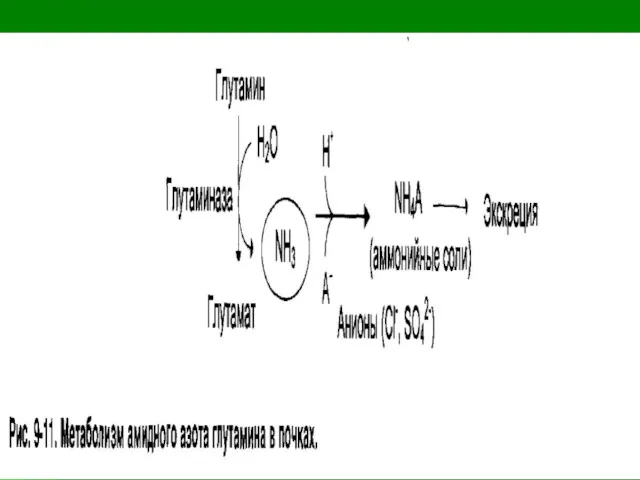
Глутаминаза
Глутаминаза
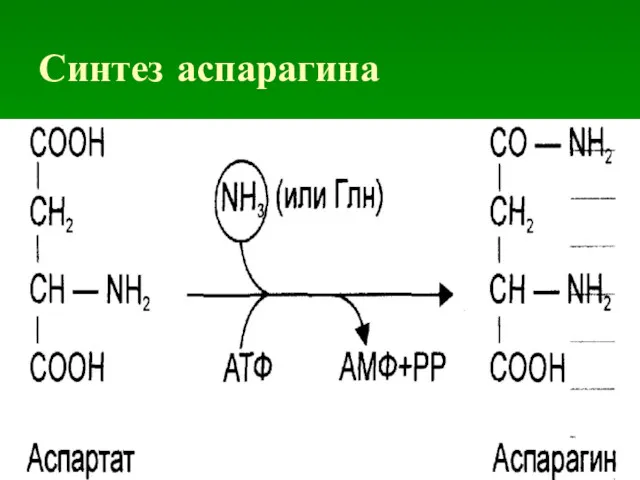
Синтез аспарагина
Синтез аспарагина

МОЧЕВИНА
У человека инактивация NH3 осуществляется за счет синтеза мочевины, часть
МОЧЕВИНА
У человека инактивация NH3 осуществляется за счет синтеза мочевины, часть

Синтез мочевины
Мочевина - это нейтральное и нетоксичное соединение. Молекула мочевины
Синтез мочевины
Мочевина - это нейтральное и нетоксичное соединение. Молекула мочевины

СТАДИИ СИНТЕЗА МОЧЕВИНЫ
Мочевина образуется в результате циклической последовательности реакций, протекающих в
СТАДИИ СИНТЕЗА МОЧЕВИНЫ
Мочевина образуется в результате циклической последовательности реакций, протекающих в

Первая реакция
На первой стадии из гидрокарбоната (НСО3-) и аммиака с потреблением
Первая реакция
На первой стадии из гидрокарбоната (НСО3-) и аммиака с потреблением
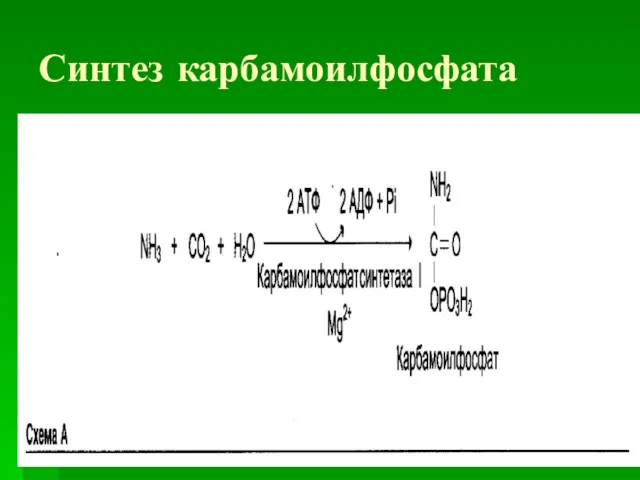
Синтез карбамоилфосфата
Синтез карбамоилфосфата

Вторая стадия
Карбамоильный остаток переносится на орнитин с образованием цитруллина. Вновь
Вторая стадия
Карбамоильный остаток переносится на орнитин с образованием цитруллина. Вновь
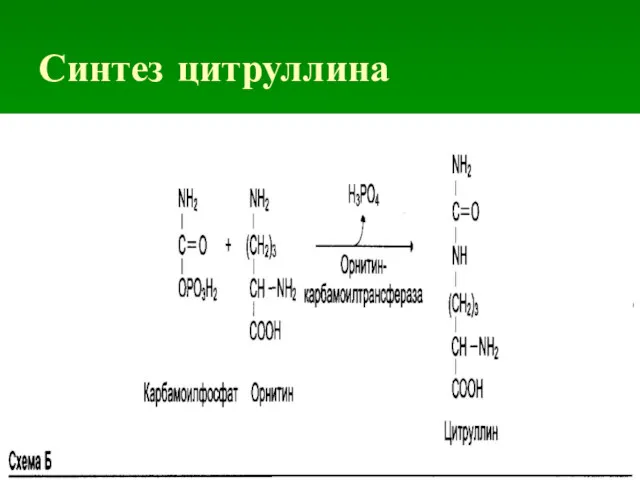
Синтез цитруллина
Синтез цитруллина

Третья стадия
Вторая аминогруппа молекулы мочевины поставляется за счет реакции аспартата с
Третья стадия
Вторая аминогруппа молекулы мочевины поставляется за счет реакции аспартата с

Синтез аргининосукцината
Синтез аргининосукцината

Для обеспечения необратимости реакции дифосфат гидролизуется полностью. Отщепление фумарата от
Для обеспечения необратимости реакции дифосфат гидролизуется полностью. Отщепление фумарата от

Четвертая стадия
Четвертая стадия

Пятая стадия
из которого в результате гидролиза образуется мочевина. Остающийся орнитин
Пятая стадия
из которого в результате гидролиза образуется мочевина. Остающийся орнитин
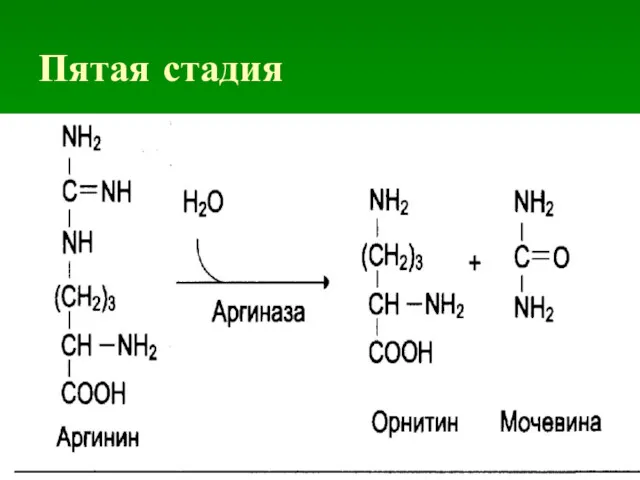
Пятая стадия
Пятая стадия
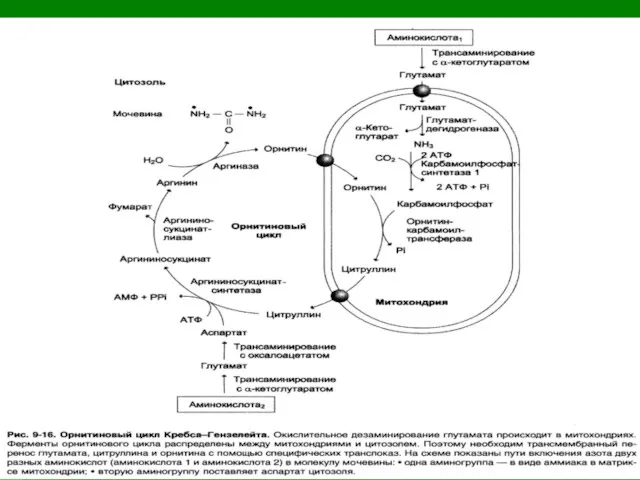

ВЕЛОСИПЕД КРЕБСА
Фумарат, образующийся в цикле мочевины, может в результате двух стадий
ВЕЛОСИПЕД КРЕБСА
Фумарат, образующийся в цикле мочевины, может в результате двух стадий
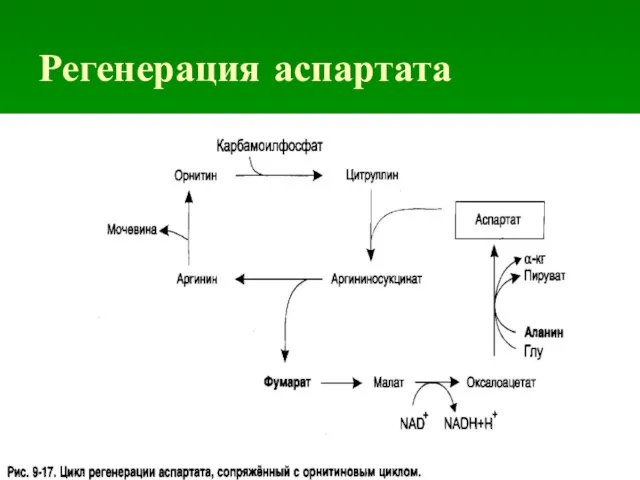
Регенерация аспартата
Регенерация аспартата
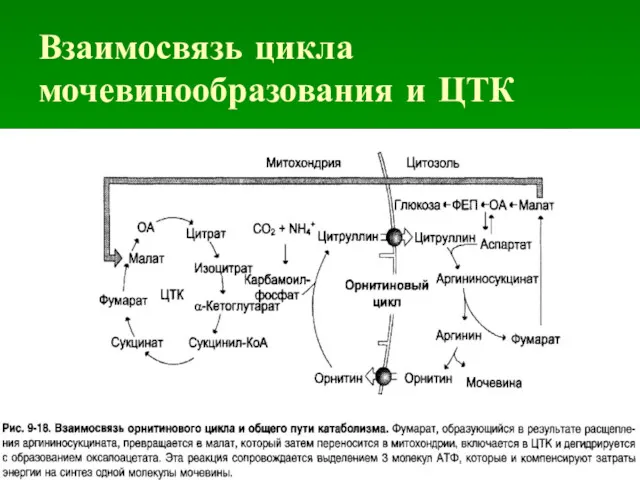
Взаимосвязь цикла мочевинообразования и ЦТК
Взаимосвязь цикла мочевинообразования и ЦТК

ЭНЕРГОЗАВИСИМЫЙ ПРОЦЕСС
Биосинтез мочевины требует больших затрат энергии. Энергия поставляется за счет
ЭНЕРГОЗАВИСИМЫЙ ПРОЦЕСС
Биосинтез мочевины требует больших затрат энергии. Энергия поставляется за счет

КОМПАРТМЕНТАЛИЗАЦИЯ
Цикл мочевины протекает исключительно в печени. Он разделен на два компартмента:
КОМПАРТМЕНТАЛИЗАЦИЯ
Цикл мочевины протекает исключительно в печени. Он разделен на два компартмента:

АЛЛОСТЕРИЧЕСКАЯ РЕГУЛЯЦИЯ СИНТЕЗА МОЧЕВИНЫ
Скорость синтеза мочевины определяется первой реакцией цикла.
АЛЛОСТЕРИЧЕСКАЯ РЕГУЛЯЦИЯ СИНТЕЗА МОЧЕВИНЫ
Скорость синтеза мочевины определяется первой реакцией цикла.

Наследственные нарушения орнитинового цикла и их симптомы
Наследственные нарушения орнитинового цикла и их симптомы

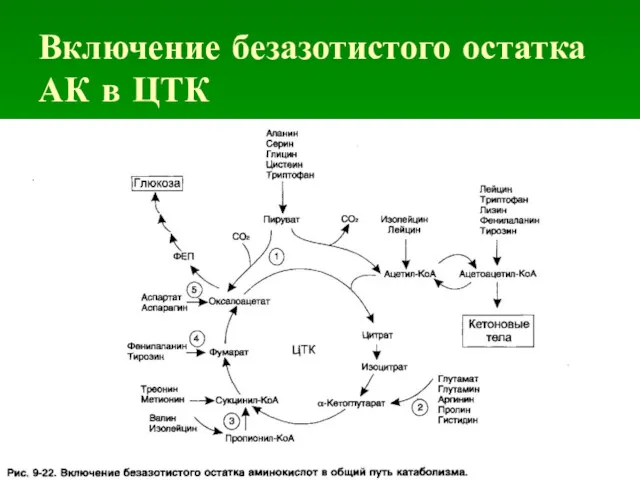
Включение безазотистого остатка АК в ЦТК
Включение безазотистого остатка АК в ЦТК

Пути биосинтеза заменимых АК
Пути биосинтеза заменимых АК
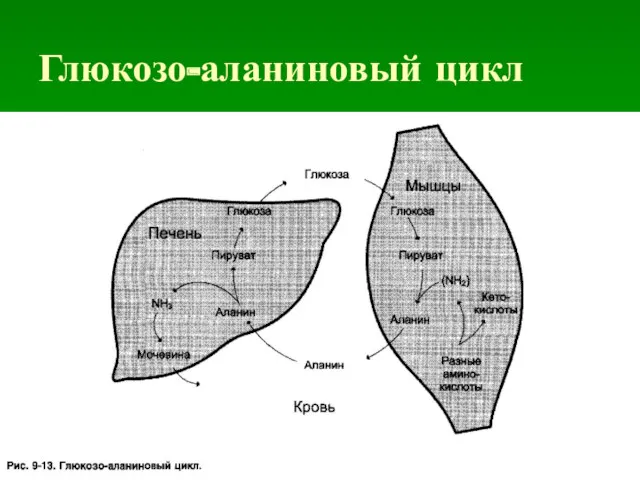
Глюкозо-аланиновый цикл
Глюкозо-аланиновый цикл
 Роль соли для организма человека
Роль соли для организма человека Мембраналық органоидтар
Мембраналық органоидтар Эволюционная теория Жана Батиста Ламарка
Эволюционная теория Жана Батиста Ламарка Презентация у уроку биологии по теме Тип Кишечнополостные (7 класс)
Презентация у уроку биологии по теме Тип Кишечнополостные (7 класс) Необычные птицы планеты
Необычные птицы планеты Witaminy - są to substancje organiczne niezbędne w niewielkich ilościach do prawidłowego przebiegu procesów
Witaminy - są to substancje organiczne niezbędne w niewielkich ilościach do prawidłowego przebiegu procesów Мутагенные факторы и их влияние на фенотип человека
Мутагенные факторы и их влияние на фенотип человека Паукообразные. Разновидности паукообразных
Паукообразные. Разновидности паукообразных Память и обучение
Память и обучение Презентация Оценка зависимости ЖЕЛ у подростков
Презентация Оценка зависимости ЖЕЛ у подростков Исследовательская работа. Выращивание дуба
Исследовательская работа. Выращивание дуба Животные и растения Астраханского края, занесённые в Красную книгу. 2 класс
Животные и растения Астраханского края, занесённые в Красную книгу. 2 класс Удивительный мир фитодизайна
Удивительный мир фитодизайна Бурый медведь
Бурый медведь Структурные уровни организации живого. Свойства живого
Структурные уровни организации живого. Свойства живого Использование ИКТ на уроках биологии как средство повышения эффективности учебно-воспитательного процесса
Использование ИКТ на уроках биологии как средство повышения эффективности учебно-воспитательного процесса Физиология мышечного сокращения
Физиология мышечного сокращения Способы культивирования вирусов
Способы культивирования вирусов Лаборатория Полимеры для биологии
Лаборатория Полимеры для биологии Развитие эволюционных идей
Развитие эволюционных идей Клеточные и неклеточные формы жизни
Клеточные и неклеточные формы жизни Закономерности влияния окружающей среды на развитие фенотипа животных. (Лекция 1)
Закономерности влияния окружающей среды на развитие фенотипа животных. (Лекция 1) презентация Вид, критерии 11 класс
презентация Вид, критерии 11 класс Врожденные формы поведения. Сравнение врожденных и приобретенных форм поведения
Врожденные формы поведения. Сравнение врожденных и приобретенных форм поведения Химический состав клетки
Химический состав клетки Развитие жизни на Земле в различные периоды
Развитие жизни на Земле в различные периоды Приспособление организмов к среде обитания
Приспособление организмов к среде обитания Клетка — элементарная структурная и функциональная единица живого
Клетка — элементарная структурная и функциональная единица живого