- Конформация биомакромолекул. Виды конформационных перестроек

Содержание
- 2. ЧТО ТАКОЕ «КОНФОРМАЦИЯ» Конформация макромолекулы - это способ укладки полимерной цепи (без разрыва ковалентных связей) за
- 3. Изменения параметров окружающей среды (температура, рН, ионная сила раствора, действие денатурирующих факторов) вызывают конформационную перестройку биомакромолекул
- 4. ПРОСТРАНСТВЕННАЯ ОРГАНИЗАЦИЯ БЕЛКА
- 5. ВИДЫ КОНФОРМАЦИЙ Четвертичная Третичная Вторичная Первичная
- 6. Первичная структура белка Белки - биополимеры, состоящие из аминокислот. Различают L- и D- формы (оптические изомеры)
- 7. правозакрученная (ϕ=132о, ψ=123о) левозакрученная (ϕ =228о, ψ=237о) Вторичная структура белка Выделяют правозакрученную и левозакрученную α-спираль и
- 8. Сверхвторичные структуры Термодинамически и кинетически стабильные комплексы α-спиралей и β-складчатых структур, образующиеся за счет взаимодействия между
- 9. Третичная структура Пространственная ориентация полипептидной спирали или способ укладки полипептидной цепи в определенном объеме. Результат –
- 10. Термодинамика образования глобулы СВОБОДНАЯ ЭНЕРГИЯ ГИББСА ΔG = ΔH - TΔS ДЛЯ САМОПРОИЗВОЛЬНЫХ ПРОЦЕССОВ ΔG ВЫГОДНО
- 11. При свертывании белковой глобулы выигрыша в числе водородных связей не происходит, т. к. одновременно утрачиваются водородные
- 12. Четвертичная структура Образуется, когда гидрофильные остатки не полностью закрывают гидрофобное ядро, что приводит к образованию надмолекулярных
- 13. ЗАКОНОМЕРНОСТИ ПРОСТРАНСТВЕННОЙ ОРИЕНТАЦИИ БИОМАКРОМОЛЕКУЛ
- 14. СПЕЦИФИКА БИОМАКРОМОЛЕКУЛ Специфика полимерных молекул в отличие от малых молекул определяется большим числом однотипных звеньев (мономеров),
- 15. Своеобразие биологических макромолекул как физического объекта заключается в тесном сочетании статистических и детерминистских (механических) особенностей их
- 16. Если будем считать, что полимерная цепь состоит из ряда прямолинейных сегментов, каждый из которых включает определенное
- 17. Разбиение реальной цепи на статистические сегменты должно происходить так, чтобы число звеньев m в составе сегмента
- 18. Свободно-сочлененная цепь (по В. Н. Цветкову, В. Е. Эскину, С. Я. Френкелю, 1964) Реальная длинная цепная
- 19. Сворачивание гибкой цепи в клубок определяется ее термодинамической гибкостью: чем ↑ гибкость, тем ↓ h2 при
- 20. ОБЪЕМНЫЕ ВЗАИМОДЕЙСТВИЯ. ПЕРЕХОД БЕЛОК-ГЛОБУЛА
- 21. Глобула – вид пространственной структуры полимера, с компактной пространственной структурой, с малыми флуктуациями плотности, с однородной
- 22. Вследствие объёмных взаимодействий сблизившиеся участки полимера могут притягиваться или отталкиваться. Повышение температуры приводит к увеличению отталкивания
- 23. В θ-точке объемные взаимодействия отсутствуют, и макромолекула представляет клубок с h~lN1/2, сохраняющийся и при температуре T
- 24. 1. Полная компенсация притяжения и отталкивания в θ -точке является специфическим свойством полимеров. Это явление связано
- 25. ТЕМПЕРАТУРНАЯ ДЕНАТУРАЦИЯ В процессе тепловой денатурации происходит изменение теплоемкости биополимера при переходе от нативного (спиральное, глобулярное
- 26. Изменение энтропии этого процесса составляет: Было показано, что молекула белка может претерпевать обратимые конформационные переходы в
- 27. ПОВОРОТНАЯ ИЗОМЕРИЯ И СТЕРИЧЕСКИЕ ОГРАНИЧЕНИЯ Зависимость конформации линейной полимерной цепи от характера взаимодействия атомных групп
- 28. Сворачивание полипептидной цепи (как и любой другой полимерной молекулы) есть статистическая и механическая форма поведения, зависящая
- 29. Энергия вращения атомных групп вокруг единичных связей дает основной вклад в общую конформационную энергию полимерной цепи.
- 30. КОНФОРМАЦИОННАЯ ЭНЕРГИЯ ПОЛИПЕПТИДНОЙ ЦЕПИ И ОСОБЕННОСТИ ЕЕ СТРОЕНИЯ Конформационная энергия полипептидной цепи определяется всеми видами взаимодействий
- 31. Последовательность углов ϕ и ψ для всех остатков белка определяет конформацию белковой цепи. Принцип, согласно которому
- 32. ПРЕДСКАЗАНИЕ И МОДЕЛИРОВАНИЕ ПРОСТРАНСТВЕННОЙ ОРГАНИЗАЦИИ БЕЛКОВ ПО ИХ ПЕРВИЧНОЙ СТРУКТУРЕ Основной особенностью пространственной структуры белка является
- 33. Эмпирический метод основан на выявлении корреляции между вторичной структурой белка и его аминокислотным составом и последовательностью.
- 34. МЕТОДЫ ПРЕДСКАЗАНИЯ СТРУКТУРЫ БЕЛКОВ ПО АМИНОКИСЛОТНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ 1. Предсказание вторичной структуры белка без укладки ее в
- 35. МЕТОДЫ ИССЛЕДОВАНИЯ ВНУТРИМОЛЕКУЛЯРНОЙ ДИНАМИКИ БЕЛКОВ Изучение быстрых внутренних движений в молекуле белка осуществляется с помощью современных
- 36. ПРОСТРАНСТВЕННАЯ ОРГАНИЗАЦИЯ НУКЛЕИНОВЫХ КИСЛОТ дезоксирибонуклеиновая (ДНК) рибонуклеиновая кислота (РНК)
- 37. Первичная структура Нити НК представляют непрерывную цепь ковалентно соединенных фосфодиэфирными связями фосфатных групп с остатками сахара.
- 38. Вторичная структура ДНК ДНК – двойная спираль, образованная двумя полинуклеотидными цепями, которые стабилизированы водородными связями между
- 39. Конформации ДНК Конформации кольца дезоксирибозы пиранозы: A – азотистое основание; x – угол ориентации азотистого основания;
- 40. Конформации ДНК B-DNA A-DNA Z-DNA B-ДНК – обусловлена C-(2ʹ)-эндоконформацией кольца дезоксирибозы; A-ДНК – обусловлена C-(3ʹ)-эндоконформацией кольца
- 41. Третичная структура ДНК
- 44. Скачать презентацию

ЧТО ТАКОЕ «КОНФОРМАЦИЯ»
Конформация макромолекулы - это способ укладки полимерной цепи (без разрыва
ЧТО ТАКОЕ «КОНФОРМАЦИЯ»
Конформация макромолекулы - это способ укладки полимерной цепи (без разрыва
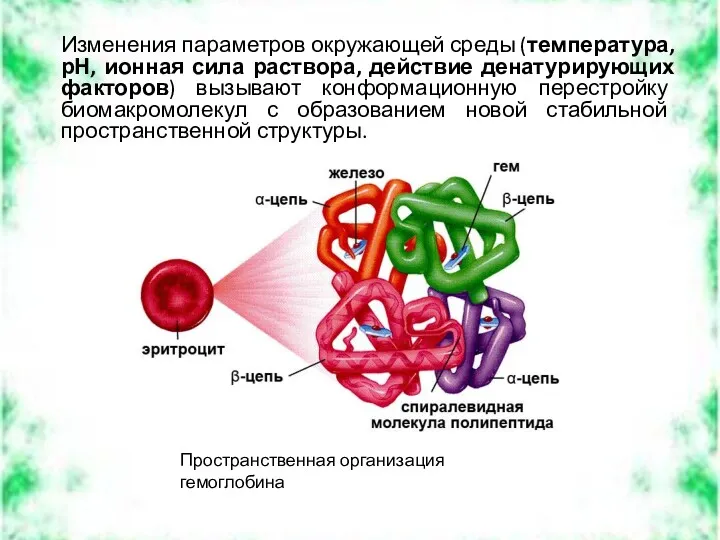
Изменения параметров окружающей среды (температура, рН, ионная сила раствора, действие денатурирующих
Изменения параметров окружающей среды (температура, рН, ионная сила раствора, действие денатурирующих

ПРОСТРАНСТВЕННАЯ ОРГАНИЗАЦИЯ БЕЛКА
ПРОСТРАНСТВЕННАЯ ОРГАНИЗАЦИЯ БЕЛКА
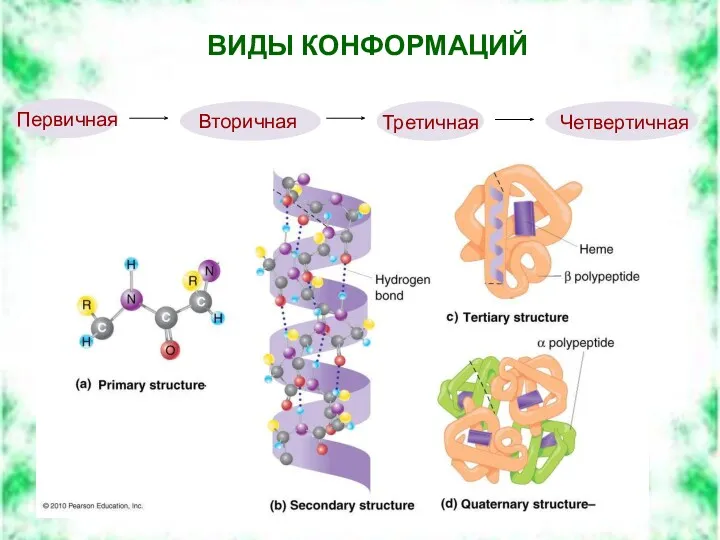
ВИДЫ КОНФОРМАЦИЙ
Четвертичная
Третичная
Вторичная
Первичная
ВИДЫ КОНФОРМАЦИЙ
Четвертичная
Третичная
Вторичная
Первичная
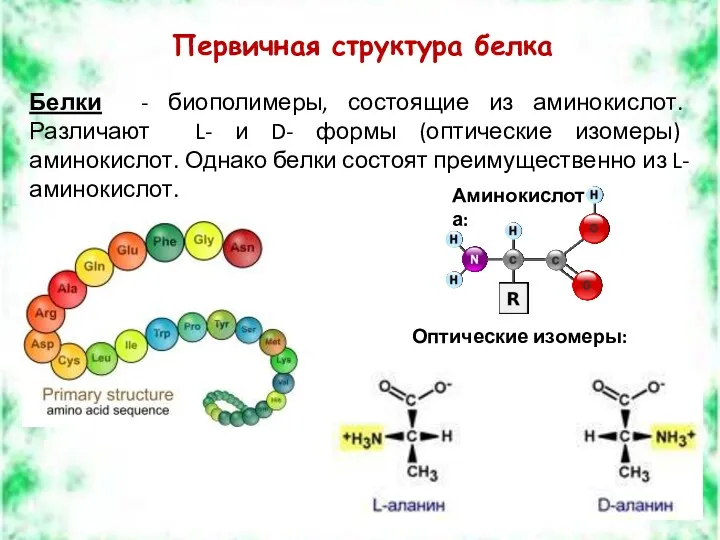
Первичная структура белка
Белки - биополимеры, состоящие из аминокислот. Различают L- и
Первичная структура белка
Белки - биополимеры, состоящие из аминокислот. Различают L- и
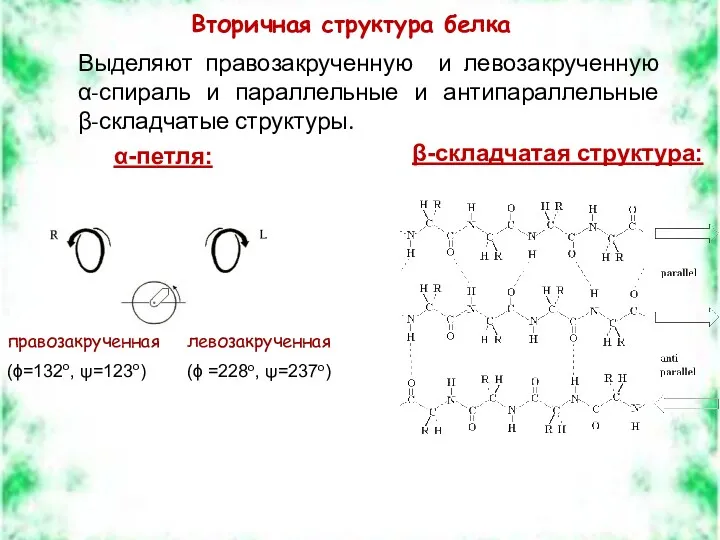
правозакрученная
(ϕ=132о, ψ=123о)
левозакрученная
(ϕ =228о, ψ=237о)
Вторичная структура белка
Выделяют правозакрученную и левозакрученную
α-спираль и параллельные и
правозакрученная
(ϕ=132о, ψ=123о)
левозакрученная
(ϕ =228о, ψ=237о)
Вторичная структура белка
Выделяют правозакрученную и левозакрученную α-спираль и параллельные и
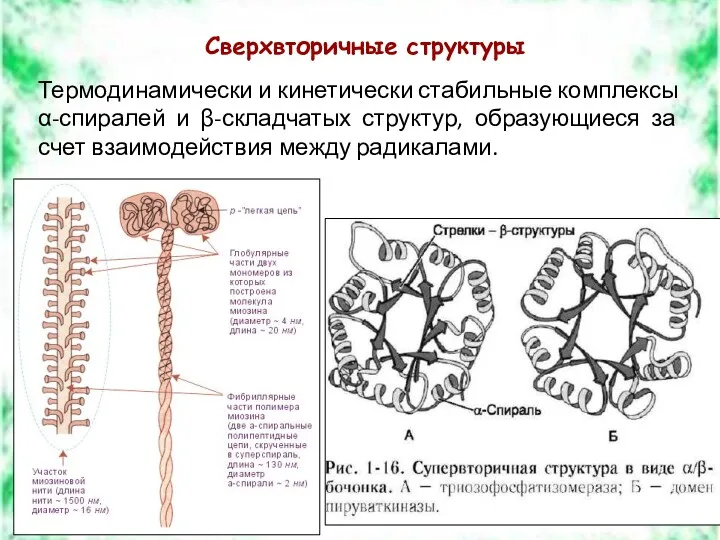
Сверхвторичные структуры
Термодинамически и кинетически стабильные комплексы
α-спиралей и β-складчатых структур, образующиеся
Сверхвторичные структуры
Термодинамически и кинетически стабильные комплексы α-спиралей и β-складчатых структур, образующиеся
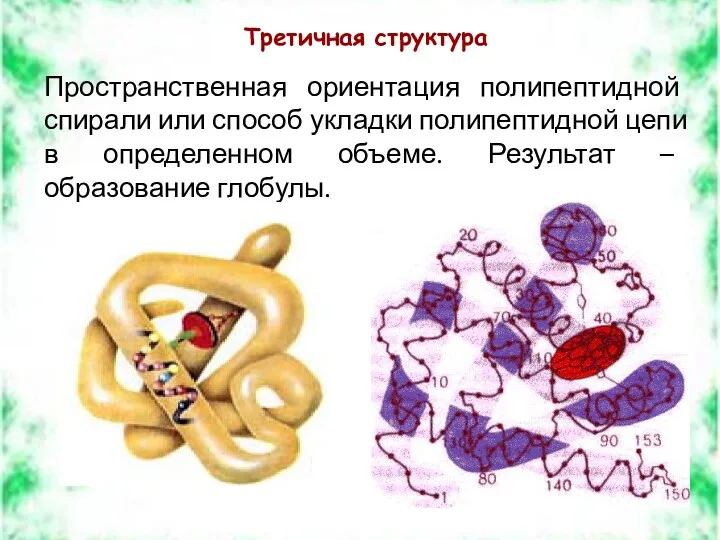
Третичная структура
Пространственная ориентация полипептидной спирали или способ укладки полипептидной цепи в
Третичная структура
Пространственная ориентация полипептидной спирали или способ укладки полипептидной цепи в
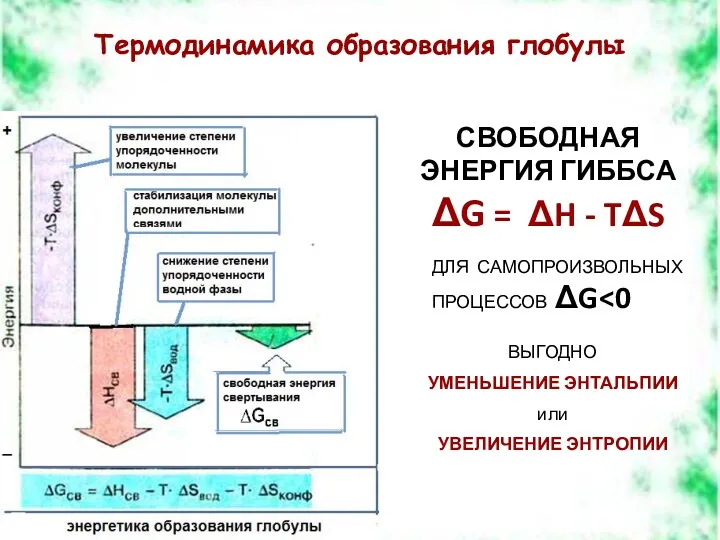
Термодинамика образования глобулы
СВОБОДНАЯ ЭНЕРГИЯ ГИББСА
ΔG = ΔH - TΔS
ДЛЯ САМОПРОИЗВОЛЬНЫХ
Термодинамика образования глобулы
СВОБОДНАЯ ЭНЕРГИЯ ГИББСА
ΔG = ΔH - TΔS
ДЛЯ САМОПРОИЗВОЛЬНЫХ

При свертывании белковой глобулы выигрыша в числе водородных связей не происходит,
При свертывании белковой глобулы выигрыша в числе водородных связей не происходит,
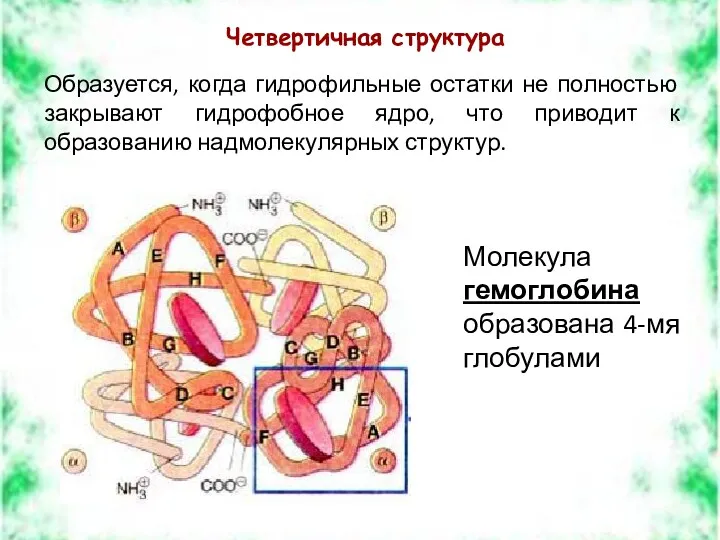
Четвертичная структура
Образуется, когда гидрофильные остатки не полностью закрывают гидрофобное ядро, что
Четвертичная структура
Образуется, когда гидрофильные остатки не полностью закрывают гидрофобное ядро, что

ЗАКОНОМЕРНОСТИ ПРОСТРАНСТВЕННОЙ ОРИЕНТАЦИИ БИОМАКРОМОЛЕКУЛ
ЗАКОНОМЕРНОСТИ ПРОСТРАНСТВЕННОЙ ОРИЕНТАЦИИ БИОМАКРОМОЛЕКУЛ

СПЕЦИФИКА БИОМАКРОМОЛЕКУЛ
Специфика полимерных молекул в отличие от малых молекул определяется большим
СПЕЦИФИКА БИОМАКРОМОЛЕКУЛ
Специфика полимерных молекул в отличие от малых молекул определяется большим

Своеобразие биологических макромолекул как физического объекта заключается в тесном сочетании статистических
Своеобразие биологических макромолекул как физического объекта заключается в тесном сочетании статистических

Если будем считать, что полимерная цепь состоит из ряда прямолинейных сегментов,
Если будем считать, что полимерная цепь состоит из ряда прямолинейных сегментов,

Разбиение реальной цепи на статистические сегменты должно происходить так, чтобы число
Разбиение реальной цепи на статистические сегменты должно происходить так, чтобы число
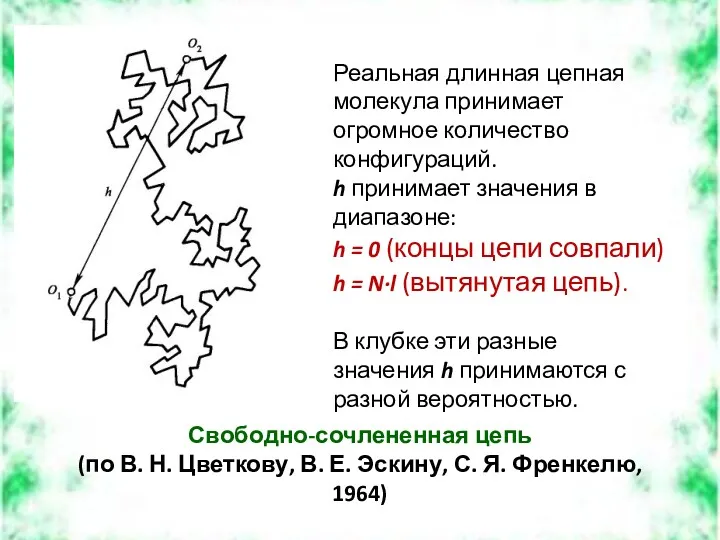
Свободно-сочлененная цепь
(по В. Н. Цветкову, В. Е. Эскину, С. Я.
Свободно-сочлененная цепь
(по В. Н. Цветкову, В. Е. Эскину, С. Я.

Сворачивание гибкой цепи в клубок определяется ее термодинамической гибкостью:
чем ↑
Сворачивание гибкой цепи в клубок определяется ее термодинамической гибкостью:
чем ↑

ОБЪЕМНЫЕ ВЗАИМОДЕЙСТВИЯ.
ПЕРЕХОД БЕЛОК-ГЛОБУЛА
ОБЪЕМНЫЕ ВЗАИМОДЕЙСТВИЯ.
ПЕРЕХОД БЕЛОК-ГЛОБУЛА

Глобула – вид пространственной структуры полимера, с компактной пространственной структурой, с
Глобула – вид пространственной структуры полимера, с компактной пространственной структурой, с
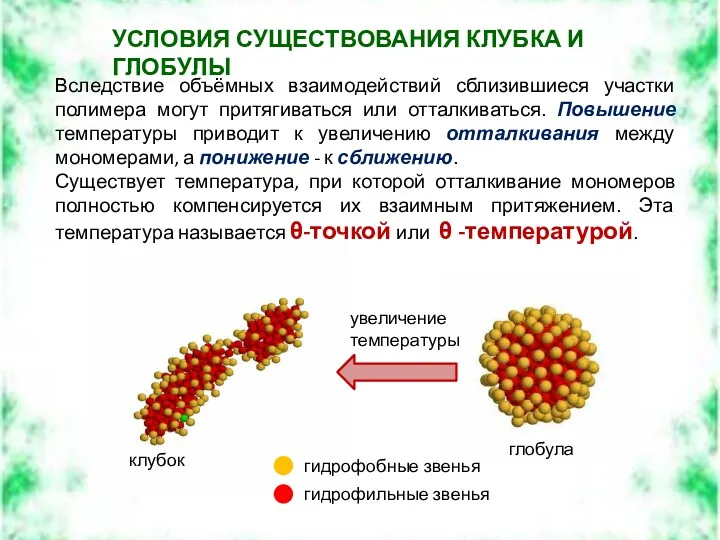
Вследствие объёмных взаимодействий сблизившиеся участки полимера могут притягиваться или отталкиваться. Повышение
Вследствие объёмных взаимодействий сблизившиеся участки полимера могут притягиваться или отталкиваться. Повышение

В θ-точке объемные взаимодействия отсутствуют, и макромолекула представляет клубок с h~lN1/2, сохраняющийся и
В θ-точке объемные взаимодействия отсутствуют, и макромолекула представляет клубок с h~lN1/2, сохраняющийся и

1. Полная компенсация притяжения и отталкивания в
θ -точке является специфическим
1. Полная компенсация притяжения и отталкивания в
θ -точке является специфическим

ТЕМПЕРАТУРНАЯ ДЕНАТУРАЦИЯ
В процессе тепловой денатурации происходит изменение теплоемкости биополимера при переходе
ТЕМПЕРАТУРНАЯ ДЕНАТУРАЦИЯ
В процессе тепловой денатурации происходит изменение теплоемкости биополимера при переходе

Изменение энтропии этого процесса составляет:
Было показано, что молекула белка может претерпевать
Изменение энтропии этого процесса составляет:
Было показано, что молекула белка может претерпевать

ПОВОРОТНАЯ ИЗОМЕРИЯ И СТЕРИЧЕСКИЕ ОГРАНИЧЕНИЯ
Зависимость конформации линейной полимерной цепи от
ПОВОРОТНАЯ ИЗОМЕРИЯ И СТЕРИЧЕСКИЕ ОГРАНИЧЕНИЯ
Зависимость конформации линейной полимерной цепи от

Сворачивание полипептидной цепи (как и любой другой полимерной молекулы) есть статистическая
Сворачивание полипептидной цепи (как и любой другой полимерной молекулы) есть статистическая

Энергия вращения атомных групп вокруг единичных связей дает основной вклад в
Энергия вращения атомных групп вокруг единичных связей дает основной вклад в
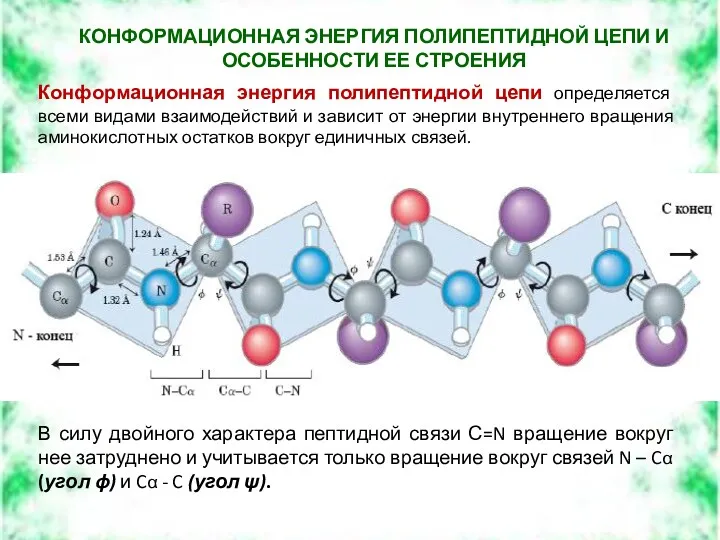
КОНФОРМАЦИОННАЯ ЭНЕРГИЯ ПОЛИПЕПТИДНОЙ ЦЕПИ И ОСОБЕННОСТИ ЕЕ СТРОЕНИЯ
Конформационная энергия полипептидной цепи
КОНФОРМАЦИОННАЯ ЭНЕРГИЯ ПОЛИПЕПТИДНОЙ ЦЕПИ И ОСОБЕННОСТИ ЕЕ СТРОЕНИЯ
Конформационная энергия полипептидной цепи

Последовательность углов ϕ и ψ для всех остатков белка определяет конформацию
Последовательность углов ϕ и ψ для всех остатков белка определяет конформацию

ПРЕДСКАЗАНИЕ И МОДЕЛИРОВАНИЕ ПРОСТРАНСТВЕННОЙ ОРГАНИЗАЦИИ БЕЛКОВ ПО ИХ ПЕРВИЧНОЙ СТРУКТУРЕ
Основной особенностью
ПРЕДСКАЗАНИЕ И МОДЕЛИРОВАНИЕ ПРОСТРАНСТВЕННОЙ ОРГАНИЗАЦИИ БЕЛКОВ ПО ИХ ПЕРВИЧНОЙ СТРУКТУРЕ
Основной особенностью

Эмпирический метод основан на выявлении корреляции между вторичной структурой белка и
Эмпирический метод основан на выявлении корреляции между вторичной структурой белка и

МЕТОДЫ ПРЕДСКАЗАНИЯ СТРУКТУРЫ БЕЛКОВ ПО АМИНОКИСЛОТНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ
1. Предсказание вторичной структуры
МЕТОДЫ ПРЕДСКАЗАНИЯ СТРУКТУРЫ БЕЛКОВ ПО АМИНОКИСЛОТНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ
1. Предсказание вторичной структуры

МЕТОДЫ ИССЛЕДОВАНИЯ ВНУТРИМОЛЕКУЛЯРНОЙ ДИНАМИКИ БЕЛКОВ
Изучение быстрых внутренних движений в молекуле белка
МЕТОДЫ ИССЛЕДОВАНИЯ ВНУТРИМОЛЕКУЛЯРНОЙ ДИНАМИКИ БЕЛКОВ
Изучение быстрых внутренних движений в молекуле белка

ПРОСТРАНСТВЕННАЯ ОРГАНИЗАЦИЯ НУКЛЕИНОВЫХ КИСЛОТ
дезоксирибонуклеиновая (ДНК)
рибонуклеиновая кислота (РНК)
ПРОСТРАНСТВЕННАЯ ОРГАНИЗАЦИЯ НУКЛЕИНОВЫХ КИСЛОТ
дезоксирибонуклеиновая (ДНК)
рибонуклеиновая кислота (РНК)
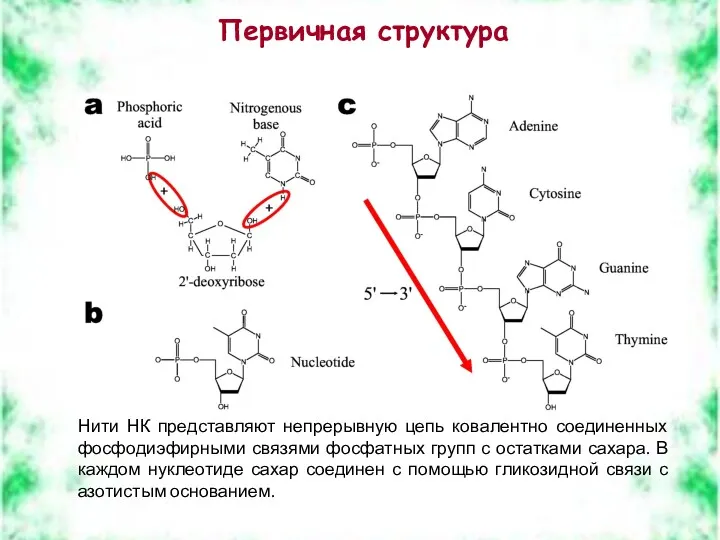
Первичная структура
Нити НК представляют непрерывную цепь ковалентно соединенных фосфодиэфирными связями фосфатных
Первичная структура
Нити НК представляют непрерывную цепь ковалентно соединенных фосфодиэфирными связями фосфатных
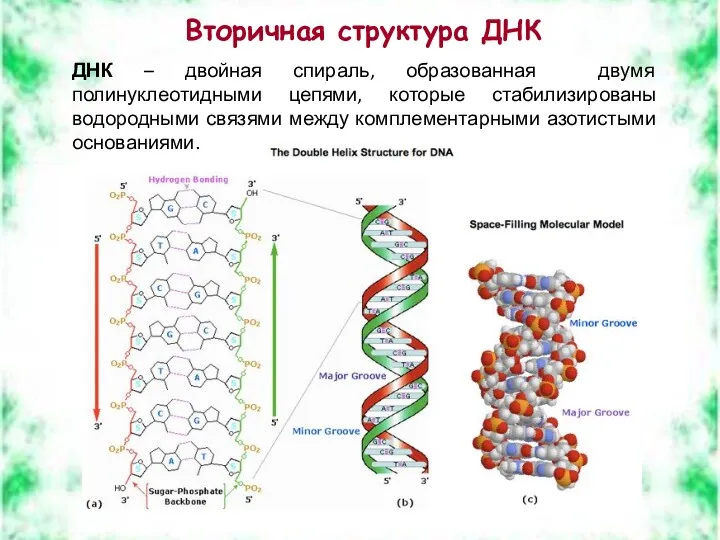
Вторичная структура ДНК
ДНК – двойная спираль, образованная двумя полинуклеотидными цепями, которые
Вторичная структура ДНК
ДНК – двойная спираль, образованная двумя полинуклеотидными цепями, которые
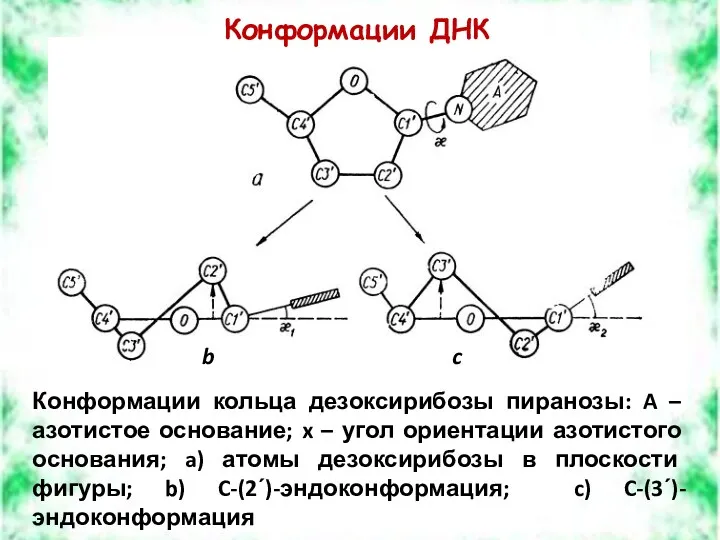
Конформации ДНК
Конформации кольца дезоксирибозы пиранозы: A – азотистое основание; x –
Конформации ДНК
Конформации кольца дезоксирибозы пиранозы: A – азотистое основание; x –
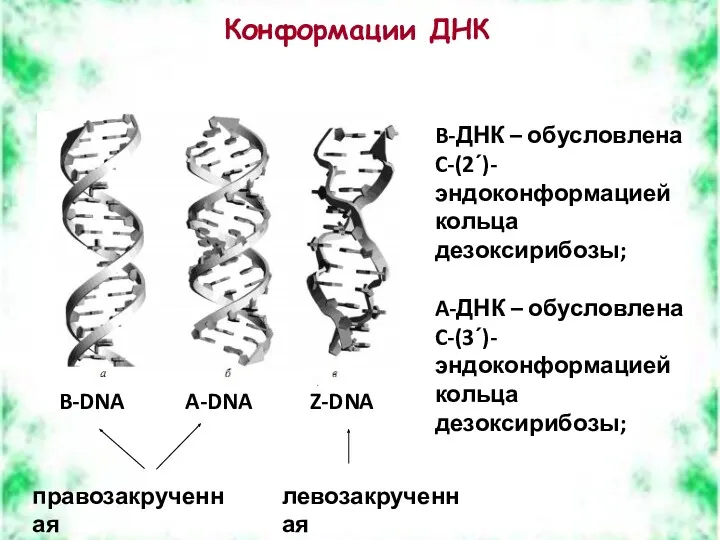
Конформации ДНК
B-DNA
A-DNA
Z-DNA
B-ДНК – обусловлена C-(2ʹ)-эндоконформацией кольца дезоксирибозы;
A-ДНК – обусловлена C-(3ʹ)-эндоконформацией кольца
Конформации ДНК
B-DNA
A-DNA
Z-DNA
B-ДНК – обусловлена C-(2ʹ)-эндоконформацией кольца дезоксирибозы;
A-ДНК – обусловлена C-(3ʹ)-эндоконформацией кольца
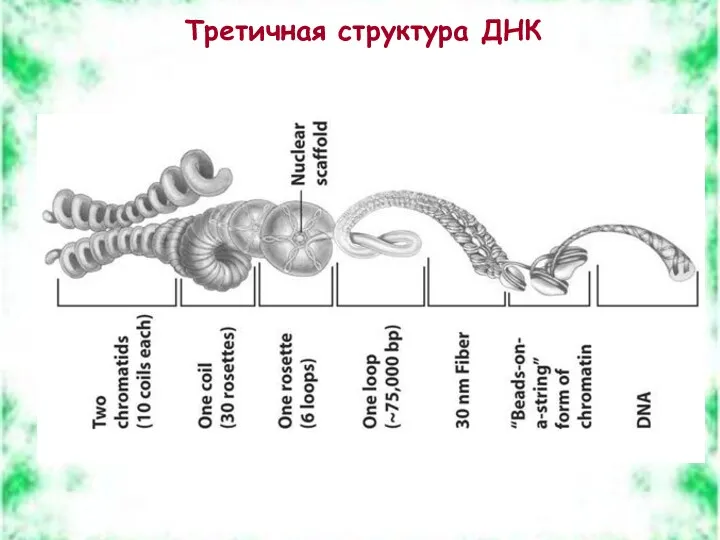
Третичная структура ДНК
Третичная структура ДНК
Похожие презентации
 Птицы Беларуси
Птицы Беларуси Биология клетки. Уровни организации жизни на Земле
Биология клетки. Уровни организации жизни на Земле Антропогенез. Происхождение человека
Антропогенез. Происхождение человека Царство животные. Тип хордовые. Класс птицы
Царство животные. Тип хордовые. Класс птицы Физиологические свойства мышц
Физиологические свойства мышц Почему вымирают животные?
Почему вымирают животные?
 Съедобные и ядовитые грибы Красноярского края
Съедобные и ядовитые грибы Красноярского края Анатомия верхних и нижних дыхательных путей
Анатомия верхних и нижних дыхательных путей Грибы, лишайники. (Тема 7)
Грибы, лишайники. (Тема 7) Ядовитые растения России
Ядовитые растения России Утомление и переутомление
Утомление и переутомление Биологическая и социальная природа человека
Биологическая и социальная природа человека Протизаплідні засоби
Протизаплідні засоби История развития биологии. Введение в общую биологию. 10 класс
История развития биологии. Введение в общую биологию. 10 класс Лимфатическая система
Лимфатическая система Нервная система
Нервная система Синичкин день
Синичкин день презентация Промысловые рыбы по теме урока Промысловые рыбы. Их использование и охрана
презентация Промысловые рыбы по теме урока Промысловые рыбы. Их использование и охрана Использование здоровье-сберегающих технологий в учебно-образовательном процессе
Использование здоровье-сберегающих технологий в учебно-образовательном процессе Дикие животные. Белка
Дикие животные. Белка Мастер-класс Первый шаг в биологию
Мастер-класс Первый шаг в биологию opasnost_ot_zhivotnykh
opasnost_ot_zhivotnykh Генно-инженерно-модифицированные организмы. Методы детекции и идентификации
Генно-инженерно-модифицированные организмы. Методы детекции и идентификации Ракообразные (лат. Crustacea)
Ракообразные (лат. Crustacea) Волк. Охота волков
Волк. Охота волков Класс млекопитающие
Класс млекопитающие Сон. Правила здорового сна
Сон. Правила здорового сна