- Кружок Основы молекулярной генетики

Содержание
- 2. Молекулярные основы наследственности. 1. Центральная догма молекулярной биологии. Типы переноса генетической информации в живых системах: общий,
- 3. Центральная догма ДНК РНК белок Репликация Транскрипция Трансляция Обратная транскрипция Репликация РНК Только РНК-вирусы Ретро-РНК-вирусы Другие
- 5. ДНК РНК белок Матричные синтезы, разрешенные по центральной догме Не обнаружен
- 6. Запрещенные матричные синтезы Белки никогда не бывают матрицами
- 7. Центральная догма ДНК РНК белок Репликация Транскрипция Трансляция Обратная транскрипция Репликация РНК
- 8. 2. Репликация ДНК Универсальный биологический процесс передачи генетической информации в поколениях клеток и организмов, благодаря созданию
- 9. Место репликации в клеточном цикле Репликация ДНК всегда предшествует делению клетки. Репликация S-период (Synthesis) Интерфаза Деление
- 11. Принципы репликации 1. Полуконсервативность 2. Комплементарность 3. Антипараллельность 4. Униполярность 5. Прерывистость
- 12. Полуконсервативность – каждая исходная (материнская) цепь ДНК выступает в качестве матрицы для синтеза дочерней цепи Полуконсервативный
- 13. Т.Е. дочерняя молекула получает одну нить от материнской ДНК, а вторую синтезирует вновь Дочерняя цепь
- 14. Комплементарность Вновь синтезируемая ( дочерняя) цепь ДНК строится по принципу комплементарности. В состав растущей цепи включается
- 15. Антипараллельность – синтез дочерней цепи ДНК происходит в противоположном от материнской цепи направлении
- 16. Униполярность: Удвоение цепи ДНК идет в направлении от 5` конца к 3` концу, следовательно новый нуклеотид
- 18. Репликон – расстояние между двумя сайтами начала репликации ori ~ 100 тыс. н.п. У прокариот вся
- 19. Прерывистость репликации ДНК одной хромосомы ori ori Репликативные вилки
- 20. Репликативная вилка 3' 5' 3' 3' Запаздывающая цепь Лидирующая цепь Направление движения вилки Фрагменты Оказаки
- 21. Репликация ДНК Скорость репликации огромна, т.к. реакция идет в нескольких местах одновременно – ориджины репликации. Сайты
- 24. ДНК ТОПОИЗОМЕРАЗА – фермент, изменяющий степень сверхспиральности, возникающее при раскручивании двух цепей в репликативной вилке ДНК
- 26. Репликация ДНК Количество раундов репликации ДНК (а значит число возможных делений клетки) зависит от длины теломерных
- 27. Репликация ДНК ДНК- полимеразы Δ и ε делают 1 ошибку на 105 - 106 нуклеотидов (ДНК-полимераза
- 29. Репликация ДНК Ошибки в ДНК (мутации) возникают спонтанно (ошибки репликации, дезаминирование нуклеотидов, депуринизация ДНК и т.д.)
- 30. Репликация ДНК Комплекс ферментов репарации узнает и вырезает поврежденные и химически измененные нуклеотиды, ДНК-полимераза β встраивает
- 32. Значение для медицины Иногда в растующую цепь случайно вклинивается неправильное основание, однако у здоровых клеток присутствует
- 33. Выводы по репликации ДНК В результате репликации каждая дочерняя клетка получает точную копию всей ДНК содержавшейся
- 34. Проблема укорочения концов у линейных ДНК Сформулирована – А.М. Оловников, 1971 При каждой репликации новые цепи
- 35. Гипотеза Оловникова Укорочение концов – это внутренние часы, отмеряющие время жизни многоклеточного организма – число отпущенных
- 36. Но почему тогда клетки зародышевой линии делятся бесконечно? Оловников: должен существовать механизм удлинения концов хромосом. Теломераза
- 37. Теломераза фермент, надстраивающий концы хромосом, содержит РНК. удлинение происходит путем обратной транскрипции: РНК → ДНК На
- 38. Теломераза активна в клетках зародышевого пути эмбриональных стволовых раковых – поэтому они бессмертны Теломераза неактивна в
- 39. Значение для медицины После каждого клеточного цикла теломеры укорачиваются на один повтор, а следовательно, количество делений
- 40. 4.Транскрипция. Механизмы транскрипции у про- и эукариот. Транскрипция - это первый этап реализации генетической информации, при
- 41. Центральная догма ДНК РНК белок Репликация Транскрипция Трансляция Обратная транскрипция Репликация РНК
- 42. Транскрипция Считывание информации с ДНК-матрицы на РНК, синтез тРНК, иРНК, рРНК с помощью одной полимеразы (у
- 43. Транскрипция Механизм РНК – полимеразной реакции тот же, что и ДНК – полимеразной, направление синтеза 5?3,
- 44. Транскрипция В ДНК – матрице выделяют транскиптоны. Участки, ограниченные промоторами и сайтами терминации, между которыми 1
- 45. Строение гена эукариот, кодирующего белок Ген ( в узком смысле слова) – это участок ДНК, в
- 46. Транскрипция 3 стадии транскрипции: инициация, элонгация и терминация. Инициация синтеза начинается с «узнавания» полимеразой промоторного сайта
- 47. Инициация транскрипции Для формирование транскрипционной вилки (раскручивание одного витка спирали ДНК-матрицы) к ТАТА-боксу присоединяется белковый фактор
- 48. Элонгация транскрипции Белковые факторы элонгации обеспечивают расплетение ДНК перед продвижением РНК-полимеразы и восстановление двойной спирали позади
- 49. Терминация транскрипции При достижении РНК - полимеразой сайта терминации белковый фактор терминации освобождает пре-РНК из комплекса
- 50. Особенности транскрипции у эукариот Транскрипция 1. Кэп и поли-А-хвост Созревание м-РНК 2.Сплайсинг цитоплазма
- 51. Созревание РНК-транскриптов
- 52. ДНК одного гена П Т Промотор Терминатор Интроны и экзоны Интроны – вставки в эукариотические гены,
- 53. ДНК одного гена П Т Промотор Терминатор пре- м-РНК Транскрипция Сплайсинг (вырезание интронов) зрелая м-РНК В
- 54. ДНК одного гена П Т Промотор Терминатор пре- м-РНК Сплайсинг в клетке 1 зрелая м-РНК Сплайсинг
- 55. Альтернативный сплайсинг в разных органах на разных стадиях развития в разных состояниях клетки 94% генов человека
- 56. Значение для медицины Иногда в некоторых транскриптах обнаруживают альтернативные механизмы сплайсинга, однако ошибки в данном процессе
- 57. Созревание РНК-транскриптов Процессингу (созреванию) подвергаются все виды РНК (и, т, р). А) Ковалентная модификация 5- и
- 58. Ковалентная модификация иРНК Гуанилил-трансфераза присоединяет ГДФ к 5- ОР концу (5-О-Р-О-5 связь), 5 – кэпирование происходит
- 59. СПЛАЙСИНГ иРНК Сплайсинг: образование зрелой мРНК: Вырезание интронных последовательностей (ограниченных AGGU- и - GAGG- последовательностями) с
- 62. 6. Трансляция. Механизмы трансляции Трансляция- это второй этап реализации генетической информации. При этом происходит перевод наследственной
- 63. Центральная догма ДНК РНК белок Репликация Транскрипция Трансляция Обратная транскрипция Репликация РНК Только РНК-вирусы Ретро-РНК-вирусы Другие
- 64. Этапы трансляции
- 65. Инициация – начало трансляции Рибосома соединяется с иРНК и захватывает два кодона (первый – инициальный -оказывается
- 66. Элонгация – синтез полипептида. Ко второму кодону иРНК подходит вторая тРНК с аминокислотой. Если антикодон тРНК
- 67. Затем первая тРНК выходит из рибосомы, рибосома перемещается на один триплет вперед. К этому триплету подходит
- 68. Терминация трансляции В аминоацильном центре оказывается нонсенс – кодон (UAG, UAA, UGA) для которого нет соответствующей
- 69. Терминация транскрипции –окончание. Рибосома доходит до стоп-кодона. Синтез полипептида останавливается. Стоп-кодон Фолдинг
- 70. иРНК Растущий полипетид старт Полисома, или полирибосома — несколько рибосом, одновременно транслирующих одну молекулу мРНК.
- 71. Значение для медицины Знание белковых продуктов различных генов позволяет успешно лечить многие болезни обмена посредством введения
- 72. Ингибиторы трансляции Стрептомицин – препятствует связыванию формилметионин- т РНК с рибосомой, нарушая инициацию трансляции. Связывается с
- 73. Действие токсинов Аманитин (токсин бледной поганки), циклический пептид, связывается с эукариотической РНК-полимеразой II, блокируя синтез м
- 75. БОНУС
- 76. Опухолевые вирусы и онкогены Некоторые опухолевые вирусы не содержат онкоген, но, встраиваясь в хромосому рядом с
- 77. Превращение клеточных протоонкогенов Превращение клеточных протоонкогенов в онкогены может происходить в результате и в результате повышения
- 78. Онкогенные вирусы Установлено, что гены и даже целые участки хромосом высших организмов могут иногда перемещаться с
- 79. Ретровирусы Как показали исследования, во многих зрелых онкогенных РНК-содержащих вирусах (ретровирусы) и в том числе в
- 80. Онкогены По фенотипическим проявлениям различают две группы онкогенов. Одна группа - ядерные (иммортелизующие) онкогены, приводящие к
- 81. Канцерогенез Некоторые опухолевые вирусы не содержат онкоген, но, встраиваясь в хромосому рядом с протоонкогеном, активируют его,
- 82. Протеин р53 В последние годы найдено еще одно, по-видимому, наиболее общее звено канцерогенеза - гены-супрессоры опухолей,
- 84. Скачать презентацию

Молекулярные основы наследственности.
1. Центральная догма молекулярной биологии.
Типы переноса генетической информации
Молекулярные основы наследственности.
1. Центральная догма молекулярной биологии.
Типы переноса генетической информации
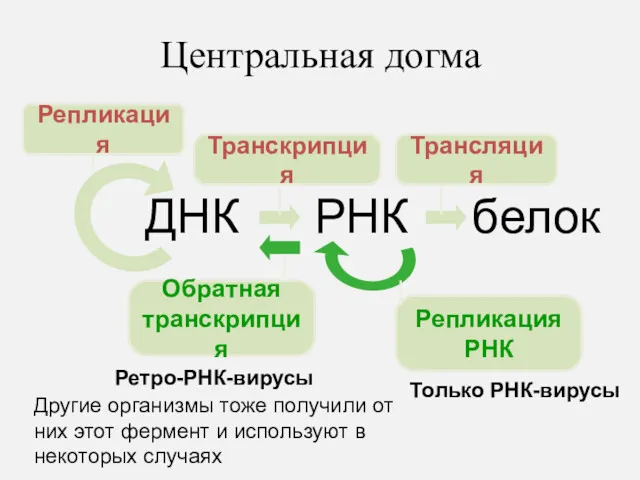
Центральная догма
ДНК
РНК
белок
Репликация
Транскрипция
Трансляция
Обратная
транскрипция
Репликация
РНК
Только РНК-вирусы
Ретро-РНК-вирусы
Другие организмы тоже получили от них этот фермент и
Центральная догма
ДНК
РНК
белок
Репликация
Транскрипция
Трансляция
Обратная
транскрипция
Репликация
РНК
Только РНК-вирусы
Ретро-РНК-вирусы
Другие организмы тоже получили от них этот фермент и
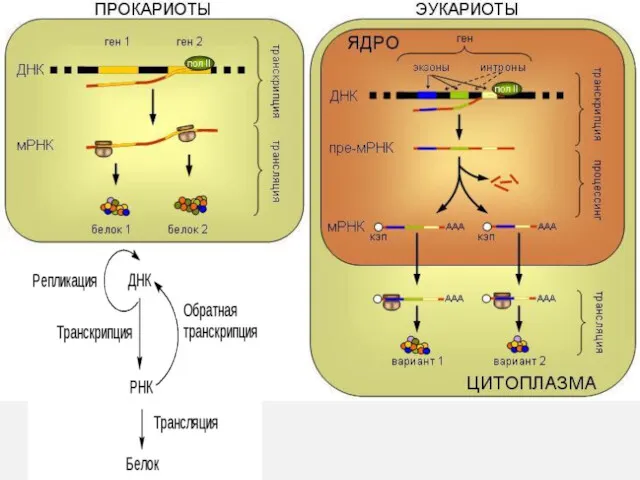
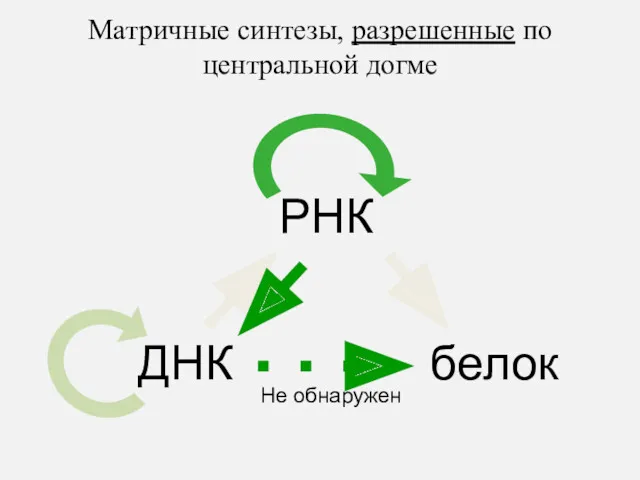
ДНК
РНК
белок
Матричные синтезы, разрешенные по центральной догме
Не обнаружен
ДНК
РНК
белок
Матричные синтезы, разрешенные по центральной догме
Не обнаружен
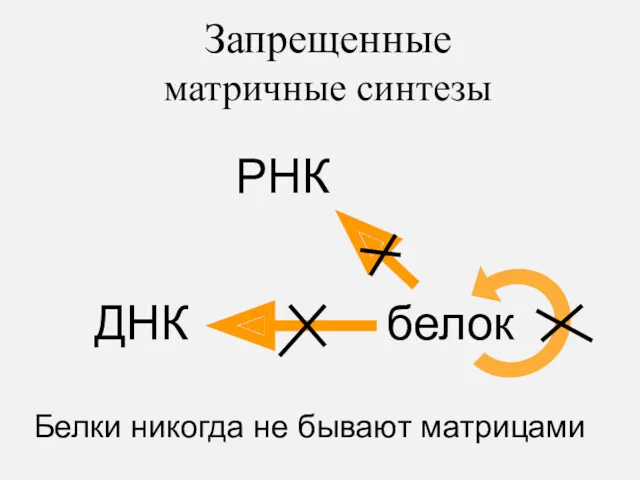
Запрещенные
матричные синтезы
Белки никогда не бывают матрицами
Запрещенные
матричные синтезы
Белки никогда не бывают матрицами
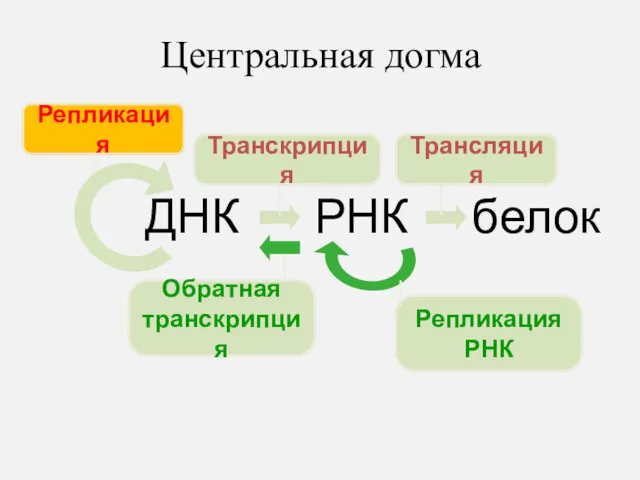
Центральная догма
ДНК
РНК
белок
Репликация
Транскрипция
Трансляция
Обратная
транскрипция
Репликация
РНК
Центральная догма
ДНК
РНК
белок
Репликация
Транскрипция
Трансляция
Обратная
транскрипция
Репликация
РНК

2. Репликация ДНК
Универсальный биологический процесс передачи генетической информации в поколениях клеток
2. Репликация ДНК
Универсальный биологический процесс передачи генетической информации в поколениях клеток
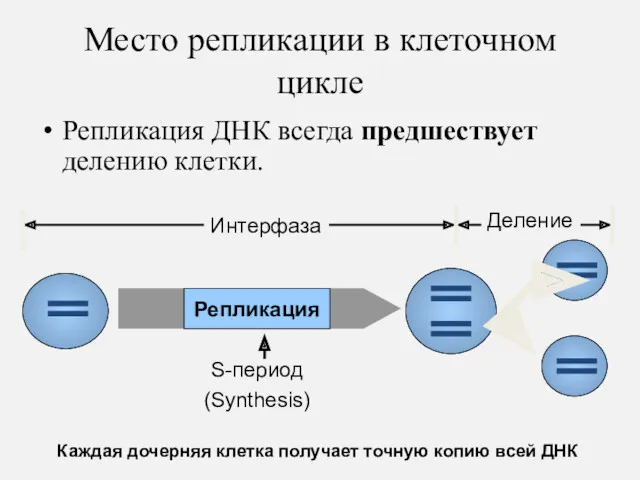
Место репликации в клеточном цикле
Репликация ДНК всегда предшествует делению клетки.
Репликация
S-период
(Synthesis)
Интерфаза
Деление
Каждая дочерняя
Место репликации в клеточном цикле
Репликация ДНК всегда предшествует делению клетки.
Репликация
S-период
(Synthesis)
Интерфаза
Деление
Каждая дочерняя

Принципы репликации
1. Полуконсервативность
2. Комплементарность
3. Антипараллельность
4. Униполярность
5. Прерывистость
Принципы репликации
1. Полуконсервативность
2. Комплементарность
3. Антипараллельность
4. Униполярность
5. Прерывистость
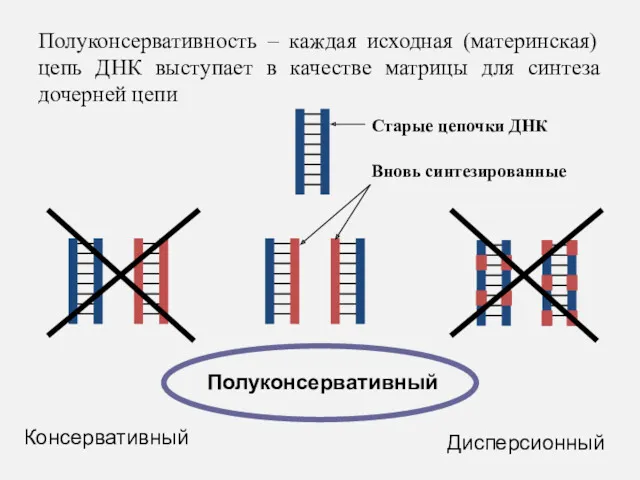
Полуконсервативность – каждая исходная (материнская) цепь ДНК выступает в качестве матрицы
Полуконсервативность – каждая исходная (материнская) цепь ДНК выступает в качестве матрицы
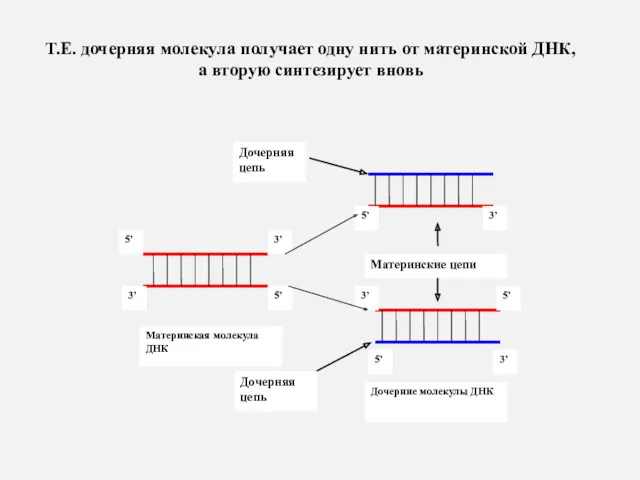
Т.Е. дочерняя молекула получает одну нить от материнской ДНК,
а вторую
Т.Е. дочерняя молекула получает одну нить от материнской ДНК, а вторую

Комплементарность
Вновь синтезируемая ( дочерняя) цепь ДНК строится по принципу комплементарности. В
Комплементарность
Вновь синтезируемая ( дочерняя) цепь ДНК строится по принципу комплементарности. В

Антипараллельность – синтез дочерней цепи ДНК происходит в противоположном от материнской
Антипараллельность – синтез дочерней цепи ДНК происходит в противоположном от материнской

Униполярность:
Удвоение цепи ДНК идет в направлении от 5` конца к 3`
Униполярность:
Удвоение цепи ДНК идет в направлении от 5` конца к 3`
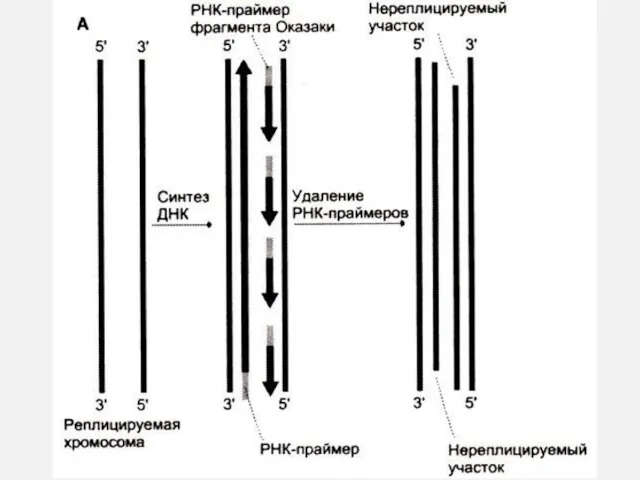
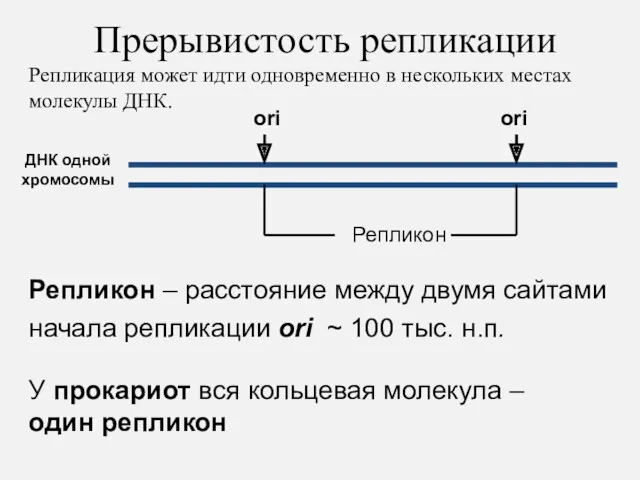
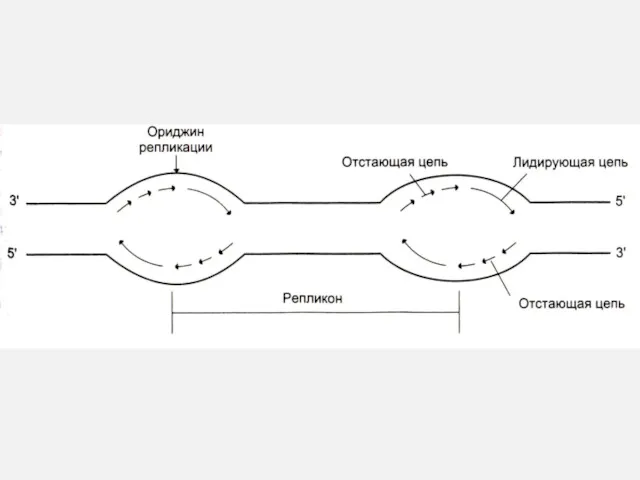
Репликон – расстояние между двумя сайтами начала репликации ori ~ 100
Репликон – расстояние между двумя сайтами начала репликации ori ~ 100
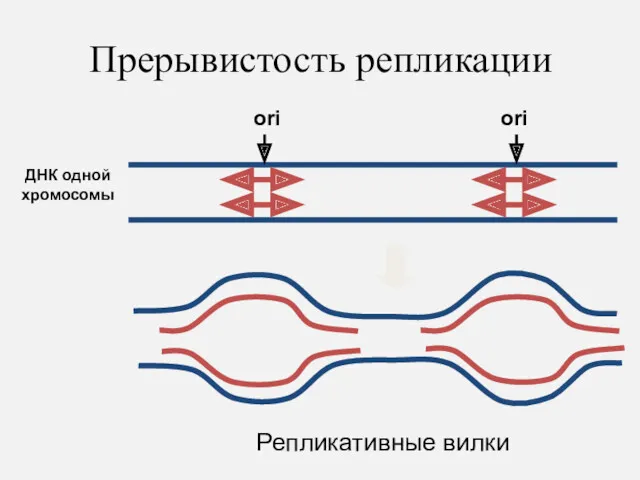
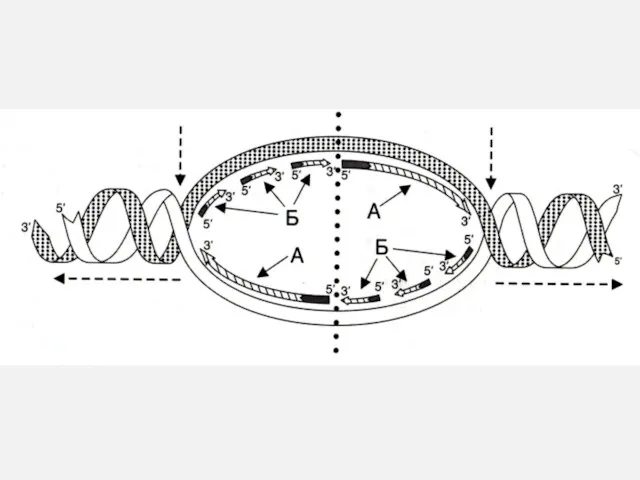
Прерывистость репликации
ДНК одной хромосомы
ori
ori
Репликативные вилки
Прерывистость репликации
ДНК одной хромосомы
ori
ori
Репликативные вилки
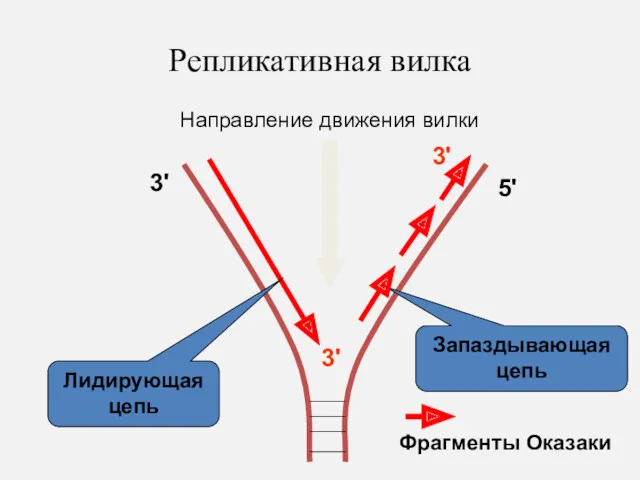
Репликативная вилка
3'
5'
3'
3'
Запаздывающая цепь
Лидирующая цепь
Направление движения вилки
Фрагменты Оказаки
Репликативная вилка
3'
5'
3'
3'
Запаздывающая цепь
Лидирующая цепь
Направление движения вилки
Фрагменты Оказаки

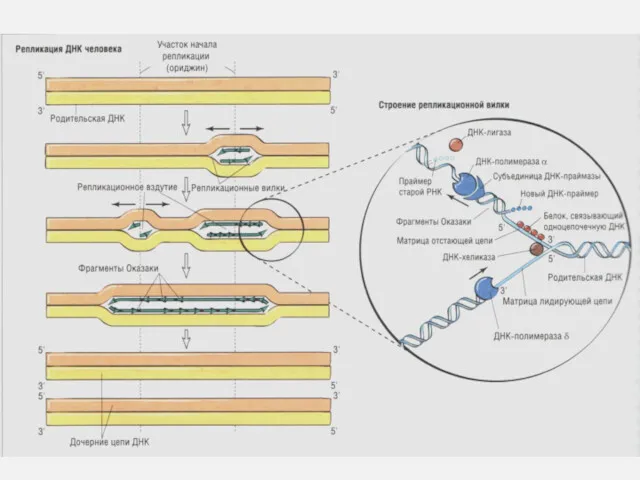
Репликация ДНК
Скорость репликации огромна, т.к. реакция идет в нескольких местах одновременно
Репликация ДНК
Скорость репликации огромна, т.к. реакция идет в нескольких местах одновременно

ДНК ТОПОИЗОМЕРАЗА – фермент, изменяющий степень сверхспиральности, возникающее при раскручивании двух
ДНК ТОПОИЗОМЕРАЗА – фермент, изменяющий степень сверхспиральности, возникающее при раскручивании двух

Репликация ДНК
Количество раундов репликации ДНК (а значит число возможных делений клетки)
Репликация ДНК
Количество раундов репликации ДНК (а значит число возможных делений клетки)

Репликация ДНК
ДНК- полимеразы Δ и ε делают 1 ошибку на 105
Репликация ДНК
ДНК- полимеразы Δ и ε делают 1 ошибку на 105
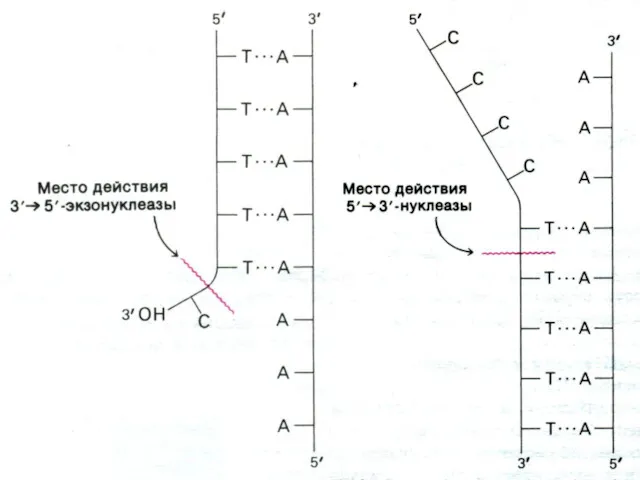

Репликация ДНК
Ошибки в ДНК (мутации) возникают спонтанно (ошибки репликации, дезаминирование нуклеотидов,
Репликация ДНК
Ошибки в ДНК (мутации) возникают спонтанно (ошибки репликации, дезаминирование нуклеотидов,

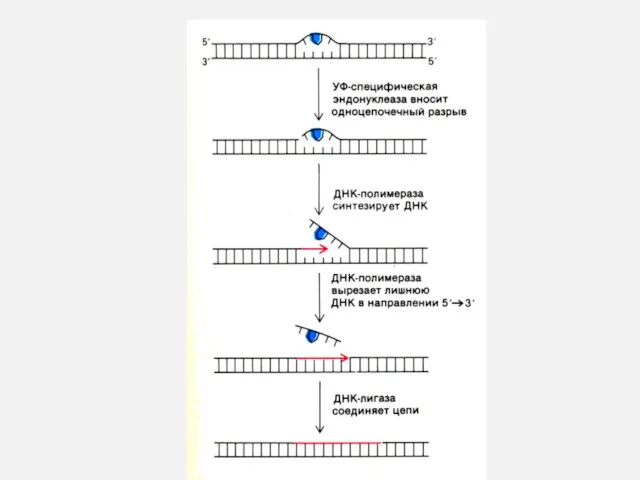
Репликация ДНК
Комплекс ферментов репарации узнает и вырезает поврежденные и химически измененные
Репликация ДНК
Комплекс ферментов репарации узнает и вырезает поврежденные и химически измененные

Значение для медицины
Иногда в растующую цепь случайно вклинивается неправильное основание,
Значение для медицины
Иногда в растующую цепь случайно вклинивается неправильное основание,

Выводы по репликации ДНК
В результате репликации каждая дочерняя клетка получает точную
Выводы по репликации ДНК
В результате репликации каждая дочерняя клетка получает точную
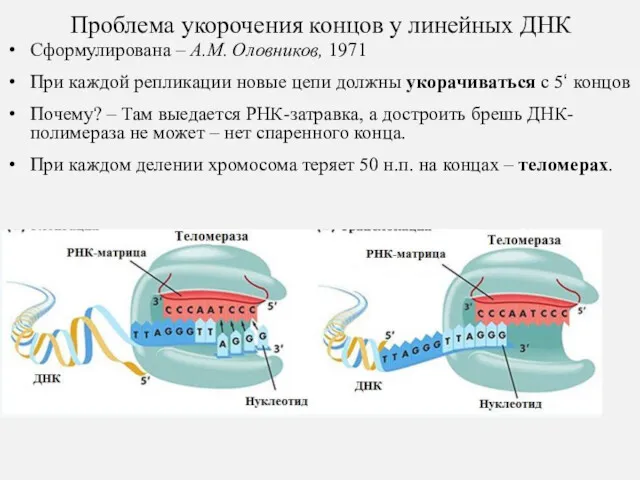
Проблема укорочения концов у линейных ДНК
Сформулирована – А.М. Оловников, 1971
При каждой
Проблема укорочения концов у линейных ДНК
Сформулирована – А.М. Оловников, 1971
При каждой
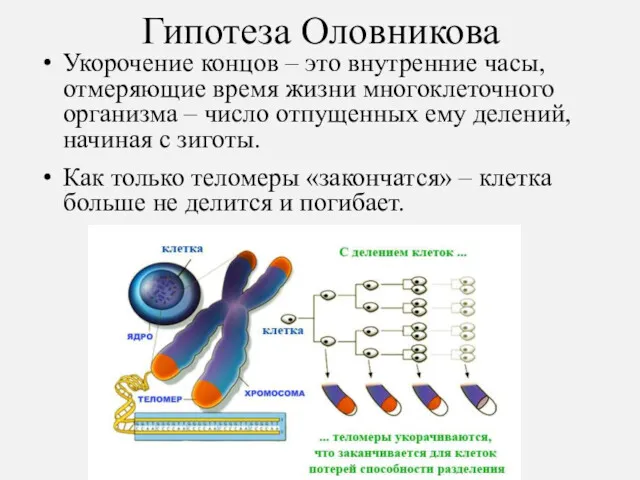
Гипотеза Оловникова
Укорочение концов – это внутренние часы, отмеряющие время жизни многоклеточного
Гипотеза Оловникова
Укорочение концов – это внутренние часы, отмеряющие время жизни многоклеточного

Но почему тогда клетки зародышевой линии делятся бесконечно?
Оловников: должен существовать механизм
Но почему тогда клетки зародышевой линии делятся бесконечно?
Оловников: должен существовать механизм

Теломераза
фермент, надстраивающий концы хромосом, содержит РНК.
удлинение происходит путем
обратной транскрипции:
РНК →
Теломераза
фермент, надстраивающий концы хромосом, содержит РНК.
удлинение происходит путем
обратной транскрипции:
РНК →

Теломераза активна в клетках
зародышевого пути
эмбриональных
стволовых
раковых –
Теломераза активна в клетках
зародышевого пути
эмбриональных
стволовых
раковых –

Значение для медицины
После каждого клеточного цикла теломеры укорачиваются на один повтор,
Значение для медицины
После каждого клеточного цикла теломеры укорачиваются на один повтор,

4.Транскрипция.
Механизмы транскрипции у про- и эукариот.
Транскрипция - это первый этап
4.Транскрипция.
Механизмы транскрипции у про- и эукариот.
Транскрипция - это первый этап
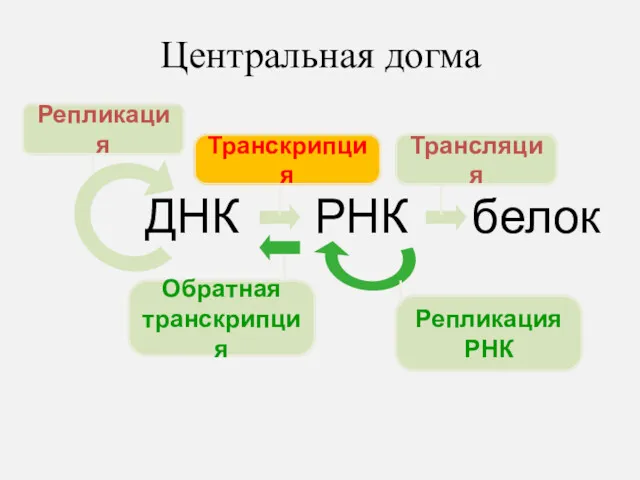
Центральная догма
ДНК
РНК
белок
Репликация
Транскрипция
Трансляция
Обратная
транскрипция
Репликация
РНК
Центральная догма
ДНК
РНК
белок
Репликация
Транскрипция
Трансляция
Обратная
транскрипция
Репликация
РНК

Транскрипция
Считывание информации с ДНК-матрицы на РНК, синтез тРНК, иРНК, рРНК с
Транскрипция
Считывание информации с ДНК-матрицы на РНК, синтез тРНК, иРНК, рРНК с

Транскрипция
Механизм РНК – полимеразной реакции тот же, что и ДНК –
Транскрипция
Механизм РНК – полимеразной реакции тот же, что и ДНК –

Транскрипция
В ДНК – матрице выделяют транскиптоны. Участки, ограниченные промоторами и сайтами
Транскрипция
В ДНК – матрице выделяют транскиптоны. Участки, ограниченные промоторами и сайтами
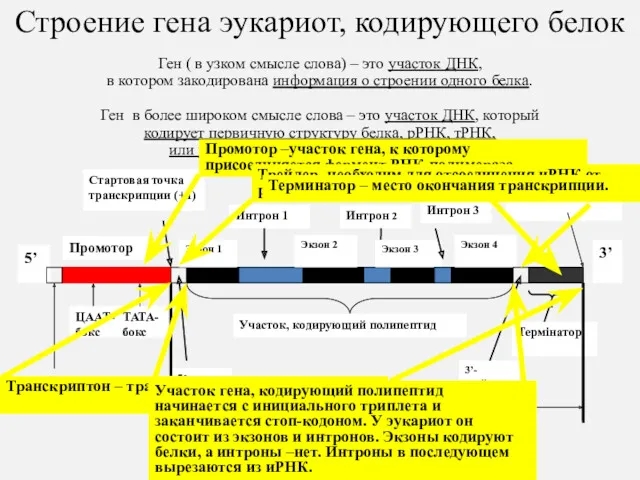
Строение гена эукариот, кодирующего белок
Ген ( в узком смысле слова)
Строение гена эукариот, кодирующего белок Ген ( в узком смысле слова)

Транскрипция
3 стадии транскрипции: инициация, элонгация и терминация.
Инициация синтеза начинается с «узнавания»
Транскрипция
3 стадии транскрипции: инициация, элонгация и терминация.
Инициация синтеза начинается с «узнавания»

Инициация транскрипции
Для формирование транскрипционной вилки (раскручивание одного витка спирали ДНК-матрицы) к
Инициация транскрипции
Для формирование транскрипционной вилки (раскручивание одного витка спирали ДНК-матрицы) к

Элонгация транскрипции
Белковые факторы элонгации обеспечивают расплетение ДНК перед продвижением РНК-полимеразы и
Элонгация транскрипции
Белковые факторы элонгации обеспечивают расплетение ДНК перед продвижением РНК-полимеразы и

Терминация транскрипции
При достижении РНК - полимеразой сайта терминации белковый фактор терминации
Терминация транскрипции
При достижении РНК - полимеразой сайта терминации белковый фактор терминации

Особенности транскрипции у эукариот
Транскрипция
1. Кэп и поли-А-хвост
Созревание м-РНК
2.Сплайсинг
цитоплазма
Особенности транскрипции у эукариот
Транскрипция
1. Кэп и поли-А-хвост
Созревание м-РНК
2.Сплайсинг
цитоплазма
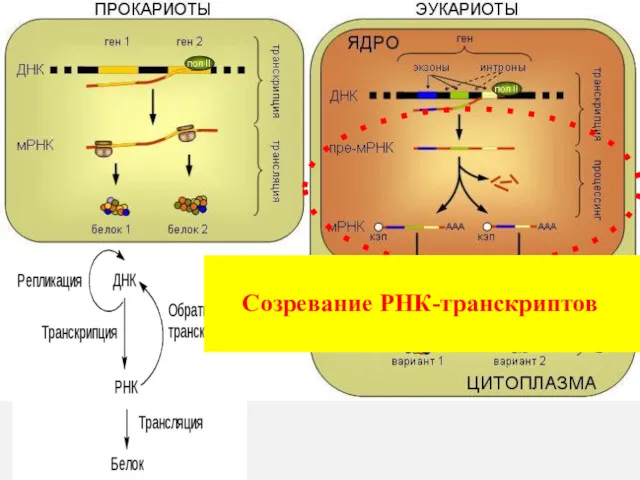
Созревание РНК-транскриптов
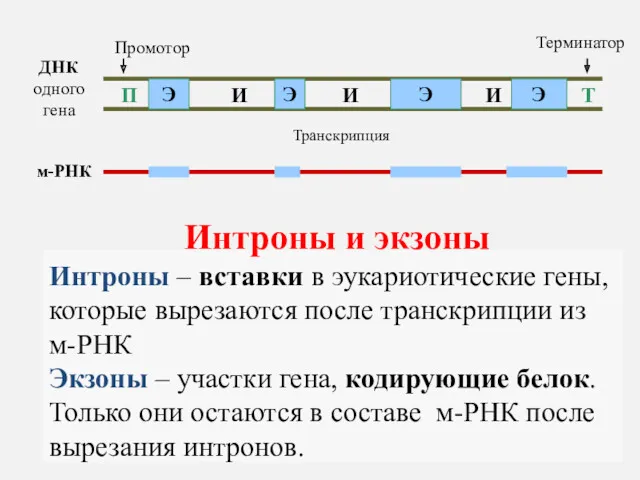
ДНК одного гена
П
Т
Промотор
Терминатор
Интроны и экзоны
Интроны – вставки в эукариотические гены,
ДНК одного гена
П
Т
Промотор
Терминатор
Интроны и экзоны
Интроны – вставки в эукариотические гены,
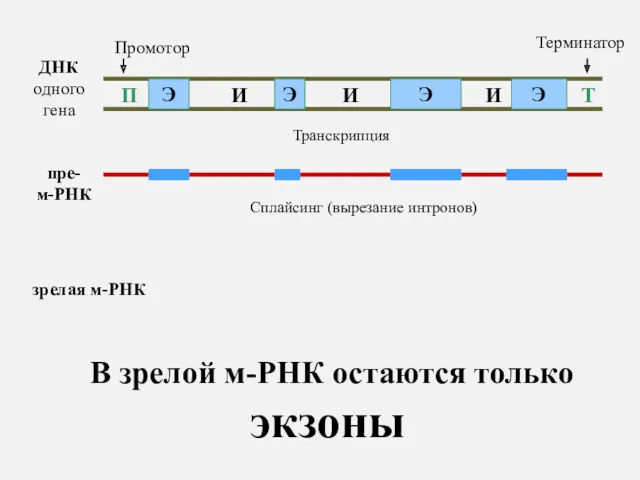
ДНК одного гена
П
Т
Промотор
Терминатор
пре-
м-РНК
Транскрипция
Сплайсинг (вырезание интронов)
зрелая м-РНК
В зрелой м-РНК остаются только
ДНК одного гена
П
Т
Промотор
Терминатор
пре-
м-РНК
Транскрипция
Сплайсинг (вырезание интронов)
зрелая м-РНК
В зрелой м-РНК остаются только
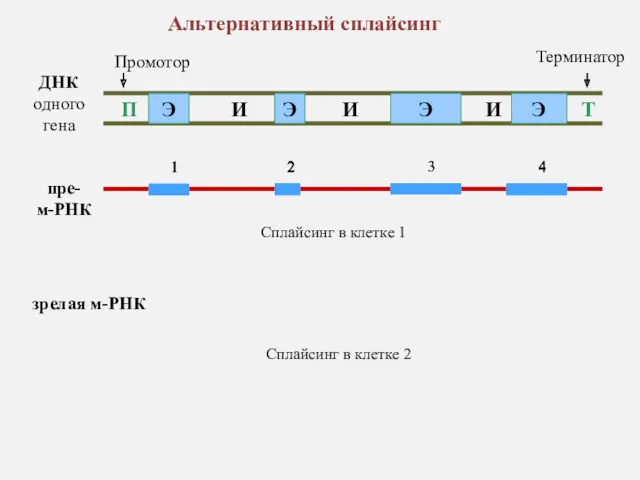
ДНК одного гена
П
Т
Промотор
Терминатор
пре-
м-РНК
Сплайсинг в клетке 1
зрелая м-РНК
Сплайсинг в клетке 2
Альтернативный сплайсинг
ДНК одного гена
П
Т
Промотор
Терминатор
пре-
м-РНК
Сплайсинг в клетке 1
зрелая м-РНК
Сплайсинг в клетке 2
Альтернативный сплайсинг

Альтернативный сплайсинг
в разных органах
на разных стадиях развития
в разных
Альтернативный сплайсинг
в разных органах
на разных стадиях развития
в разных

Значение для медицины
Иногда в некоторых транскриптах обнаруживают альтернативные механизмы сплайсинга, однако
Значение для медицины
Иногда в некоторых транскриптах обнаруживают альтернативные механизмы сплайсинга, однако

Созревание РНК-транскриптов
Процессингу (созреванию) подвергаются все виды РНК (и, т, р).
А) Ковалентная
Созревание РНК-транскриптов
Процессингу (созреванию) подвергаются все виды РНК (и, т, р).
А) Ковалентная

Ковалентная модификация иРНК
Гуанилил-трансфераза присоединяет ГДФ к 5- ОР концу (5-О-Р-О-5 связь),
5
Ковалентная модификация иРНК
Гуанилил-трансфераза присоединяет ГДФ к 5- ОР концу (5-О-Р-О-5 связь),
5

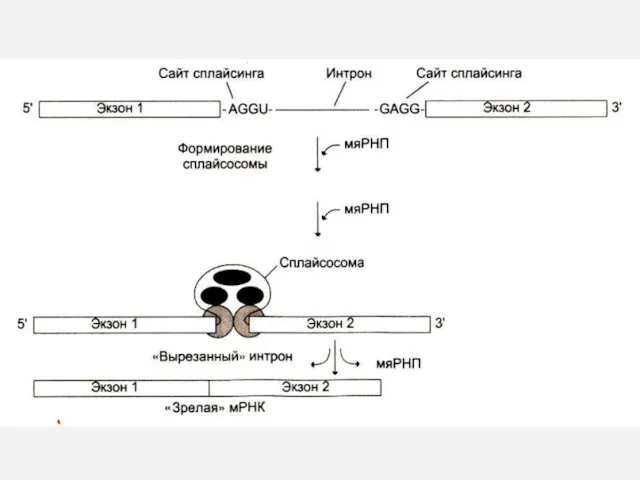
СПЛАЙСИНГ иРНК
Сплайсинг: образование зрелой мРНК:
Вырезание интронных последовательностей (ограниченных AGGU- и -
СПЛАЙСИНГ иРНК
Сплайсинг: образование зрелой мРНК:
Вырезание интронных последовательностей (ограниченных AGGU- и -
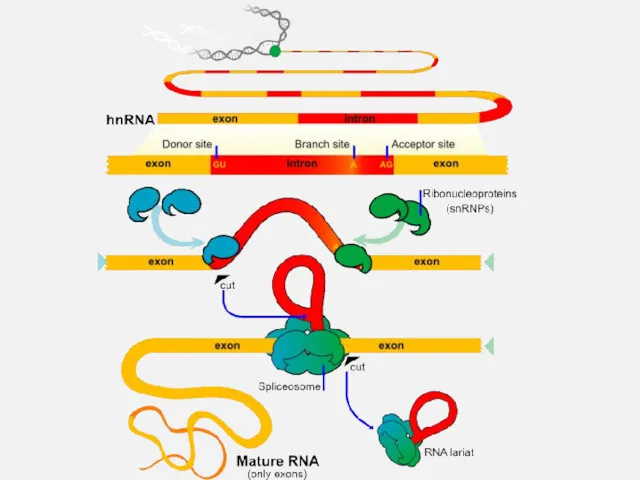
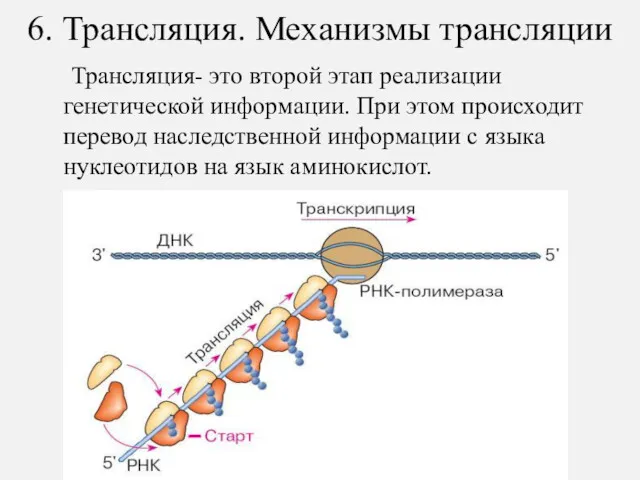
6. Трансляция. Механизмы трансляции
Трансляция- это второй этап реализации генетической информации.
6. Трансляция. Механизмы трансляции
Трансляция- это второй этап реализации генетической информации.
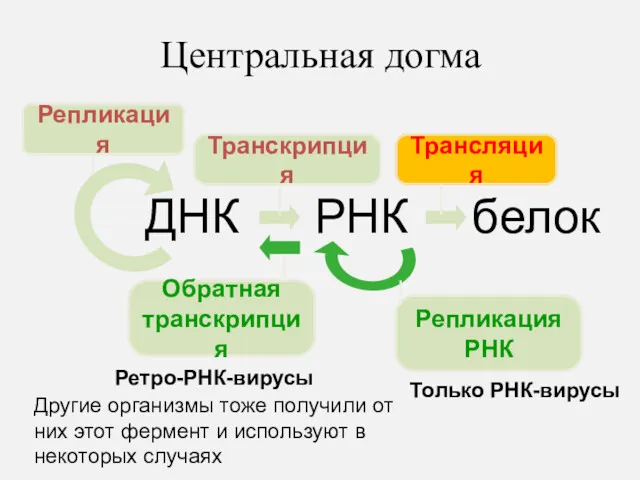
Центральная догма
ДНК
РНК
белок
Репликация
Транскрипция
Трансляция
Обратная
транскрипция
Репликация
РНК
Только РНК-вирусы
Ретро-РНК-вирусы
Другие организмы тоже получили от них этот фермент и
Центральная догма
ДНК
РНК
белок
Репликация
Транскрипция
Трансляция
Обратная
транскрипция
Репликация
РНК
Только РНК-вирусы
Ретро-РНК-вирусы
Другие организмы тоже получили от них этот фермент и
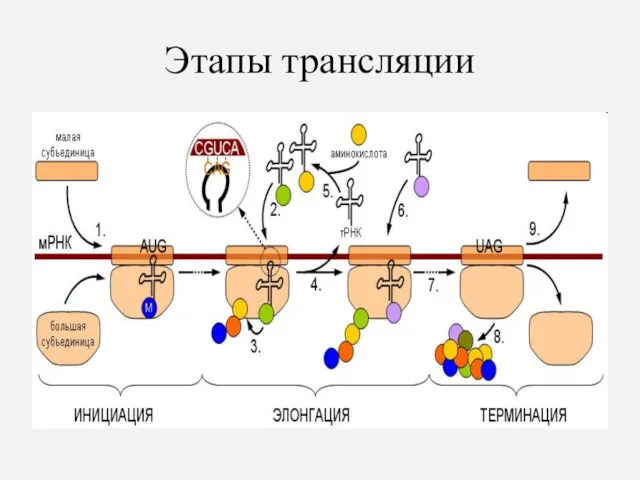
Этапы трансляции
Этапы трансляции

Инициация – начало трансляции
Рибосома соединяется с иРНК и захватывает два кодона
Инициация – начало трансляции
Рибосома соединяется с иРНК и захватывает два кодона
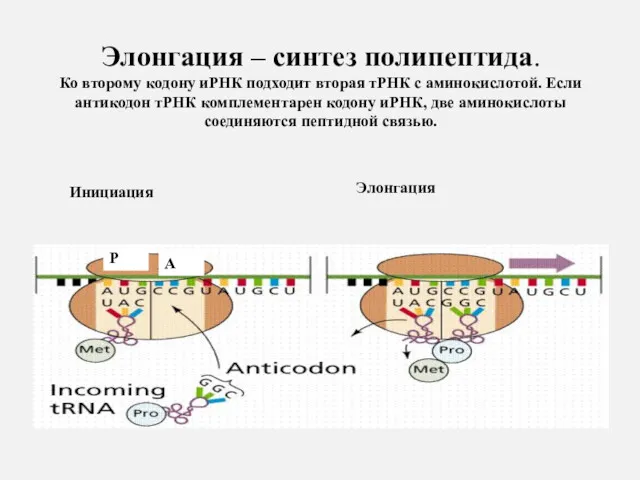
Элонгация – синтез полипептида.
Ко второму кодону иРНК подходит вторая тРНК
Элонгация – синтез полипептида. Ко второму кодону иРНК подходит вторая тРНК
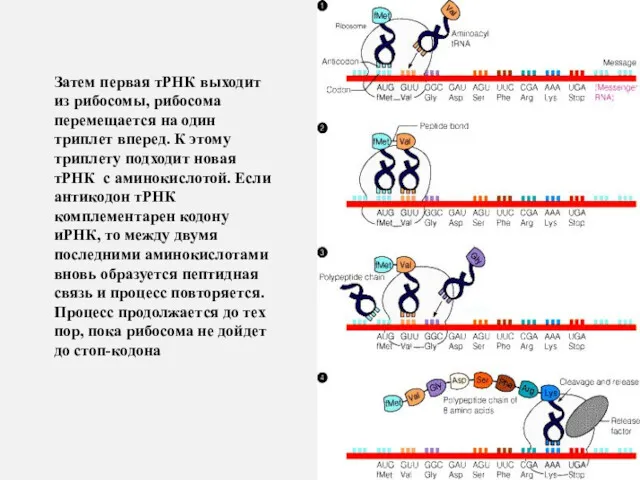
Затем первая тРНК выходит из рибосомы, рибосома перемещается на один триплет
Затем первая тРНК выходит из рибосомы, рибосома перемещается на один триплет

Терминация трансляции
В аминоацильном центре оказывается нонсенс – кодон (UAG, UAA, UGA)
Терминация трансляции
В аминоацильном центре оказывается нонсенс – кодон (UAG, UAA, UGA)
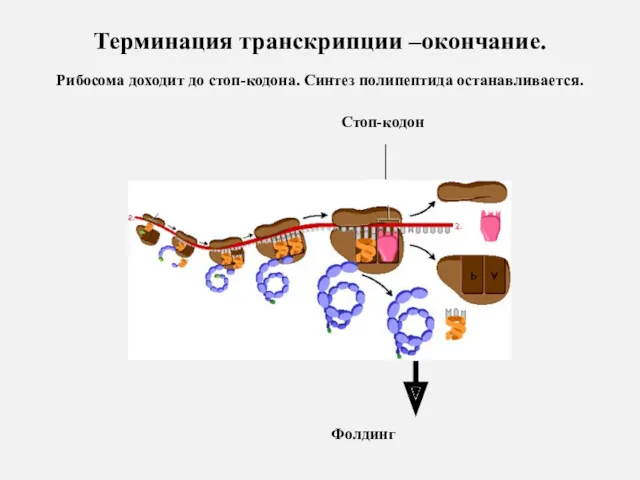
Терминация транскрипции –окончание.
Рибосома доходит до стоп-кодона. Синтез полипептида останавливается.
Стоп-кодон
Фолдинг
Терминация транскрипции –окончание.
Рибосома доходит до стоп-кодона. Синтез полипептида останавливается.
Стоп-кодон
Фолдинг
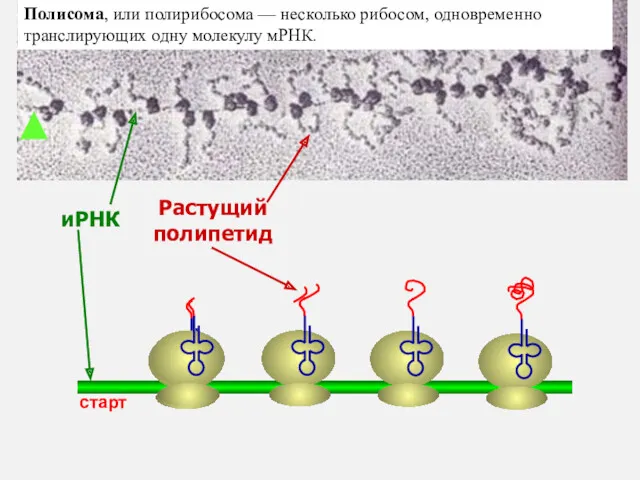
иРНК
Растущий
полипетид
старт
Полисома, или полирибосома — несколько рибосом, одновременно транслирующих одну молекулу мРНК.
иРНК
Растущий
полипетид
старт
Полисома, или полирибосома — несколько рибосом, одновременно транслирующих одну молекулу мРНК.

Значение для медицины
Знание белковых продуктов различных генов позволяет успешно лечить
Значение для медицины
Знание белковых продуктов различных генов позволяет успешно лечить

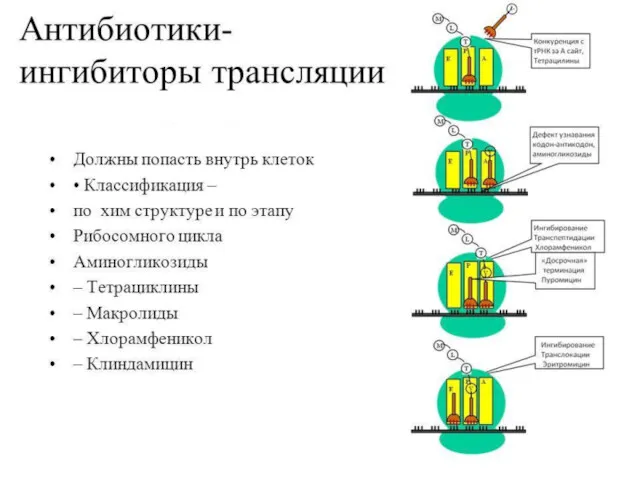
Ингибиторы трансляции
Стрептомицин – препятствует связыванию формилметионин- т РНК с рибосомой, нарушая
Ингибиторы трансляции
Стрептомицин – препятствует связыванию формилметионин- т РНК с рибосомой, нарушая

Действие токсинов
Аманитин (токсин бледной поганки), циклический пептид, связывается с эукариотической РНК-полимеразой
Действие токсинов
Аманитин (токсин бледной поганки), циклический пептид, связывается с эукариотической РНК-полимеразой

БОНУС
БОНУС

Опухолевые вирусы и онкогены
Некоторые опухолевые вирусы не содержат онкоген, но, встраиваясь
Опухолевые вирусы и онкогены
Некоторые опухолевые вирусы не содержат онкоген, но, встраиваясь

Превращение клеточных протоонкогенов
Превращение клеточных протоонкогенов в онкогены может происходить в результате
Превращение клеточных протоонкогенов
Превращение клеточных протоонкогенов в онкогены может происходить в результате

Онкогенные вирусы
Установлено, что гены и даже целые участки хромосом высших организмов
Онкогенные вирусы
Установлено, что гены и даже целые участки хромосом высших организмов

Ретровирусы
Как показали исследования, во многих зрелых онкогенных РНК-содержащих вирусах (ретровирусы) и
Ретровирусы
Как показали исследования, во многих зрелых онкогенных РНК-содержащих вирусах (ретровирусы) и

Онкогены
По фенотипическим проявлениям различают две группы онкогенов.
Одна группа - ядерные
Онкогены
По фенотипическим проявлениям различают две группы онкогенов.
Одна группа - ядерные

Канцерогенез
Некоторые опухолевые вирусы не содержат онкоген, но, встраиваясь в хромосому рядом
Канцерогенез
Некоторые опухолевые вирусы не содержат онкоген, но, встраиваясь в хромосому рядом

Протеин р53
В последние годы найдено еще одно, по-видимому, наиболее общее
Протеин р53
В последние годы найдено еще одно, по-видимому, наиболее общее
 Самостоятельная творческая работа по теме РНК- рибонуклеиновая кислота
Самостоятельная творческая работа по теме РНК- рибонуклеиновая кислота Популяционно-статистические методы
Популяционно-статистические методы Гетерозис ғылымының теориясы мен болжамы
Гетерозис ғылымының теориясы мен болжамы Скелет конечностей позвоночных. Эволюция и общие закономерности
Скелет конечностей позвоночных. Эволюция и общие закономерности Загадка человека Урок 2
Загадка человека Урок 2 Строение семян однодольных и двудольных растений. Морфология проростка. Прорастание семян
Строение семян однодольных и двудольных растений. Морфология проростка. Прорастание семян Полевые цветы
Полевые цветы The Chemical Context of Life
The Chemical Context of Life Физиология высшей нервной деятельности. Формирование поведения в онтогенезе
Физиология высшей нервной деятельности. Формирование поведения в онтогенезе Естественнонаучные предпосылки возникновения учения Ч. Дарвина
Естественнонаучные предпосылки возникновения учения Ч. Дарвина Использование современного оборудования в образовательном процессе
Использование современного оборудования в образовательном процессе Ароматерапия. Действие эфирных масел
Ароматерапия. Действие эфирных масел Кровоснабжение стенок и органов полостей
Кровоснабжение стенок и органов полостей Магний и его роль в орпганизме.
Магний и его роль в орпганизме. Черепные нервы. Лекция №26
Черепные нервы. Лекция №26 Кроссворд по биологии по теме: Антропогенез
Кроссворд по биологии по теме: Антропогенез Сүйектің байланысу түрлері
Сүйектің байланысу түрлері Байкал – фабрика чистой воды. Приложение 4
Байкал – фабрика чистой воды. Приложение 4 Приспособленность организмов и ее относительность
Приспособленность организмов и ее относительность Схемы. Онтогенез и регуляция
Схемы. Онтогенез и регуляция Ядро клетки
Ядро клетки Wonderful wild animals
Wonderful wild animals Будова тварин. Клітини, тканини
Будова тварин. Клітини, тканини Факторы эволюции человека. Человеческие расы
Факторы эволюции человека. Человеческие расы Структурно-функциональные особенности цитоскелета и его производных
Структурно-функциональные особенности цитоскелета и его производных Флора и фауна луга
Флора и фауна луга Тип черви
Тип черви Высшая нервная деятельность. Лекция № 28
Высшая нервная деятельность. Лекция № 28