Lecture presented on the 4th International Plant Protection Symposium at Debrecen University презентация
- Lecture presented on the 4th International Plant Protection Symposium at Debrecen University

Содержание
- 2. First data on the sibling species of the common green lacewings in Spain (Neuroptera: Chrysopidae) A.
- 3. Summary Common green lacewings are good candidates for use in IPM programs because they are distributed
- 4. The taxonomic status of the most important lacewing species in Andaluzia
- 5. Introduction Chrysopids and among them the common green lacewings are not only attractive and wonderful insects
- 6. Systematic troubles The original name of the species was Chrysopa carnea. However, because of taxonomical accuracy
- 7. Attempts for clearing the taxonomic status of Chrysoperla carnea s.l.
- 8. Courtship sonification (Henry, 1994; Henry et al., 2002) Silent inaudible sing: the male places himself in
- 9. Weak points of courtship sonification nobody could verify the method except Henry and colleagues the methodology
- 10. Genetic studies with multilocus electrophoresis and molecular systematics Regarding multilocus electrophoresis there are differences but variation
- 11. Morphological characterization of adults (Thierry et al., 1992) colour of ventral setae on the distal portion
- 12. Morphological characterization of adults (Henry et al., 2002) fine structure of the male distal abdomen Weak
- 13. Ecological differences (Thierry et al., (1994), Henry et al., (2002) Ch. carnea is an arboreal species
- 14. Ecophysiological variability Thierry et al. (1994) Overwintering places Ch. carnea hibernates in rolled dry leaves and
- 15. AChE tolerance (Bozsik et al., 2002 unpublished) Variation in tolerance level can be important. Further work
- 16. The different evidences supported the existence of various cryptic species Ch. carnea former Chrysoperla kolthoffi (Navás,
- 17. Oscillographs of some European Chrysoperla spp. Source: http://www.eeb.uconn.edu/people/chenry/Cryptic_songs.html The Cryptic Song Species of Chrysoperla Charles S.
- 18. Oscillograph of Ch. affinis (Cc4, motorboat)
- 19. Oscillograph of Ch. carnea s.str. (Cc2, slow-motorboat)
- 20. Oscillograph of Ch. lucasina
- 21. Oscillograph of Ch. agilis (Cc3, Maltese)
- 22. Morphology of Chrysoperla spp.
- 23. Ch. affinis
- 24. Ch. affinis (female)
- 25. Ch. affinis
- 26. Ch. carnea s.str.
- 27. Ch. carnea s.str.
- 28. Ch. carnea s.str.
- 29. Ch. lucasina
- 30. Ch. lucasina
- 31. Ch. lucasina
- 32. Ch. lucasina (female)
- 33. Ch. lucasina
- 34. Practical troubles One should not forget the natural enemy role of Ch. carnea, which is used
- 35. A practical example of the study of Ch. carnea complex Management of the most important natural
- 36. Natural control for olive moth Olive moth (Prays oleae (Bernard) is one of the most important
- 37. Questions Which sibling species is the really efficient taxon? In some years when the density of
- 38. Future tasks a. - identification of the lacewing species (sibling species) controlling olive moth caterpillars, -
- 39. Basic data of collection in southern Spain
- 40. A typical olive grove landscape (Photo by R. González Ruiz)
- 41. Olive trees (Photo by R. González Ruiz)
- 42. Identification Individuals preserved in ethanol were identified according to the descriptions of Thierry et al. (1992),
- 43. Number and proportion of sibling species of common green lacewings in Andaluzia
- 44. Collection sites of Ch. agilis (on the data of Henry et al., 2003) (No data =
- 45. Conclusions Ch. agilis is the dominant species in Southern Spain and the olive groves as well.
- 46. By biologic control and IPM we can save landscapes like that
- 47. References Al-Asaad, D. S. (2004): Viabilidad de los chrisópidos (Neuroptera: Chrysopidae) del olivar. Influencia del entorno
- 48. Henry, Ch. S., Brooks, S.J., Thierry, D., Duelli, P. & Johnson, J.B. (2001): The common green
- 50. Скачать презентацию

First data on the sibling species of the common green lacewings
First data on the sibling species of the common green lacewings

Summary
Common green lacewings are good candidates for use in IPM programs
Summary
Common green lacewings are good candidates for use in IPM programs

The taxonomic status of the most important lacewing species in Andaluzia
The taxonomic status of the most important lacewing species in Andaluzia

Introduction
Chrysopids and among them the common green lacewings are not
Introduction
Chrysopids and among them the common green lacewings are not

Systematic troubles
The original name of the species was Chrysopa carnea.
Systematic troubles
The original name of the species was Chrysopa carnea.

Attempts for clearing the taxonomic status of Chrysoperla carnea s.l.
Attempts for clearing the taxonomic status of Chrysoperla carnea s.l.

Courtship sonification (Henry, 1994; Henry et al., 2002)
Silent inaudible sing:
Courtship sonification (Henry, 1994; Henry et al., 2002)
Silent inaudible sing:

Weak points of courtship sonification
nobody could verify the method except Henry
Weak points of courtship sonification
nobody could verify the method except Henry

Genetic studies with multilocus electrophoresis
and molecular systematics
Regarding multilocus electrophoresis
Genetic studies with multilocus electrophoresis
and molecular systematics
Regarding multilocus electrophoresis

Morphological characterization of adults (Thierry et al., 1992)
colour of ventral setae
Morphological characterization of adults (Thierry et al., 1992)
colour of ventral setae

Morphological characterization of adults (Henry et al., 2002)
fine structure of
Morphological characterization of adults (Henry et al., 2002)
fine structure of

Ecological differences (Thierry et al., (1994), Henry et al., (2002)
Ch. carnea
Ecological differences (Thierry et al., (1994), Henry et al., (2002)
Ch. carnea

Ecophysiological variability Thierry et al. (1994)
Overwintering places
Ch. carnea hibernates in rolled
Ecophysiological variability Thierry et al. (1994)
Overwintering places
Ch. carnea hibernates in rolled

AChE tolerance (Bozsik et al., 2002 unpublished)
Variation in tolerance level can
AChE tolerance (Bozsik et al., 2002 unpublished)
Variation in tolerance level can

The different evidences supported the existence of various cryptic species
Ch.
The different evidences supported the existence of various cryptic species
Ch.

Oscillographs of some European Chrysoperla spp.
Source:
http://www.eeb.uconn.edu/people/chenry/Cryptic_songs.html
The Cryptic Song Species
Oscillographs of some European Chrysoperla spp.
Source:
http://www.eeb.uconn.edu/people/chenry/Cryptic_songs.html
The Cryptic Song Species
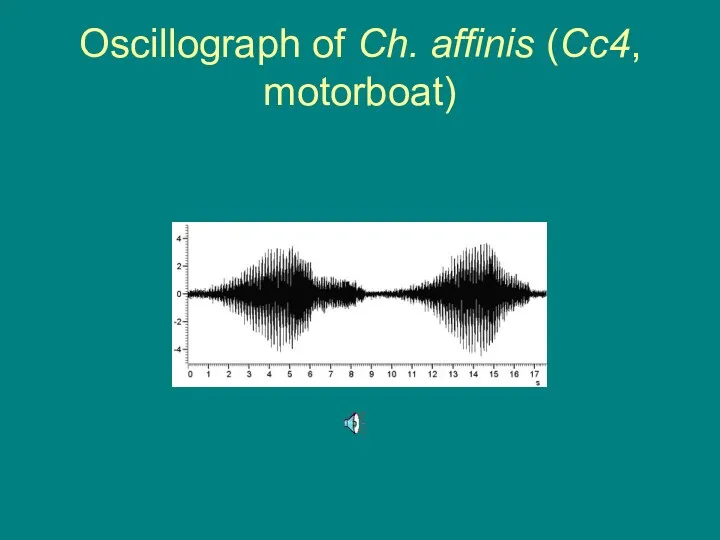
Oscillograph of Ch. affinis (Cc4, motorboat)
Oscillograph of Ch. affinis (Cc4, motorboat)
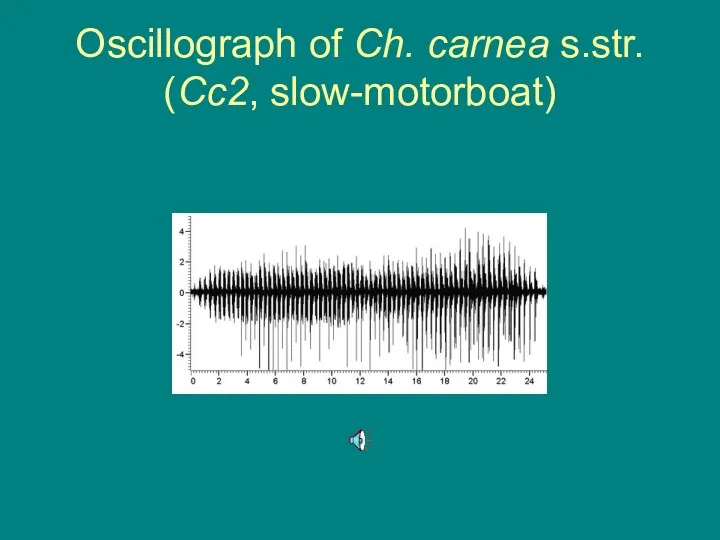
Oscillograph of Ch. carnea s.str. (Cc2, slow-motorboat)
Oscillograph of Ch. carnea s.str. (Cc2, slow-motorboat)

Oscillograph of Ch. lucasina
Oscillograph of Ch. lucasina
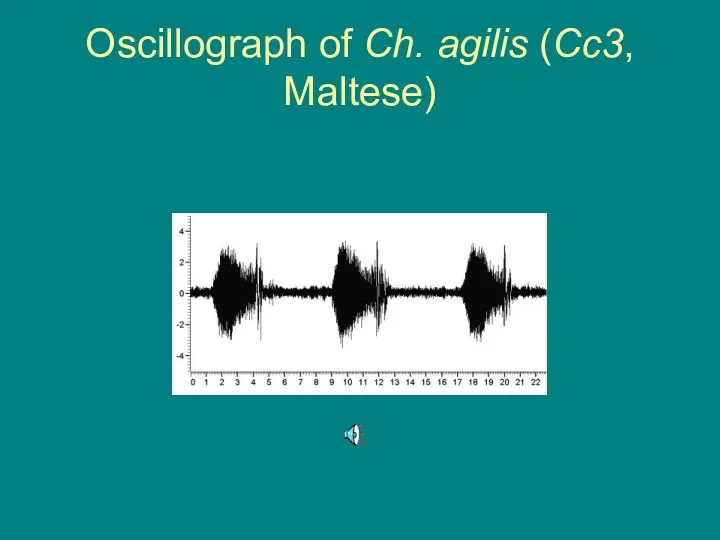
Oscillograph of Ch. agilis (Cc3, Maltese)
Oscillograph of Ch. agilis (Cc3, Maltese)

Morphology of Chrysoperla spp.
Morphology of Chrysoperla spp.

Ch. affinis
Ch. affinis

Ch. affinis (female)
Ch. affinis (female)

Ch. affinis
Ch. affinis

Ch. carnea s.str.
Ch. carnea s.str.

Ch. carnea s.str.
Ch. carnea s.str.

Ch. carnea s.str.
Ch. carnea s.str.

Ch. lucasina
Ch. lucasina

Ch. lucasina
Ch. lucasina

Ch. lucasina
Ch. lucasina

Ch. lucasina (female)
Ch. lucasina (female)

Ch. lucasina
Ch. lucasina

Practical troubles
One should not forget the natural enemy role of
Practical troubles
One should not forget the natural enemy role of

A practical example of the study of Ch. carnea complex
Management of
A practical example of the study of Ch. carnea complex
Management of

Natural control for olive moth
Olive moth (Prays oleae (Bernard) is one
Natural control for olive moth
Olive moth (Prays oleae (Bernard) is one

Questions
Which sibling species is the really efficient taxon?
In some years
Questions
Which sibling species is the really efficient taxon?
In some years

Future tasks
a.
- identification of the lacewing species (sibling species) controlling
Future tasks
a.
- identification of the lacewing species (sibling species) controlling
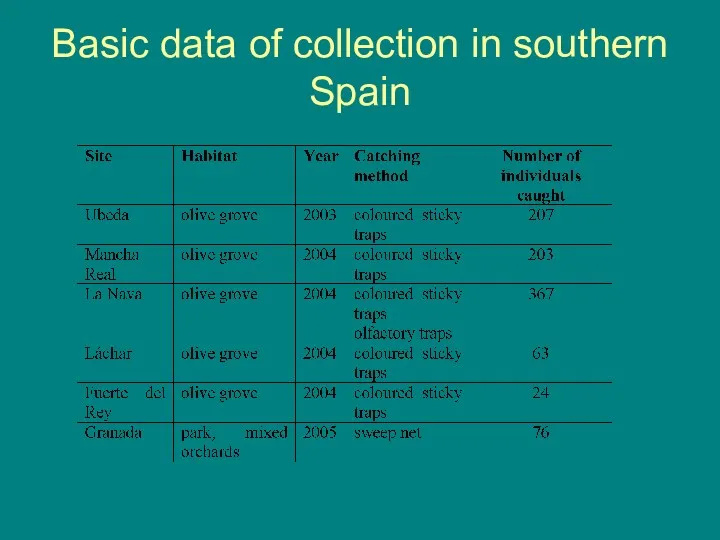
Basic data of collection in southern Spain
Basic data of collection in southern Spain

A typical olive grove
landscape
(Photo by R. González Ruiz)
A typical olive grove
landscape
(Photo by R. González Ruiz)

Olive trees
(Photo by R. González Ruiz)
Olive trees
(Photo by R. González Ruiz)

Identification
Individuals preserved in ethanol were identified according to the descriptions of
Identification
Individuals preserved in ethanol were identified according to the descriptions of
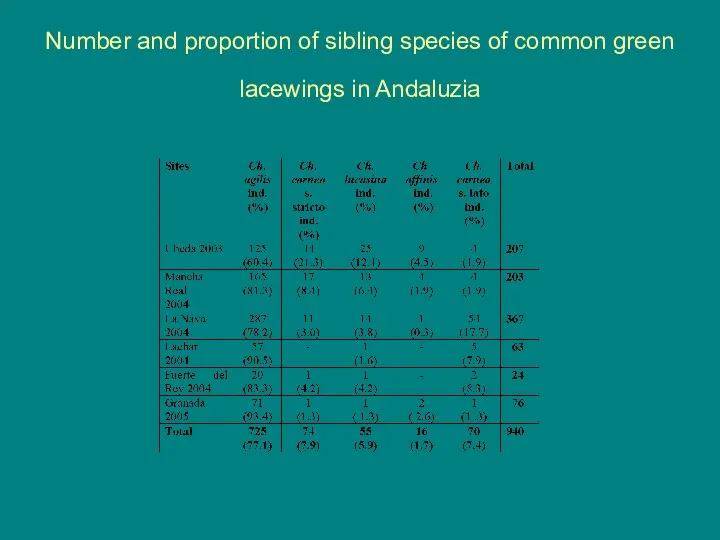
Number and proportion of sibling species of common green lacewings in
Number and proportion of sibling species of common green lacewings in

Collection sites of Ch. agilis (on the data of Henry et
Collection sites of Ch. agilis (on the data of Henry et

Conclusions
Ch. agilis is the dominant species in Southern Spain and the
Conclusions
Ch. agilis is the dominant species in Southern Spain and the

By biologic control and IPM we can save landscapes like that
By biologic control and IPM we can save landscapes like that

References
Al-Asaad, D. S. (2004): Viabilidad de los chrisópidos (Neuroptera: Chrysopidae) del
References
Al-Asaad, D. S. (2004): Viabilidad de los chrisópidos (Neuroptera: Chrysopidae) del

Henry, Ch. S., Brooks, S.J., Thierry, D., Duelli, P. & Johnson,
Henry, Ch. S., Brooks, S.J., Thierry, D., Duelli, P. & Johnson,
 Класс Двустворчатые моллюски
Класс Двустворчатые моллюски Строение и функции головного мозга
Строение и функции головного мозга Методы изучения природы
Методы изучения природы Курс общей биологии (размножение организмов)
Курс общей биологии (размножение организмов) Селекция
Селекция Розмноження на клітинному рівні. (Лекція 3)
Розмноження на клітинному рівні. (Лекція 3) Животные красной книги. Манул
Животные красной книги. Манул Биогенные элементы. Классификация. Топография биогенных элементов в организме человека
Биогенные элементы. Классификация. Топография биогенных элементов в организме человека Investigate DNA Extraction
Investigate DNA Extraction Молекулярно-генетические методы. Основы ПЦР
Молекулярно-генетические методы. Основы ПЦР Өсімдіктерден экономикалық маңызды заттарды өндірудің клеткалық технологиялары
Өсімдіктерден экономикалық маңызды заттарды өндірудің клеткалық технологиялары Презентация Костные рыбы (содержит видеофайл)
Презентация Костные рыбы (содержит видеофайл) Вирусология и открытие вирусов
Вирусология и открытие вирусов Зерновые в культуре стран: от Древности до наших дней
Зерновые в культуре стран: от Древности до наших дней Социальное партнёрство в профориентационном воспитании при изучении биологии
Социальное партнёрство в профориентационном воспитании при изучении биологии Гориллы
Гориллы Популяционная организация и динамика численности
Популяционная организация и динамика численности Эмбриология человека
Эмбриология человека Основы разведения животных
Основы разведения животных Помидор – полезный овощ. 2 класс
Помидор – полезный овощ. 2 класс Рослинний і тваринний світ України
Рослинний і тваринний світ України Синтез белка
Синтез белка Викторина Неделя леса - Сахаинский лес
Викторина Неделя леса - Сахаинский лес Отряды насекомых
Отряды насекомых Как растут фрукты
Как растут фрукты Чому мігруюють тварини
Чому мігруюють тварини Защитные функции крови
Защитные функции крови Среда обитания
Среда обитания