- Межклеточные сигнальные вещества

Содержание
- 2. Гормональная регуляция обмена веществ Гормоны – биологически активные вещества разной химической природы, секретируются железами внутренней секреции
- 3. Химическая природа гормонов 1. Гидрофильные: белки, пептиды, производные аминокислот (кроме тиреоидов) 2. Гидрофобные: стероиды, производные жирных
- 4. Этапы реализации действия гормонов 1. Регуляция синтеза гормонов (контроль нервной системы; тропная регуляция; механизм обратной связи)
- 5. Этапы реализации действия гормонов 5. Эффекторное звено: взаимоотношения с рецепторами. Непосредственное действие гормонов или образование вторичных
- 6. Как гормоны изменяют метаболизм в клетках- мишенях? Непосредственно или через вторичные мессенджеры: 1. Изменяют функциональную активность
- 7. Как гормоны изменяют метаболизм в клетках- мишенях? 2. Влияют на транскрипцию и последующий процессинг иРНК 3.
- 8. Основные типы мембранных рецепторов Рецепторы (> 200) сопряженные с G -белковыми комплексами (G –protein coupled receptors).
- 9. G – белковые комплексы > 20 G- белков (обладают GTP-азной активностью) Гетеротримеры (α, β, γ –
- 10. G – белковые комплексы Разные типы G –белков: Gs – стимулирует аденилатциклазу Gi – ингибирует аденилатциклазу
- 11. Вторичные мессенджеры биоактивных веществ Мессенджер Источник Эффект цАМФ –аденилатциклаза - Активирует протеинкиназу А цГМФ – гуанилатциклаза
- 12. Вторичные мессенджеры биоактивных веществ Инозитолтрифосфат – фосфолипаза С – активирует Ca2+ -каналы Диацилглицерол – фосфолипаза С
- 15. Фосфопротеинкиназы Активация протеинкиназ и последующее фосфорилирование ими разнообразных белковых субстратов обусловливает широкий спектр эффектов вторичных мессенджеров
- 16. Фосфопротеинкиназы ФПКG – гомодимер, активируется аллостерически 4 молекулами цГМФ. ФПК С – гетеродимер (RC), R –
- 17. Рецепторы = ионные каналы Связывание с лигандом приводит к изменению конформации рецептора, что позволяет специфическим ионам
- 18. Рецепторы, обладающие ферментативной активностью 1. Рецепторы, ассоциированные с гуанилатциклазной активностью 2. Рецепторы, ассоциированные с фосфатазной активностью
- 19. Рецепторы, не обладающие собственной каталитической активностью После связывания лиганда рецепторы связывают цитоплазматичекие протеинкиназы, фосфорилирующие рецептор (
- 20. Передача сигнала через внутриклеточные рецепторы Тиреоидные гормоны (иодированные производные тирозина) и стероиды проникают через цитоплазматическую мембрану
- 21. Гормоны гипоталамуса Нейропептиды гипоталамуса объединяют высшие отделы ЦНС и эндокринную систему. По системе портальных сосудов поступают
- 22. Гормоны гипоталамуса Либерины: тиреолиберин (трипептид), гонадолиберин (декапептид), кортиколиберин (41 аминокислота), соматолиберин (40 -44 аминокислотных остатка); Статины:
- 23. Гормоны гипоталамуса Нейропептиды рецептируются на поверхности соответствующих клеток гипофиза и активируют (либерины) или ингибируют (статины) аденилатциклазу.
- 24. Гормоны гипоталамуса Нейрогормоны (вазопрессин(АДГ) и окситоцин) попадают через аксоны в заднюю долю гипофиза (в комплексе с
- 25. Гормоны гипоталамуса Мишенью для окситоцина являются клетки гладких мышц (более всего рецепторов на мускулатуре матки и
- 26. Гормоны гипофиза Соматотропный гормон и пролактин – сходные по структуре белки (191 и 199 остатков аминокислот).
- 27. Гормон роста, СТГ Единственный видоспецифичный из всех гормонов гипофиза. Синтез и секреция стимулируются соматолиберином, тиреолиберином, эндорфином,
- 28. Гормон роста, СТГ Рецепторы – на плазматической мембране скелетных и мышечных тканей и практически всех внутренних
- 29. Гормон роста, СТГ Первичные эффекты СТГ сходны с инсулином: запасание клетками глюкозы и активация липогенеза. Вторичные
- 30. Гормон роста, СТГ СТГ способствует биосинтезу инсулина в поджелудочной железе и соматомединов (ИФР) в клетках-мишенях. ИФР
- 31. Пролактин Эстрогены в период беременности резко увеличивают количество лактотрофных клеток гипофиза. Импульсная секреция пролактина возрастает во
- 32. ТТГ, ФСГ, ЛГ, ХГ Гликопротеины, димеры из α –и β- субъединиц. α - субъединицы идентичны, β
- 33. Проопиомеланокортин (ПОМК) Под влиянием кортиколиберина гипоталамуса в аденогипофизе синтезируется крупный белковый предшественник (265 аминокислот). Лимитированный протеолиз
- 34. Адренокортикотропный гормон (АКТГ) Пептид (39 аминокислот). Стимулируется синтез кортиколиберином, стрессовыми воздействиями. Циркадный ритм: минимум – вечером,
- 35. Адренокортикотропный гормон (АКТГ) ХЛ ?ПРЕГНЕНОЛОН АКТГ имеет срочные и отдаленные механизмы, надежно обеспечивающие эту реакцию: Стимулируя
- 36. Гормоны щитовидной железы Под влиянием ТТГ в фолликулах синтезируется тиреоглобулин (гликопротеин) и секретируется во внеклеточное пространство,
- 37. Три- и тетраиодтиронин (тироксин) Большая часть гормонов находится в плазме в связанном с белком (неактивном) состоянии.
- 38. Три- и тетраиодтиронин (тироксин) Липофильные иодированные производные тирозина проникают в клетки и связываются с ядерными рецепторами.
- 39. Паратиреоидный гормон Белок, 84 аминокислоты (синтез идет в виде препрогормона 115 АК). Секреция стимулируется снижением уровня
- 40. Паратиреоидный гормон Стимулированные паратгормоном остеобласты секретируют ИФР и цитокины, индуцирующие в остеокластах синтез коллагеназы и фосфатазы
- 41. Кальцитонин Низкомолекулярный белок (предшественник из 136 аминокислот). Синтезируется в щитовидной железе, паращитовидной, тимусе, опухолевых клетках. Секретируется
- 42. Гормоны поджелудочной железы В островках Лангерганса: А клетки – глюкагон; В –клетки- инсулин Д клетки –
- 43. инсулин Белок из двух полипептидных цепей (21 и 30 аминокислот), соединенных 2 дисульфидными мостиками. Может образовывать
- 44. Инсулин Стимулируется синтез и секреция под влиянием глюкозы, аргинина и лизина, гормонов ЖКТ, СТГ, кортизола, эстрогенов.
- 45. Рецептор к инсулину Гликопротеиновый рецептор (IR) с тирозинкиназной активностью и способностью к автофосфорилированию и фосфорилированию целого
- 46. Глюкагон Полипептид (35 аминокислот). Препроглюкагон – белок (124 аминокислоты). Не найден транспортный белок. Угнетается секреция приемом
- 47. надпочечники Адреналин оказался первым гормоном, который удалось выделить, идентифицировать, синтезировать in vitro (1900г., Абель) 1899г. (Левандовский)
- 48. адреналин Катехоламины (адреналин в большей степени) осуществляют первую линию защиты организма при стрессе (гормон «бегства и
- 49. адреналин Секреция адреналина стимулируется тревожным состоянием, психическим возбуждением, гипоксией, гипогликемией. Тиреоидные гормоны увеличивают число β −
- 50. Синтез катехоламинов фенилаланин –> ДОФА –> дофамин – норадреналин –> адреналин (НАДФ – гидроксилазы, метилтрансфераза). Катехоламины
- 51. Инактивация катехоламинов Концентрация в крови – 0,05 нг/л, при стрессе – до 0,3 нг/л. при тяжелом
- 52. Метаболические эффекты адреналина Печень: активация гликогенфосфорилазы, нейрогенная острая гипергликемия. Одновременно – снижение секреции инсулина и продление
- 53. Стероиды коры надпочечников Под влиянием кортиколиберина гипоталамуса и АКТГ гипофиза (max утром и min вечером) из
- 54. Инактивация стероидов Период полужизни: 0,5 – 1,5 часа. Кортикостероиды и андрогены выделяются в виде 17-кетостероидов (окисление
- 55. кортикостероиды Для кортикостероидов характерна дозозависимость, вплоть до противоположных эффектов, различное влияние на разные мишени (тканеспецифичность). Метаболические
- 56. Метаболические эффекты кортикостероидов Белковый обмен: ↓поглощения аминокислот (кроме печени), ↑протеолиза, ↓транскрипции и трансляции (иммунодепрессия), ↑экскреции аминокислот,
- 57. альдостерон Регуляция синтеза и секреции: АКТГ (в меньшей степени), в большей степени ↓[Na+] в крови и
- 58. Половые стероиды Гонадолиберины гипоталамуса и гонадотропные гормоны гипофиза стимулируют синтез (через стимуляцию стероидгидроксилазных ферментов) и секрецию
- 60. Скачать презентацию

Гормональная регуляция обмена веществ
Гормоны – биологически активные вещества разной химической природы,
Гормональная регуляция обмена веществ
Гормоны – биологически активные вещества разной химической природы,

Химическая природа гормонов
1. Гидрофильные: белки, пептиды, производные аминокислот (кроме тиреоидов)
2. Гидрофобные:
Химическая природа гормонов
1. Гидрофильные: белки, пептиды, производные аминокислот (кроме тиреоидов)
2. Гидрофобные:

Этапы реализации действия гормонов
1. Регуляция синтеза гормонов (контроль нервной системы; тропная
Этапы реализации действия гормонов
1. Регуляция синтеза гормонов (контроль нервной системы; тропная

Этапы реализации действия гормонов
5. Эффекторное звено: взаимоотношения с рецепторами. Непосредственное действие
Этапы реализации действия гормонов
5. Эффекторное звено: взаимоотношения с рецепторами. Непосредственное действие

Как гормоны изменяют метаболизм в клетках- мишенях?
Непосредственно или через вторичные мессенджеры:
Как гормоны изменяют метаболизм в клетках- мишенях?
Непосредственно или через вторичные мессенджеры:

Как гормоны изменяют метаболизм в клетках- мишенях?
2. Влияют на транскрипцию и
Как гормоны изменяют метаболизм в клетках- мишенях?
2. Влияют на транскрипцию и

Основные типы мембранных рецепторов
Рецепторы (> 200) сопряженные с G -белковыми комплексами
Основные типы мембранных рецепторов
Рецепторы (> 200) сопряженные с G -белковыми комплексами

G – белковые комплексы
> 20 G- белков (обладают GTP-азной активностью)
Гетеротримеры (α,
G – белковые комплексы
> 20 G- белков (обладают GTP-азной активностью)
Гетеротримеры (α,

G – белковые комплексы
Разные типы G –белков:
Gs – стимулирует аденилатциклазу
Gi –
G – белковые комплексы
Разные типы G –белков:
Gs – стимулирует аденилатциклазу
Gi –

Вторичные мессенджеры биоактивных веществ
Мессенджер Источник Эффект
цАМФ –аденилатциклаза - Активирует протеинкиназу А
цГМФ
Вторичные мессенджеры биоактивных веществ
Мессенджер Источник Эффект
цАМФ –аденилатциклаза - Активирует протеинкиназу А
цГМФ

Вторичные мессенджеры биоактивных веществ
Инозитолтрифосфат – фосфолипаза С – активирует Ca2+ -каналы
Диацилглицерол
Вторичные мессенджеры биоактивных веществ
Инозитолтрифосфат – фосфолипаза С – активирует Ca2+ -каналы
Диацилглицерол
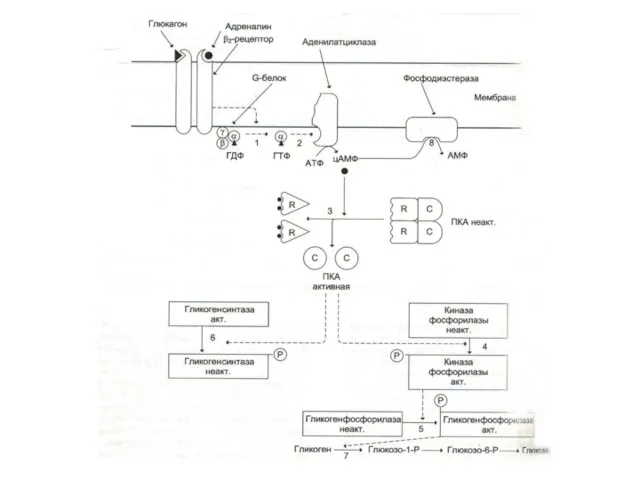
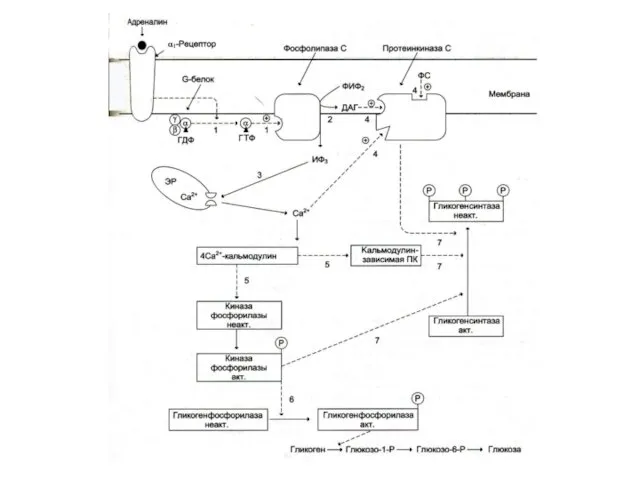

Фосфопротеинкиназы
Активация протеинкиназ и последующее фосфорилирование ими разнообразных белковых субстратов обусловливает широкий
Фосфопротеинкиназы
Активация протеинкиназ и последующее фосфорилирование ими разнообразных белковых субстратов обусловливает широкий

Фосфопротеинкиназы
ФПКG – гомодимер, активируется аллостерически 4 молекулами цГМФ.
ФПК С – гетеродимер
Фосфопротеинкиназы
ФПКG – гомодимер, активируется аллостерически 4 молекулами цГМФ.
ФПК С – гетеродимер

Рецепторы = ионные каналы
Связывание с лигандом приводит к изменению конформации рецептора,
Рецепторы = ионные каналы
Связывание с лигандом приводит к изменению конформации рецептора,

Рецепторы, обладающие ферментативной активностью
1. Рецепторы, ассоциированные с гуанилатциклазной активностью
2. Рецепторы,
Рецепторы, обладающие ферментативной активностью
1. Рецепторы, ассоциированные с гуанилатциклазной активностью
2. Рецепторы,

Рецепторы, не обладающие собственной каталитической активностью
После связывания лиганда рецепторы связывают цитоплазматичекие
Рецепторы, не обладающие собственной каталитической активностью
После связывания лиганда рецепторы связывают цитоплазматичекие

Передача сигнала через внутриклеточные рецепторы
Тиреоидные гормоны (иодированные производные тирозина) и стероиды
Передача сигнала через внутриклеточные рецепторы
Тиреоидные гормоны (иодированные производные тирозина) и стероиды

Гормоны гипоталамуса
Нейропептиды гипоталамуса объединяют высшие отделы ЦНС и эндокринную систему.
По системе
Гормоны гипоталамуса
Нейропептиды гипоталамуса объединяют высшие отделы ЦНС и эндокринную систему.
По системе

Гормоны гипоталамуса
Либерины: тиреолиберин (трипептид), гонадолиберин (декапептид), кортиколиберин (41 аминокислота), соматолиберин (40
Гормоны гипоталамуса
Либерины: тиреолиберин (трипептид), гонадолиберин (декапептид), кортиколиберин (41 аминокислота), соматолиберин (40

Гормоны гипоталамуса
Нейропептиды рецептируются на поверхности соответствующих клеток гипофиза и активируют (либерины)
Гормоны гипоталамуса
Нейропептиды рецептируются на поверхности соответствующих клеток гипофиза и активируют (либерины)

Гормоны гипоталамуса
Нейрогормоны (вазопрессин(АДГ) и окситоцин) попадают через аксоны в заднюю долю
Гормоны гипоталамуса
Нейрогормоны (вазопрессин(АДГ) и окситоцин) попадают через аксоны в заднюю долю

Гормоны гипоталамуса
Мишенью для окситоцина являются клетки гладких мышц (более всего рецепторов
Гормоны гипоталамуса
Мишенью для окситоцина являются клетки гладких мышц (более всего рецепторов

Гормоны гипофиза
Соматотропный гормон и пролактин – сходные по структуре белки (191
Гормоны гипофиза
Соматотропный гормон и пролактин – сходные по структуре белки (191

Гормон роста, СТГ
Единственный видоспецифичный из всех гормонов гипофиза.
Синтез и секреция стимулируются
Гормон роста, СТГ
Единственный видоспецифичный из всех гормонов гипофиза.
Синтез и секреция стимулируются

Гормон роста, СТГ
Рецепторы – на плазматической мембране скелетных и мышечных тканей
Гормон роста, СТГ
Рецепторы – на плазматической мембране скелетных и мышечных тканей

Гормон роста, СТГ
Первичные эффекты СТГ сходны с инсулином: запасание клетками глюкозы
Гормон роста, СТГ
Первичные эффекты СТГ сходны с инсулином: запасание клетками глюкозы

Гормон роста, СТГ
СТГ способствует биосинтезу инсулина в поджелудочной железе и соматомединов
Гормон роста, СТГ
СТГ способствует биосинтезу инсулина в поджелудочной железе и соматомединов

Пролактин
Эстрогены в период беременности резко увеличивают количество лактотрофных клеток гипофиза.
Импульсная секреция
Пролактин
Эстрогены в период беременности резко увеличивают количество лактотрофных клеток гипофиза.
Импульсная секреция

ТТГ, ФСГ, ЛГ, ХГ
Гликопротеины, димеры из α –и β- субъединиц.
α -
ТТГ, ФСГ, ЛГ, ХГ
Гликопротеины, димеры из α –и β- субъединиц.
α -

Проопиомеланокортин (ПОМК)
Под влиянием кортиколиберина гипоталамуса в аденогипофизе синтезируется крупный белковый предшественник
Проопиомеланокортин (ПОМК)
Под влиянием кортиколиберина гипоталамуса в аденогипофизе синтезируется крупный белковый предшественник

Адренокортикотропный гормон (АКТГ)
Пептид (39 аминокислот).
Стимулируется синтез кортиколиберином, стрессовыми воздействиями.
Циркадный ритм:
Адренокортикотропный гормон (АКТГ)
Пептид (39 аминокислот).
Стимулируется синтез кортиколиберином, стрессовыми воздействиями.
Циркадный ритм:

Адренокортикотропный гормон (АКТГ)
ХЛ ?ПРЕГНЕНОЛОН
АКТГ имеет срочные и отдаленные механизмы, надежно обеспечивающие
Адренокортикотропный гормон (АКТГ)
ХЛ ?ПРЕГНЕНОЛОН
АКТГ имеет срочные и отдаленные механизмы, надежно обеспечивающие

Гормоны щитовидной железы
Под влиянием ТТГ в фолликулах синтезируется тиреоглобулин (гликопротеин) и
Гормоны щитовидной железы
Под влиянием ТТГ в фолликулах синтезируется тиреоглобулин (гликопротеин) и

Три- и тетраиодтиронин (тироксин)
Большая часть гормонов находится в плазме в связанном
Три- и тетраиодтиронин (тироксин)
Большая часть гормонов находится в плазме в связанном

Три- и тетраиодтиронин (тироксин)
Липофильные иодированные производные тирозина проникают в клетки и
Три- и тетраиодтиронин (тироксин)
Липофильные иодированные производные тирозина проникают в клетки и

Паратиреоидный гормон
Белок, 84 аминокислоты (синтез идет в виде препрогормона 115 АК).
Секреция
Паратиреоидный гормон
Белок, 84 аминокислоты (синтез идет в виде препрогормона 115 АК).
Секреция

Паратиреоидный гормон
Стимулированные паратгормоном остеобласты секретируют ИФР и цитокины, индуцирующие в остеокластах
Паратиреоидный гормон
Стимулированные паратгормоном остеобласты секретируют ИФР и цитокины, индуцирующие в остеокластах

Кальцитонин
Низкомолекулярный белок (предшественник из 136 аминокислот).
Синтезируется в щитовидной железе, паращитовидной, тимусе,
Кальцитонин
Низкомолекулярный белок (предшественник из 136 аминокислот).
Синтезируется в щитовидной железе, паращитовидной, тимусе,

Гормоны поджелудочной железы
В островках Лангерганса:
А клетки – глюкагон;
В –клетки- инсулин
Д клетки
Гормоны поджелудочной железы
В островках Лангерганса:
А клетки – глюкагон;
В –клетки- инсулин
Д клетки

инсулин
Белок из двух полипептидных цепей (21 и 30 аминокислот), соединенных 2
инсулин
Белок из двух полипептидных цепей (21 и 30 аминокислот), соединенных 2

Инсулин
Стимулируется синтез и секреция под влиянием глюкозы, аргинина и лизина, гормонов
Инсулин
Стимулируется синтез и секреция под влиянием глюкозы, аргинина и лизина, гормонов

Рецептор к инсулину
Гликопротеиновый рецептор (IR) с тирозинкиназной активностью и способностью к
Рецептор к инсулину
Гликопротеиновый рецептор (IR) с тирозинкиназной активностью и способностью к

Глюкагон
Полипептид (35 аминокислот).
Препроглюкагон – белок (124 аминокислоты).
Не найден транспортный белок.
Угнетается секреция
Глюкагон
Полипептид (35 аминокислот).
Препроглюкагон – белок (124 аминокислоты).
Не найден транспортный белок.
Угнетается секреция

надпочечники
Адреналин оказался первым гормоном, который удалось выделить, идентифицировать, синтезировать in vitro
надпочечники
Адреналин оказался первым гормоном, который удалось выделить, идентифицировать, синтезировать in vitro

адреналин
Катехоламины (адреналин в большей степени) осуществляют первую линию защиты организма при
адреналин
Катехоламины (адреналин в большей степени) осуществляют первую линию защиты организма при

адреналин
Секреция адреналина стимулируется тревожным состоянием, психическим возбуждением, гипоксией, гипогликемией.
Тиреоидные гормоны увеличивают
адреналин
Секреция адреналина стимулируется тревожным состоянием, психическим возбуждением, гипоксией, гипогликемией.
Тиреоидные гормоны увеличивают

Синтез катехоламинов
фенилаланин –> ДОФА –> дофамин – норадреналин –> адреналин (НАДФ
Синтез катехоламинов
фенилаланин –> ДОФА –> дофамин – норадреналин –> адреналин (НАДФ

Инактивация катехоламинов
Концентрация в крови – 0,05 нг/л, при стрессе – до
Инактивация катехоламинов
Концентрация в крови – 0,05 нг/л, при стрессе – до

Метаболические эффекты адреналина
Печень: активация гликогенфосфорилазы, нейрогенная острая гипергликемия.
Одновременно – снижение секреции
Метаболические эффекты адреналина
Печень: активация гликогенфосфорилазы, нейрогенная острая гипергликемия.
Одновременно – снижение секреции

Стероиды коры надпочечников
Под влиянием кортиколиберина гипоталамуса и АКТГ гипофиза (max утром
Стероиды коры надпочечников
Под влиянием кортиколиберина гипоталамуса и АКТГ гипофиза (max утром

Инактивация стероидов
Период полужизни: 0,5 – 1,5 часа.
Кортикостероиды и андрогены выделяются в
Инактивация стероидов
Период полужизни: 0,5 – 1,5 часа.
Кортикостероиды и андрогены выделяются в

кортикостероиды
Для кортикостероидов характерна дозозависимость, вплоть до противоположных эффектов, различное влияние на
кортикостероиды
Для кортикостероидов характерна дозозависимость, вплоть до противоположных эффектов, различное влияние на

Метаболические эффекты кортикостероидов
Белковый обмен:
↓поглощения аминокислот (кроме печени), ↑протеолиза, ↓транскрипции и трансляции
Метаболические эффекты кортикостероидов
Белковый обмен:
↓поглощения аминокислот (кроме печени), ↑протеолиза, ↓транскрипции и трансляции

альдостерон
Регуляция синтеза и секреции: АКТГ (в меньшей степени), в большей степени
альдостерон
Регуляция синтеза и секреции: АКТГ (в меньшей степени), в большей степени

Половые стероиды
Гонадолиберины гипоталамуса и гонадотропные гормоны гипофиза стимулируют синтез (через стимуляцию
Половые стероиды
Гонадолиберины гипоталамуса и гонадотропные гормоны гипофиза стимулируют синтез (через стимуляцию
 Наш живой уголок
Наш живой уголок ПРЕЗЕНТАЦИИ 5 КЛАСС 1 ЧЕТВЕРТЬ
ПРЕЗЕНТАЦИИ 5 КЛАСС 1 ЧЕТВЕРТЬ Орган зрения
Орган зрения Популяции
Популяции Викторина: Фрукты
Викторина: Фрукты ДНҚ-ның метилденуі
ДНҚ-ның метилденуі Голосеменные растения
Голосеменные растения Загадки на тему Фрукты
Загадки на тему Фрукты Physiology of Bacteria
Physiology of Bacteria Удод
Удод Семейство: Гвоздичные
Семейство: Гвоздичные 20181126_razvitie_nasekomyh
20181126_razvitie_nasekomyh Позвоночные животные
Позвоночные животные Мутации. Классификации. Мутации и мобильные элементы. Мобильные элементы генома, классификация и их генетическая роль
Мутации. Классификации. Мутации и мобильные элементы. Мобильные элементы генома, классификация и их генетическая роль Надкласс Рыбы
Надкласс Рыбы Doom and Boom on a Resilient Reef: Climate Change, Algal Overgrowth and Coral Recovery
Doom and Boom on a Resilient Reef: Climate Change, Algal Overgrowth and Coral Recovery Скелет человека
Скелет человека Популяция. Свойства популяций
Популяция. Свойства популяций Свинья. Биологические особенности
Свинья. Биологические особенности Презентация по биологии Мутагены. Их влияние на природу и человека
Презентация по биологии Мутагены. Их влияние на природу и человека Побег. Почка
Побег. Почка Содержание селена и элементов антагонистов в почве и продуктах пчеловодства
Содержание селена и элементов антагонистов в почве и продуктах пчеловодства Типы питания живых организмов
Типы питания живых организмов Модификационная изменчивость и её характеристики. Биометрические методы оценки степени и характера изменчивости
Модификационная изменчивость и её характеристики. Биометрические методы оценки степени и характера изменчивости Гистологическое строение печени
Гистологическое строение печени Безусловные рефлексы
Безусловные рефлексы Глотка. Пищевод. Желудок
Глотка. Пищевод. Желудок Мир животных - мир звуков
Мир животных - мир звуков