- Молекулярно-генетические механизмы развития корня

Содержание
- 2. Строение апикальной части корня Корневая апикальная меристема (КАМ, RAM)
- 3. Сравнение организации апикальной меристемы побега и корня Sarkar et al., Nature (2007) V. 446: 811-814 cтволовые
- 4. Эмбриональное происхождение корня Arabidopsis 1. первое зиготическое деление 2. стадия октанта 4. проросток 3. стадия сердцевидного
- 5. Ткани корня арабидопсиса Scheres Arabidopsis Book 2002
- 6. Организация инициальных клеток корня Ткани корня развиваются из инициальных (стволовых) клеток: -инициали эпидермы -перицикл/васкулярные инициали -
- 7. Число клеток ПЦ различается у разных видов: у арабидопсиса ПЦ состоит из 4 клеток, у кукурузы
- 8. Clowes (1975) Обнаружение покоящегося центра в корне кукурузы в экспериментах по введению 3H-тимидина Клетки ПЦ в
- 9. Гранулы крахмала в клетках колумеллы арабидопсиса (окрашивание люголем) В инициалях колумеллы отсутствуют гранулы крахмала Дифференцировка инициалей
- 10. Клетки ПЦ ингибируют дифференцировку клеток кортико-эндодермальных инициалей При удаление лазером 1 клетки ПЦ прилегающая кортико-эндодермальная инициальная
- 11. Факторы, отвечающие за формирование ПЦ и поддержание стволовых клеток в меристеме корня
- 12. Поддержание стволовых клеток в побеговой и корневой апикальных меристемах (a) Продольный срез апикальной меристемы побега. Показана
- 13. Роль гена WOX5 в АМ корня Экспрессия WOX5 в ПЦ Корень мутанта wox5-1 характеризуется увеличением размера
- 14. Индукция экспрессии WOX5 Блокирование дифференцировки клеток колумеллы + активация их деления Активность WOX5 достаточна для подавления
- 15. Экспрессия гена WOX5 (гомолог гена WUS) в эмбриогенезе (гибридизация in situ) Haeker et al., Development 2004
- 16. WUS WOX5 WUS и WOX5 экспрессируются в «организующих центрах» АМ побега и корня, соответственно АМ побега
- 17. Являются ли гены WUS и WOX5 функционально эквивалентными? wox 5-1 РWOX5-WUS/wox5-1 wus-1 РWUS-WOX5/wus-1 WOX5-WUS восстанавливает нормальный
- 18. ТФ WOX5 обеспечивает «покой» ПЦ, негативно регулируя гены CYCD Подавление делений клеток ПЦ зависит от активности
- 19. ТФ WOX5 перемещается в окружающие стволовые клетки (на примере инициалей колумеллы) Мишень WOX5-зависимой репрессии: ген, кодирующий
- 20. Опыты по обнаружению перемещения белка WOX5 в инициали колумеллы Pi et al., Developmental Cell, 2015 Промотор
- 21. Фенотип мутации cle40 Роль гена CLE40 в меристеме корня CLE40 экспрессируется в стеле в клетках колумеллы
- 22. Влияние CLE40 на число клеток, экспрессирующих WOX5, и на содержание гранул крахмала в клетках колумеллы Stahl
- 23. Гомеостаз коллумеллы и ПЦ зависит от противоположного действия CLE40 и WOX5
- 24. Ген ACR4 кодирует RLK, рецептор пептида CLE40 В меристеме корня мутантов acr4 увеличено число клеток, экспрессирующих
- 26. ПAM КAM Апикальные меристемы побега и корня имеют сходный план строения и механизмы регуляции WUS WOX5
- 27. pCyclinB1::CyclinB1-GFP CLE-пептиды CLE14 и CLE20 подавляют клеточные деления в меристеме корня и могут связываться с рецепторным
- 28. Подавлять активность меристемы корня могут почти все CLE-пептиды группы А Влияние сверхэкспрессии CLE19 на фенотип корня:
- 29. Участие систем WOX-CLV в регуляции первичных разных меристем Тип меристемы Ген WOX Компонент системы CLAVATA Апикальная
- 30. Scheres Arabidopsis Book 2002 + NPA + NPA А Б В Г А – Экспрессия DR5::GUS
- 31. Роль белков PIN в транспорте ауксина в корне PIN1 PIN2 PIN1 PIN2
- 32. Petrasek and Friml, 2009 Роль белков PIN в формировании максимума ауксина в кончике корня корне DR5::GUS
- 33. Нарушение апикально-базальной структуры корня у мутантов по генам, вовлеченным в метаболизм ауксина: gnom (gn) monopteros (mp)
- 34. DR5::GFP Распределение ауксина в зародыше на ранних стадиях развития: Вывод: Локальный максимум концентрации ауксина нужен для
- 35. Распределение ауксина в зародыше на ранних стадиях развития:
- 36. Экспрессия DR5::GUSа кончике корня проростков арабидопсиса 1 - Окрашивание 30 сек 2 - Окрашивание 16 часов
- 37. Области клеточных делений и ориентация клеточных стенок у дочерних клеток + 10 мкм NPA + 50
- 38. 1. WT 2. plt1-3 3. plt1-4 4. plt2-2 5. plt1-3 plt2-2 6. plt1-4 plt2-2. 1 2
- 39. PLETHORA MP PIN plt1plt2plt3 wt PLT1 PLT2 PIN1 PIN3 PIN2 mp-G12 wt PLETHORA Связь между PLETHORA
- 40. Поиск мишеней ТФ PLETHORA Santuiari et al., Plant Cell 2016
- 41. Поиск мишеней ТФ PLETHORA Santuiari et al., Plant Cell 2016
- 42. scr-1 wt Роль гена SCARECROW (SCR) в формировании ПЦ и в поддержании стволовых клеток scr-1,UAS::SCR,QC46 Ген
- 43. Эктопическая экспрессия SHORTROOT (pSCR::SHR) приводит к увеличению слоев клеток эндодермы Дополнительные слои клеток эндодермы (маркированы активностью
- 44. Экспрессия SCR в ПЦ у мутантов scr-1 восстановление структуры и функции ПЦ Sabatini et al., GENES
- 45. scr-1 / N9094 / UAS::SCR Экспрессия SCR в коре и эндодерме у мутантов scr-1 восстановление радиальной
- 46. Роль SCARECROW в апикальной меристеме корня J2341 – SCR экспрессируется в области ПЦ => восстановление структуры
- 47. Регуляторная сеть, контролирующая функционирование меристемы корня Sablowski, Current Opinion in Plant Biology 2007
- 48. ФОРМИРОВАНИЕ РАДИАЛЬНОЙ СТРУКТУРЫ КОРНЯ
- 49. кодируют транскрипционные факторы семейства GRAS Основные гены, отвечающие за развитие эндодермы и коры (контроль делений инициалей
- 50. Мутация scarecrow C - кортекс; En - эндодерма; Ep - эпидермис; M - мутантный клеточный слой
- 51. Scheres et al., Arabidopsis Book 2002 Экспрессия гена SCR
- 52. Экспрессия гена SHR (А) pSHR::GFP, WT (В) pSHR::SHR::GFP shr-2
- 53. Модель транспорта белка SHORTROOT через плазмодесмы и его участия в формировании радиальной структуры корня Barton Cell
- 54. Перемещение белка SHR из стелы во внешние слои клеток негативно регулируется при взаимодействии с SCR
- 55. Гены-мишени SCR и SHR Sozzani et al., 2010 CYCD6;1 SHR
- 56. Регуляция клеточного цикла растений
- 57. Экспрессия pCYCD6;1::GFP в клетках-инициалях коры/эндодермы CYCD6;1 – изоформа СycD, специфичная для клеток-инициалей коры/эндодермы, регулирует асимметричное деление
- 58. У мутанта cycd6;1 – нарушено деление клеток-инициалей коры/эндодермы в эмбриогенезе Sozzani et al., 2010
- 59. Генетический контроль дифференцировки клеток эпидермы
- 60. Генетический контроль дифференцировки клеток эпидермы Поперечный срез корня арабидопсиса Сканирующая электронная микрофотография корневого волоска Показано расположение
- 61. Фенотипы мутаций по генам, определяющим дифференцировку клеток эпидермы WT gl2 ttg wer 35S::CPC cpc Поперечный срез
- 62. Фенотипы мутаций по генам, определяющим дифференцировку клеток эпидермы A. Wild type B. cpc C. 35S::CPC D.
- 63. Экспрессия в эпидерме Основные гены, вовлеченные в контроль дифференцировки клеток эпидермы у арабидопсиса: GLABRA2 (GL2) –транскрипционный
- 64. Экспрессия GL2::GUS «Неволосковые» клетки
- 65. Позиционный сигнал, поступающий из клеток кортекса, определяет судьбу клеток эпидермиса: эпидермальная клетки в “H”-положении располагается между
- 66. По тому же принципу регулируется формирование трихомов на стебле (но там GL2 является ПОЗИТИВНЫМ регулятором)
- 67. Модель дифференцировки клеток эпидермиса в корне арабидопсиса JACKDAW – белок с Zn-fingers, регулирует синтез гипотетического сигнала-
- 68. Механизмы роста волосковой клетки
- 69. Механизмы роста волосковой клетки
- 70. Развитие бокового корня
- 71. Формирование бокового корня из клеток перицикла у арабидопсиса У арабидопсиса боковой корень образуется из клеток перицикла,
- 72. Развитие бокового корня у пшеницы Демченко, 2002 У пшеницы боковой корень образуется из клеток перицикла, расположенных
- 73. zone A – the root apical meristem where active cell divisions take place. zone B –
- 74. меристема Triticum aestivum L. (мягкая пшеница) Cucurbita pepo L (тыква) Различные стратегии инициации бокового корня Демченко,
- 75. Стадии развития бокового корня Malamy and Benfey Development 1997
- 76. Экспрессия DR5::GUS при развитии бокового корня Роль ауксина в развитии бокового корня
- 77. Транспорт ауксина при развитии бокового корня
- 78. Колебания концентрации ауксина в меристеме корня определяют периодичность закладки боковых корней De Smet et al. 2007
- 79. ТФ NAC1 опосредует ауксиновый ответ при развитии бокового корня Гены ауксинового ответа DBP, AIR3 TIR NAC1
- 80. У мутантов alf4 снижена экспрессия циклина CYCB1;1 (G2-M) и и повышена экспрессия CDKB.1 DiDonato et al.,
- 81. Основные гормоны, регулирующие развитие корня: -ауксин (формирование покоящегося центра, стимуляция развития боковых корней) -цитокинин (подавление развития
- 82. AHK3 ARR1 и ARR12 Роль цитокинина в развитии корня
- 83. Роль цитокинина в развитии корня цитокинин Perilli, 2012 ARR1 и ARR12 (регуляторы цитокининового ответа B-типа) SHY
- 84. Асимметричное деление клеток перицикла градиент ауксина осцилляция ауксинового ответа Клетки-основательницы БК GATA23 Выход БК на поверхность
- 85. Конверсия примордиев боковых корней (БК) в меристему побега (ПАМ) по действием цитокинина 10 мкмМ НУК (ауксин),
- 88. Скачать презентацию
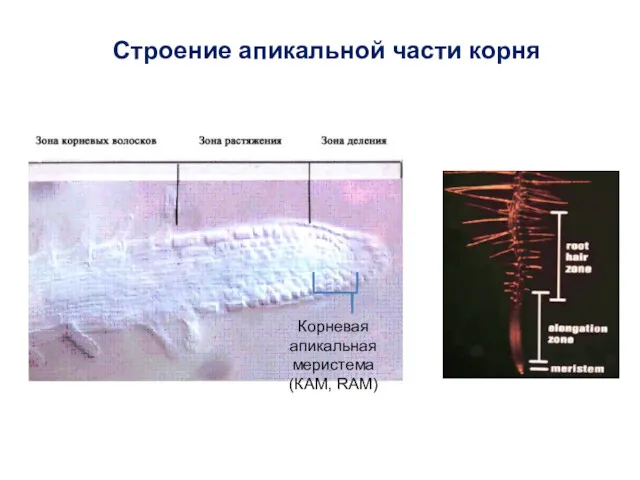
Строение апикальной части корня
Корневая апикальная меристема (КАМ, RAM)
Строение апикальной части корня
Корневая апикальная меристема (КАМ, RAM)
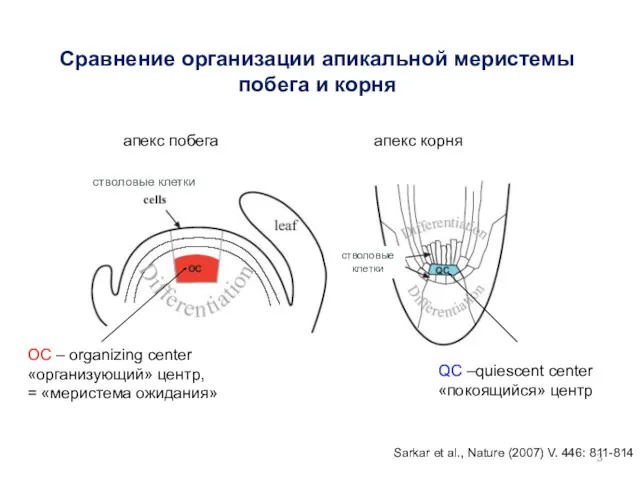
Сравнение организации апикальной меристемы побега и корня
Sarkar et al., Nature
Сравнение организации апикальной меристемы побега и корня
Sarkar et al., Nature
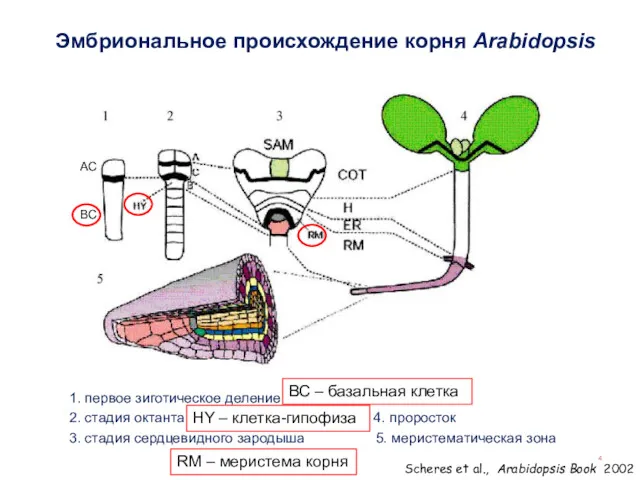
Эмбриональное происхождение корня Arabidopsis
1. первое зиготическое деление
2. стадия октанта 4. проросток
Эмбриональное происхождение корня Arabidopsis
1. первое зиготическое деление
2. стадия октанта 4. проросток
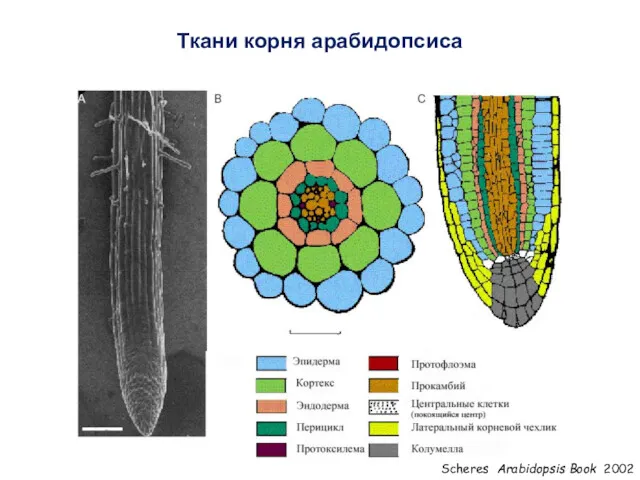
Ткани корня арабидопсиса
Scheres Arabidopsis Book 2002
Ткани корня арабидопсиса
Scheres Arabidopsis Book 2002
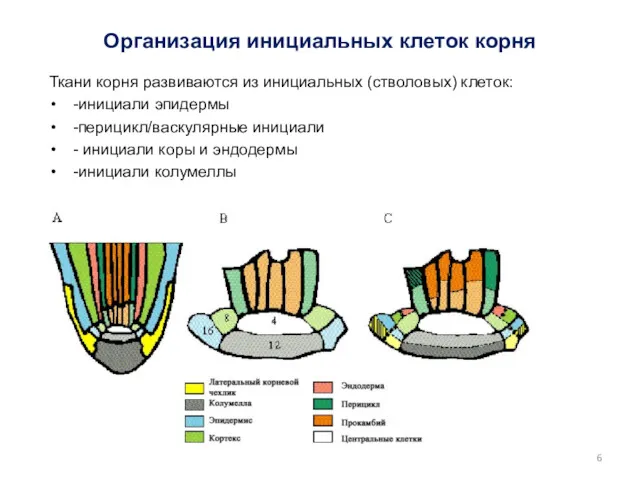
Организация инициальных клеток корня
Ткани корня развиваются из инициальных (стволовых) клеток:
-инициали эпидермы
-перицикл/васкулярные
Организация инициальных клеток корня
Ткани корня развиваются из инициальных (стволовых) клеток:
-инициали эпидермы
-перицикл/васкулярные
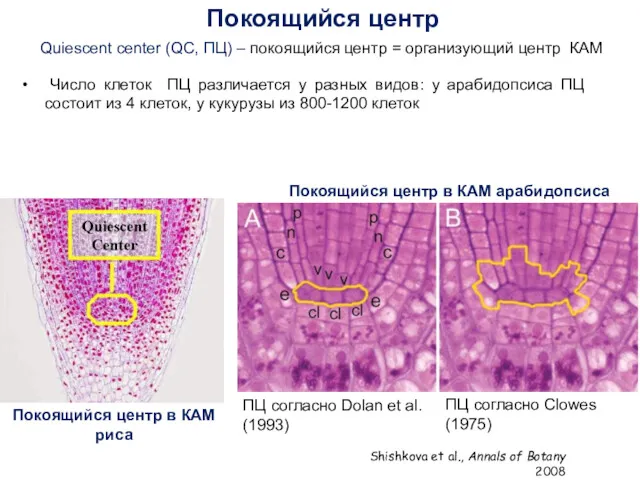
Число клеток ПЦ различается у разных видов: у арабидопсиса ПЦ состоит
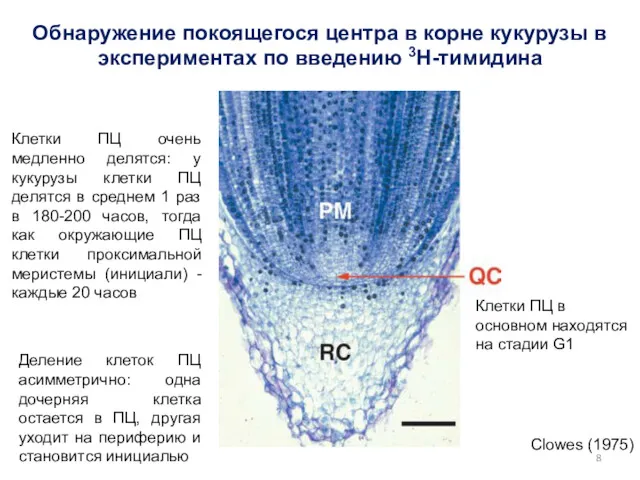
Clowes (1975)
Обнаружение покоящегося центра в корне кукурузы в экспериментах по введению
Clowes (1975)
Обнаружение покоящегося центра в корне кукурузы в экспериментах по введению
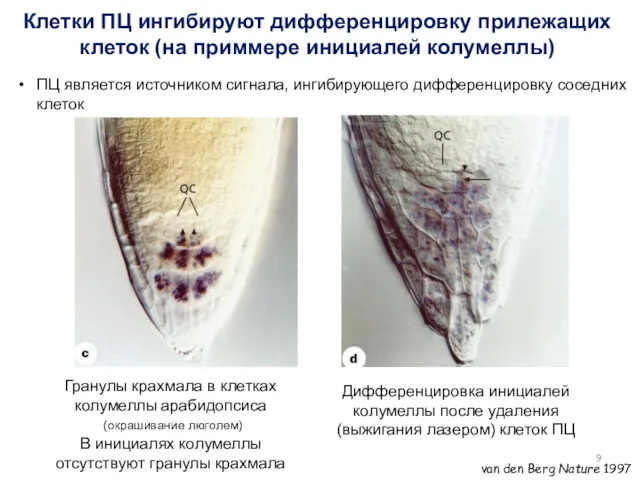
Гранулы крахмала в клетках колумеллы арабидопсиса
(окрашивание люголем)
В инициалях колумеллы
Гранулы крахмала в клетках колумеллы арабидопсиса
(окрашивание люголем)
В инициалях колумеллы
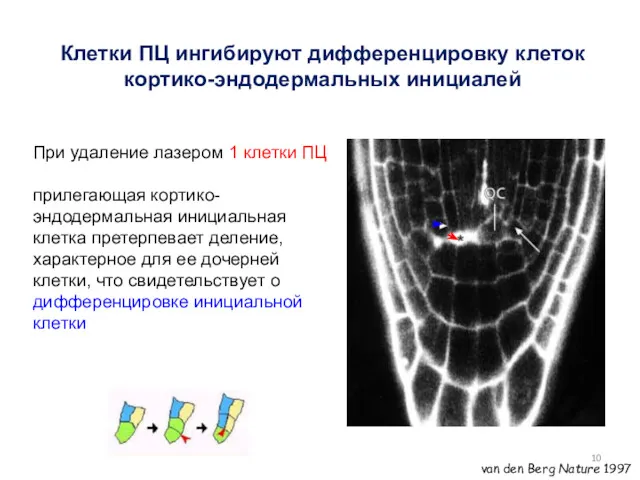
Клетки ПЦ ингибируют дифференцировку клеток кортико-эндодермальных инициалей
При удаление лазером 1 клетки
Клетки ПЦ ингибируют дифференцировку клеток кортико-эндодермальных инициалей
При удаление лазером 1 клетки

Факторы, отвечающие за формирование ПЦ и поддержание стволовых клеток в меристеме
Факторы, отвечающие за формирование ПЦ и поддержание стволовых клеток в меристеме
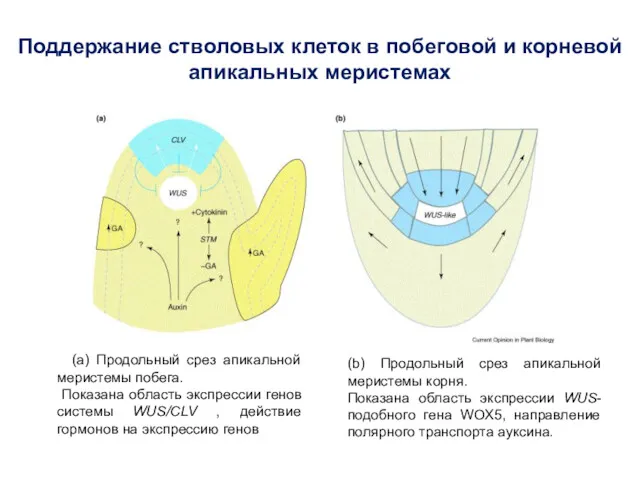
Поддержание стволовых клеток в побеговой и корневой апикальных меристемах
(a) Продольный
Поддержание стволовых клеток в побеговой и корневой апикальных меристемах
(a) Продольный
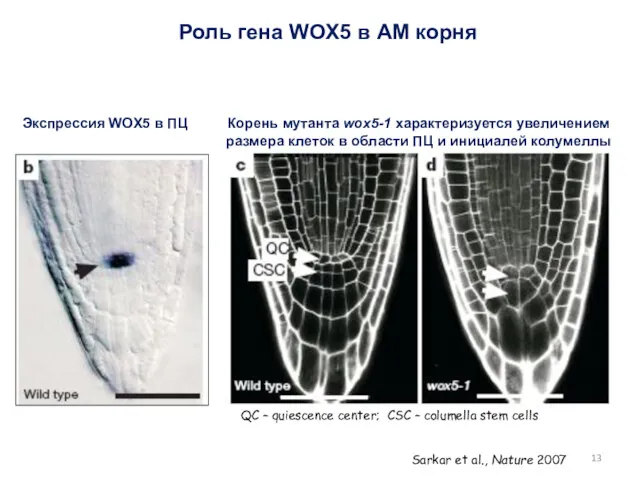
Роль гена WOX5 в АМ корня
Экспрессия WOX5 в ПЦ
Корень мутанта wox5-1
Роль гена WOX5 в АМ корня
Экспрессия WOX5 в ПЦ
Корень мутанта wox5-1

Индукция экспрессии WOX5
Блокирование дифференцировки клеток колумеллы + активация их деления
Активность
Индукция экспрессии WOX5
Блокирование дифференцировки клеток колумеллы + активация их деления
Активность
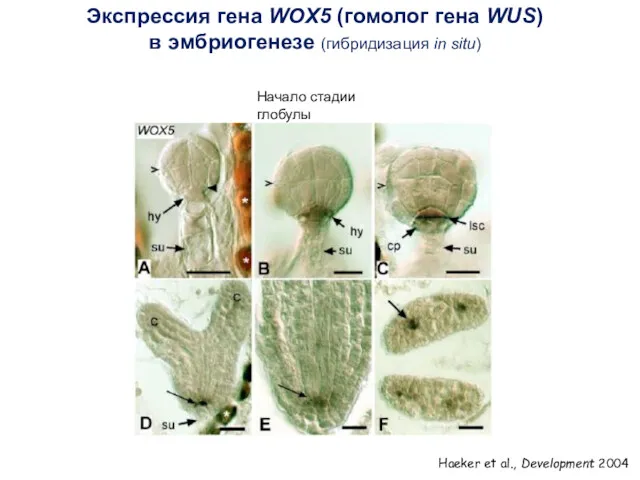
Экспрессия гена WOX5 (гомолог гена WUS)
в эмбриогенезе (гибридизация in situ)
Экспрессия гена WOX5 (гомолог гена WUS)
в эмбриогенезе (гибридизация in situ)
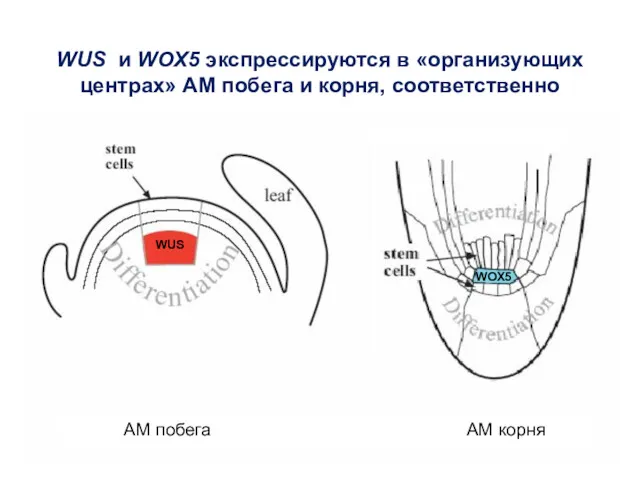
WUS
WOX5
WUS и WOX5 экспрессируются в «организующих центрах» АМ побега и корня,
WUS
WOX5
WUS и WOX5 экспрессируются в «организующих центрах» АМ побега и корня,
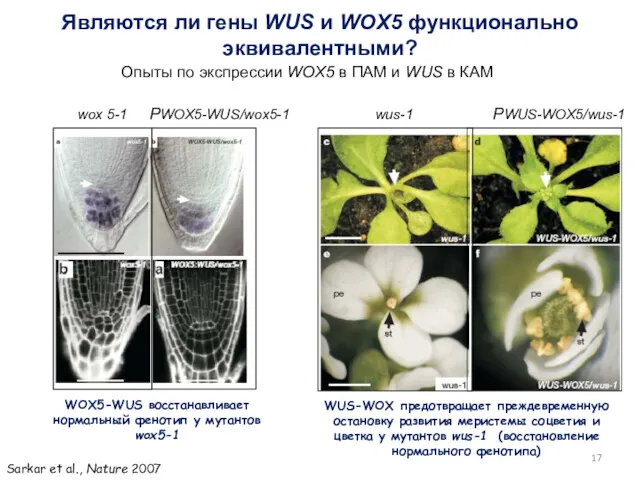
Являются ли гены WUS и WOX5 функционально эквивалентными?
wox 5-1 РWOX5-WUS/wox5-1
wus-1
Являются ли гены WUS и WOX5 функционально эквивалентными?
wox 5-1 РWOX5-WUS/wox5-1
wus-1
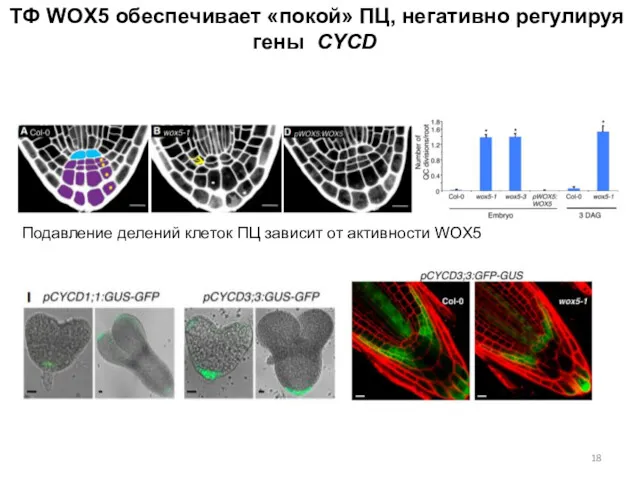
ТФ WOX5 обеспечивает «покой» ПЦ, негативно регулируя гены CYCD
Подавление
ТФ WOX5 обеспечивает «покой» ПЦ, негативно регулируя гены CYCD
Подавление
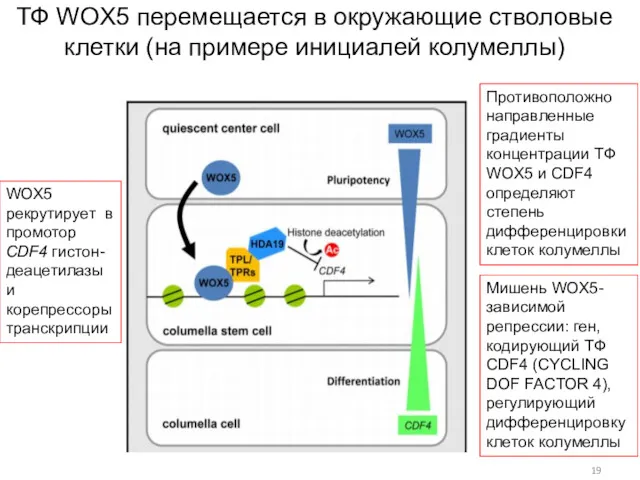
ТФ WOX5 перемещается в окружающие стволовые клетки (на примере инициалей колумеллы)
Мишень
ТФ WOX5 перемещается в окружающие стволовые клетки (на примере инициалей колумеллы)
Мишень
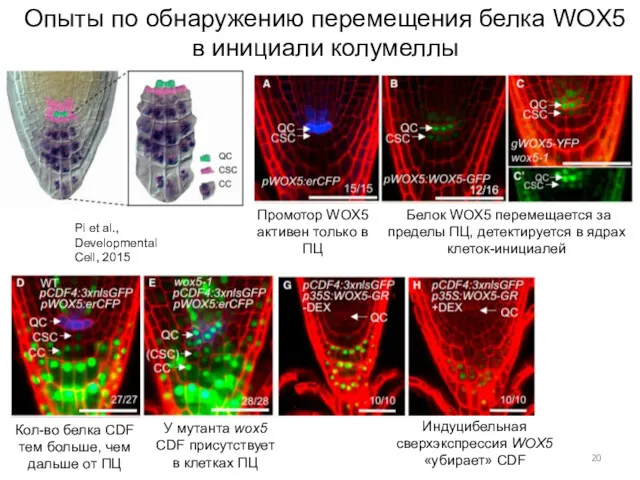
Опыты по обнаружению перемещения белка WOX5 в инициали колумеллы
Pi et al.,
Опыты по обнаружению перемещения белка WOX5 в инициали колумеллы
Pi et al.,
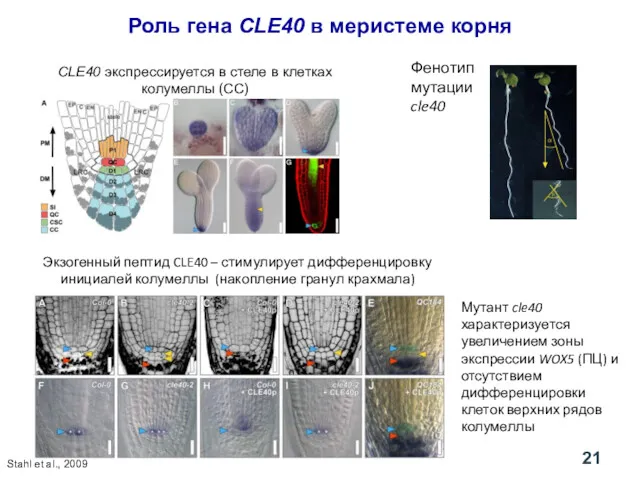
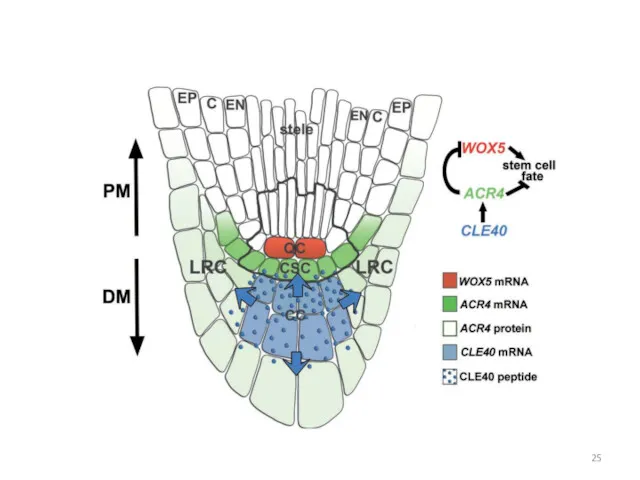
Фенотип мутации cle40
Роль гена CLE40 в меристеме корня
CLE40 экспрессируется в стеле
Фенотип мутации cle40
Роль гена CLE40 в меристеме корня
CLE40 экспрессируется в стеле

Влияние CLE40 на число клеток, экспрессирующих WOX5, и на содержание гранул
Влияние CLE40 на число клеток, экспрессирующих WOX5, и на содержание гранул
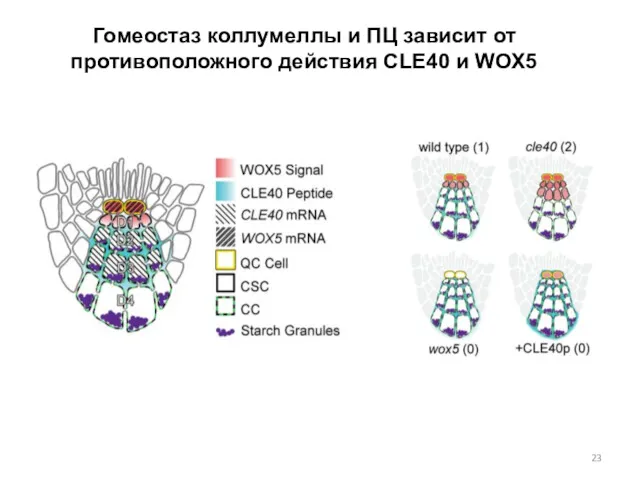
Гомеостаз коллумеллы и ПЦ зависит от противоположного действия CLE40 и WOX5
Гомеостаз коллумеллы и ПЦ зависит от противоположного действия CLE40 и WOX5
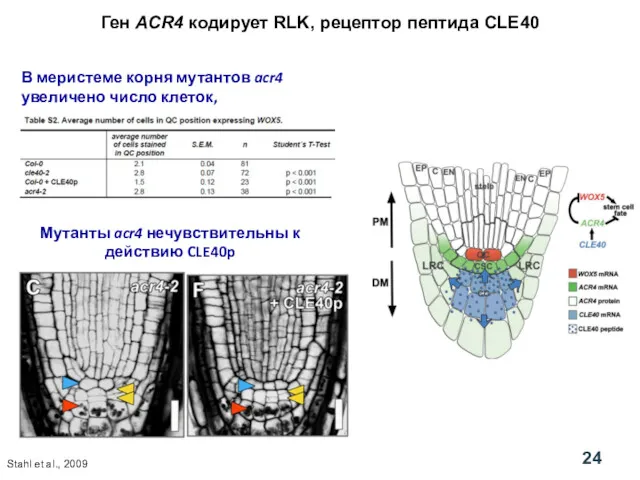
Ген ACR4 кодирует RLK, рецептор пептида CLE40
В меристеме корня мутантов acr4
Ген ACR4 кодирует RLK, рецептор пептида CLE40
В меристеме корня мутантов acr4
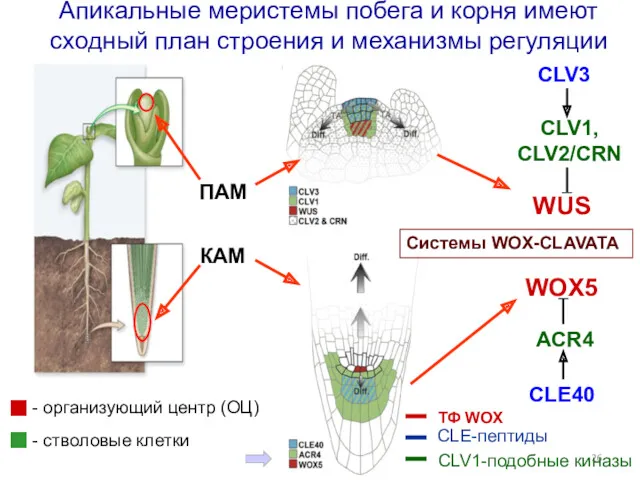
ПAM
КAM
Апикальные меристемы побега и корня имеют сходный план строения и механизмы
ПAM
КAM
Апикальные меристемы побега и корня имеют сходный план строения и механизмы
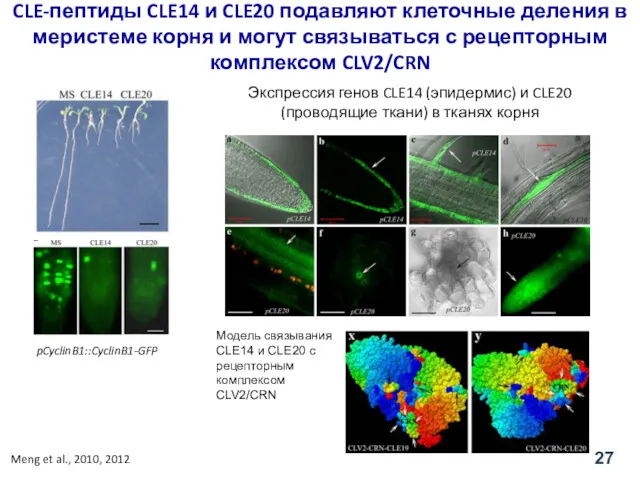
pCyclinB1::CyclinB1-GFP
CLE-пептиды CLE14 и CLE20 подавляют клеточные деления в меристеме корня и
pCyclinB1::CyclinB1-GFP
CLE-пептиды CLE14 и CLE20 подавляют клеточные деления в меристеме корня и
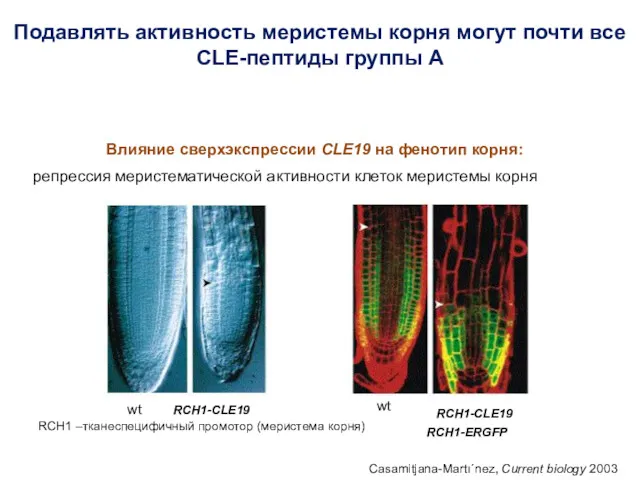
Подавлять активность меристемы корня могут почти все CLE-пептиды группы А
Влияние сверхэкспрессии
Подавлять активность меристемы корня могут почти все CLE-пептиды группы А
Влияние сверхэкспрессии
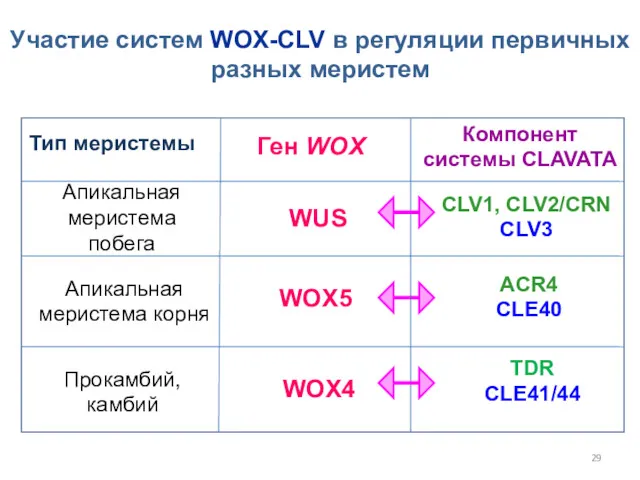
Участие систем WOX-CLV в регуляции первичных разных меристем
Тип меристемы
Ген WOX
Компонент
системы
Участие систем WOX-CLV в регуляции первичных разных меристем
Тип меристемы
Ген WOX
Компонент
системы
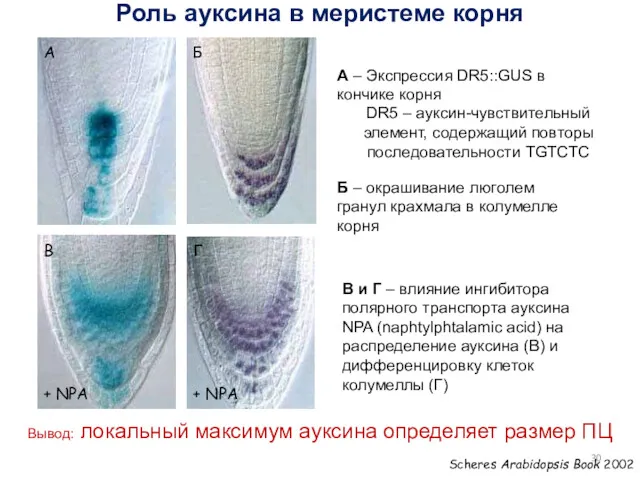
Scheres Arabidopsis Book 2002
+ NPA
+ NPA
А
Б
В
Г
А – Экспрессия DR5::GUS
Scheres Arabidopsis Book 2002
+ NPA
+ NPA
А
Б
В
Г
А – Экспрессия DR5::GUS
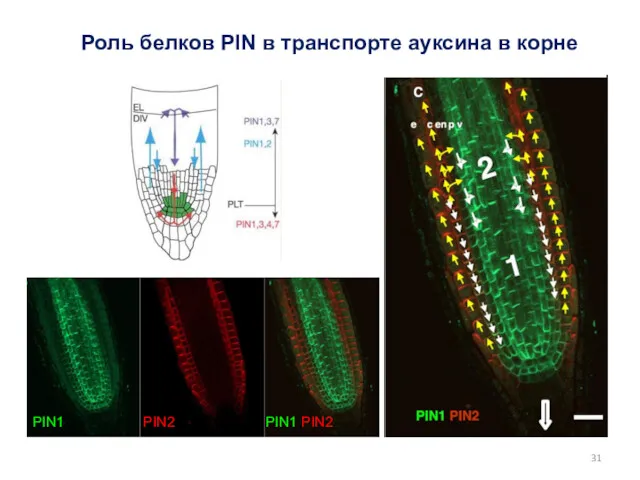
Роль белков PIN в транспорте ауксина в корне
PIN1 PIN2 PIN1 PIN2
Роль белков PIN в транспорте ауксина в корне
PIN1 PIN2 PIN1 PIN2
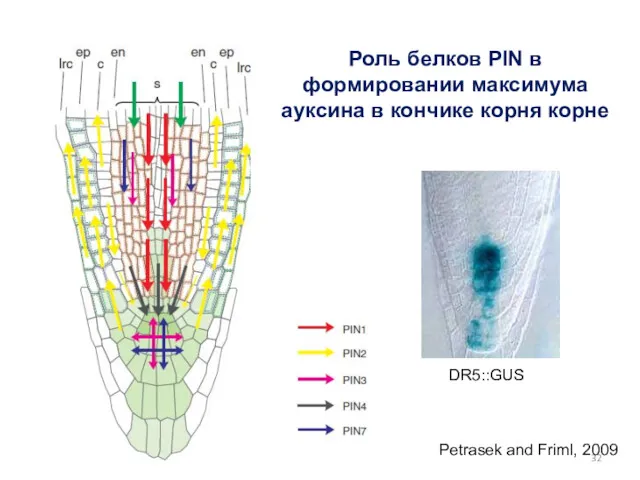
Petrasek and Friml, 2009
Роль белков PIN в формировании максимума ауксина
Petrasek and Friml, 2009
Роль белков PIN в формировании максимума ауксина

Нарушение апикально-базальной структуры корня у мутантов по генам, вовлеченным в метаболизм
Нарушение апикально-базальной структуры корня у мутантов по генам, вовлеченным в метаболизм
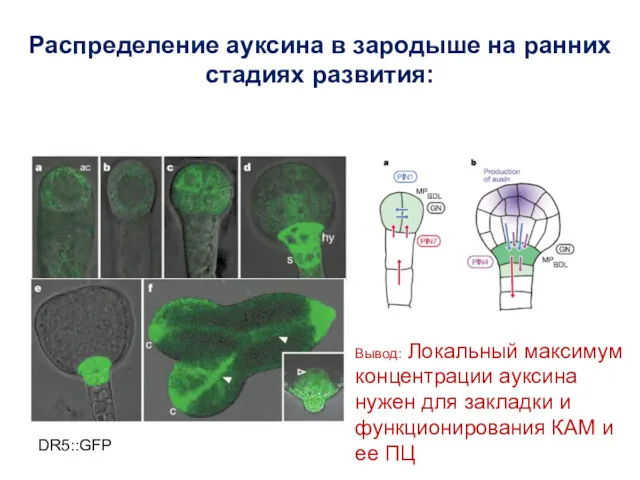
DR5::GFP
Распределение ауксина в зародыше на ранних стадиях развития:
Вывод: Локальный максимум концентрации
DR5::GFP
Распределение ауксина в зародыше на ранних стадиях развития:
Вывод: Локальный максимум концентрации
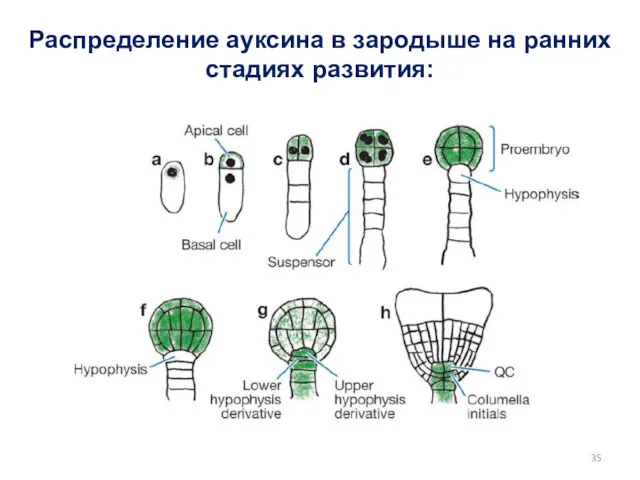
Распределение ауксина в зародыше на ранних стадиях развития:
Распределение ауксина в зародыше на ранних стадиях развития:
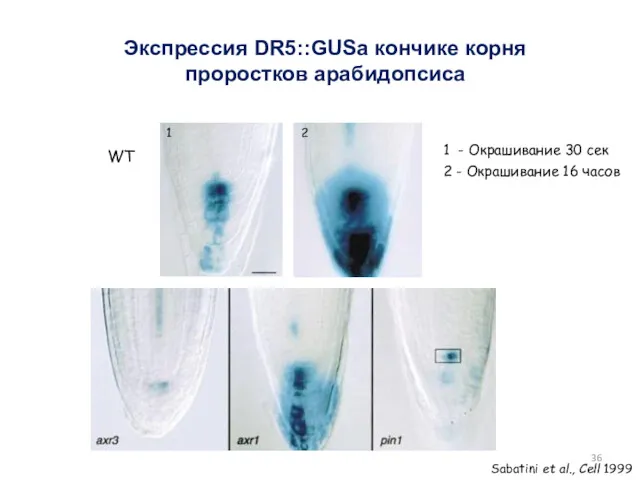
Экспрессия DR5::GUSа кончике корня проростков арабидопсиса
1 - Окрашивание 30 сек
2 -
Экспрессия DR5::GUSа кончике корня проростков арабидопсиса
1 - Окрашивание 30 сек
2 -
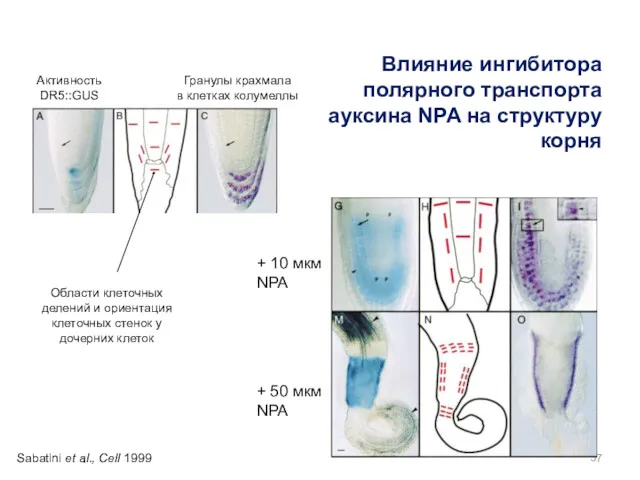
Области клеточных делений и ориентация клеточных стенок у дочерних клеток
+
Области клеточных делений и ориентация клеточных стенок у дочерних клеток
+
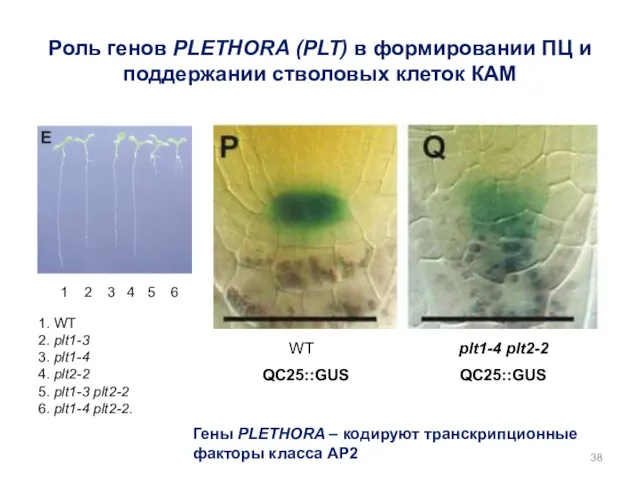
1. WT
2. plt1-3
3. plt1-4
4. plt2-2
5. plt1-3 plt2-2
6.
1. WT
2. plt1-3
3. plt1-4
4. plt2-2
5. plt1-3 plt2-2
6.
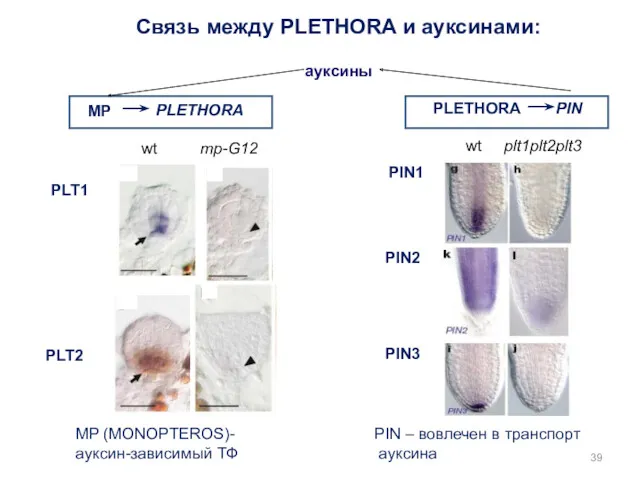
PLETHORA
MP
PIN
plt1plt2plt3
wt
PLT1
PLT2
PIN1
PIN3
PIN2
mp-G12
wt
PLETHORA
Связь между PLETHORA и ауксинами:
MP (MONOPTEROS)-
ауксин-зависимый ТФ
PIN – вовлечен в
PLETHORA
MP
PIN
plt1plt2plt3
wt
PLT1
PLT2
PIN1
PIN3
PIN2
mp-G12
wt
PLETHORA
Связь между PLETHORA и ауксинами:
MP (MONOPTEROS)-
ауксин-зависимый ТФ
PIN – вовлечен в
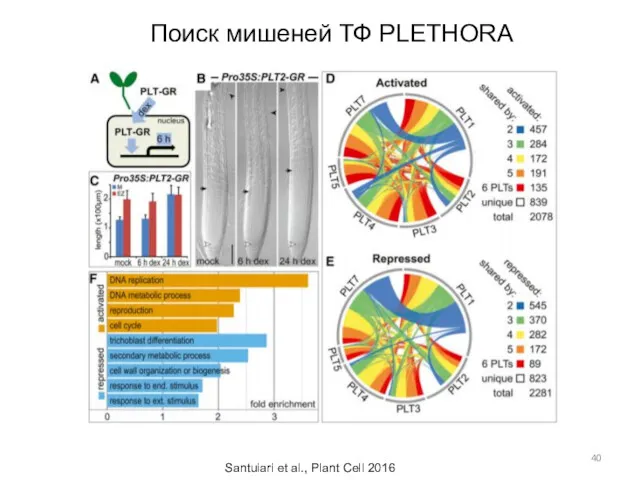
Поиск мишеней ТФ PLETHORA
Santuiari et al., Plant Cell 2016
Поиск мишеней ТФ PLETHORA
Santuiari et al., Plant Cell 2016
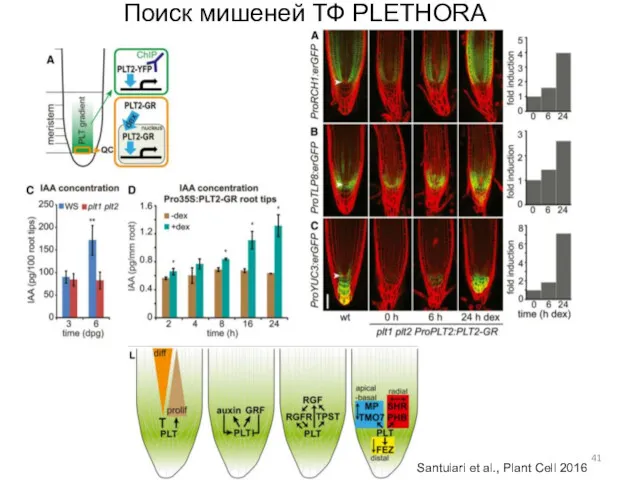
Поиск мишеней ТФ PLETHORA
Santuiari et al., Plant Cell 2016
Поиск мишеней ТФ PLETHORA
Santuiari et al., Plant Cell 2016
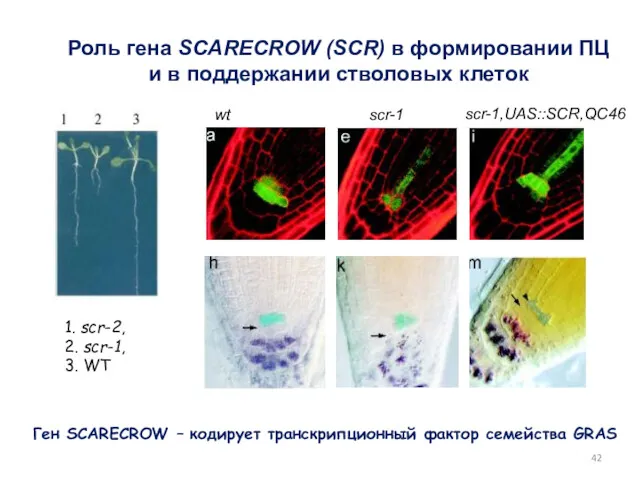
scr-1
wt
Роль гена SCARECROW (SCR) в формировании ПЦ и в поддержании стволовых
scr-1
wt
Роль гена SCARECROW (SCR) в формировании ПЦ и в поддержании стволовых
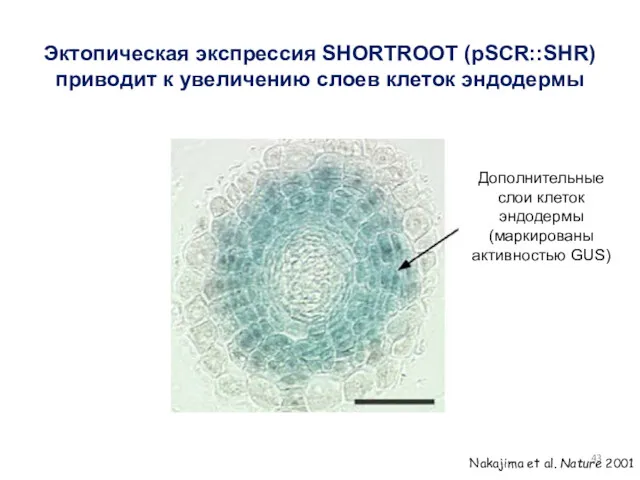
Эктопическая экспрессия SHORTROOT (pSCR::SHR) приводит к увеличению слоев клеток эндодермы
Дополнительные
Эктопическая экспрессия SHORTROOT (pSCR::SHR) приводит к увеличению слоев клеток эндодермы
Дополнительные
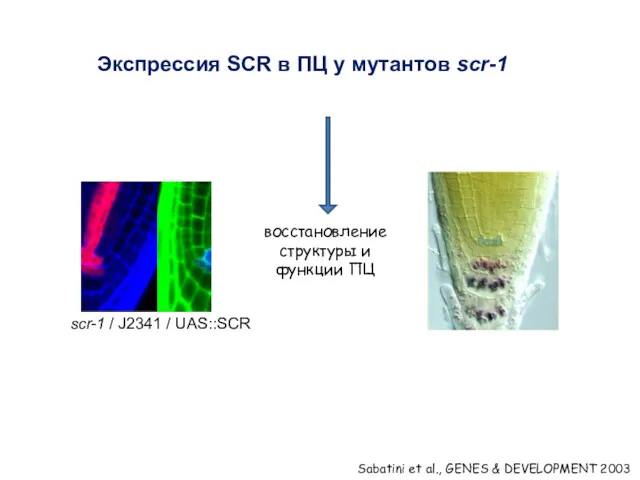
Экспрессия SCR в ПЦ у мутантов scr-1
восстановление структуры и функции ПЦ
Sabatini
Экспрессия SCR в ПЦ у мутантов scr-1
восстановление структуры и функции ПЦ
Sabatini
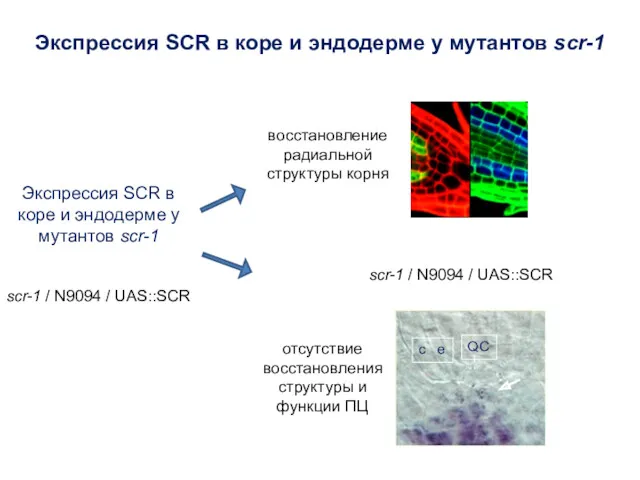
scr-1 / N9094 / UAS::SCR
Экспрессия SCR в коре и эндодерме у
scr-1 / N9094 / UAS::SCR
Экспрессия SCR в коре и эндодерме у
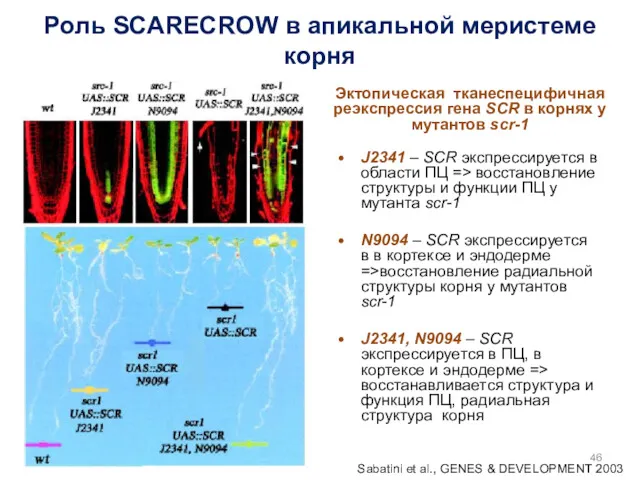
Роль SCARECROW в апикальной меристеме корня
J2341 – SCR экспрессируется в
Роль SCARECROW в апикальной меристеме корня
J2341 – SCR экспрессируется в
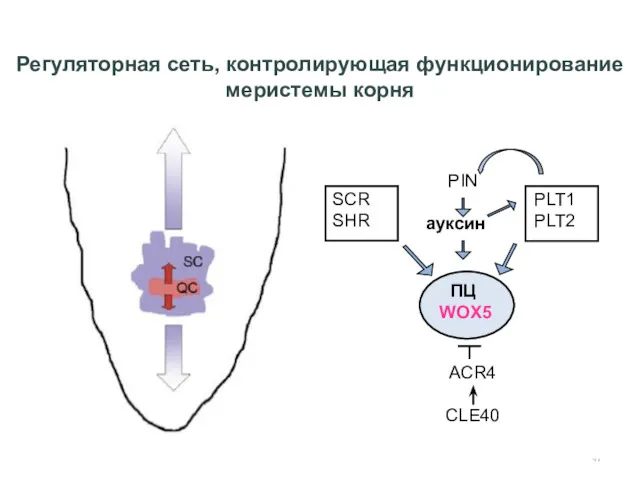
Регуляторная сеть, контролирующая функционирование меристемы корня
Sablowski, Current Opinion in Plant Biology
Регуляторная сеть, контролирующая функционирование меристемы корня
Sablowski, Current Opinion in Plant Biology
ФОРМИРОВАНИЕ РАДИАЛЬНОЙ СТРУКТУРЫ КОРНЯ
ФОРМИРОВАНИЕ РАДИАЛЬНОЙ СТРУКТУРЫ КОРНЯ
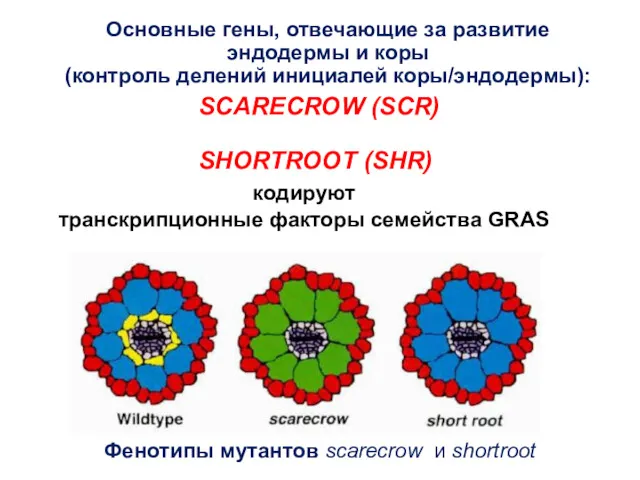
кодируют
транскрипционные факторы семейства GRAS
Основные гены, отвечающие за развитие
эндодермы
транскрипционные факторы семейства GRAS
Основные гены, отвечающие за развитие
эндодермы
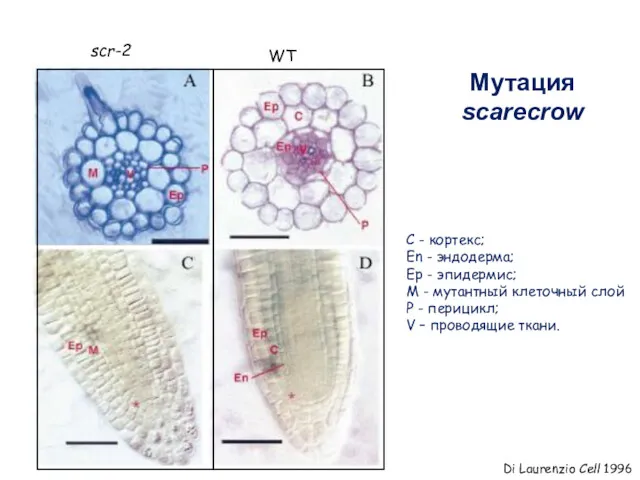
Мутация scarecrow
C - кортекс;
En - эндодерма;
Ep - эпидермис;
M -
Мутация scarecrow
C - кортекс;
En - эндодерма;
Ep - эпидермис;
M -
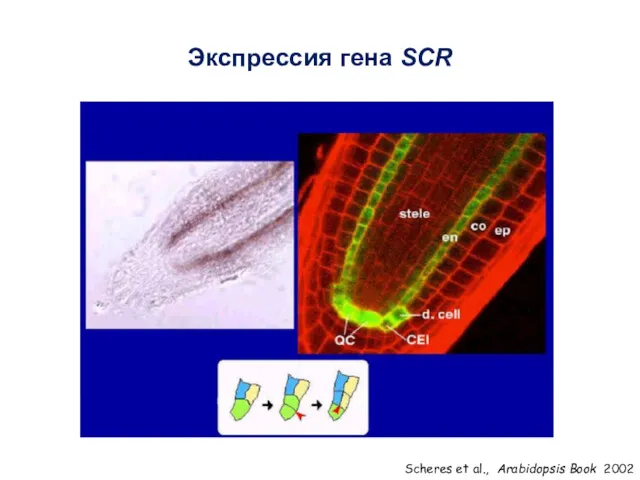
Scheres et al., Arabidopsis Book 2002
Экспрессия гена SCR
Scheres et al., Arabidopsis Book 2002
Экспрессия гена SCR
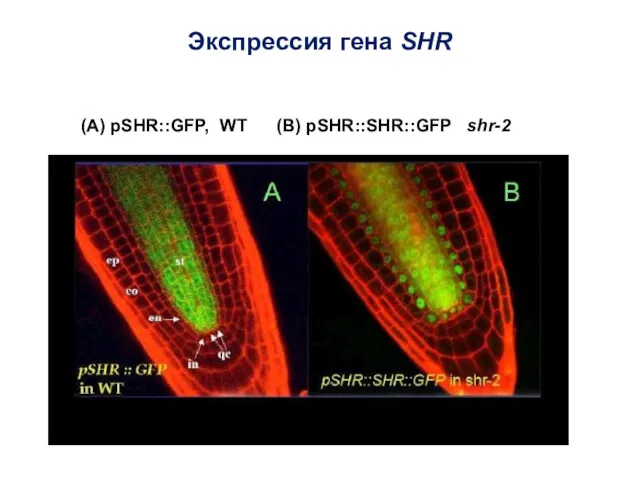
Экспрессия гена SHR
(А) pSHR::GFP, WT (В) pSHR::SHR::GFP shr-2
Экспрессия гена SHR
(А) pSHR::GFP, WT (В) pSHR::SHR::GFP shr-2
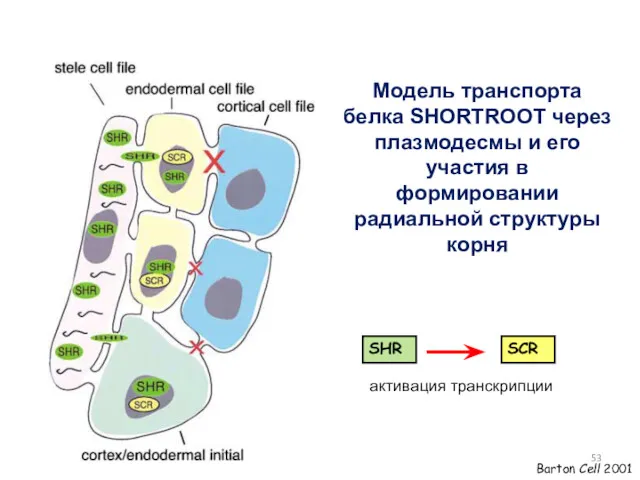
Модель транспорта белка SHORTROOT через плазмодесмы и его участия в формировании
Модель транспорта белка SHORTROOT через плазмодесмы и его участия в формировании
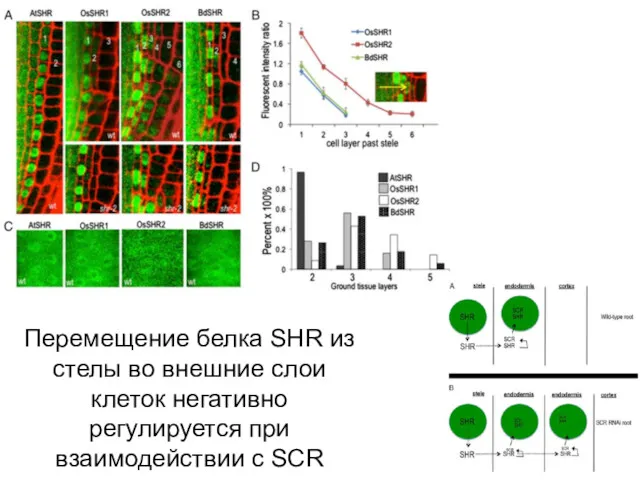
Перемещение белка SHR из стелы во внешние слои клеток негативно регулируется
Перемещение белка SHR из стелы во внешние слои клеток негативно регулируется

Гены-мишени SCR и SHR
Sozzani et al., 2010
CYCD6;1
SHR
Гены-мишени SCR и SHR
Sozzani et al., 2010
CYCD6;1
SHR
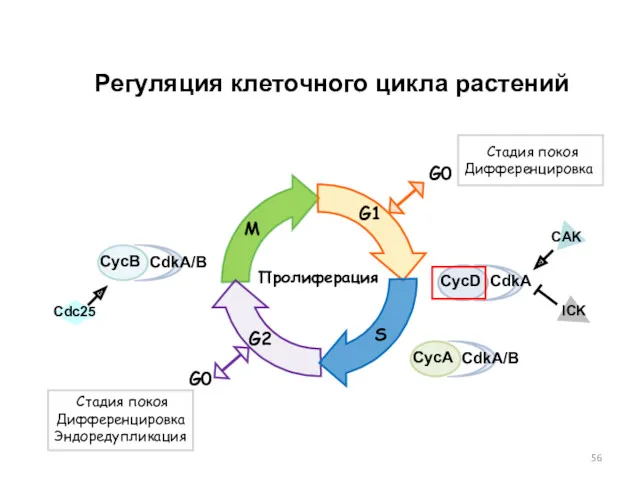
Регуляция клеточного цикла растений
Регуляция клеточного цикла растений
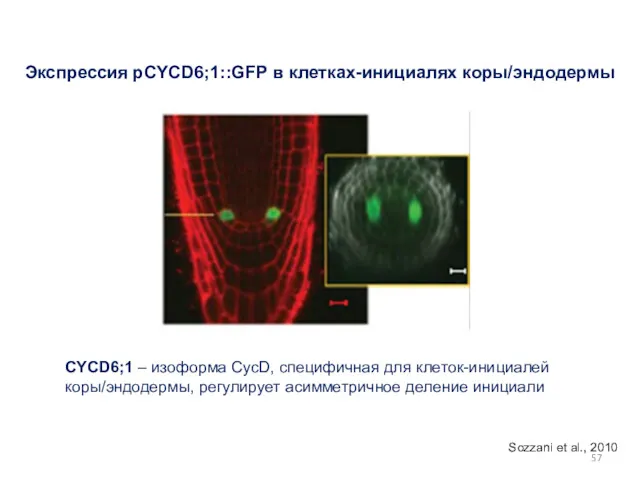
Экспрессия pCYCD6;1::GFP в клетках-инициалях коры/эндодермы
CYCD6;1 – изоформа СycD, специфичная для клеток-инициалей
Экспрессия pCYCD6;1::GFP в клетках-инициалях коры/эндодермы
CYCD6;1 – изоформа СycD, специфичная для клеток-инициалей
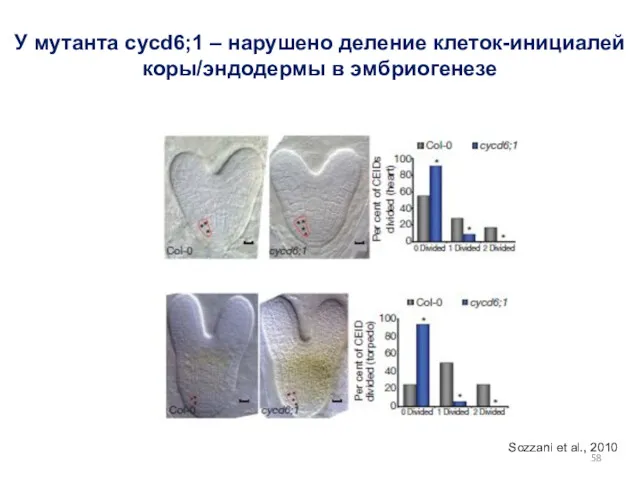
У мутанта cycd6;1 – нарушено деление клеток-инициалей коры/эндодермы в эмбриогенезе
Sozzani et
У мутанта cycd6;1 – нарушено деление клеток-инициалей коры/эндодермы в эмбриогенезе
Sozzani et

Генетический контроль дифференцировки клеток эпидермы
Генетический контроль дифференцировки клеток эпидермы
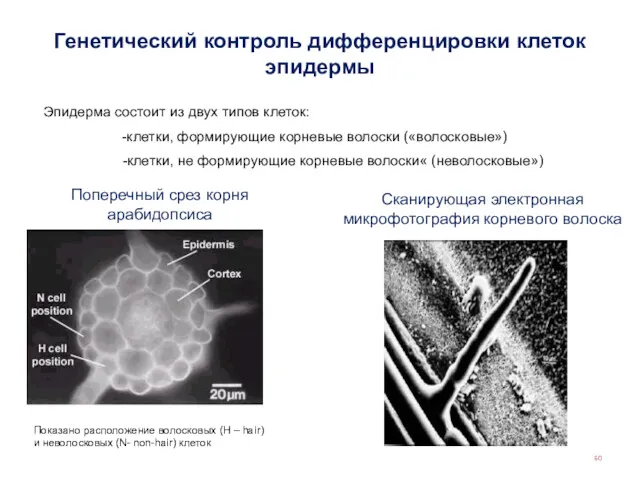
Генетический контроль дифференцировки клеток эпидермы
Поперечный срез корня арабидопсиса
Сканирующая электронная микрофотография корневого
Генетический контроль дифференцировки клеток эпидермы
Поперечный срез корня арабидопсиса
Сканирующая электронная микрофотография корневого
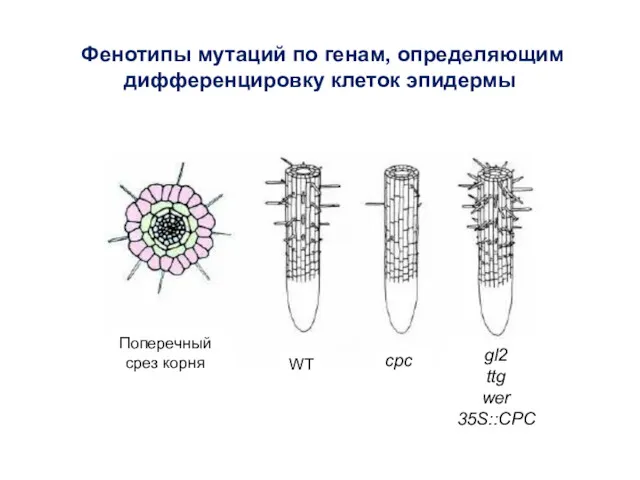
Фенотипы мутаций по генам, определяющим дифференцировку клеток эпидермы
WT
gl2
ttg
wer
35S::CPC
cpc
Поперечный срез корня
Фенотипы мутаций по генам, определяющим дифференцировку клеток эпидермы
WT
gl2
ttg
wer
35S::CPC
cpc
Поперечный срез корня

Фенотипы мутаций по генам, определяющим дифференцировку клеток эпидермы
A. Wild type
Фенотипы мутаций по генам, определяющим дифференцировку клеток эпидермы
A. Wild type

Экспрессия в эпидерме
Основные гены, вовлеченные в контроль дифференцировки клеток эпидермы у
Экспрессия в эпидерме
Основные гены, вовлеченные в контроль дифференцировки клеток эпидермы у
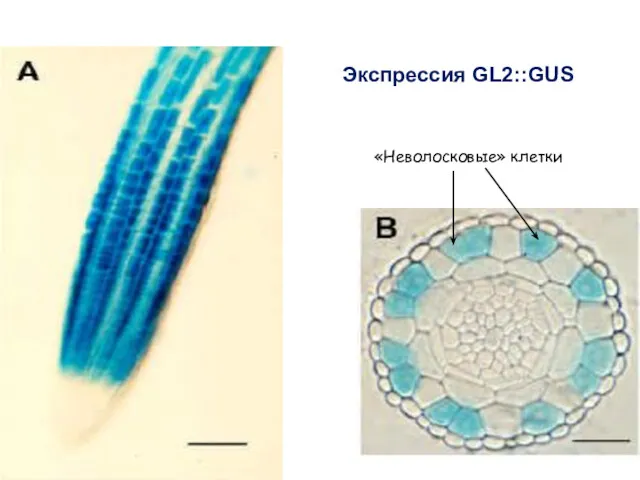
Экспрессия GL2::GUS
«Неволосковые» клетки
Экспрессия GL2::GUS
«Неволосковые» клетки
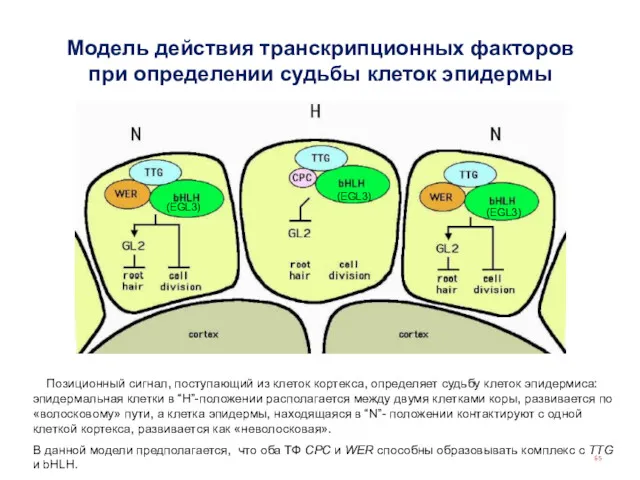
Позиционный сигнал, поступающий из клеток кортекса, определяет судьбу клеток эпидермиса:
Позиционный сигнал, поступающий из клеток кортекса, определяет судьбу клеток эпидермиса:
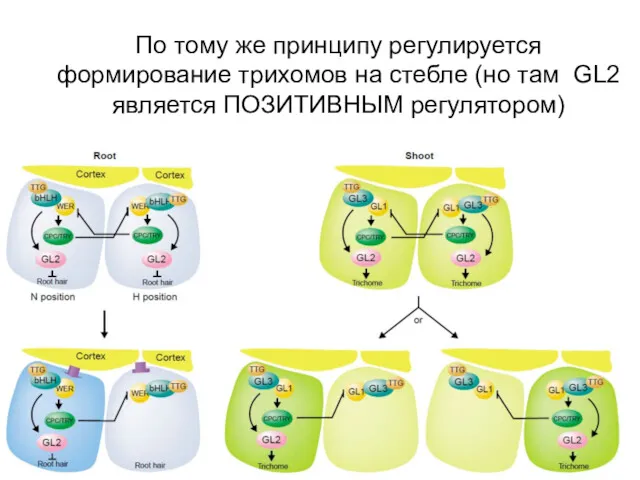
По тому же принципу регулируется формирование трихомов на стебле (но там
По тому же принципу регулируется формирование трихомов на стебле (но там
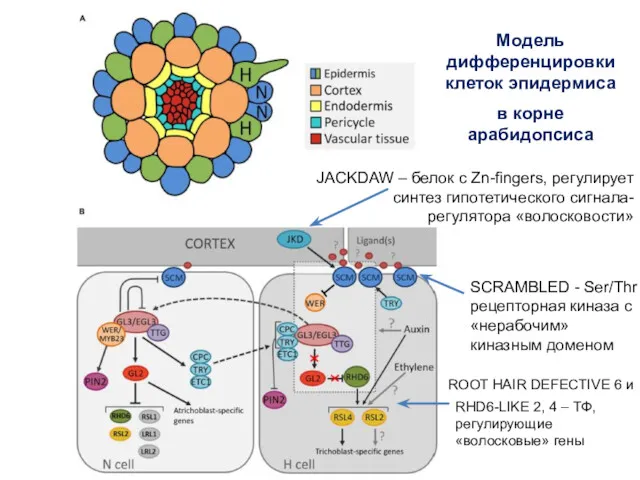
Модель дифференцировки клеток эпидермиса
в корне арабидопсиса
JACKDAW – белок с Zn-fingers,
Модель дифференцировки клеток эпидермиса
в корне арабидопсиса
JACKDAW – белок с Zn-fingers,
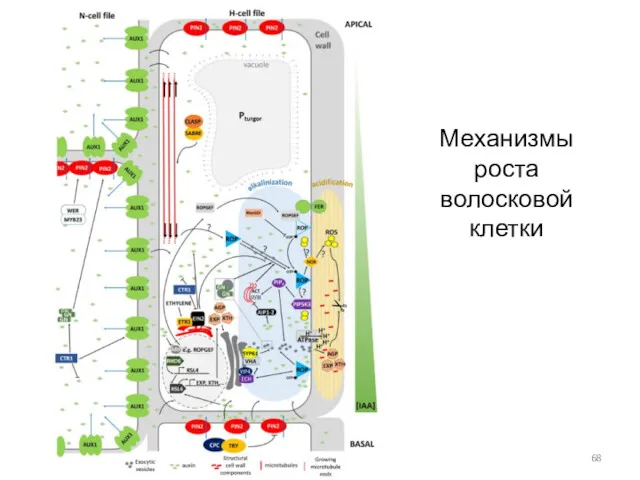
Механизмы роста волосковой клетки
Механизмы роста волосковой клетки
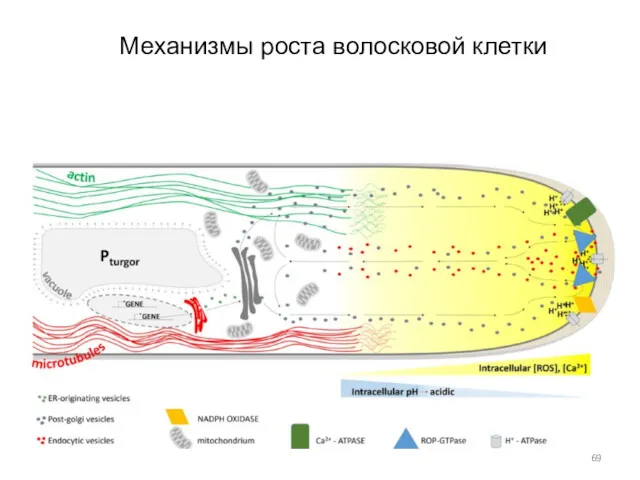
Механизмы роста волосковой клетки
Механизмы роста волосковой клетки
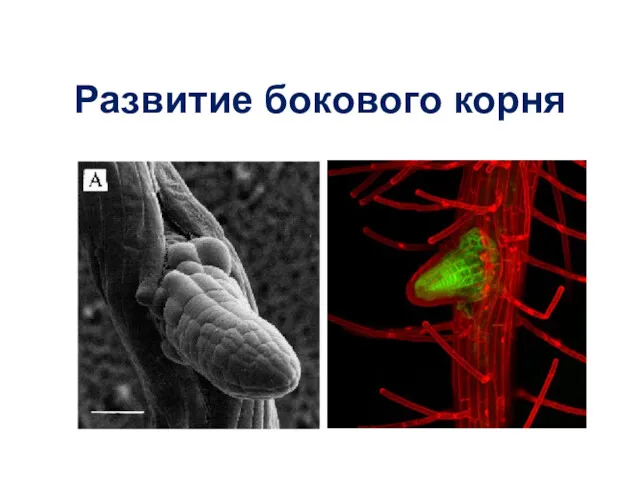
Развитие бокового корня
Развитие бокового корня
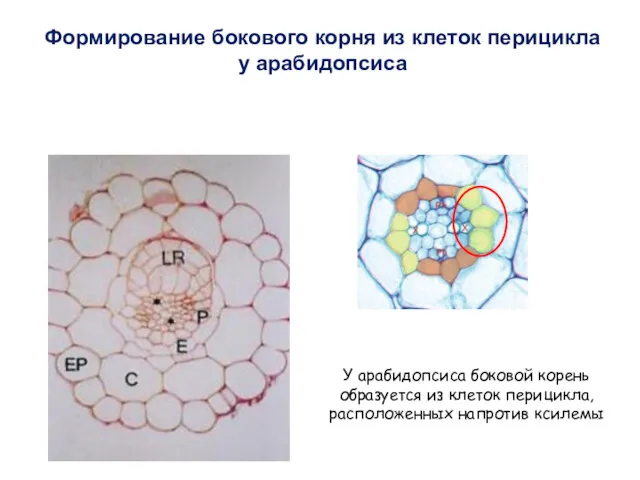
Формирование бокового корня из клеток перицикла
у арабидопсиса
У арабидопсиса боковой корень образуется
Формирование бокового корня из клеток перицикла
у арабидопсиса
У арабидопсиса боковой корень образуется
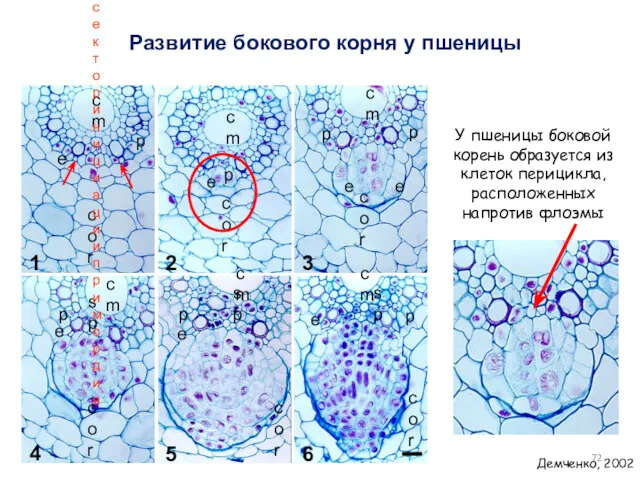
Развитие бокового корня у пшеницы
Демченко, 2002
У пшеницы боковой корень образуется из
Развитие бокового корня у пшеницы
Демченко, 2002
У пшеницы боковой корень образуется из
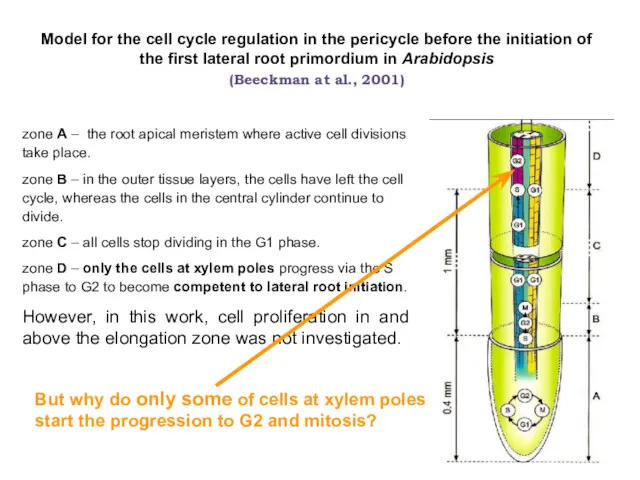
zone A – the root apical meristem where active cell divisions
zone A – the root apical meristem where active cell divisions
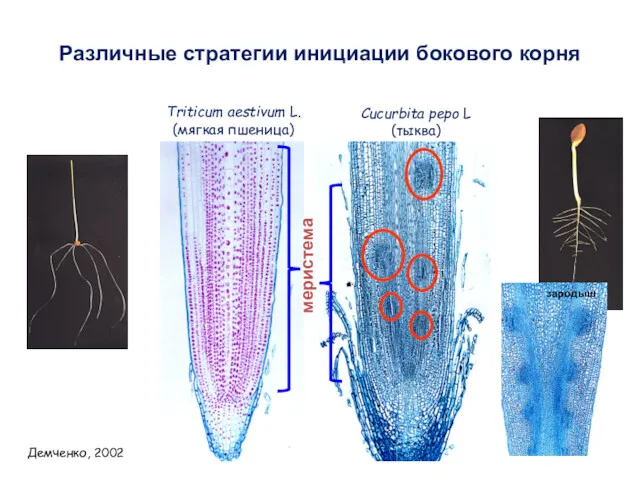
меристема
Triticum aestivum L.
(мягкая пшеница)
Cucurbita pepo L
(тыква)
Различные стратегии инициации бокового корня
Демченко,
меристема
Triticum aestivum L.
(мягкая пшеница)
Cucurbita pepo L
(тыква)
Различные стратегии инициации бокового корня
Демченко,
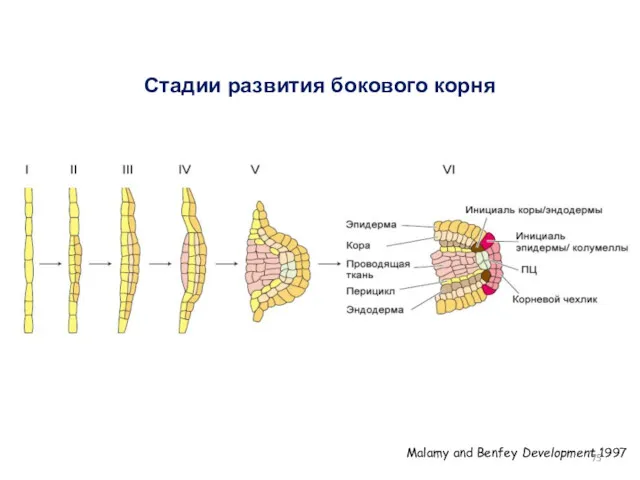
Стадии развития бокового корня
Malamy and Benfey Development 1997
Стадии развития бокового корня
Malamy and Benfey Development 1997
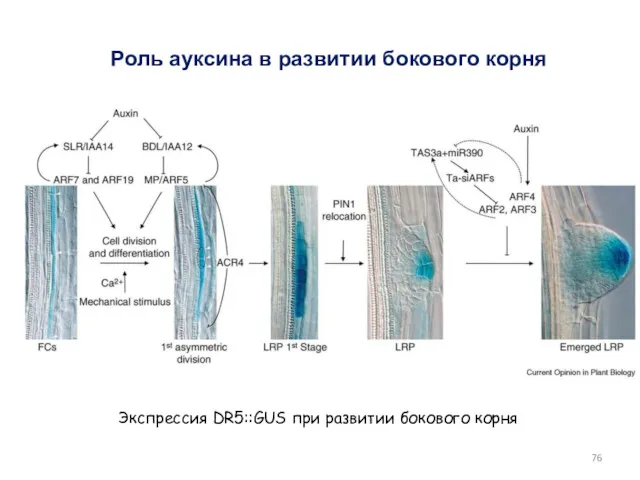
Экспрессия DR5::GUS при развитии бокового корня
Роль ауксина в развитии бокового корня
Экспрессия DR5::GUS при развитии бокового корня
Роль ауксина в развитии бокового корня
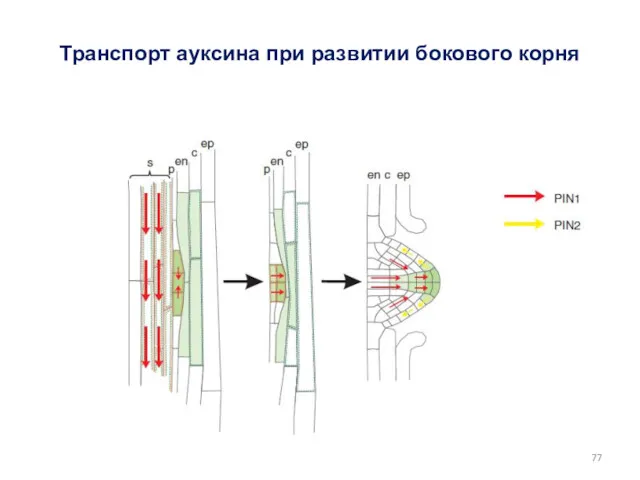
Транспорт ауксина при развитии бокового корня
Транспорт ауксина при развитии бокового корня
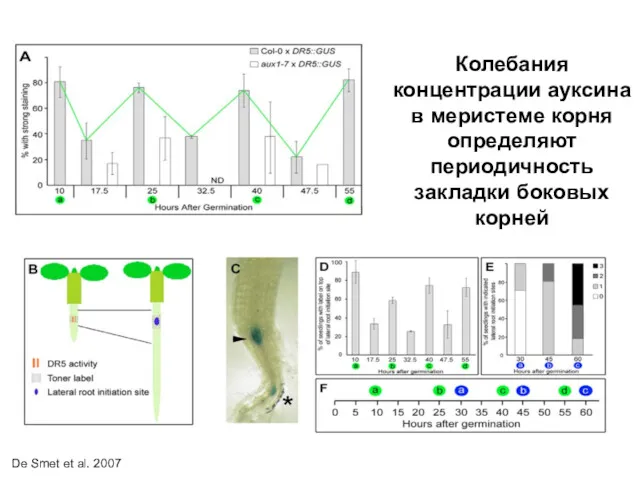
Колебания концентрации ауксина в меристеме корня определяют периодичность закладки боковых корней
De
Колебания концентрации ауксина в меристеме корня определяют периодичность закладки боковых корней
De
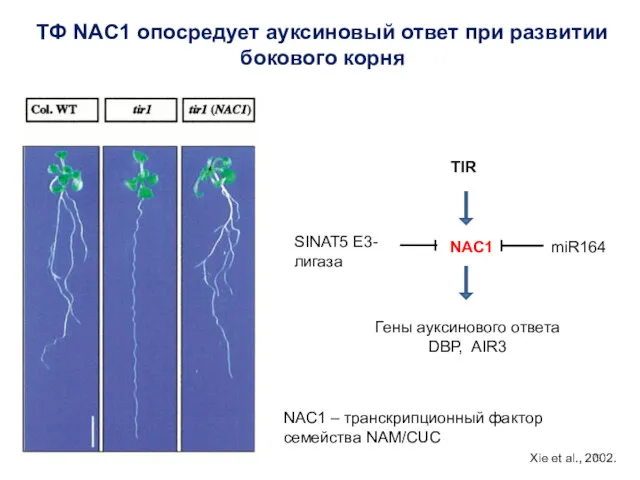
ТФ NAC1 опосредует ауксиновый ответ при развитии бокового корня
Гены ауксинового
ТФ NAC1 опосредует ауксиновый ответ при развитии бокового корня
Гены ауксинового
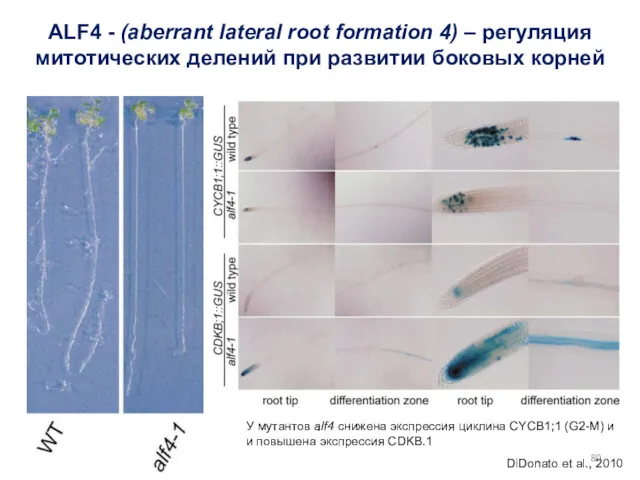
У мутантов alf4 снижена экспрессия циклина CYCB1;1 (G2-M) и
и повышена экспрессия
У мутантов alf4 снижена экспрессия циклина CYCB1;1 (G2-M) и
и повышена экспрессия

Основные гормоны, регулирующие развитие корня:
-ауксин (формирование покоящегося центра, стимуляция развития боковых
Основные гормоны, регулирующие развитие корня:
-ауксин (формирование покоящегося центра, стимуляция развития боковых
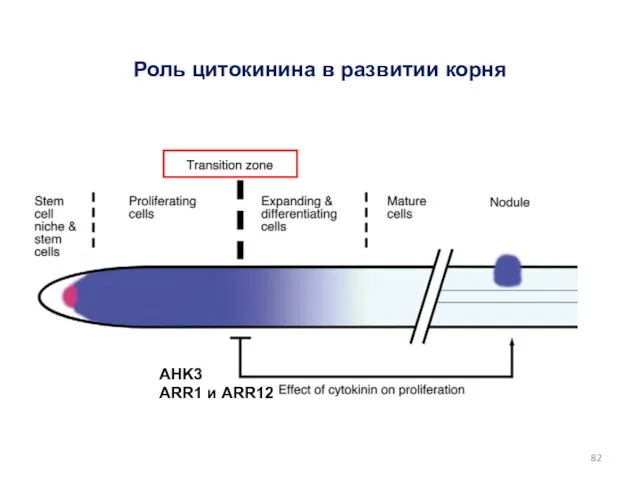
AHK3
ARR1 и ARR12
Роль цитокинина в развитии корня
AHK3
ARR1 и ARR12
Роль цитокинина в развитии корня
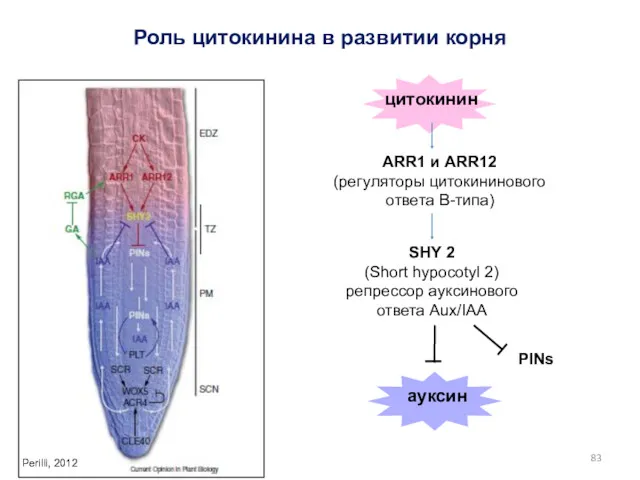
Роль цитокинина в развитии корня
цитокинин
Perilli, 2012
ARR1 и ARR12
(регуляторы цитокининового ответа
Роль цитокинина в развитии корня
цитокинин
Perilli, 2012
ARR1 и ARR12
(регуляторы цитокининового ответа
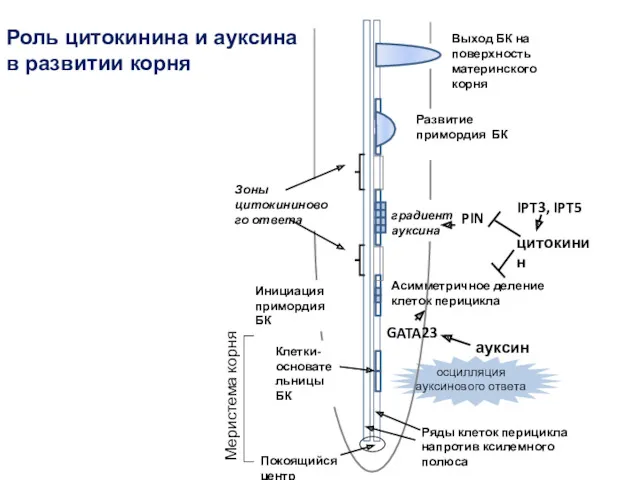
Асимметричное деление клеток перицикла
градиент ауксина
осцилляция ауксинового ответа
Клетки-основательницы БК
GATA23
Выход БК на поверхность
Асимметричное деление клеток перицикла
градиент ауксина
осцилляция ауксинового ответа
Клетки-основательницы БК
GATA23
Выход БК на поверхность
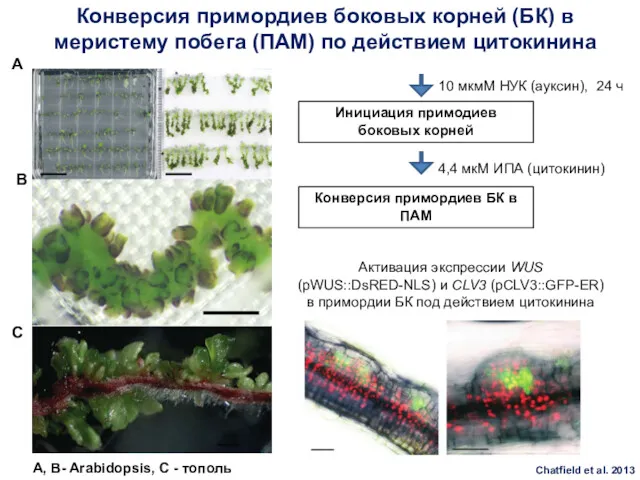
Конверсия примордиев боковых корней (БК) в меристему побега (ПАМ) по действием
Конверсия примордиев боковых корней (БК) в меристему побега (ПАМ) по действием
 Экология растений. Жизненные формы и стратегии (Лекция №6)
Экология растений. Жизненные формы и стратегии (Лекция №6)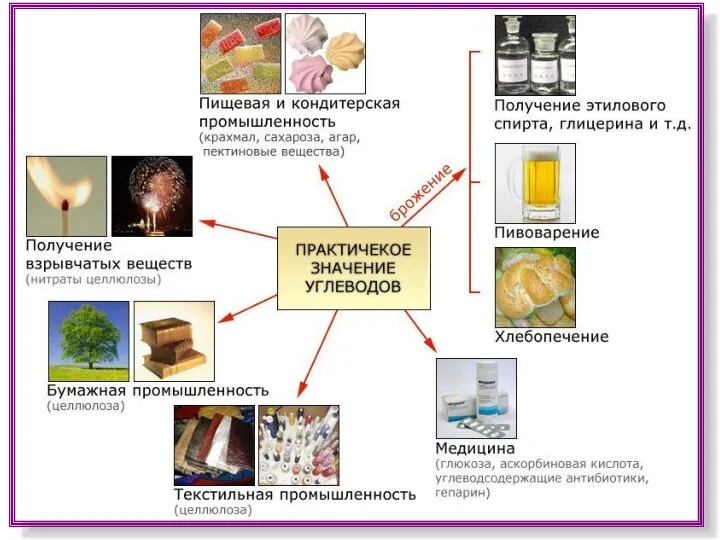 Практическое значение углеводов
Практическое значение углеводов Развитие индюка от яйца до взрослой особи
Развитие индюка от яйца до взрослой особи Тип Кольчатые черви. класс малощетинковые черви
Тип Кольчатые черви. класс малощетинковые черви Быстрота, как физическое качество человека
Быстрота, как физическое качество человека Эмбриология. Нейруляция
Эмбриология. Нейруляция История первобытного общества. Антропосоциогенез. Лекция 1
История первобытного общества. Антропосоциогенез. Лекция 1 Белки. Превращение белков, жиров и углеводов
Белки. Превращение белков, жиров и углеводов Физиологические основы психики
Физиологические основы психики Групповой проект как средство формирование коммуникативных УУД.
Групповой проект как средство формирование коммуникативных УУД. Эмбриогенез головного мозга
Эмбриогенез головного мозга Значение бактерий в природе и для человека
Значение бактерий в природе и для человека Молекулярно-генетические методы диагностики
Молекулярно-генетические методы диагностики Нервная регуляция
Нервная регуляция Покровы тела. Сравнительная характеристика
Покровы тела. Сравнительная характеристика Роль пресмыкающихся в природе и в жизни человека. Охрана пресмыкающихся. Древние пресмыкающиеся
Роль пресмыкающихся в природе и в жизни человека. Охрана пресмыкающихся. Древние пресмыкающиеся Необычные рыбы
Необычные рыбы Царство грибы
Царство грибы Белка - лесной зверек
Белка - лесной зверек Придатки кожи
Придатки кожи Формирование исследовательских умений и навыков на уроках биологии и химии и во внеурочное время
Формирование исследовательских умений и навыков на уроках биологии и химии и во внеурочное время Семейство Бобовые: общая характеристика, многообразие и значение
Семейство Бобовые: общая характеристика, многообразие и значение Теория эволюции. Промежуточные этапы развития
Теория эволюции. Промежуточные этапы развития Многообразие организмов и их классификация. Систематика и её задачи
Многообразие организмов и их классификация. Систематика и её задачи DNA RNA Protein
DNA RNA Protein Ядовитые грибы и растения
Ядовитые грибы и растения Структура и функции клетки. Цитоплазма. Митохондрии, пластиды, органоиды движения, включения. (Глава 2.9)
Структура и функции клетки. Цитоплазма. Митохондрии, пластиды, органоиды движения, включения. (Глава 2.9) Классы и семейства покрытосеменных растений
Классы и семейства покрытосеменных растений