- Теория эволюции. Промежуточные этапы развития

Содержание
- 2. Система взглядов на историю мироздания далеко не новая и трудно сказать, что появилось раньше: проблески эволюционного
- 3. Существует много примеров, когда то или иное, наблюдаемое в природе, явление не находит объяснений с позиций
- 4. Синтетическая теория эволюции В настоящее время СТЭ можно считать и наиболее разработанной, и имеющей наибольшее число
- 5. 2. Материалом для эволюции служат мелкие дискретные наследственные изменения – мутации; среди них преимущественное внимание уделяется
- 6. 4. Эволюция носит дивергентный характер, т.е. от одного таксона могут возникнуть несколько дочерних таксонов. Если говорить
- 7. 6. Эволюция носит постепенный (градуалистический) характер, и ее результаты могут быть замечены лишь по прошествии длительных
- 8. 8. Вид представляет собой сложную полиморфную систему, составленную множеством соподчиненных единиц – подвидов и популяций. Такое
- 9. Нам представляется нецелесообразным выделять эту констатацию фактов в отдельный постулат, так как это всего лишь иллюстрация
- 10. Этот постулат СТЭ также не выдерживает критики. В свете новейших данных законы индивидуального и исторического развития
- 11. Далее В.И. Назаров высказывает следующую мысль: «Возможно, … проницательный читатель сможет убедиться, что в объяснении целого
- 12. Критика синтетической теории эволюции по В.А. Красилову СТЭ в большей степени, чем классический дарвинизм, построена по
- 13. Я уже пытался объяснить в предыдущем разделе, почему я не считаю оцепенение науки, скованной аксиомами и
- 14. В СТЭ с её «популяционным мышлением» редукции подвергся в первую очередь организм (отсюда невнимание к индивидуальной
- 15. Какова роль этих экспериментов в развитии биологии и, в частности, теории эволюции? Мне она представляется второстепенной,
- 16. СТЭ принадлежит к числу сверхсильных парадигм, которые столь успешно подавляют конкурирующие теории, что продолжают удерживать свои
- 17. Ничуть не преуменьшая значение индустриального меланизма и отношений между однотонными и полосатыми улитками, отметим, что они
- 18. СТЭ, как и классическая эволюционная теория Дарвина, разработана главным образом для процессов, протекающих в устойчивых условиях.
- 19. Возникновение Земли и жизни на ней Формирование Земли Сведения, содержащиеся в этом разделе, почерпнуты в основном
- 20. Известно, что почти 9/10 массы Земли приходится на долю всего четырёх химических элементов: кислорода, входящего в
- 21. Расчеты геофизиков показывают, что эта энергия составляет чудовищную величину 4х1030 кал (что эквивалентно триллиону суммарных ядерных
- 22. В толще этой «жидкости» постоянно происходят чрезвычайно медленные, но немыслимо мощные движения колоссальных масс вещества, с
- 23. Структура недр Земли со схематическим вулканом (из Еськов, 2014)
- 24. Между корой и ядром располагается «промежуточный» слой – мантия; ее породы имеют плотность около 3,5 г/см3
- 25. Первичная атмосфера была еще очень тонкой, температура на поверхности Земли равнялась температуре лучистого равновесия, получающейся при
- 26. Как же изменялся объем гидросферы на протяжении ее истории? В расплавленном базальте (в астеносфере) при температуре
- 27. Если спросить человека: «Отчего море соленое?», он почти наверняка ответит: «Оттого же, отчего солоны бессточные озера
- 28. Что касается атмосферы, то ее состав стал меняться в протерозое, когда фотосинтезирующие организмы начали вырабатывать (в
- 29. Помимо кислорода и азота, в атмосфере содержится небольшое количество так называемых парниковых газов – углекислый газ,
- 30. Возникновение жизни на Земле Следуя далее нити повествования К.Ю. Еськова, мы принимаем, что эволюция биосферы и
- 31. В 20-е годы прошлого века А.И. Опарин и Дж. Холдейн экспериментально показали, что в растворах высокомолекулярных
- 32. Многие органические соединения представляют собой смесь двух оптических изомеров – веществ, имеющих совершенно одинаковые химические свойства,
- 33. Между тем даже успешный синтез «живых» макромолекул (до которого еще, как говорится, «семь верст – и
- 34. Сейчас всем понятно, что это просто несерьезно: потребное для этого время (его вполне можно рассчитать) намного
- 35. Концепцию панспермии обычно упрекают в том, что она не дает принципиального ответа на вопрос о путях
- 37. Скачать презентацию

Система взглядов на историю мироздания далеко не новая и трудно сказать,
Система взглядов на историю мироздания далеко не новая и трудно сказать,

Существует много примеров, когда то или иное, наблюдаемое в природе, явление
Существует много примеров, когда то или иное, наблюдаемое в природе, явление

Синтетическая теория эволюции
В настоящее время СТЭ можно считать и наиболее разработанной,
Синтетическая теория эволюции
В настоящее время СТЭ можно считать и наиболее разработанной,

2. Материалом для эволюции служат мелкие дискретные наследственные изменения – мутации;
2. Материалом для эволюции служат мелкие дискретные наследственные изменения – мутации;

4. Эволюция носит дивергентный характер, т.е. от одного таксона могут возникнуть
4. Эволюция носит дивергентный характер, т.е. от одного таксона могут возникнуть

6. Эволюция носит постепенный (градуалистический) характер, и ее результаты могут быть
6. Эволюция носит постепенный (градуалистический) характер, и ее результаты могут быть

8. Вид представляет собой сложную полиморфную систему, составленную множеством соподчиненных единиц
8. Вид представляет собой сложную полиморфную систему, составленную множеством соподчиненных единиц

Нам представляется нецелесообразным выделять эту констатацию фактов в отдельный постулат, так
Нам представляется нецелесообразным выделять эту констатацию фактов в отдельный постулат, так

Этот постулат СТЭ также не выдерживает критики. В свете новейших данных
Этот постулат СТЭ также не выдерживает критики. В свете новейших данных

Далее В.И. Назаров высказывает следующую мысль: «Возможно, … проницательный читатель сможет
Далее В.И. Назаров высказывает следующую мысль: «Возможно, … проницательный читатель сможет

Критика синтетической теории эволюции по В.А. Красилову
СТЭ в большей степени, чем
Критика синтетической теории эволюции по В.А. Красилову
СТЭ в большей степени, чем

Я уже пытался объяснить в предыдущем разделе, почему я не считаю
Я уже пытался объяснить в предыдущем разделе, почему я не считаю

В СТЭ с её «популяционным мышлением» редукции подвергся в первую очередь
В СТЭ с её «популяционным мышлением» редукции подвергся в первую очередь

Какова роль этих экспериментов в развитии биологии и, в частности, теории
Какова роль этих экспериментов в развитии биологии и, в частности, теории

СТЭ принадлежит к числу сверхсильных парадигм, которые столь успешно подавляют конкурирующие
СТЭ принадлежит к числу сверхсильных парадигм, которые столь успешно подавляют конкурирующие

Ничуть не преуменьшая значение индустриального меланизма и отношений между однотонными и
Ничуть не преуменьшая значение индустриального меланизма и отношений между однотонными и

СТЭ, как и классическая эволюционная теория Дарвина, разработана главным образом для
СТЭ, как и классическая эволюционная теория Дарвина, разработана главным образом для

Возникновение Земли и жизни на ней
Формирование Земли
Сведения, содержащиеся в этом разделе,
Возникновение Земли и жизни на ней
Формирование Земли
Сведения, содержащиеся в этом разделе,

Известно, что почти 9/10 массы Земли приходится на долю всего четырёх
Известно, что почти 9/10 массы Земли приходится на долю всего четырёх

Расчеты геофизиков показывают, что эта энергия составляет чудовищную величину 4х1030 кал
Расчеты геофизиков показывают, что эта энергия составляет чудовищную величину 4х1030 кал

В толще этой «жидкости» постоянно происходят чрезвычайно медленные, но немыслимо мощные
В толще этой «жидкости» постоянно происходят чрезвычайно медленные, но немыслимо мощные
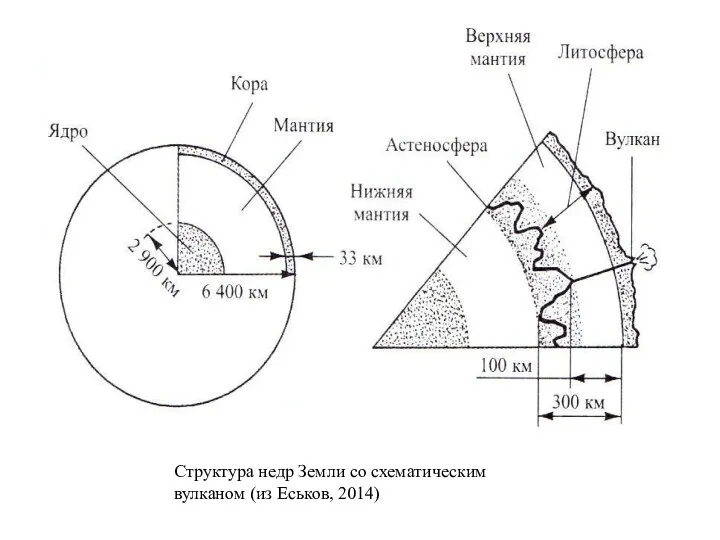
Структура недр Земли со схематическим вулканом (из Еськов, 2014)
Структура недр Земли со схематическим вулканом (из Еськов, 2014)

Между корой и ядром располагается «промежуточный» слой – мантия; ее породы
Между корой и ядром располагается «промежуточный» слой – мантия; ее породы

Первичная атмосфера была еще очень тонкой, температура на поверхности Земли равнялась
Первичная атмосфера была еще очень тонкой, температура на поверхности Земли равнялась

Как же изменялся объем гидросферы на протяжении ее истории? В расплавленном
Как же изменялся объем гидросферы на протяжении ее истории? В расплавленном

Если спросить человека: «Отчего море соленое?», он почти наверняка ответит: «Оттого
Если спросить человека: «Отчего море соленое?», он почти наверняка ответит: «Оттого

Что касается атмосферы, то ее состав стал меняться в протерозое, когда
Что касается атмосферы, то ее состав стал меняться в протерозое, когда

Помимо кислорода и азота, в атмосфере содержится небольшое количество так называемых
Помимо кислорода и азота, в атмосфере содержится небольшое количество так называемых

Возникновение жизни на Земле
Следуя далее нити повествования К.Ю. Еськова, мы принимаем,
Возникновение жизни на Земле
Следуя далее нити повествования К.Ю. Еськова, мы принимаем,

В 20-е годы прошлого века А.И. Опарин и Дж. Холдейн экспериментально
В 20-е годы прошлого века А.И. Опарин и Дж. Холдейн экспериментально

Многие органические соединения представляют собой смесь двух оптических изомеров – веществ,
Многие органические соединения представляют собой смесь двух оптических изомеров – веществ,

Между тем даже успешный синтез «живых» макромолекул (до которого еще, как
Между тем даже успешный синтез «живых» макромолекул (до которого еще, как

Сейчас всем понятно, что это просто несерьезно: потребное для этого время
Сейчас всем понятно, что это просто несерьезно: потребное для этого время

Концепцию панспермии обычно упрекают в том, что она не дает принципиального
Концепцию панспермии обычно упрекают в том, что она не дает принципиального
 Выделительная система человека
Выделительная система человека Викторина для знатоков природы
Викторина для знатоков природы Белки
Белки Ризосферные плазмидосодержащие бактерии рода Pseudomonas для био/фиторемедиации загрязненных почв
Ризосферные плазмидосодержащие бактерии рода Pseudomonas для био/фиторемедиации загрязненных почв Определение свинца в золе пищевых продуктов
Определение свинца в золе пищевых продуктов Организмы царства грибы и лишайники
Организмы царства грибы и лишайники Семейство розоцветных.
Семейство розоцветных. Дүниесі: Жануарлар
Дүниесі: Жануарлар Ферменты, их свойства и классификация
Ферменты, их свойства и классификация Буферные системы. Классификация буферных растворов
Буферные системы. Классификация буферных растворов Анатомия женской половой системы человека
Анатомия женской половой системы человека Общая физиология сенсорных систем
Общая физиология сенсорных систем Культивирование растительных клеток
Культивирование растительных клеток Як бачать тварини
Як бачать тварини Жасуша және оның құрылысы
Жасуша және оның құрылысы Фотобиологические процессы. Основные определения
Фотобиологические процессы. Основные определения Птахи. Загальна характеристика птахів
Птахи. Загальна характеристика птахів How wonderful smells
How wonderful smells Общая характеристика грибов
Общая характеристика грибов 20231130_8_klass_ptitsy._obshchie_priznaki._osobennosti_stroeniya._1
20231130_8_klass_ptitsy._obshchie_priznaki._osobennosti_stroeniya._1 презентация по биологии на тему Основные понятия генетики для 9 класса
презентация по биологии на тему Основные понятия генетики для 9 класса Вода в жизни человека
Вода в жизни человека Устройство микроскопа и приёмы работы с ним. 5 класс
Устройство микроскопа и приёмы работы с ним. 5 класс Основы косметологии MIXIT (new)
Основы косметологии MIXIT (new) Грегор Мендель 1822 1884г.г
Грегор Мендель 1822 1884г.г Систематика растений. Многообразие растений
Систематика растений. Многообразие растений Половая система самок животных
Половая система самок животных Характеристика сложных белков. Структура и свойства
Характеристика сложных белков. Структура и свойства