- Молекулярные аспекты биологического действия ионизирующих излучений

Содержание
- 2. Радиационные повреждения ДНК При облучении клетки поражаются все ее структуры. Вероятность поражения тех или иных молекул
- 3. Радиационные повреждения ДНК Основой молекулы ДНК являются две нити (также называемых цепями, или цепочками), построенные из
- 4. Строение одной из двух нитей ДНК 1- сахаро-фосфатный скелет; 2- дезоксирибоза; 3- азотистые основания аденин, гуанин,
- 5. Радиационные повреждения ДНК В клетке различные участки ДНК одной и той же молекулы находятся очень близко
- 6. Радиационные повреждения ДНК Следующие уровни упаковки хроматина — петли шириной 300 нм, из которых формируются петли
- 7. Строение участка молекулы ДНК и ее пространственная упаковка в интерфазном ядре и (после конденсации) в хромосоме
- 8. Радиационные повреждения ДНК Однонитевое и двунитевое повреждения ДНК В результате прямой ионизации самой молекулы ДНК и
- 9. Радиационные повреждения ДНК Известно, что одиночные разрывы постоянно возникают в клетке и без всякой связи с
- 10. Радиационные повреждения ДНК Разрыв межатомных связей в сахаро-фосфатном скелете ведет к нарушению непрерывности молекулы ДНК, что
- 11. Механизм индукции и реализации смерти клетки (апоптоза) Еще одним следствием повреждения молекул ДНК является включение процесса
- 12. Механизм индукции и реализации смерти клетки (апоптоза) Именно путем апоптоза происходит удаление трансформированных клеток. В клетке
- 13. Механизм индукции и реализации смерти клетки (апоптоза) Очень четко развитие апоптоза проявляется при наблюдении за клетками,
- 14. АПОПТОЗ До начала апоптоза ядро имеет равномерную синюю окраску, указывающую на равномерное распределение ДНК по его
- 15. АПОПТОЗ Финальная стадия апоптоза
- 16. Механизм индукции и реализации смерти клетки (апоптоза) Клетки могут значительно различаться по времени гибели после облучения.
- 17. Последовательность ультраструктурных изменений при апоптозе (справа) и некрозе (слева) 1 – нормальная клетка; АПОПТОЗ 2 –
- 18. Морфологические проявления апоптоза Для клетки, подвергающейся апоптозу характерно: Сжатие клетки. Клетка уменьшается в размерах; цитоплазма уплотняется;
- 19. Морфологические проявления апоптоза Фагоцитоз апоптотических клеток или телец осуществляется окружающими здоровыми клетками, или паренхиматозными, или макрофагами.
- 20. Хромосомные аберрации Хромосомные аберрации (перестройки) являются классическим проявлением лучевого поражения клеток. Появление аберраций отражает образование разрывов
- 21. Схема строения хромосомы в поздней профазе — метафазе митоза 1—хроматида; 2—центромера; 3—короткое плечо; 4—длинное плечо. Отклонения
- 22. Хромосомные аберрации Неверное воссоединение разрывов, когда при репарации происходит соединение участков ДНК из разных мест одной
- 23. Механизм упорядоченного расположения хромосом в метафазе и их расхождение в анафазе при первом (a) и втором
- 24. Хромосомные аберрации Возникающие в клетке аберрации подразделяют на хромосомные и хроматидные. Хромосомные аберрации возникают в случае,
- 25. Хромосомные аберрации Разрыв обеих хроматид может проявиться различным образом — образованием двух фрагментов, их слиянием в
- 26. Хромосомные аберрации Схема образования робертсоновской транслокации (а), изохромосом (б) и кольцевой хромосомы (в), A и В
- 27. Хромосомные аберрации На фото: красным кружком обведены хромосомные аномалии - маркеры радиационного поражения клетки
- 29. Скачать презентацию

Радиационные повреждения ДНК
При облучении клетки поражаются все ее структуры.
Вероятность поражения
Радиационные повреждения ДНК
При облучении клетки поражаются все ее структуры.
Вероятность поражения

Радиационные повреждения ДНК
Основой молекулы ДНК являются две нити (также называемых цепями,
Радиационные повреждения ДНК
Основой молекулы ДНК являются две нити (также называемых цепями,
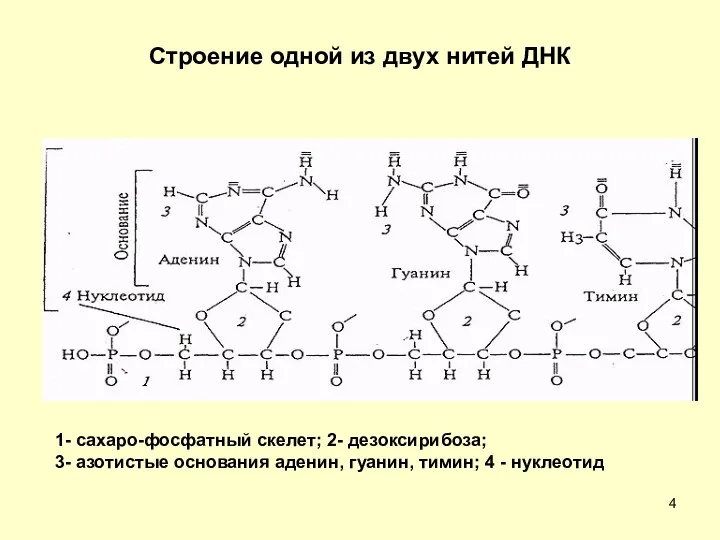
Строение одной из двух нитей ДНК
1- сахаро-фосфатный скелет; 2- дезоксирибоза;
3-
Строение одной из двух нитей ДНК
1- сахаро-фосфатный скелет; 2- дезоксирибоза; 3-

Радиационные повреждения ДНК
В клетке различные участки ДНК одной и той же
Радиационные повреждения ДНК
В клетке различные участки ДНК одной и той же

Радиационные повреждения ДНК
Следующие уровни упаковки хроматина — петли шириной 300 нм,
Радиационные повреждения ДНК
Следующие уровни упаковки хроматина — петли шириной 300 нм,
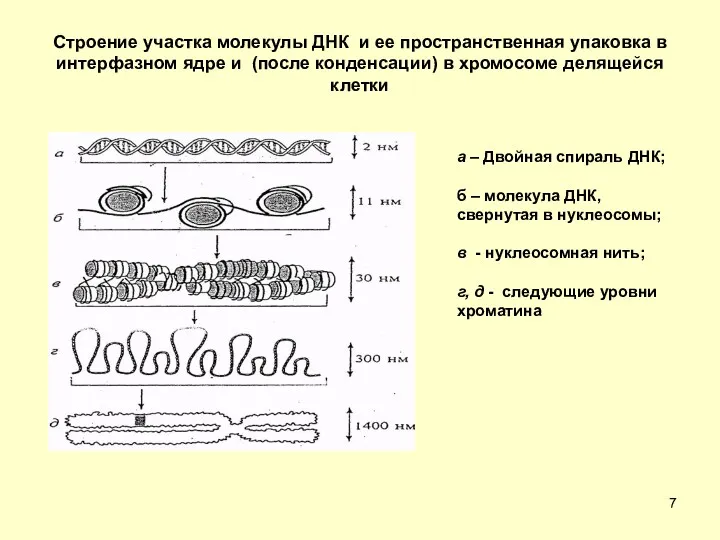
Строение участка молекулы ДНК и ее пространственная упаковка в интерфазном ядре
Строение участка молекулы ДНК и ее пространственная упаковка в интерфазном ядре
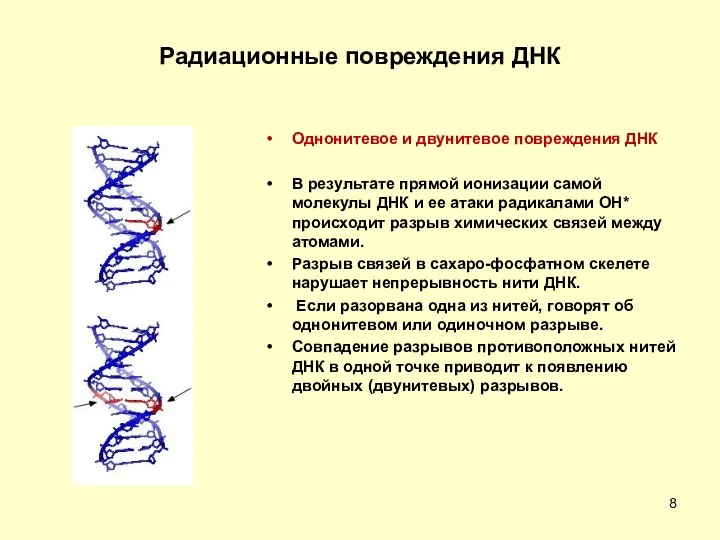
Радиационные повреждения ДНК
Однонитевое и двунитевое повреждения ДНК
В результате прямой ионизации самой
Радиационные повреждения ДНК
Однонитевое и двунитевое повреждения ДНК
В результате прямой ионизации самой

Радиационные повреждения ДНК
Известно, что одиночные разрывы постоянно возникают в клетке и
Радиационные повреждения ДНК
Известно, что одиночные разрывы постоянно возникают в клетке и
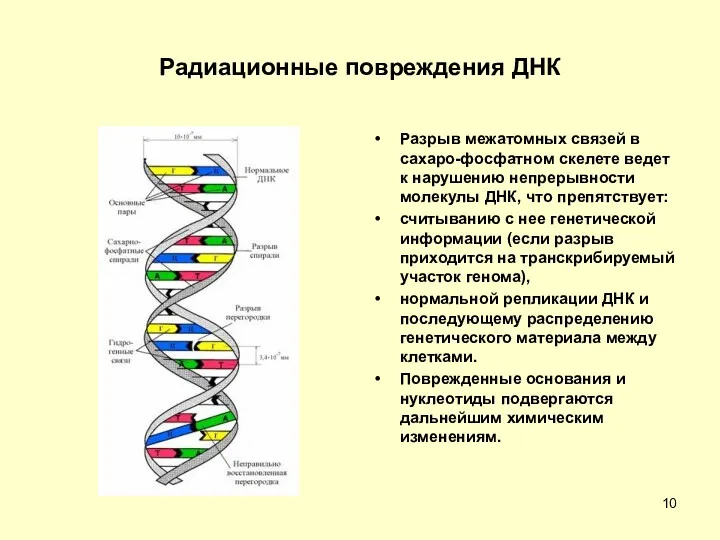
Радиационные повреждения ДНК
Разрыв межатомных связей в сахаро-фосфатном скелете ведет к нарушению
Радиационные повреждения ДНК
Разрыв межатомных связей в сахаро-фосфатном скелете ведет к нарушению

Механизм индукции и реализации смерти клетки (апоптоза)
Еще одним следствием повреждения молекул
Механизм индукции и реализации смерти клетки (апоптоза)
Еще одним следствием повреждения молекул

Механизм индукции и реализации смерти клетки (апоптоза)
Именно путем апоптоза происходит удаление
Механизм индукции и реализации смерти клетки (апоптоза)
Именно путем апоптоза происходит удаление

Механизм индукции и реализации смерти клетки (апоптоза)
Очень четко развитие апоптоза проявляется
Механизм индукции и реализации смерти клетки (апоптоза)
Очень четко развитие апоптоза проявляется
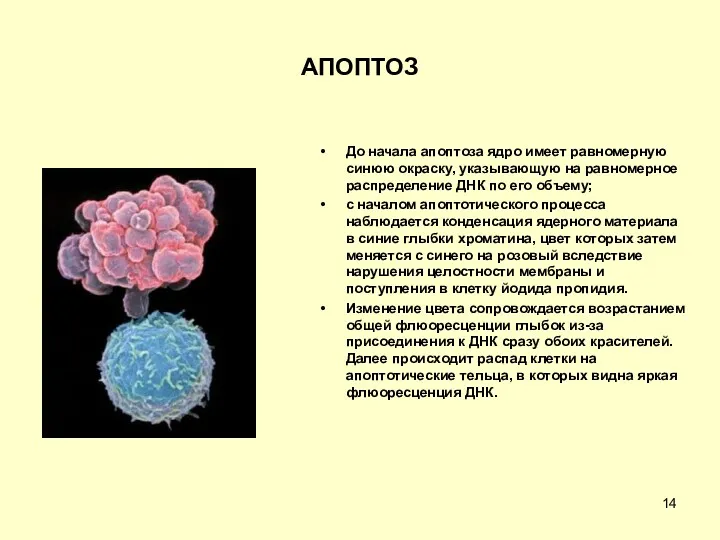
АПОПТОЗ
До начала апоптоза ядро имеет равномерную синюю окраску, указывающую на равномерное
АПОПТОЗ
До начала апоптоза ядро имеет равномерную синюю окраску, указывающую на равномерное
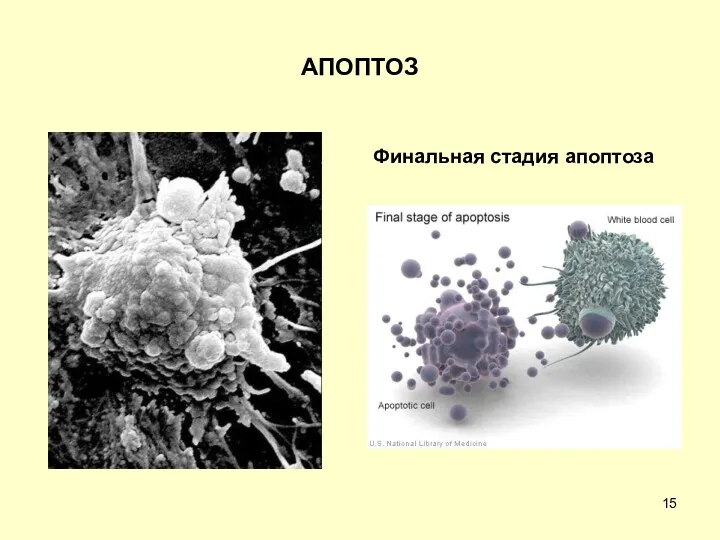
АПОПТОЗ
Финальная стадия апоптоза
АПОПТОЗ
Финальная стадия апоптоза

Механизм индукции и реализации смерти клетки (апоптоза)
Клетки могут значительно различаться по
Механизм индукции и реализации смерти клетки (апоптоза)
Клетки могут значительно различаться по
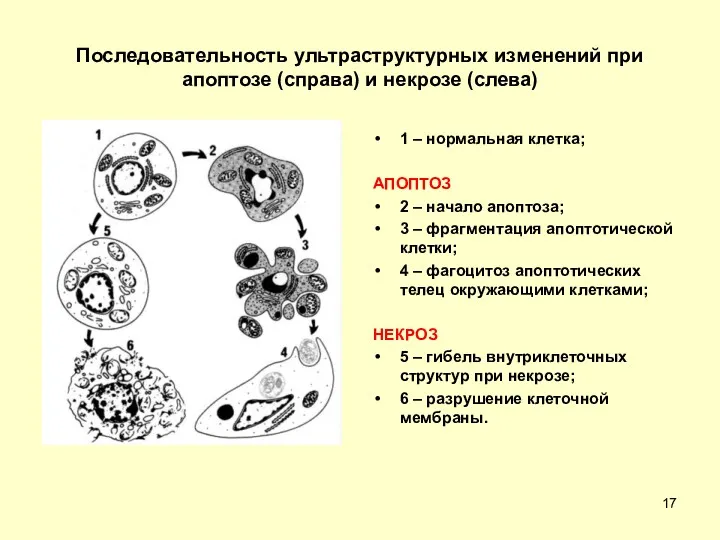
Последовательность ультраструктурных изменений при апоптозе (справа) и некрозе (слева)
1 – нормальная
Последовательность ультраструктурных изменений при апоптозе (справа) и некрозе (слева)
1 – нормальная

Морфологические проявления апоптоза
Для клетки, подвергающейся апоптозу характерно:
Сжатие клетки. Клетка уменьшается
Морфологические проявления апоптоза
Для клетки, подвергающейся апоптозу характерно:
Сжатие клетки. Клетка уменьшается

Морфологические проявления апоптоза
Фагоцитоз апоптотических клеток или телец осуществляется окружающими здоровыми клетками,
Морфологические проявления апоптоза
Фагоцитоз апоптотических клеток или телец осуществляется окружающими здоровыми клетками,

Хромосомные аберрации
Хромосомные аберрации (перестройки) являются классическим проявлением лучевого поражения клеток.
Появление аберраций
Хромосомные аберрации
Хромосомные аберрации (перестройки) являются классическим проявлением лучевого поражения клеток.
Появление аберраций
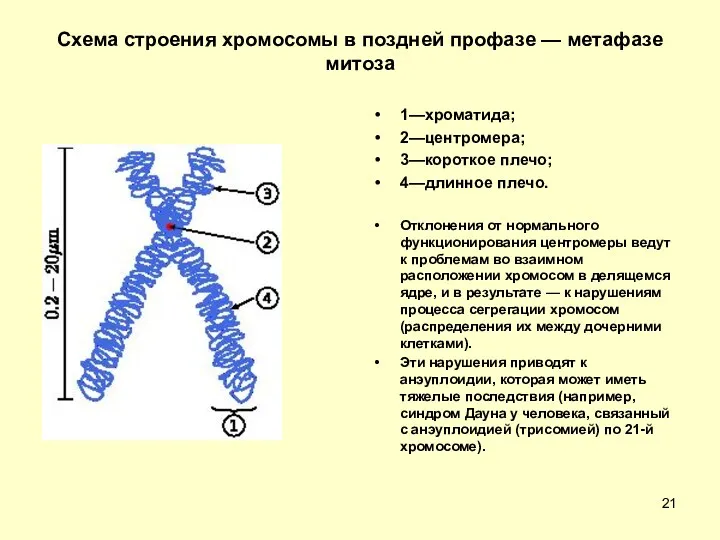
Схема строения хромосомы в поздней профазе — метафазе митоза
1—хроматида;
2—центромера;
Схема строения хромосомы в поздней профазе — метафазе митоза
1—хроматида;
2—центромера;

Хромосомные аберрации
Неверное воссоединение разрывов, когда при репарации происходит соединение участков ДНК
Хромосомные аберрации
Неверное воссоединение разрывов, когда при репарации происходит соединение участков ДНК
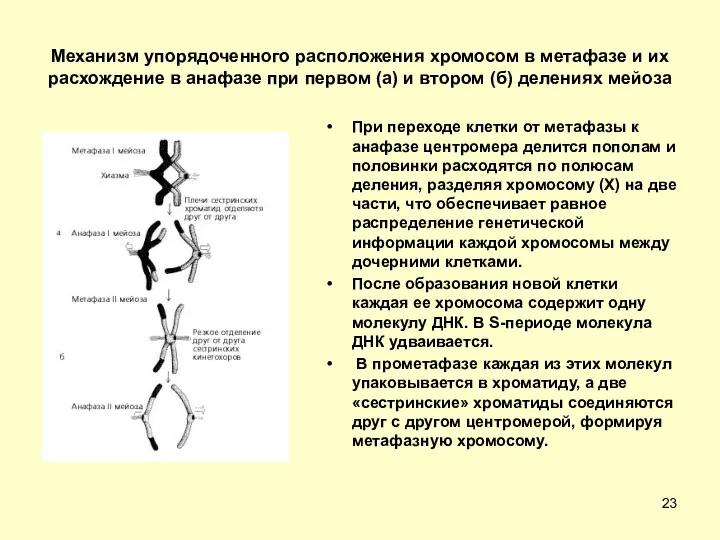
Механизм упорядоченного расположения хромосом в метафазе и их расхождение в анафазе
Механизм упорядоченного расположения хромосом в метафазе и их расхождение в анафазе

Хромосомные аберрации
Возникающие в клетке аберрации подразделяют на хромосомные и хроматидные.
Хромосомные
Хромосомные аберрации
Возникающие в клетке аберрации подразделяют на хромосомные и хроматидные.
Хромосомные

Хромосомные аберрации
Разрыв обеих хроматид может проявиться различным образом — образованием двух
Хромосомные аберрации
Разрыв обеих хроматид может проявиться различным образом — образованием двух
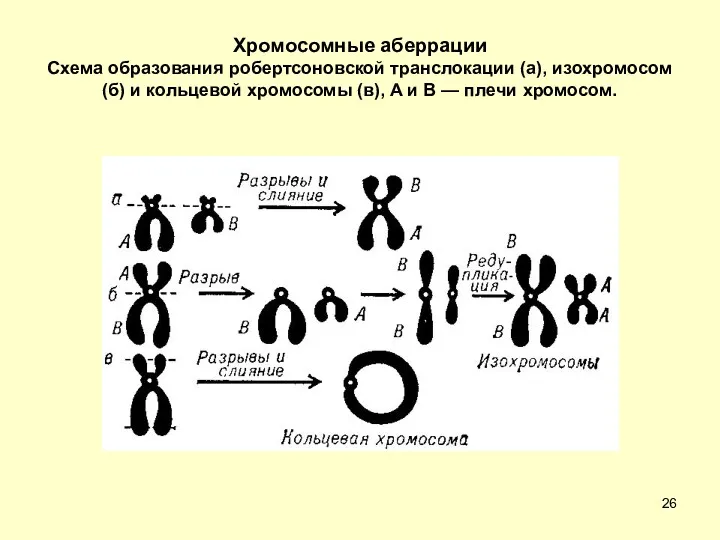
Хромосомные аберрации
Схема образования робертсоновской транслокации (а), изохромосом (б) и кольцевой
Хромосомные аберрации Схема образования робертсоновской транслокации (а), изохромосом (б) и кольцевой
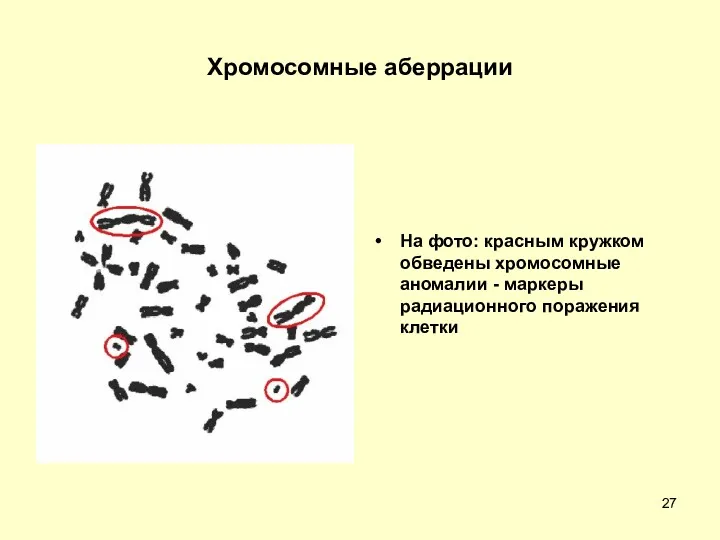
Хромосомные аберрации
На фото: красным кружком обведены хромосомные аномалии - маркеры радиационного
Хромосомные аберрации
На фото: красным кружком обведены хромосомные аномалии - маркеры радиационного
 Віруси. Будова і життєві цикли. Небезпечні вірусні хвороби людини. Урок для 10 класу
Віруси. Будова і життєві цикли. Небезпечні вірусні хвороби людини. Урок для 10 класу Бөлме гүлдері - адамның досы
Бөлме гүлдері - адамның досы Рычаги птицы
Рычаги птицы Приёмы работы с текстом
Приёмы работы с текстом Память и внимание
Память и внимание Ямчатость стебля – обзор, физиология
Ямчатость стебля – обзор, физиология Органы чувств человека
Органы чувств человека Химический состав клеток. Углеводы. Липиды
Химический состав клеток. Углеводы. Липиды Обыкновенный песец или полярная лисица
Обыкновенный песец или полярная лисица Середовища існування організмів
Середовища існування організмів Покровная система
Покровная система Структура и функции биологических мембран
Структура и функции биологических мембран Воздушные и тепловые свойства почв
Воздушные и тепловые свойства почв Психрофитті және криофитті аймақ өсімдіктері
Психрофитті және криофитті аймақ өсімдіктері Урок по теме Многообразие рыб
Урок по теме Многообразие рыб Иглокожие. Лекция 15
Иглокожие. Лекция 15 Презентация Основные методы селекции
Презентация Основные методы селекции Экспрессия генов
Экспрессия генов Насекомые. Отгадай загадки
Насекомые. Отгадай загадки Обмен углеводов-1
Обмен углеводов-1 Микробиология, как наука. Морфология и классификация бактерий
Микробиология, как наука. Морфология и классификация бактерий Анатомия наружного уха
Анатомия наружного уха Tissues of the human body
Tissues of the human body Растения Красной книги
Растения Красной книги Систематика млекопитающих
Систематика млекопитающих Современная биологическая картина мира
Современная биологическая картина мира Регуляция синтеза белка у прокариот и эукариот
Регуляция синтеза белка у прокариот и эукариот Клуб садоводов и огородников Здоровый огород
Клуб садоводов и огородников Здоровый огород