- Морфология прокариот

Содержание
- 2. Размеры прокариот Средний размер клеток прокариот – 0,5 – 3 мкм. У Escherichia coli – 0,4—0,8
- 3. Гигантские клетки прокариот Epulopiscium fishelsoni – гигантская бактерия размером — 80 × 600 мкм (до 700
- 4. «Карликовые» прокариоты (нанобактерии) имеют размер Микоплазмы - 0,1 – 0,15 мкм Хламидии - 0,5 мкм Риккетсии
- 5. Форма клеток прокариот Три основные группы прокариот по морфологии: Кокки – клетки округлой формы (от лат.
- 6. Кокки Кокки могут быть объединены в различные агрегаты: Одиночные кокки – р. Micrococcus. Micrococcus luteus
- 7. Диплококки – попарно соединенные кокки – р. Neisseria N. gonorrhoeae N. meningitidis (возбудитель гонореи) (возбудитель менингита)
- 8. Стрептококки – цепочки кокков (р. Streptococcus). S. pyogenes (возбудитель тонзилита) S. cremoris (сливочный стрептококк)
- 9. Тетракокки - тетрады клеток. Gaffkya tetragena
- 10. Стафилококки – кокки, соединенные в неправильные скопления в виде «виноградной грозди». Staphylococcus aureus
- 11. Сарцины – кокки, соединенные в «пакеты» по 8, 16, 32, 64 клетки (р. Sarcina).
- 12. Палочковидные бактерии Палочки – короткие и длинные, толстые и тонкие, одиночные, соединенные по 2 и в
- 13. Неспорообразующие палочки Escherichia coli Pseudomonas aeruginosa
- 14. Спорообразующие палочки Bacillus anthracis Bacillus thuringiensis возбудитель сибирской язвы Кристаллы δ-эндотоксина Bacillus thuringiensis
- 15. Типы спорообразования Бациллярный Клостридиальный Плектридиальный р. Clostridium (анаэробные спорообразующие палочки)) р. Bacillus (аэробные спорообразующие палочки)
- 16. Расположение спор в клетке Различное расположение эндоспор в материнской клетке: 1, 4 — центральное; 2, 3,
- 17. Извитые формы Вибрионы (от лат. «vibrio» - изгибаюсь) – изогнутые палочки (форма «запятой») (р. Vibrio). Vibrio
- 18. Спириллы (от лат. «spira» - завиток) – клетки, имеющие 2-3 изгиба. Р. Spirillum.
- 19. Спирохеты – тонкие длинные клетки со множеством завитков и изгибов. Treponema pallidum Borrelia burgdorferi
- 20. Бактерии, образующие простеки (выросты клетки) и стебельки (слизистые придатки клетки) 1 – Caulobacter; 2 – Hyphomicrobium;
- 21. Нитчатые бактерии – клетки собраны в длинные нити, погруженные в общий слизистый чехол. 1 - Beggiatoa;
- 22. Ветвящиеся бактерии Актиномицеты (р. Streptomyces) 1 – клетка 2 - мицелий
- 23. Другие формы клеток A – клетки треугольной формы, B – плоские прямоугольные клетки, C – лентовидные
- 24. Типы жгутикования Монополярный монотрихальный (р. Vibrio) Монополярный политрихальный (р. Pseudomonas) Биполярный политрихальный (р. Spirillum) Перитрихальный р.
- 25. СТРОЕНИЕ БАКТЕРИАЛЬНОЙ КЛЕТКИ Вопросы: 1.Внутриклеточные структуры. 2. Клеточная оболочка. 3. Поверхностные структуры клетки. 4. Типы движения
- 26. Клетка прокариот представлена одной полостью, образуемой ЦПМ. Нет вторичных полостей. Отсутствуют органеллы, типичные для эукариот. Функционально
- 27. 1 – гранулы поли-β-оксимасляной кислоты; 2 – жировые капельки; 3 – включения серы; 4 – трубчатые
- 28. Прокариотная клетка состоит из трех компартментов: 1. Поверхностные структуры - капсула, S-слой, жгутики, пили, адгезины. 2.
- 29. 1. Внутриклеточные структуры Цитоплазма, рибосомы, генетический аппарат, включения, внутрицитоплазматические мембраны (ВЦМ).
- 30. Цитоплазма Цитоплазма – содержимое клетки, окруженное ЦПМ. Коллоидная фаза цитоплазмы (цитозоль) – растворимая фаза. Содержит белки,
- 31. Рибосомы 70-S типа, размер - 15-20 нм. Состоят из двух субъединиц: 30 S и 50 S.
- 33. Генетический аппарат Геном у прокариот представлен бактериальной хромосомой (БХ) и внехромосомными ДНК. Бактериальная клетка гаплоидна, имеет
- 35. М. м. БХ – в среднем 1010 Да (5×106 пар оснований) (1Да = 1,66033×10-27 кг). Составляет
- 36. Внехромосомные молекулы ДНК: Плазмиды – кольцевые, реже, линейные фрагменты ДНК с м. м. 106-108 Да, располагаются
- 37. Внутрицитоплазматические включения 1) Активно функционирующие структуры: Хлоросомы (у зеленых бактерий) – продолговатые пузырьки, окруженные липидной мембраной,
- 38. Фикобилисомы (у цианобактерий) - гранулы размером 28-55 нм, располагаются на фотосинтетических мембранах, содержат фикобилипротеины.
- 39. Карбоксисомы - у фототрофных и хемолитотрофных бактерий, фиксирующих СО2. Форма - многогранник. Состоят из рибулозо-1,5-бифосфат-карбоксилазы.
- 40. Аэросомы (газовые везикулы) – у водных бактерий – состоят из газовых пузырьков, которые окружены белковой мембраной;
- 41. Магнетосомы (у магнитных бактерий) состоят из магнетита Fe2O3, определяют магнитотаксис.
- 42. 2) Запасные вещества используются как источник углерода, азота, фосфора и энергии Липиды – в виде гранул,
- 43. Сера (у серных бактерий) - в виде глобул. Белки (цианофицин) – у цианобактерий. Белковые включения Bacillus
- 44. Bacillus thuringiensis используется для борьбы с вредными насекомыми
- 45. Внутрицитоплазматические мембраны (ВЦМ) ВЦМ образуются в результате локальной инвагинации ЦПМ в цитоплазму и ее разрастания, сохраняют
- 46. а); в); с) – тилакоиды цианобактерий; d) – тубулярные ВЦМ у Thiocapsa sp. (пурпурная бактерия); е)
- 47. 2. Клеточная оболочка ЦПМ Химический состав: белки – 50 - 75 %, липиды – 15 -
- 48. Модель строения элементарной биологической мембраны: 1 — молекулы липидов: а — гидрофильная "голова"; б — гидрофобный
- 49. Мембраны архей Схема бислойной и монослойной мембран архей, образованных ди- (А) и тетраэфирами (Б) глицерина: 1
- 50. Функции ЦПМ 1. Барьерная. 2. Транспортная. 3. Осмотическая. 4. Метаболическая (участвует в энергетических процессах, в ЦПМ
- 51. Клеточная стенка (КС) КС – обязательный структурный элемент, придает клеткам форму. КС отсутствует у микоплазм и
- 52. Пептидогликан (муреин) – специфический гетерополимер КС бактерий, отсутствует у эукариот. Построен из чередующихся остатков N-ацетилглюкозамина и
- 53. Пептидогликан – линейный полимер, его молекулы образуют сеть из параллельно расположенных полисахаридных цепей, соединенных пептидными хвостами.
- 54. Прокариоты в зависимости от структуры и химического состава клеточной стенки делят на две группы: грам(+) и
- 55. Строение клеточной стенки грам(+) и грам(-) бактерий Схематическое строение клеточной стенки грам(+) (А) и грам(-) (Б)
- 56. Химический состав Структура пептидогликан (п/гл) – 40-90 %, тейхоевые кислоты (полимеры из 8-50 остатков глицерина или
- 57. ГРАМ(+) БАКТЕРИИ: р. Bacillus – B. anthracis (возбудитель сибирской язвы), B. thuringiensis (поражает насекомых), B. mesentericus
- 58. КЛЕТОЧНАЯ СТЕНКА ГРАМ(-) БАКТЕРИЙ Химический состав Структура пептидогликан (п/гл) – 1 - 10 %, липополисахариды, фосфолипиды,
- 59. ГРАМ(-) БАКТЕРИИ: Escherichia coli Pseudomonas aeruginosa – синегнойная палочка Neisseria gonorrhoeae - возбудитель гонореи Neisseria meningitidis
- 60. У архей КС трех типов: Состоящие из псевдомуреина - грам (+). Состоящие из гетерополисахарида - грам
- 61. Функции клеточной стенки прокариот: 1. Придает клеткам определенную форму. 2. Механическая. 3. Осмотическая. 3. Транспортная. 4.
- 62. 3. Поверхностные структуры клетки Капсула - слизистое образование, располагается поверх клеточной стенки, сохраняет связь с клеткой:
- 63. Капсулы бактерий
- 64. Слизистые слои – имеют аморфное строение, легко отделяются от клетки. У роящихся клеток (Myxococcus), слизь способствует
- 65. Жгутики Имеются у подвижных бактерий. Жгутик состоит из 3 компонентов: Длинная спиральная нить – фибрилла из
- 66. У грам(+) – М- и S-кольцо: М-кольцо в ЦПМ, S- - в пептидогликановом слое клеточной стенки.
- 67. Движущей силой вращения колец служит протонный градиент. При вращении колец их движение сообщается связанной с ним
- 68. Микроворсинки (фимбрии, пили) Это нитевидные клеточные придатки диаметром 3-10 нм, длиной - 0,2-2 мкм; состоят из
- 69. Пили
- 70. 4. Типы движения прокариот и таксисы Типы движения: 1. Плавание – передвижение бактерий при помощи жгутиков
- 71. 4. Подтягивающее движение - движение по твердой поверхности с участием пили особого типа, располагающихся на полюсах
- 72. Таксисы 1. Хемотаксис – движение бактерий в определенном направлении относительно источника химического соединения - эффектора: 1)
- 73. 5. Магнитотаксис – у магнитобактерий, содержат магнетосомы, движение определяется силовыми линиями магнитного поля Земли. 6. Вискозитаксис
- 74. МОРФОЛОГИЧЕСКИ ДИФФЕРЕНЦИРОВАННЫЕ КЛЕТКИ ПРОКАРИОТ (МДК)
- 75. Вопросы: Цисты и акинеты. Эндоспоры бактерий. Экзоспоры бактерий и актиномицетов.
- 76. Вегетативные клетки многих прокариот при определенных условиях дают начало структурам, которые морфологически отличаются от исходных. Их
- 77. К МДК прокариот относятся: Покоящиеся формы – эндоспоры грам(+) бактерий, цисты, акинеты цианобактерий, экзоспоры некоторых бактерий,
- 78. 1. Цисты и акинеты Цисты образуются у миксобактерий, азотобактера, риккетсий и др. У Azotobacter цисты округлой
- 79. Акинеты цианобактерий Акинеты крупнее вегетативных клеток, продолговатой или сферической формы, с толстой клеточной оболочкой. Содержат меньше
- 80. Акинета Видны гранулы цианофицина (белок), зерна волютина (полифостфаты) – запасные питательные вещества.
- 81. 2. Эндоспоры бактерий Бактериальные эндоспоры – особый тип покоящихся клеток в основном Грам(+) бактерий. Формируются эндогенно,
- 82. Строение бактериальной эндоспоры 1 — нуклеоид; 2 — цитоплазма; 3 — внутренняя мембрана споры; 4 —
- 83. Химический состав эндоспор Белки эндоспор богаты цистеином и гидрофобными аминокислотами. Содержание ДНК ниже, чем в исходной
- 84. Содержание РНК в спорах ниже, чем в вегетативных клетках. В спорах повышенное содержание ионов Са2+, Mg2+,
- 85. Кортекс построен из молекул особого типа пептидогликана. При прорастании споры из него формируется клеточная стенка вегетативной
- 86. Устойчивость эндоспор Устойчивы к высоким (+80 °С) и низким температурам, обезвоживанию, высокой кислотности среды, радиации, к
- 87. Факторы, обеспечивающие устойчивость эндоспор: 1. Нахождение споровой цитоплазмы в обезвоженном состоянии. 2. Термостойкость споровых ферментов. 3.
- 88. 3. Экзоспоры бактерий и актиномицетов Экзоспоры у бактерий образуются редко, например, у некоторых почкующихся бактерий. Образуются
- 89. Отпочковывание экзоспор от одного из полюсов клеток Methylosinus (почкующаяся метилотрофная бактерия) Экзоспора Клетка
- 90. Экзоспоры имеют более плотную и утолщенную клеточную стенку. В экзоспорах нет дипиколиновой кислоты и характерных для
- 91. Экзоспоры актиномицетов Актиномицеты – это актинобактерии, имеют ветвящиеся клетки, образуют мицелий.
- 92. Споры актиномицетов Экзоспоры у актиномицетов образуются путем фрагментации гифы. Имеют плотную и утолщенную клеточную стенку. Устойчивы
- 94. Скачать презентацию

Размеры прокариот
Средний размер клеток прокариот – 0,5 – 3 мкм.
У
Размеры прокариот
Средний размер клеток прокариот – 0,5 – 3 мкм.
У
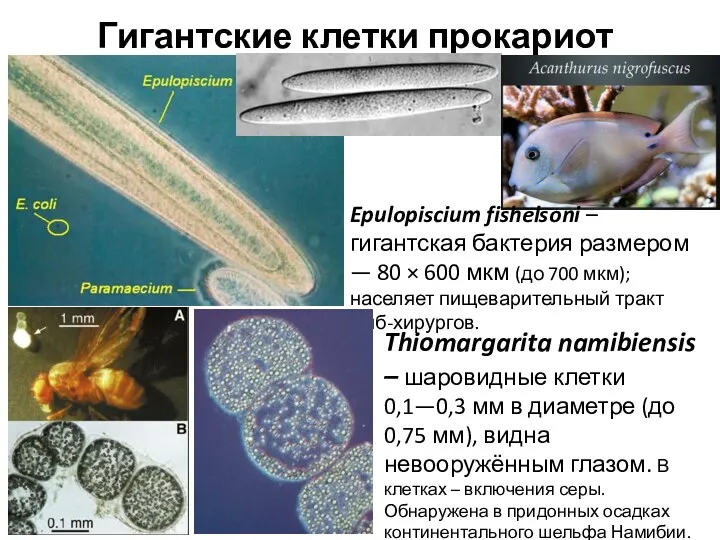
Гигантские клетки прокариот
Epulopiscium fishelsoni – гигантская бактерия размером — 80 ×
Гигантские клетки прокариот
Epulopiscium fishelsoni – гигантская бактерия размером — 80 ×
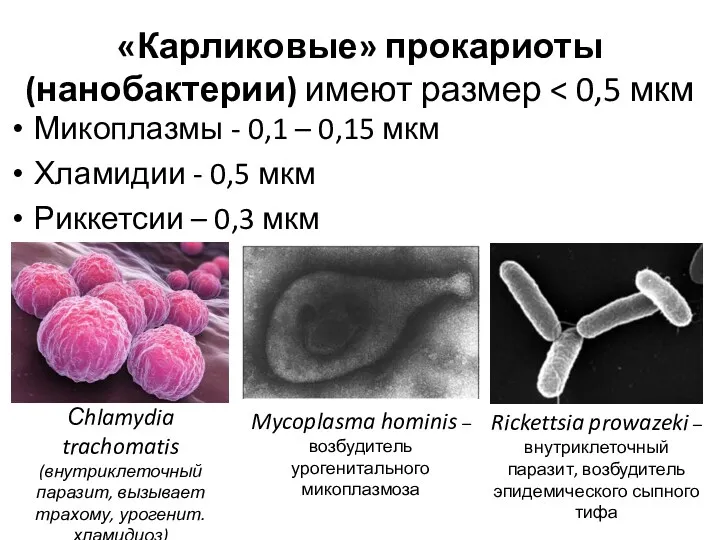
«Карликовые» прокариоты (нанобактерии) имеют размер < 0,5 мкм
Микоплазмы - 0,1 –
«Карликовые» прокариоты (нанобактерии) имеют размер < 0,5 мкм
Микоплазмы - 0,1 –

Форма клеток прокариот
Три основные группы прокариот по морфологии:
Кокки – клетки округлой
Форма клеток прокариот
Три основные группы прокариот по морфологии:
Кокки – клетки округлой
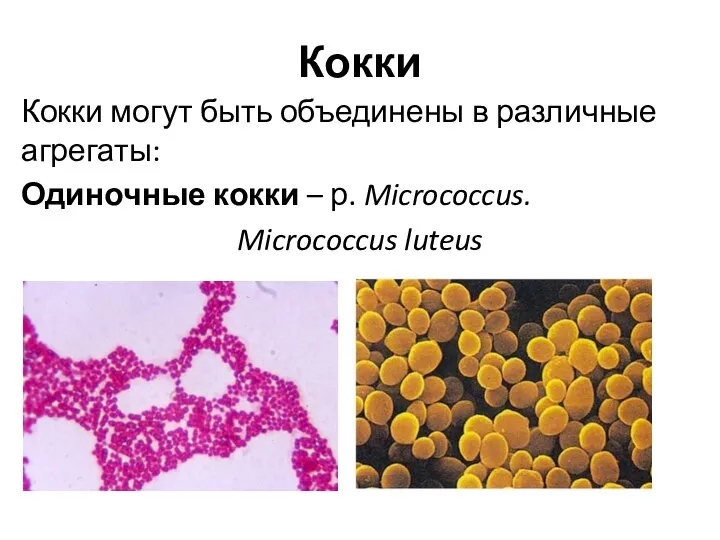
Кокки
Кокки могут быть объединены в различные агрегаты:
Одиночные кокки – р. Micrococcus.
Micrococcus
Кокки
Кокки могут быть объединены в различные агрегаты:
Одиночные кокки – р. Micrococcus.
Micrococcus
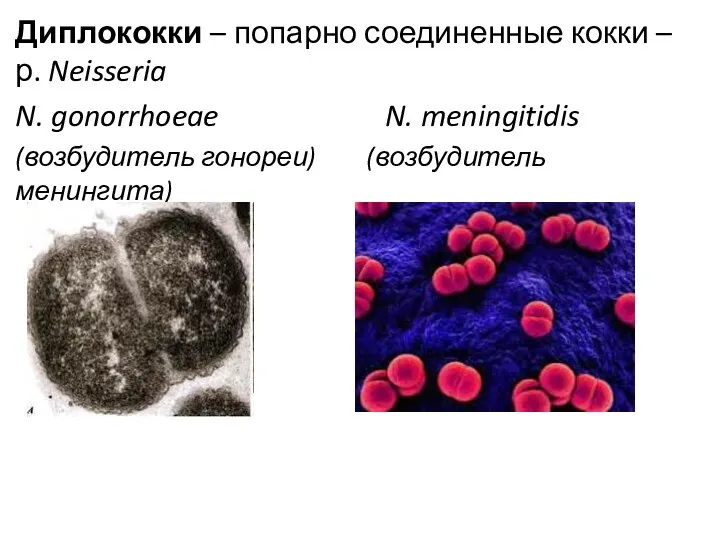
Диплококки – попарно соединенные кокки – р. Neisseria
N. gonorrhoeae N.
Диплококки – попарно соединенные кокки – р. Neisseria
N. gonorrhoeae N.
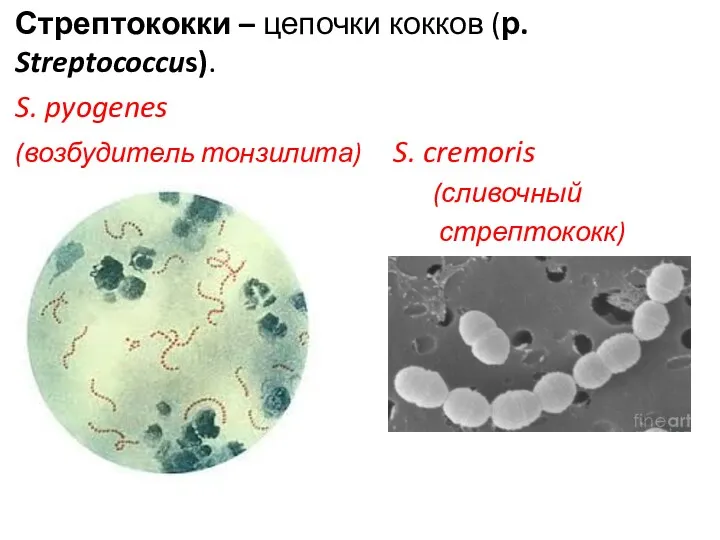
Стрептококки – цепочки кокков (р. Streptococcus).
S. pyogenes
(возбудитель тонзилита) S. cremoris
Стрептококки – цепочки кокков (р. Streptococcus).
S. pyogenes
(возбудитель тонзилита) S. cremoris
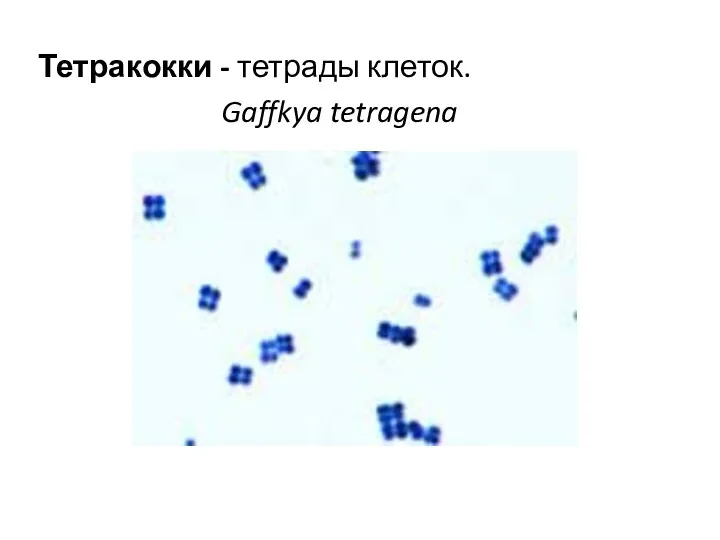
Тетракокки - тетрады клеток.
Gaffkya tetragena
Тетракокки - тетрады клеток.
Gaffkya tetragena
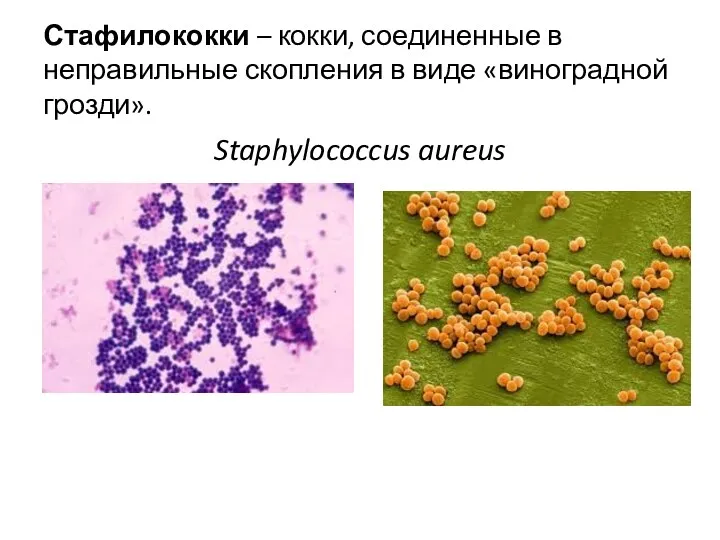
Стафилококки – кокки, соединенные в неправильные скопления в виде «виноградной грозди».
Staphylococcus
Стафилококки – кокки, соединенные в неправильные скопления в виде «виноградной грозди».
Staphylococcus
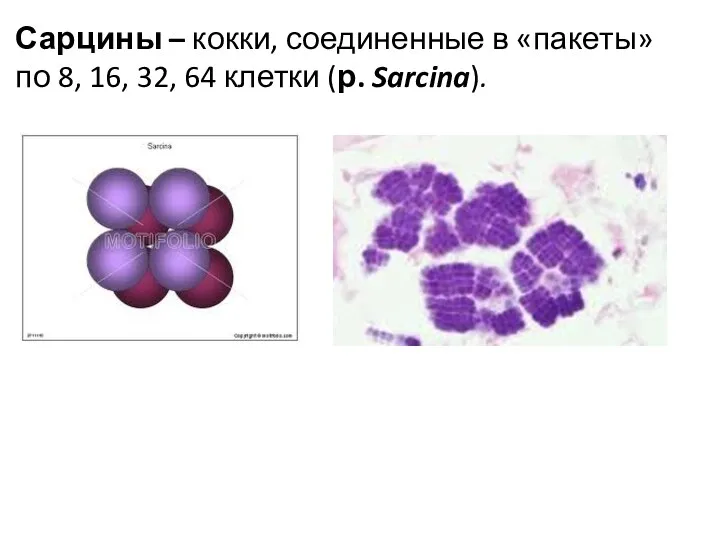
Сарцины – кокки, соединенные в «пакеты» по 8, 16, 32, 64
Сарцины – кокки, соединенные в «пакеты» по 8, 16, 32, 64
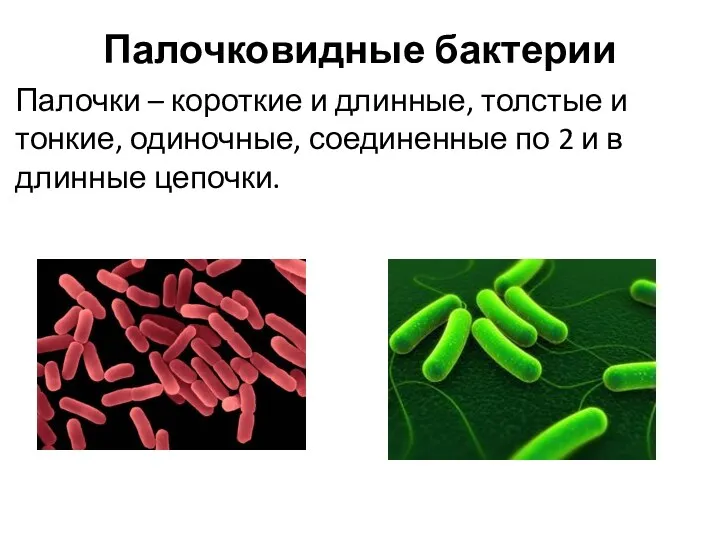
Палочковидные бактерии
Палочки – короткие и длинные, толстые и тонкие, одиночные, соединенные
Палочковидные бактерии
Палочки – короткие и длинные, толстые и тонкие, одиночные, соединенные
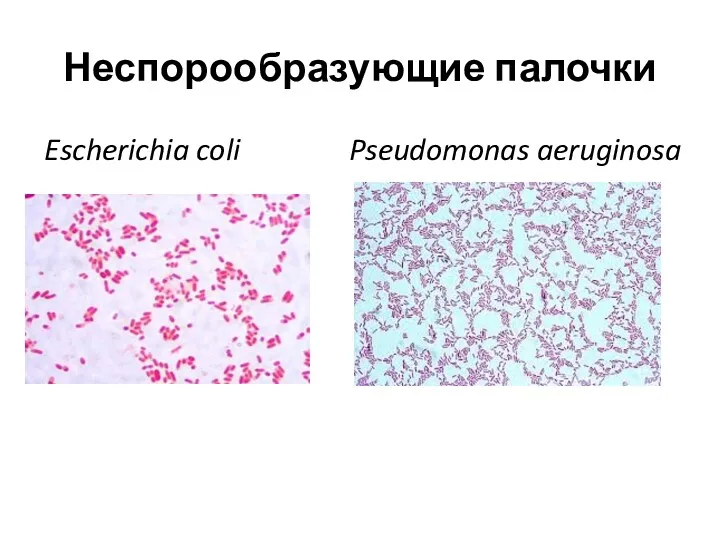
Неспорообразующие палочки
Escherichia coli Pseudomonas aeruginosa
Неспорообразующие палочки
Escherichia coli Pseudomonas aeruginosa
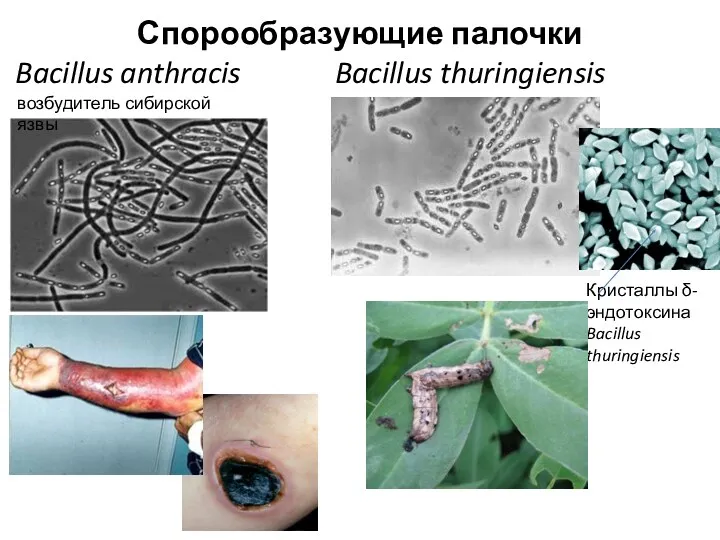
Спорообразующие палочки
Bacillus anthracis Bacillus thuringiensis
возбудитель сибирской язвы
Кристаллы δ-эндотоксина Bacillus thuringiensis
Спорообразующие палочки
Bacillus anthracis Bacillus thuringiensis
возбудитель сибирской язвы
Кристаллы δ-эндотоксина Bacillus thuringiensis
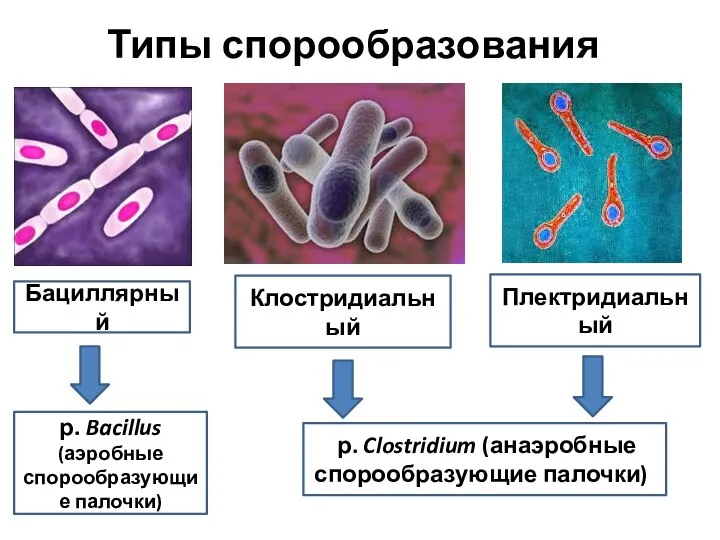
Типы спорообразования
Бациллярный
Клостридиальный
Плектридиальный
р. Clostridium (анаэробные спорообразующие палочки))
р. Bacillus (аэробные спорообразующие
Типы спорообразования
Бациллярный
Клостридиальный
Плектридиальный
р. Clostridium (анаэробные спорообразующие палочки))
р. Bacillus (аэробные спорообразующие
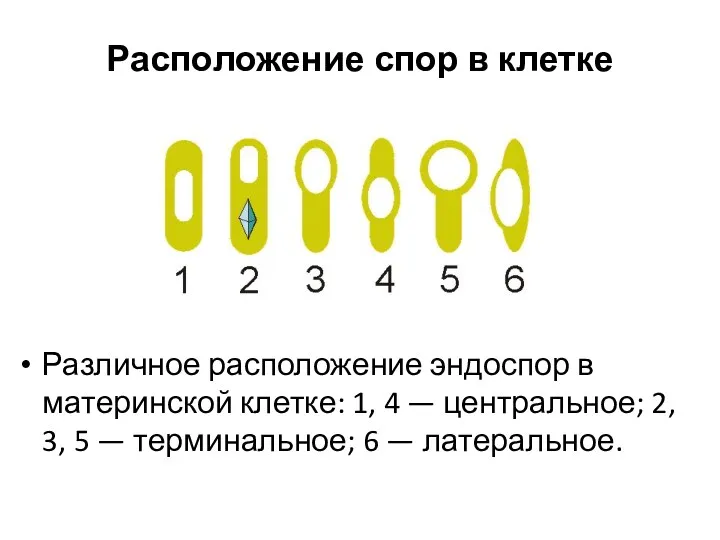
Расположение спор в клетке
Различное расположение эндоспор в материнской клетке: 1, 4
Расположение спор в клетке
Различное расположение эндоспор в материнской клетке: 1, 4
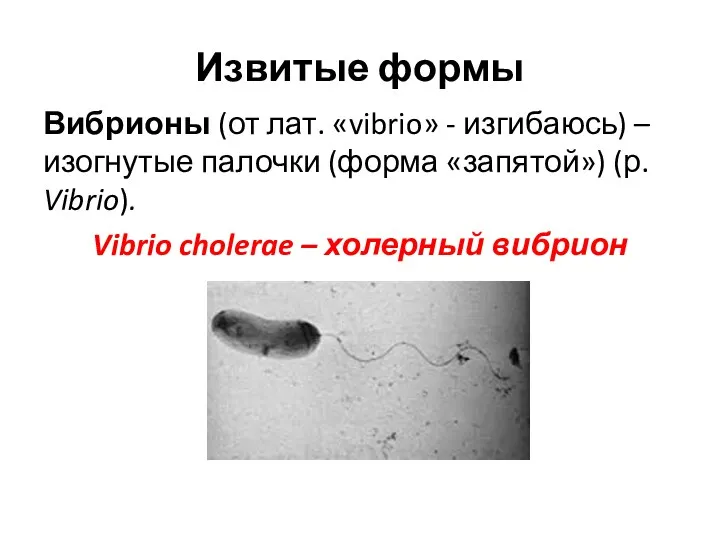
Извитые формы
Вибрионы (от лат. «vibrio» - изгибаюсь) – изогнутые палочки (форма
Извитые формы
Вибрионы (от лат. «vibrio» - изгибаюсь) – изогнутые палочки (форма
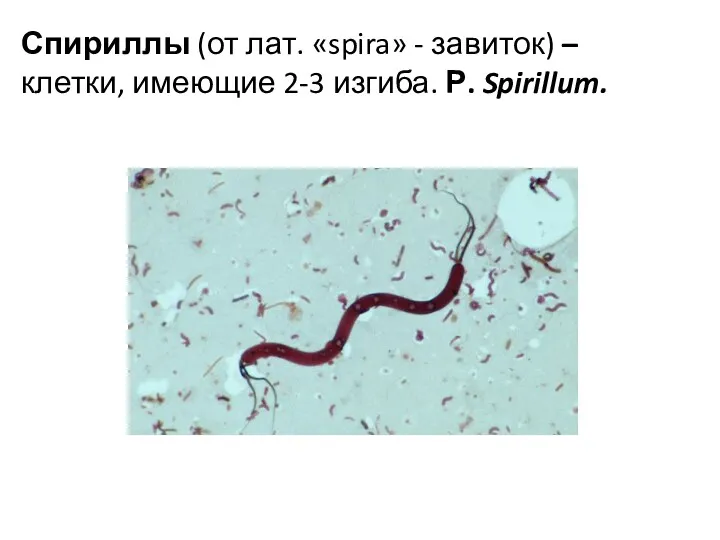
Спириллы (от лат. «spira» - завиток) – клетки, имеющие 2-3 изгиба.
Спириллы (от лат. «spira» - завиток) – клетки, имеющие 2-3 изгиба.
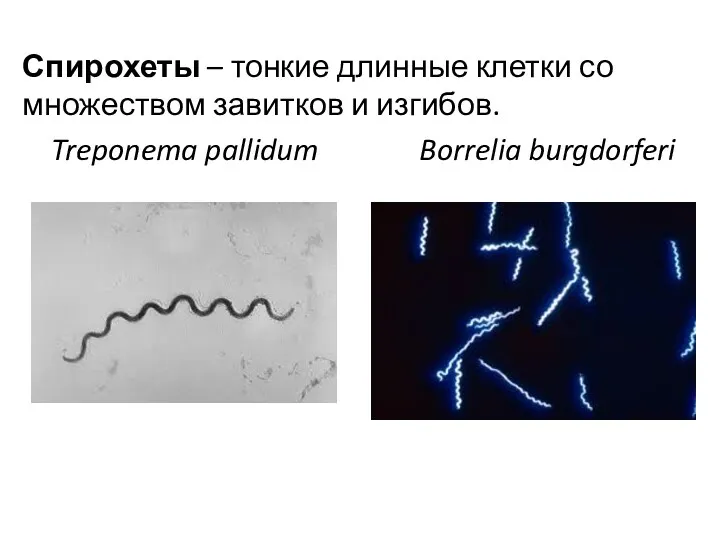
Спирохеты – тонкие длинные клетки со множеством завитков и изгибов.
Спирохеты – тонкие длинные клетки со множеством завитков и изгибов.
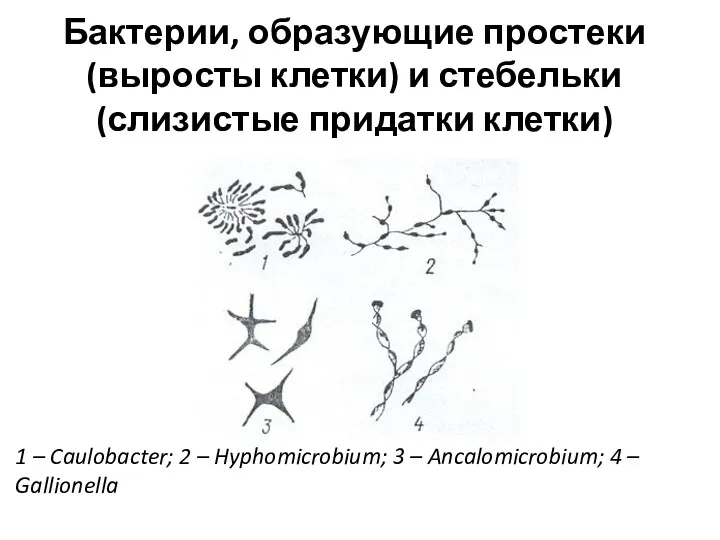
Бактерии, образующие простеки (выросты клетки) и стебельки (слизистые придатки клетки)
1 –
Бактерии, образующие простеки (выросты клетки) и стебельки (слизистые придатки клетки)
1 –

Нитчатые бактерии – клетки собраны в длинные нити, погруженные в общий
Нитчатые бактерии – клетки собраны в длинные нити, погруженные в общий
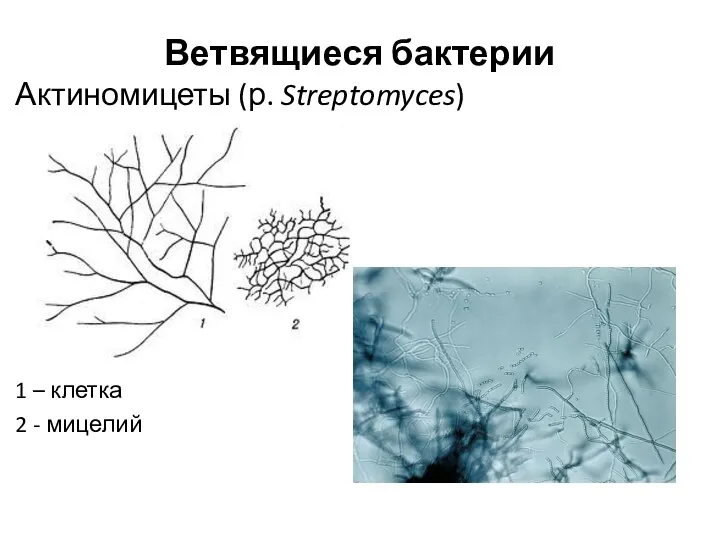
Ветвящиеся бактерии
Актиномицеты (р. Streptomyces)
1 – клетка
2 - мицелий
Ветвящиеся бактерии
Актиномицеты (р. Streptomyces)
1 – клетка
2 - мицелий
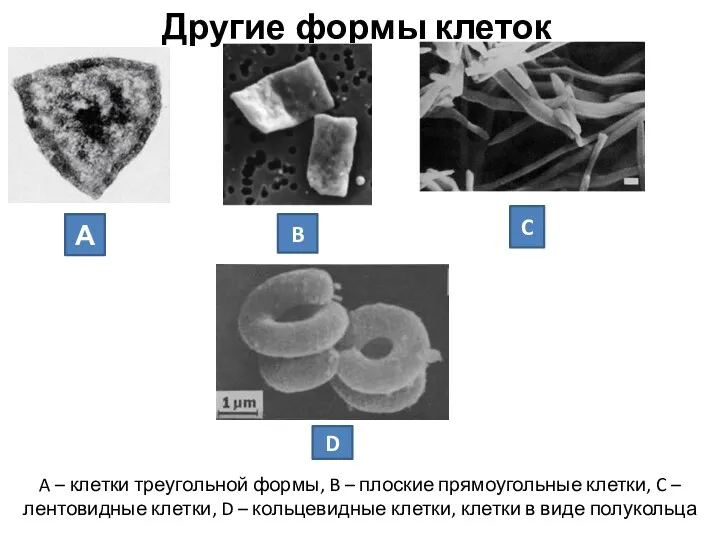
Другие формы клеток
A – клетки треугольной формы, B – плоские прямоугольные
Другие формы клеток
A – клетки треугольной формы, B – плоские прямоугольные
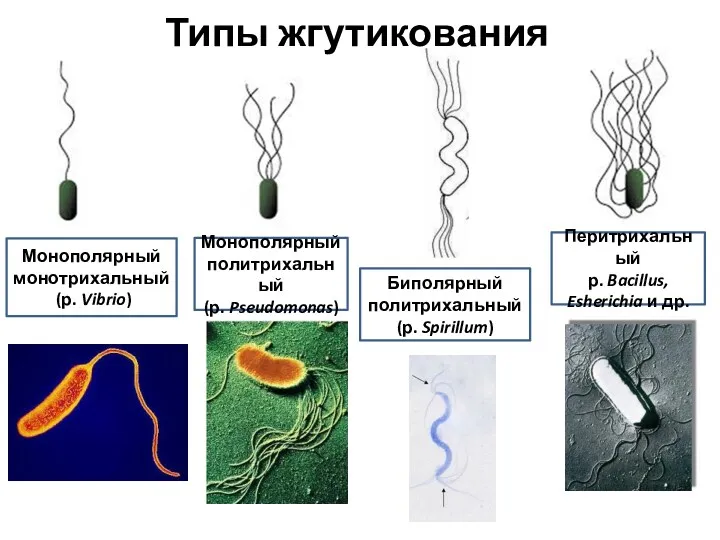
Типы жгутикования
Монополярный
монотрихальный
(р. Vibrio)
Монополярный политрихальный
(р. Pseudomonas)
Биполярный политрихальный
(р. Spirillum)
Перитрихальный
р.
Типы жгутикования
Монополярный
монотрихальный
(р. Vibrio)
Монополярный политрихальный
(р. Pseudomonas)
Биполярный политрихальный
(р. Spirillum)
Перитрихальный
р.

СТРОЕНИЕ БАКТЕРИАЛЬНОЙ КЛЕТКИ
Вопросы:
1.Внутриклеточные структуры.
2. Клеточная оболочка.
3. Поверхностные структуры клетки.
4. Типы
СТРОЕНИЕ БАКТЕРИАЛЬНОЙ КЛЕТКИ
Вопросы:
1.Внутриклеточные структуры.
2. Клеточная оболочка.
3. Поверхностные структуры клетки.
4. Типы

Клетка прокариот представлена одной полостью, образуемой ЦПМ.
Нет вторичных полостей.
Отсутствуют
Клетка прокариот представлена одной полостью, образуемой ЦПМ.
Нет вторичных полостей.
Отсутствуют
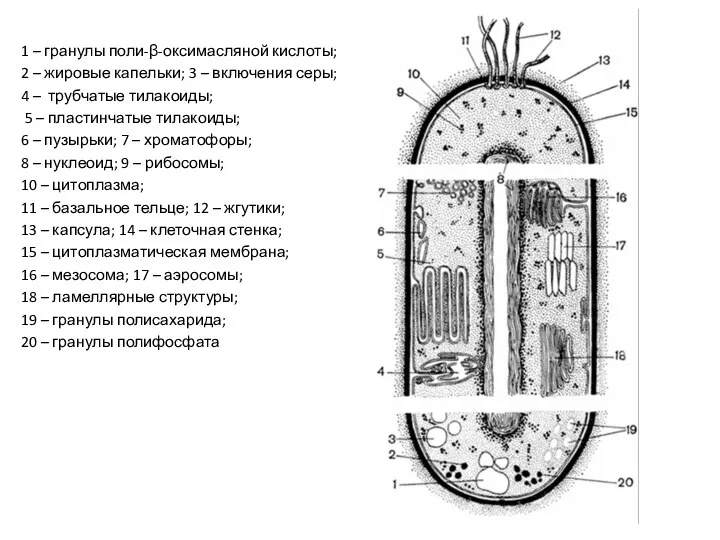
1 – гранулы поли-β-оксимасляной кислоты;
2 – жировые капельки; 3 –
1 – гранулы поли-β-оксимасляной кислоты;
2 – жировые капельки; 3 –

Прокариотная клетка состоит из трех компартментов:
1. Поверхностные структуры - капсула, S-слой,
Прокариотная клетка состоит из трех компартментов:
1. Поверхностные структуры - капсула, S-слой,

1. Внутриклеточные структуры
Цитоплазма,
рибосомы,
генетический аппарат,
включения,
внутрицитоплазматические мембраны (ВЦМ).
1. Внутриклеточные структуры
Цитоплазма,
рибосомы,
генетический аппарат,
включения,
внутрицитоплазматические мембраны (ВЦМ).

Цитоплазма
Цитоплазма – содержимое клетки, окруженное ЦПМ.
Коллоидная фаза цитоплазмы (цитозоль) – растворимая
Цитоплазма
Цитоплазма – содержимое клетки, окруженное ЦПМ.
Коллоидная фаза цитоплазмы (цитозоль) – растворимая

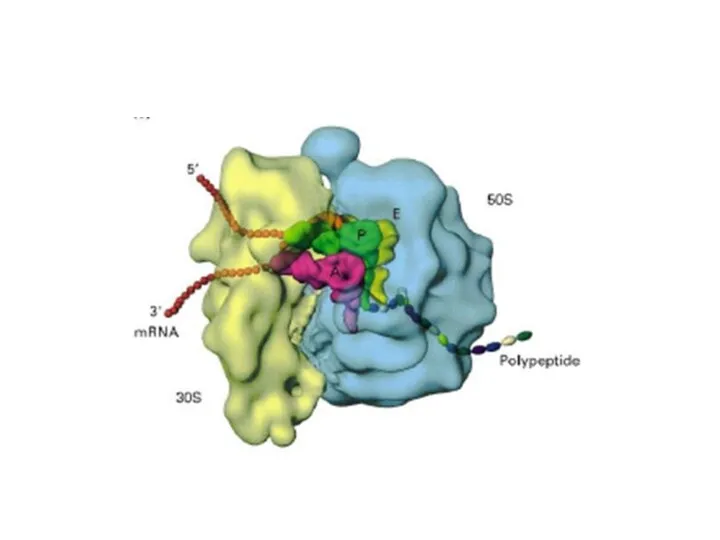
Рибосомы
70-S типа, размер - 15-20 нм.
Состоят из двух субъединиц: 30
Рибосомы
70-S типа, размер - 15-20 нм.
Состоят из двух субъединиц: 30

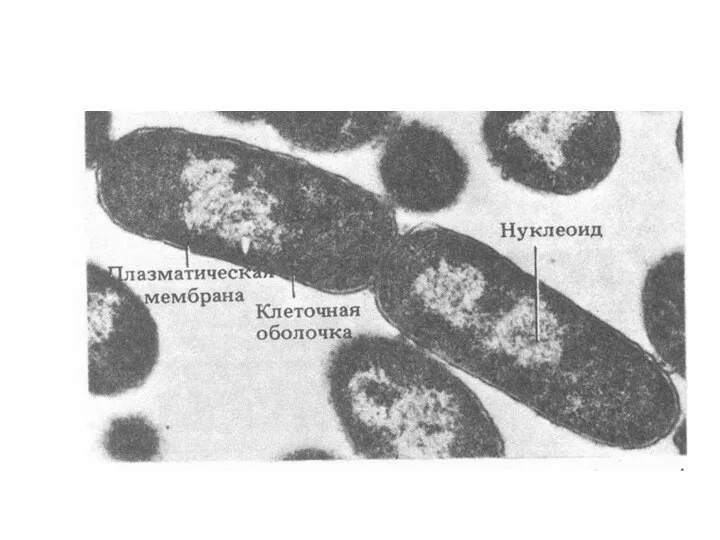
Генетический аппарат
Геном у прокариот представлен бактериальной хромосомой (БХ) и внехромосомными ДНК.
Генетический аппарат
Геном у прокариот представлен бактериальной хромосомой (БХ) и внехромосомными ДНК.

М. м. БХ – в среднем 1010 Да (5×106 пар оснований)
М. м. БХ – в среднем 1010 Да (5×106 пар оснований)

Внехромосомные молекулы ДНК:
Плазмиды – кольцевые, реже, линейные фрагменты ДНК с м.
Внехромосомные молекулы ДНК:
Плазмиды – кольцевые, реже, линейные фрагменты ДНК с м.

Внутрицитоплазматические включения
1) Активно функционирующие структуры:
Хлоросомы (у зеленых бактерий) – продолговатые пузырьки,
Внутрицитоплазматические включения
1) Активно функционирующие структуры:
Хлоросомы (у зеленых бактерий) – продолговатые пузырьки,

Фикобилисомы (у цианобактерий) - гранулы размером 28-55 нм, располагаются на фотосинтетических
Фикобилисомы (у цианобактерий) - гранулы размером 28-55 нм, располагаются на фотосинтетических

Карбоксисомы - у фототрофных и хемолитотрофных бактерий, фиксирующих СО2. Форма -
Карбоксисомы - у фототрофных и хемолитотрофных бактерий, фиксирующих СО2. Форма -

Аэросомы (газовые везикулы) – у водных бактерий – состоят из газовых
Аэросомы (газовые везикулы) – у водных бактерий – состоят из газовых
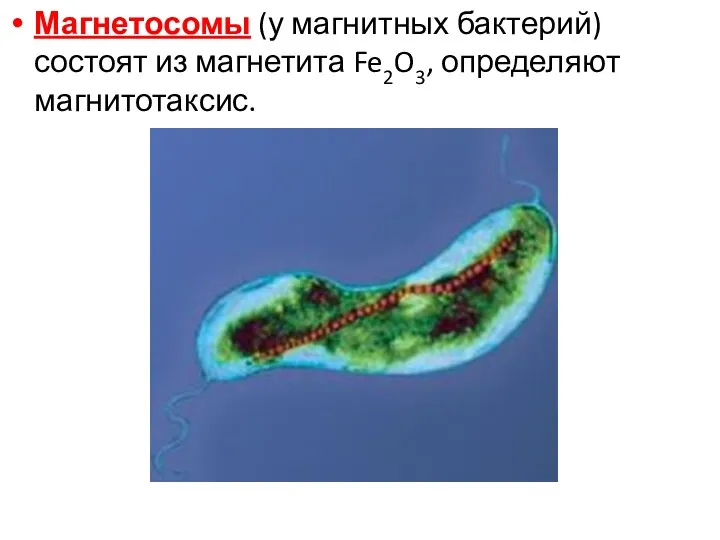
Магнетосомы (у магнитных бактерий) состоят из магнетита Fe2O3, определяют магнитотаксис.
Магнетосомы (у магнитных бактерий) состоят из магнетита Fe2O3, определяют магнитотаксис.
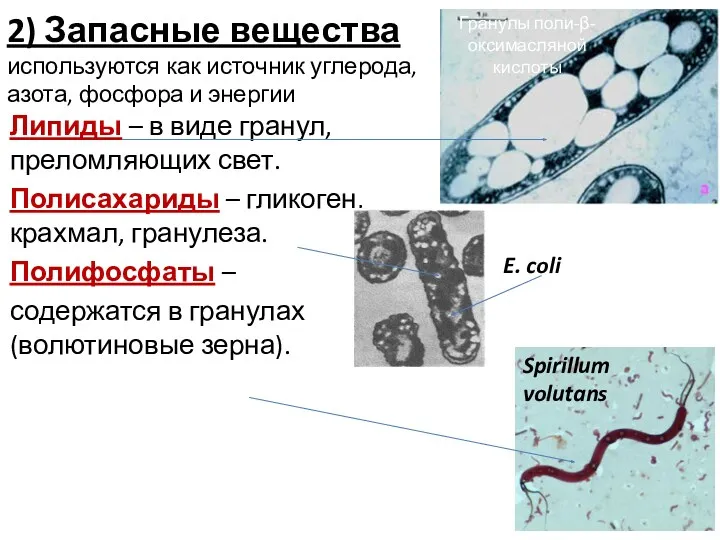
2) Запасные вещества используются как источник углерода, азота, фосфора и энергии
Липиды
2) Запасные вещества используются как источник углерода, азота, фосфора и энергии
Липиды

Сера (у серных бактерий) - в виде глобул.
Белки (цианофицин) – у
Сера (у серных бактерий) - в виде глобул.
Белки (цианофицин) – у

Bacillus thuringiensis используется для борьбы с вредными насекомыми
Bacillus thuringiensis используется для борьбы с вредными насекомыми

Внутрицитоплазматические мембраны (ВЦМ)
ВЦМ образуются в результате локальной инвагинации ЦПМ в
Внутрицитоплазматические мембраны (ВЦМ)
ВЦМ образуются в результате локальной инвагинации ЦПМ в
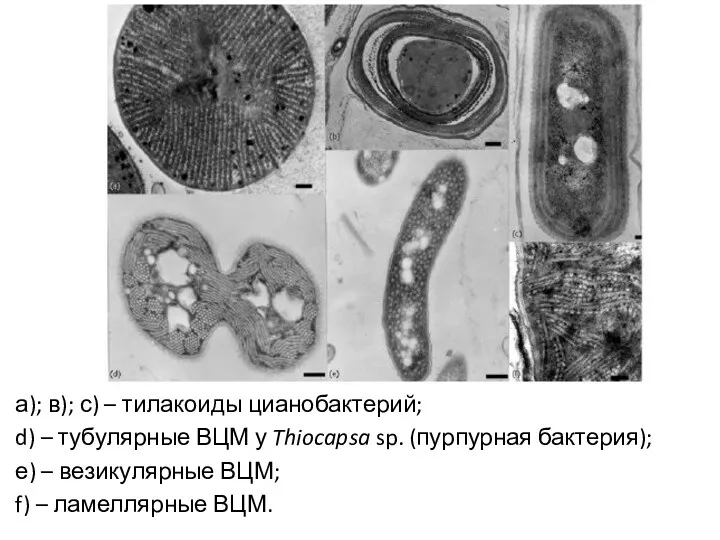
а); в); с) – тилакоиды цианобактерий;
d) – тубулярные ВЦМ у Thiocapsa
а); в); с) – тилакоиды цианобактерий;
d) – тубулярные ВЦМ у Thiocapsa

2. Клеточная оболочка
ЦПМ
Химический состав:
белки – 50 - 75 %,
2. Клеточная оболочка
ЦПМ
Химический состав:
белки – 50 - 75 %,
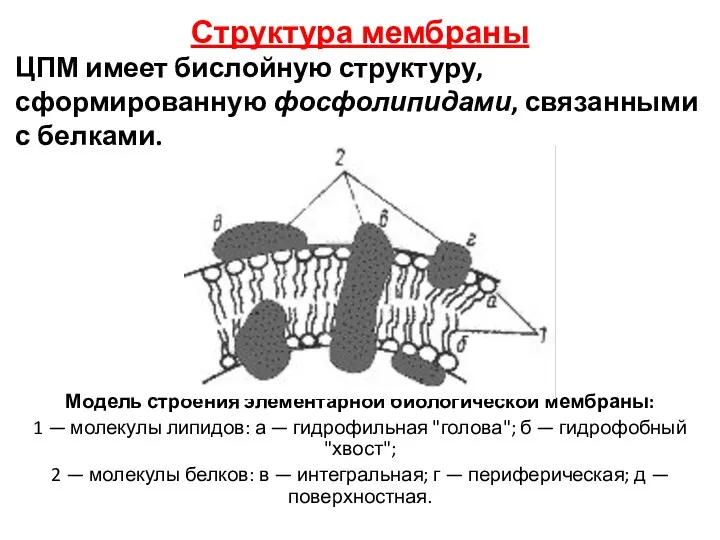
Модель строения элементарной биологической мембраны:
1 — молекулы липидов: а —
Модель строения элементарной биологической мембраны:
1 — молекулы липидов: а —
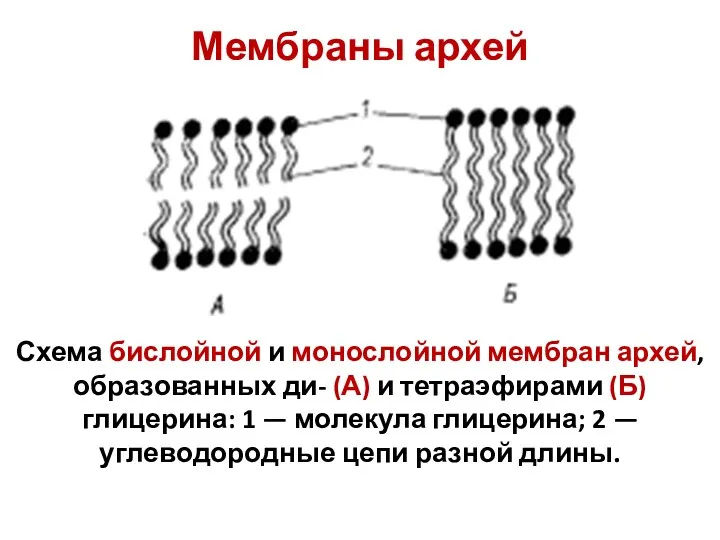
Мембраны архей
Схема бислойной и монослойной мембран архей, образованных ди- (А) и
Мембраны архей
Схема бислойной и монослойной мембран архей, образованных ди- (А) и

Функции ЦПМ
1. Барьерная.
2. Транспортная.
3. Осмотическая.
4. Метаболическая (участвует в энергетических процессах,
Функции ЦПМ
1. Барьерная.
2. Транспортная.
3. Осмотическая.
4. Метаболическая (участвует в энергетических процессах,

Клеточная стенка (КС)
КС – обязательный структурный элемент, придает клеткам форму.
КС
Клеточная стенка (КС)
КС – обязательный структурный элемент, придает клеткам форму.
КС
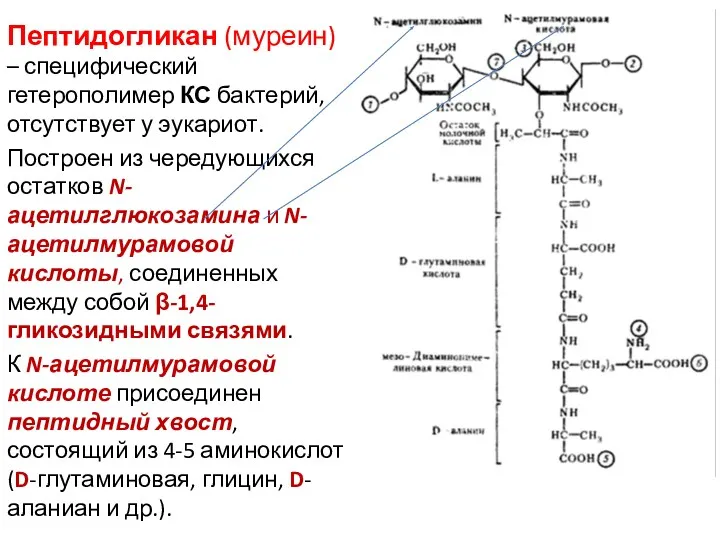
Пептидогликан (муреин) – специфический гетерополимер КС бактерий, отсутствует у эукариот.
Построен из
Пептидогликан (муреин) – специфический гетерополимер КС бактерий, отсутствует у эукариот.
Построен из

Пептидогликан – линейный полимер, его молекулы образуют сеть из параллельно расположенных
Пептидогликан – линейный полимер, его молекулы образуют сеть из параллельно расположенных

Прокариоты в зависимости от структуры и химического состава клеточной стенки делят
Прокариоты в зависимости от структуры и химического состава клеточной стенки делят
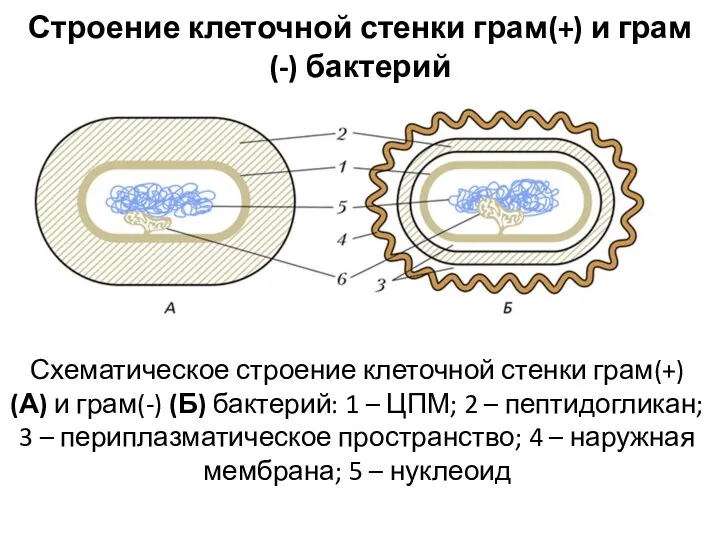
Строение клеточной стенки грам(+) и грам(-) бактерий
Схематическое строение клеточной стенки грам(+)
Строение клеточной стенки грам(+) и грам(-) бактерий
Схематическое строение клеточной стенки грам(+)

Химический состав
Структура
пептидогликан (п/гл) – 40-90 %,
тейхоевые кислоты (полимеры из
Химический состав
Структура
пептидогликан (п/гл) – 40-90 %,
тейхоевые кислоты (полимеры из

ГРАМ(+) БАКТЕРИИ:
р. Bacillus – B. anthracis (возбудитель сибирской язвы), B.
ГРАМ(+) БАКТЕРИИ:
р. Bacillus – B. anthracis (возбудитель сибирской язвы), B.

КЛЕТОЧНАЯ СТЕНКА ГРАМ(-) БАКТЕРИЙ
Химический состав
Структура
пептидогликан (п/гл) –
1 - 10
КЛЕТОЧНАЯ СТЕНКА ГРАМ(-) БАКТЕРИЙ
Химический состав
Структура
пептидогликан (п/гл) –
1 - 10

ГРАМ(-) БАКТЕРИИ:
Escherichia coli
Pseudomonas aeruginosa – синегнойная палочка
Neisseria
ГРАМ(-) БАКТЕРИИ:
Escherichia coli
Pseudomonas aeruginosa – синегнойная палочка
Neisseria
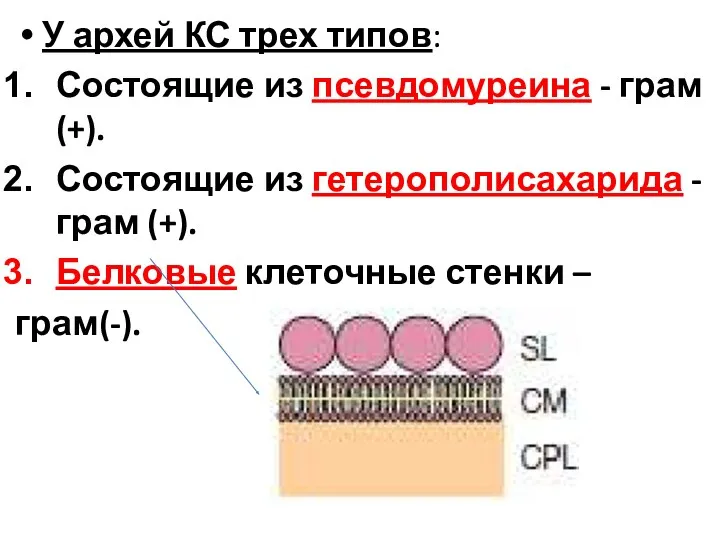
У архей КС трех типов:
Состоящие из псевдомуреина - грам (+).
Состоящие из
У архей КС трех типов:
Состоящие из псевдомуреина - грам (+).
Состоящие из

Функции клеточной стенки прокариот:
1. Придает клеткам определенную форму.
2. Механическая.
3. Осмотическая.
3. Транспортная.
4.
Функции клеточной стенки прокариот:
1. Придает клеткам определенную форму.
2. Механическая.
3. Осмотическая.
3. Транспортная.
4.

3. Поверхностные структуры клетки
Капсула - слизистое образование, располагается поверх клеточной стенки,
3. Поверхностные структуры клетки
Капсула - слизистое образование, располагается поверх клеточной стенки,
Капсулы бактерий
Капсулы бактерий

Слизистые слои – имеют аморфное строение, легко отделяются от клетки.
У роящихся
Слизистые слои – имеют аморфное строение, легко отделяются от клетки.
У роящихся

Жгутики
Имеются у подвижных бактерий.
Жгутик состоит из 3 компонентов:
Длинная спиральная
Жгутики
Имеются у подвижных бактерий.
Жгутик состоит из 3 компонентов:
Длинная спиральная

У грам(+) – М- и S-кольцо:
М-кольцо в ЦПМ,
S- - в
У грам(+) – М- и S-кольцо:
М-кольцо в ЦПМ,
S- - в

Движущей силой вращения колец служит протонный градиент.
При вращении колец их движение
Движущей силой вращения колец служит протонный градиент.
При вращении колец их движение

Микроворсинки (фимбрии, пили)
Это нитевидные клеточные придатки диаметром 3-10 нм, длиной -
Микроворсинки (фимбрии, пили)
Это нитевидные клеточные придатки диаметром 3-10 нм, длиной -
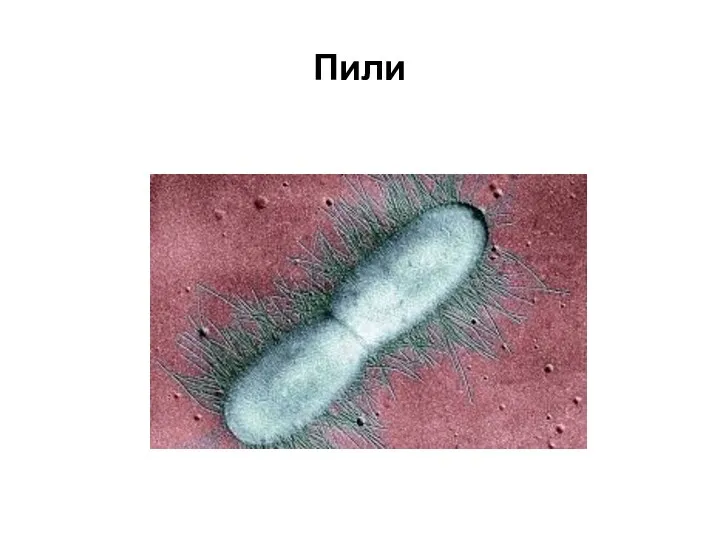
Пили
Пили

4. Типы движения прокариот и таксисы
Типы движения:
1. Плавание – передвижение бактерий
4. Типы движения прокариот и таксисы
Типы движения:
1. Плавание – передвижение бактерий

4. Подтягивающее движение - движение по твердой поверхности с участием пили
4. Подтягивающее движение - движение по твердой поверхности с участием пили

Таксисы
1. Хемотаксис – движение бактерий в определенном направлении относительно источника химического
Таксисы
1. Хемотаксис – движение бактерий в определенном направлении относительно источника химического

5. Магнитотаксис – у магнитобактерий, содержат магнетосомы, движение определяется силовыми линиями
5. Магнитотаксис – у магнитобактерий, содержат магнетосомы, движение определяется силовыми линиями

МОРФОЛОГИЧЕСКИ ДИФФЕРЕНЦИРОВАННЫЕ КЛЕТКИ ПРОКАРИОТ (МДК)
МОРФОЛОГИЧЕСКИ ДИФФЕРЕНЦИРОВАННЫЕ КЛЕТКИ ПРОКАРИОТ (МДК)

Вопросы:
Цисты и акинеты.
Эндоспоры бактерий.
Экзоспоры бактерий и актиномицетов.
Вопросы:
Цисты и акинеты.
Эндоспоры бактерий.
Экзоспоры бактерий и актиномицетов.

Вегетативные клетки многих прокариот при определенных условиях дают начало структурам, которые
Вегетативные клетки многих прокариот при определенных условиях дают начало структурам, которые

К МДК прокариот относятся:
Покоящиеся формы – эндоспоры грам(+) бактерий, цисты, акинеты
К МДК прокариот относятся:
Покоящиеся формы – эндоспоры грам(+) бактерий, цисты, акинеты

1. Цисты и акинеты
Цисты образуются у миксобактерий, азотобактера, риккетсий и др.
У
1. Цисты и акинеты
Цисты образуются у миксобактерий, азотобактера, риккетсий и др.
У

Акинеты цианобактерий
Акинеты крупнее вегетативных клеток, продолговатой или сферической формы, с
Акинеты цианобактерий
Акинеты крупнее вегетативных клеток, продолговатой или сферической формы, с
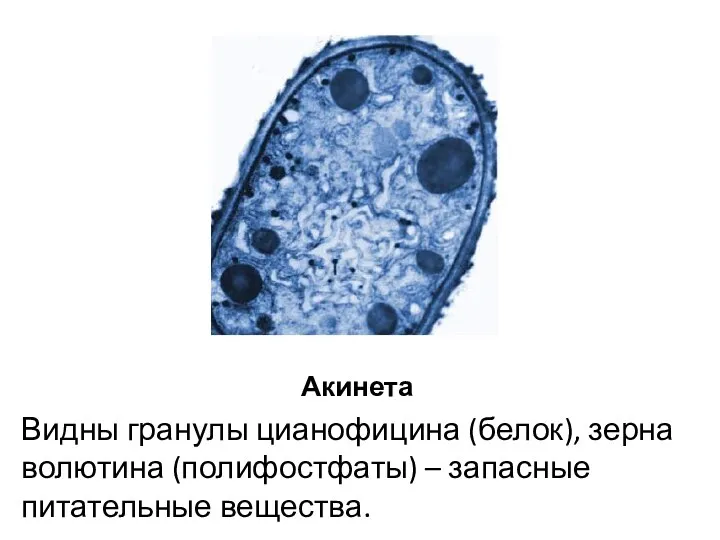
Акинета
Видны гранулы цианофицина (белок), зерна волютина (полифостфаты) – запасные питательные вещества.
Акинета
Видны гранулы цианофицина (белок), зерна волютина (полифостфаты) – запасные питательные вещества.

2. Эндоспоры бактерий
Бактериальные эндоспоры – особый тип покоящихся клеток в основном
2. Эндоспоры бактерий
Бактериальные эндоспоры – особый тип покоящихся клеток в основном
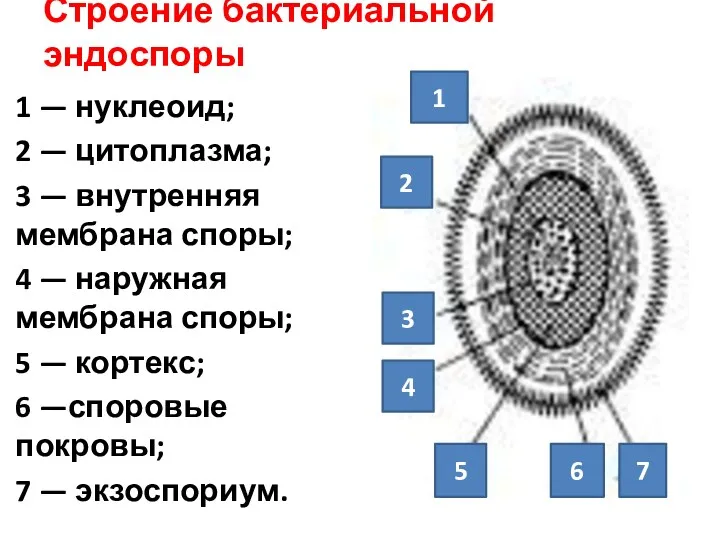
Строение бактериальной эндоспоры
1 — нуклеоид;
2 — цитоплазма;
3 — внутренняя
Строение бактериальной эндоспоры
1 — нуклеоид;
2 — цитоплазма;
3 — внутренняя

Химический состав эндоспор
Белки эндоспор богаты цистеином и гидрофобными аминокислотами.
Содержание ДНК ниже,
Химический состав эндоспор
Белки эндоспор богаты цистеином и гидрофобными аминокислотами.
Содержание ДНК ниже,

Содержание РНК в спорах ниже, чем в вегетативных клетках.
В спорах
Содержание РНК в спорах ниже, чем в вегетативных клетках.
В спорах

Кортекс построен из молекул особого типа пептидогликана. При прорастании споры из
Кортекс построен из молекул особого типа пептидогликана. При прорастании споры из

Устойчивость эндоспор
Устойчивы к высоким (+80 °С) и низким температурам, обезвоживанию, высокой
Устойчивость эндоспор
Устойчивы к высоким (+80 °С) и низким температурам, обезвоживанию, высокой

Факторы, обеспечивающие устойчивость эндоспор:
1. Нахождение споровой цитоплазмы в обезвоженном состоянии.
2. Термостойкость споровых ферментов.
3. Наличие
Факторы, обеспечивающие устойчивость эндоспор:
1. Нахождение споровой цитоплазмы в обезвоженном состоянии.
2. Термостойкость споровых ферментов.
3. Наличие

3. Экзоспоры бактерий и актиномицетов
Экзоспоры у бактерий образуются редко, например, у
3. Экзоспоры бактерий и актиномицетов
Экзоспоры у бактерий образуются редко, например, у
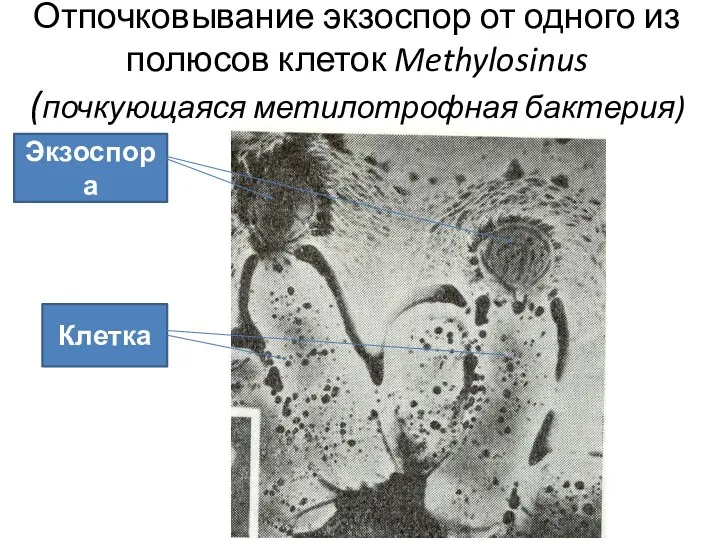
Отпочковывание экзоспор от одного из полюсов клеток Methylosinus (почкующаяся метилотрофная бактерия)
Экзоспора
Клетка
Отпочковывание экзоспор от одного из полюсов клеток Methylosinus (почкующаяся метилотрофная бактерия)
Экзоспора
Клетка

Экзоспоры имеют более плотную и утолщенную клеточную стенку.
В экзоспорах нет дипиколиновой
Экзоспоры имеют более плотную и утолщенную клеточную стенку.
В экзоспорах нет дипиколиновой
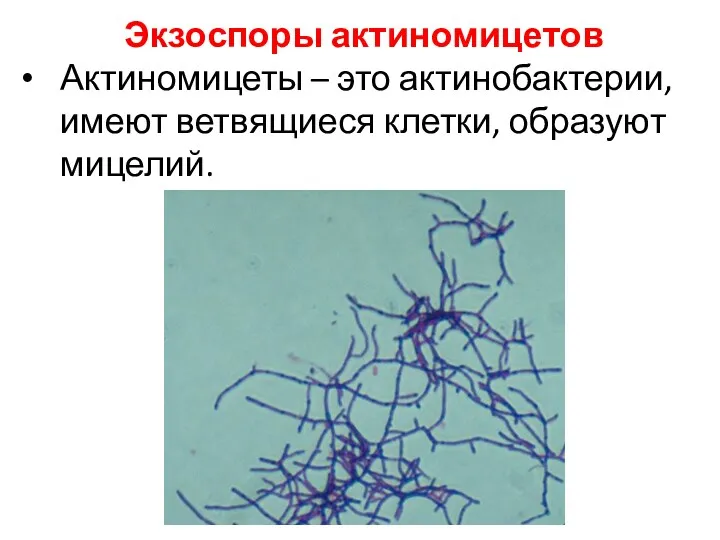
Экзоспоры актиномицетов
Актиномицеты – это актинобактерии, имеют ветвящиеся клетки, образуют мицелий.
Экзоспоры актиномицетов
Актиномицеты – это актинобактерии, имеют ветвящиеся клетки, образуют мицелий.
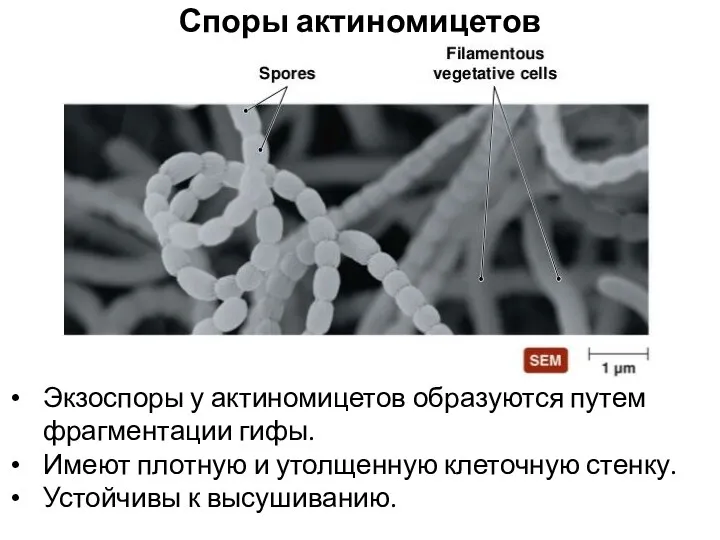
Споры актиномицетов
Экзоспоры у актиномицетов образуются путем фрагментации гифы.
Имеют плотную и утолщенную
Споры актиномицетов
Экзоспоры у актиномицетов образуются путем фрагментации гифы.
Имеют плотную и утолщенную
 Зеленые водоросли
Зеленые водоросли Строение и функции органов дыхания
Строение и функции органов дыхания Клас Павукоподібні
Клас Павукоподібні Среды обитания живых организмов
Среды обитания живых организмов Что мы не знаем о грибах.
Что мы не знаем о грибах. Моногибридное скрещивание.Задачи.
Моногибридное скрещивание.Задачи. Пищеварительная система. Общий план строения стенки пишеварительной трубки
Пищеварительная система. Общий план строения стенки пишеварительной трубки Вегетативті жүйке жүйесі
Вегетативті жүйке жүйесі Служебные собаки
Служебные собаки Жиры, Белки и Углеводы человека
Жиры, Белки и Углеводы человека Есте сақтау қабілетінің биохимиялық механизмі
Есте сақтау қабілетінің биохимиялық механизмі Викторина по биологии и химии (вопросы)
Викторина по биологии и химии (вопросы) Біологія та майбутнє людства
Біологія та майбутнє людства Такие разные полезные бактерии
Такие разные полезные бактерии Бабочка Адмирал
Бабочка Адмирал Безусловные рефлексы, их значение. Условные рефлексы, образование и торможение
Безусловные рефлексы, их значение. Условные рефлексы, образование и торможение Регуляція вісцеральних функцій організму. Фізіологія автономної нервової системи
Регуляція вісцеральних функцій організму. Фізіологія автономної нервової системи Понятие об организме. Доядерные и ядерные организмы
Понятие об организме. Доядерные и ядерные организмы Органы и системы органов
Органы и системы органов ПРЕДПРОФИЛЬНЫЙ КУРС ХИМИЯ И ПРАКТИЧЕСКОЕ РАТЕНИЕВОДСТВО КАК СРЕДСТВО ФОРМИРОВАНИЯ УУД И РАЗВИТИЯ ТВОРЧЕСКОЙ АКТИВНОСТИ ОБУЧАЮЩИХСЯ
ПРЕДПРОФИЛЬНЫЙ КУРС ХИМИЯ И ПРАКТИЧЕСКОЕ РАТЕНИЕВОДСТВО КАК СРЕДСТВО ФОРМИРОВАНИЯ УУД И РАЗВИТИЯ ТВОРЧЕСКОЙ АКТИВНОСТИ ОБУЧАЮЩИХСЯ Гистология. Электронный альбом
Гистология. Электронный альбом Энзимология. Структура и механизм действия ферментов. (Лекция 2)
Энзимология. Структура и механизм действия ферментов. (Лекция 2) Предпосылки эволюционных преобразований органов. Аномалии и пороки развития
Предпосылки эволюционных преобразований органов. Аномалии и пороки развития Определение пола. Наследование признаков, сцепленных с полом
Определение пола. Наследование признаков, сцепленных с полом Приспособления организмов к среде обитания
Приспособления организмов к среде обитания Structure of DNA and its function
Structure of DNA and its function Кровь
Кровь Разработка урока по биологии Углеводы и липиды 9 класс (проектная деятельность)
Разработка урока по биологии Углеводы и липиды 9 класс (проектная деятельность)