- Есте сақтау қабілетінің биохимиялық механизмі

Содержание
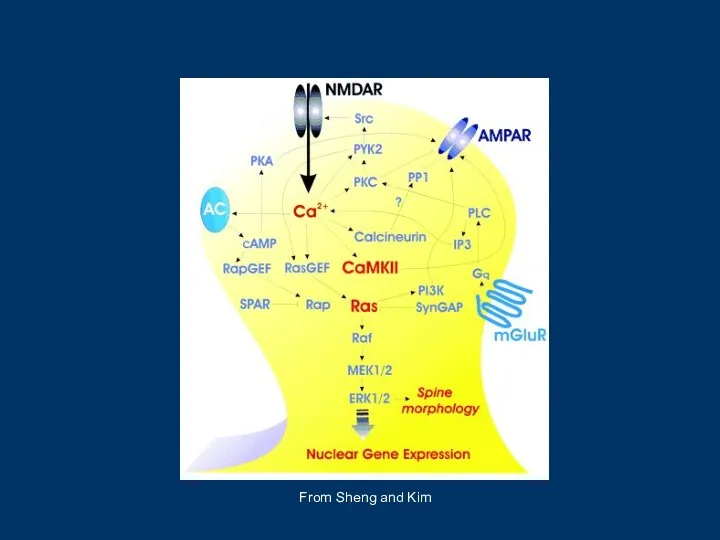
- 3. From Sheng and Kim
- 4. Neurotransmitter Receptor NMDA Receptor AMPA Receptor K Channels Ca++ Channels IP3 Receptor 1 2 2 3
- 5. 1. NMDA рецепторының функциясын тікелей реттейтін NMDA рецепторларының үстіндегі механизмдер. 2. Мембранадағы деполяризацияны басқаратын NMDA рецепторларының
- 7. Husi et al. (2001) Nature Neuroscience 3: 661-669.
- 8. TABLE I – DIRECT MODULATORS OF THE NMDA RECEPTOR
- 9. Receptor Modulation of the NMDA receptor
- 10. TABLE II – MECHANISMS UPSTREAM OF THE NMDA RECEPTOR INVOLVED IN MEMBRANE DEPOLARIZATION
- 11. Үш жолды сәйкестендіру ↓Kv4.2 Strong Input Back propagating Action Potential ACh CA1 Pyramidal Neuron NMDAR Glu
- 12. TABLE III – COMPONENTS OF THE SYNAPTIC INFRASTRUCTURE NECESSARY FOR NMDA RECEPTOR FUNCTION
- 13. TABLE III – COMPONENTS OF THE SYNAPTIC INFRASTRUCTURE NECESSARY FOR NMDA RECEPTOR FUNCTION ( Continued)
- 14. PSD-95 as an Anchoring Protein for NMDA Receptors
- 15. From Sheng and Kim
- 20. Fig. 1RIM1 және синаптические везикулды біріктіру қабынуы. (а) Қондырылғаннан кейін синапстык везикул (SV) ядродағы Раб3-ні (N)
- 22. Жоғарғы панельдер CA1 гиппокампусынан (А) және ферментті-эозин фототрансформациясын пайдалана отырып, F-актинге арналған электронды-томографиялық омыртқа көлемінің жеке
- 23. Figure 1. LIMK Influences Postsynaptic and Presynaptic Function through Modulation of Actin Filaments Dendritic spines are
- 24. Интегриндер мен жасуша ішіндегі эффекторлар арасындағы өзара әрекеттесулер
- 25. Кальций механизмі
- 27. Кальций реакциясының сыртқы сигналдарын модификациялау
- 28. Model for the cAMP Gate Sweatt (2001) Curr. Biol. 11:R391-394.
- 29. Phospholipase C Neurogranin Neurogranin PO4 + DAG PKC Calmodulin Calmodulin Metabotropic Receptor PKC Phosphorylation of Neurogranin
- 30. Augmented PKC cAMP GATE Initial Ca++ Signal Increased Ca++/CaM Augmented CaMKII Activity Adenylyl Cyclase NMDAR Neurogranin
- 32. Скачать презентацию
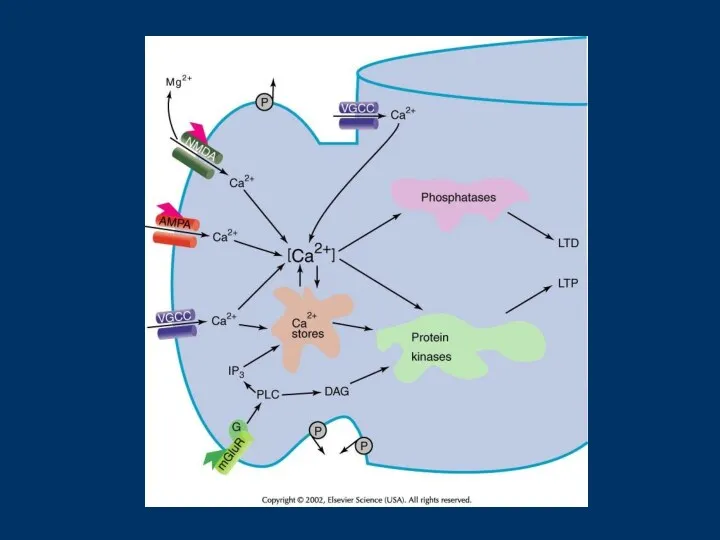
From Sheng and Kim
From Sheng and Kim
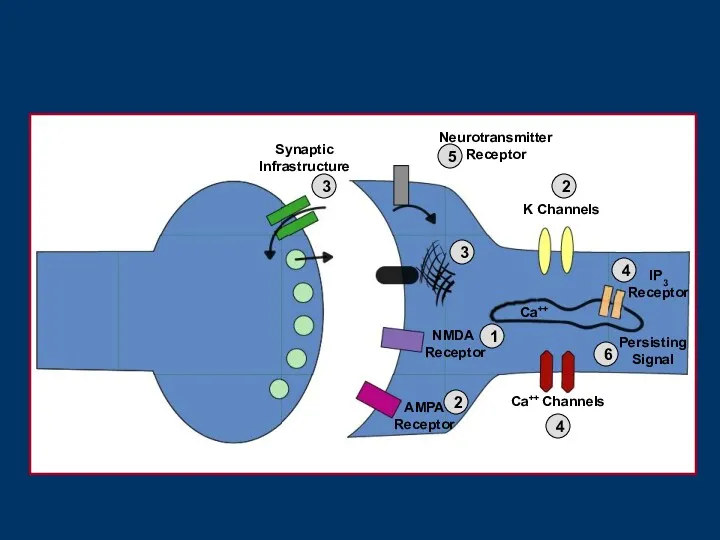
Neurotransmitter
Receptor
NMDA
Receptor
AMPA
Receptor
K Channels
Ca++ Channels
IP3
Receptor
1
2
2
3
3
4
4
5
Synaptic
Infrastructure
Ca++
6
Persisting
Signal
Neurotransmitter
Receptor
NMDA
Receptor
AMPA
Receptor
K Channels
Ca++ Channels
IP3
Receptor
1
2
2
3
3
4
4
5
Synaptic
Infrastructure
Ca++
6
Persisting
Signal

1. NMDA рецепторының функциясын тікелей реттейтін NMDA рецепторларының үстіндегі механизмдер.
2. Мембранадағы
2. Мембранадағы

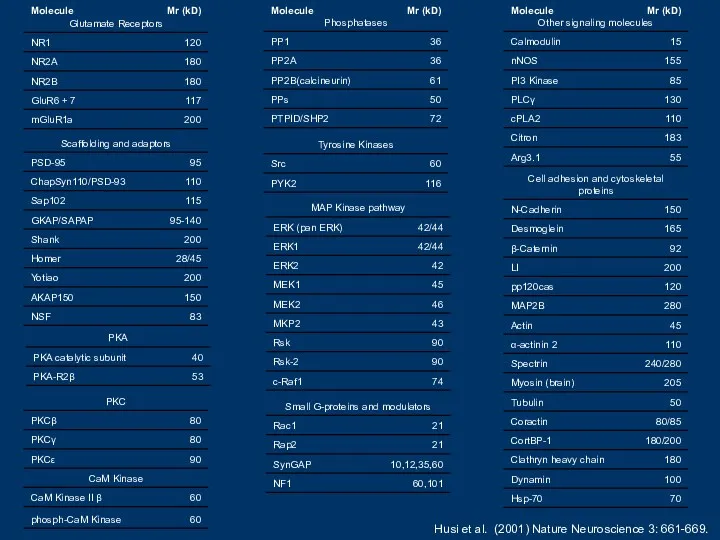
Husi et al. (2001) Nature Neuroscience 3: 661-669.
Husi et al. (2001) Nature Neuroscience 3: 661-669.
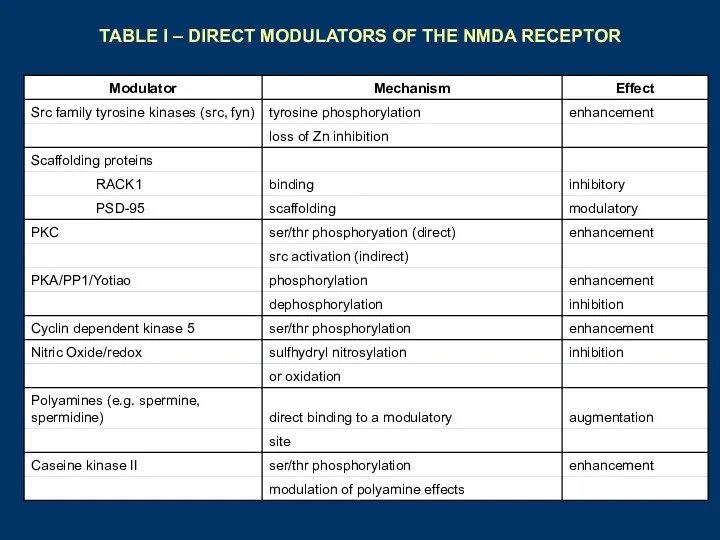
TABLE I – DIRECT MODULATORS OF THE NMDA RECEPTOR
TABLE I – DIRECT MODULATORS OF THE NMDA RECEPTOR
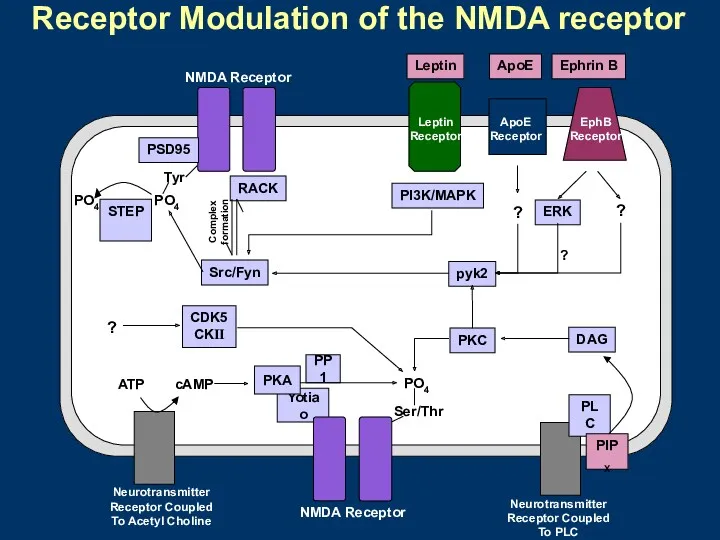
Receptor Modulation of the NMDA receptor
Receptor Modulation of the NMDA receptor
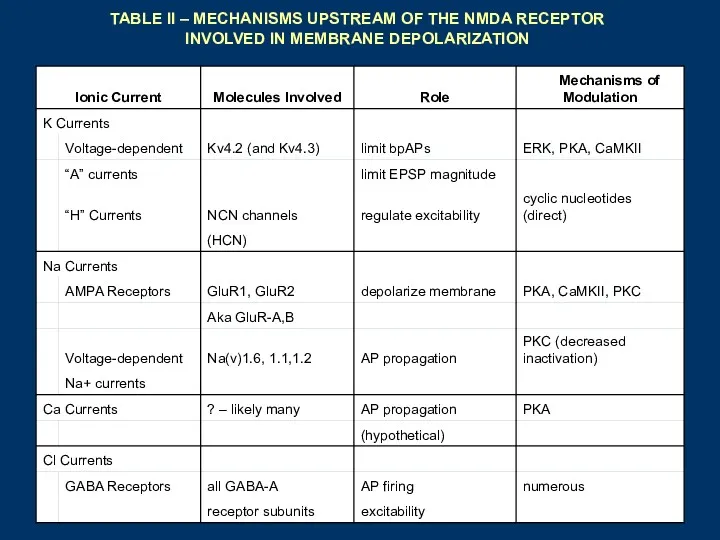
TABLE II – MECHANISMS UPSTREAM OF THE NMDA RECEPTOR INVOLVED IN
TABLE II – MECHANISMS UPSTREAM OF THE NMDA RECEPTOR INVOLVED IN
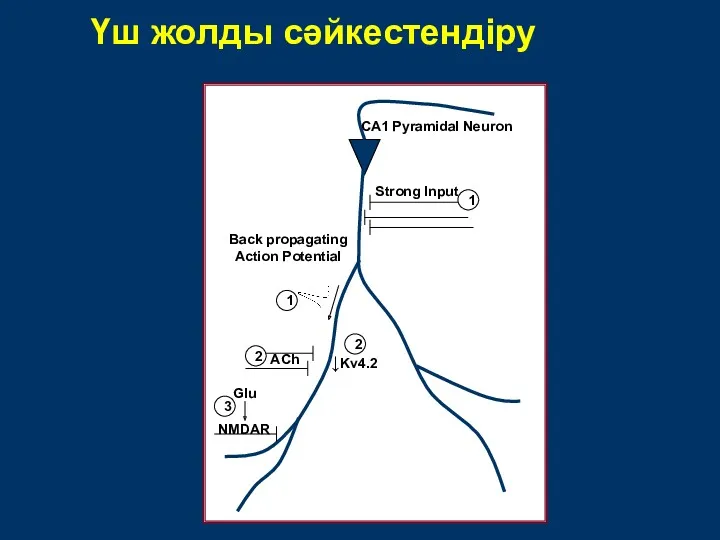
Үш жолды сәйкестендіру
↓Kv4.2
Strong Input
Back propagating
Action Potential
ACh
CA1 Pyramidal Neuron
NMDAR
Glu
1
1
2
2
3
Үш жолды сәйкестендіру
↓Kv4.2
Strong Input
Back propagating
Action Potential
ACh
CA1 Pyramidal Neuron
NMDAR
Glu
1
1
2
2
3
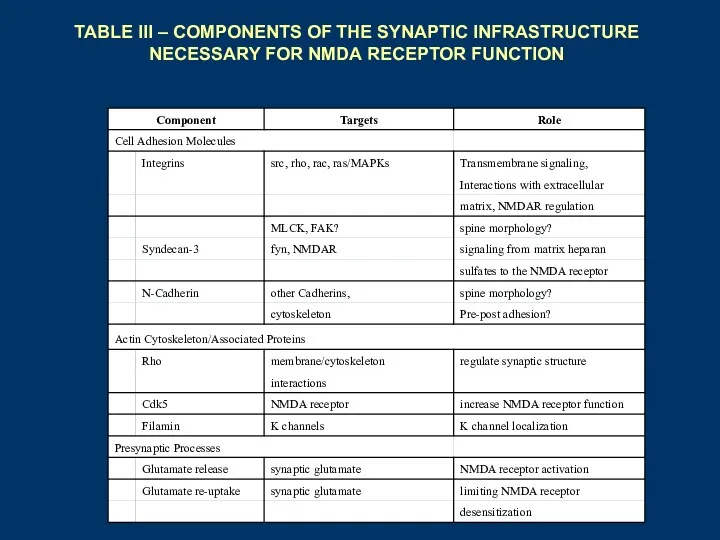
TABLE III – COMPONENTS OF THE SYNAPTIC INFRASTRUCTURE NECESSARY FOR NMDA
TABLE III – COMPONENTS OF THE SYNAPTIC INFRASTRUCTURE NECESSARY FOR NMDA
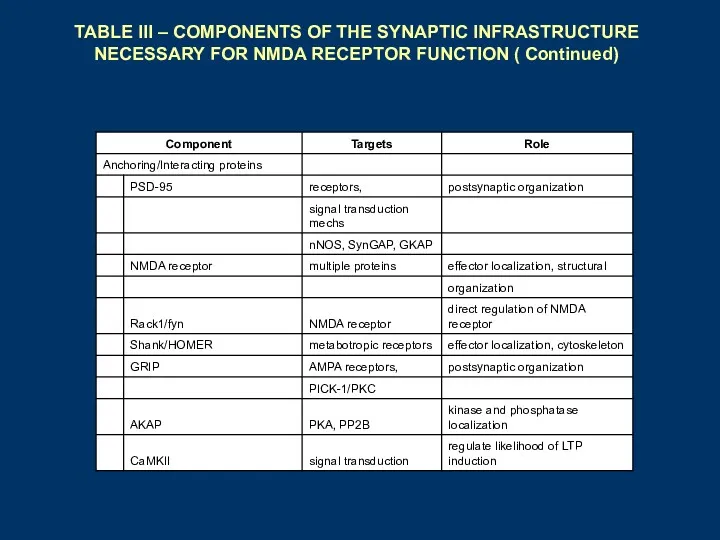
TABLE III – COMPONENTS OF THE SYNAPTIC INFRASTRUCTURE NECESSARY FOR NMDA
TABLE III – COMPONENTS OF THE SYNAPTIC INFRASTRUCTURE NECESSARY FOR NMDA
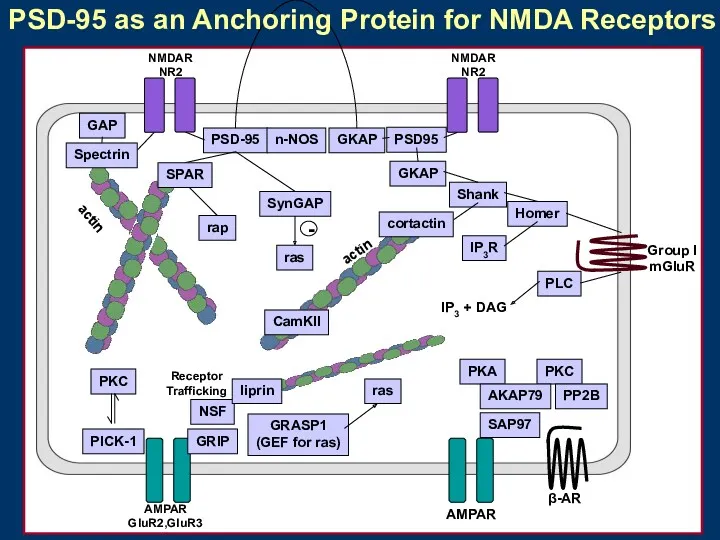
PSD-95 as an Anchoring Protein for NMDA Receptors
PSD-95 as an Anchoring Protein for NMDA Receptors
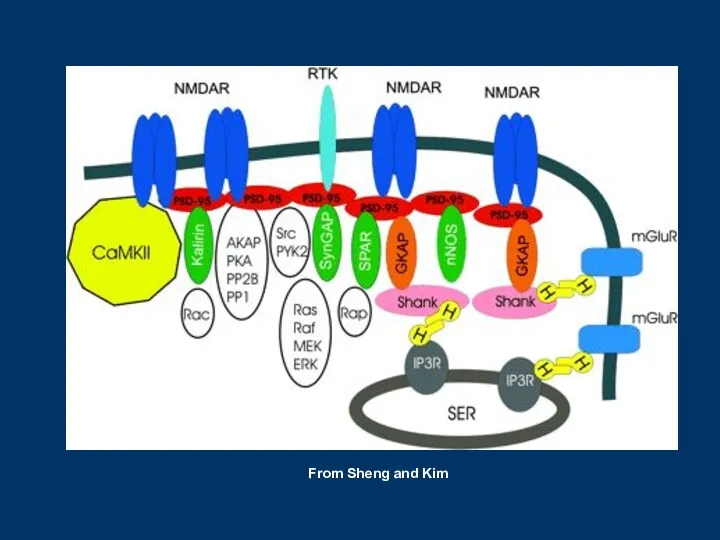
From Sheng and Kim
From Sheng and Kim

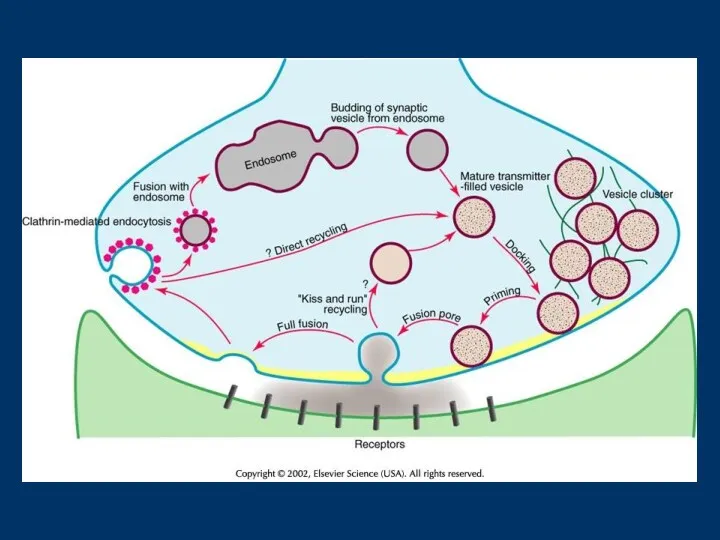


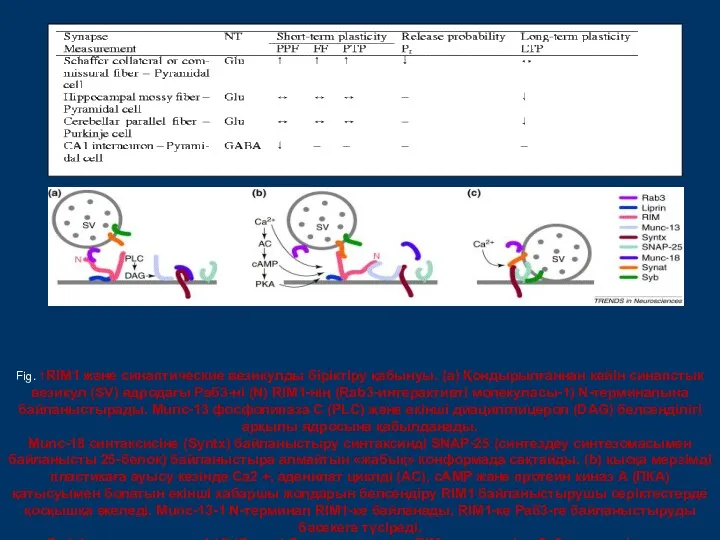
Fig. 1RIM1 және синаптические везикулды біріктіру қабынуы. (а) Қондырылғаннан кейін синапстык
Fig. 1RIM1 және синаптические везикулды біріктіру қабынуы. (а) Қондырылғаннан кейін синапстык

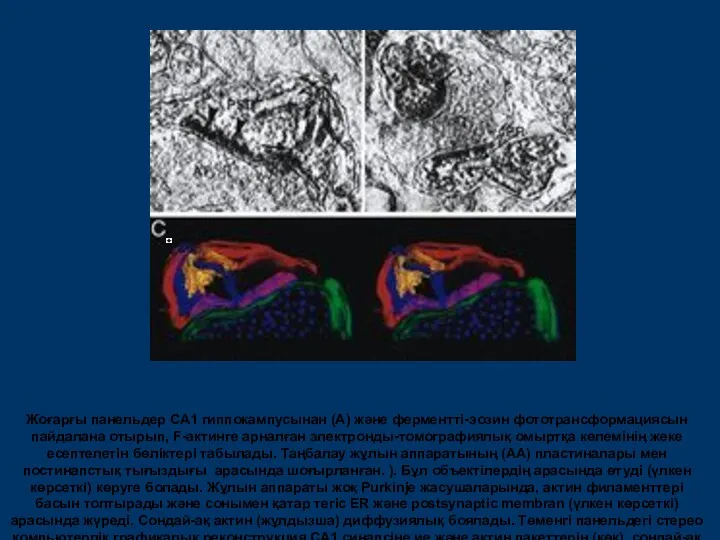
Жоғарғы панельдер CA1 гиппокампусынан (А) және ферментті-эозин фототрансформациясын пайдалана отырып, F-актинге
Жоғарғы панельдер CA1 гиппокампусынан (А) және ферментті-эозин фототрансформациясын пайдалана отырып, F-актинге
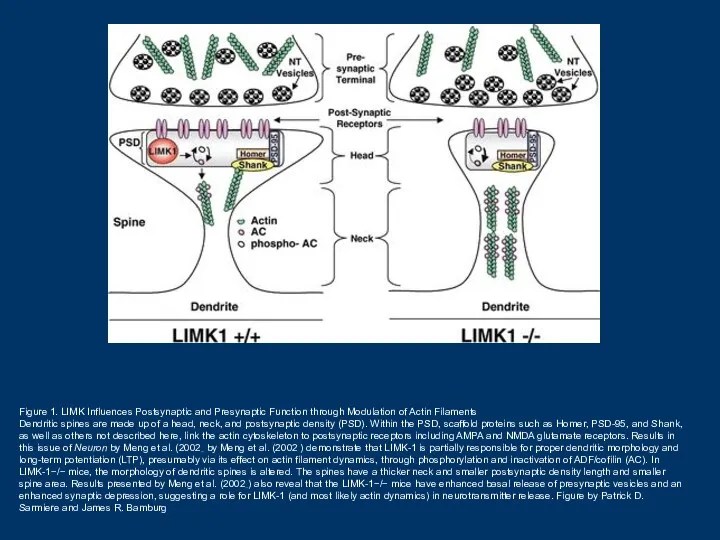
Figure 1. LIMK Influences Postsynaptic and Presynaptic Function through Modulation of
Figure 1. LIMK Influences Postsynaptic and Presynaptic Function through Modulation of
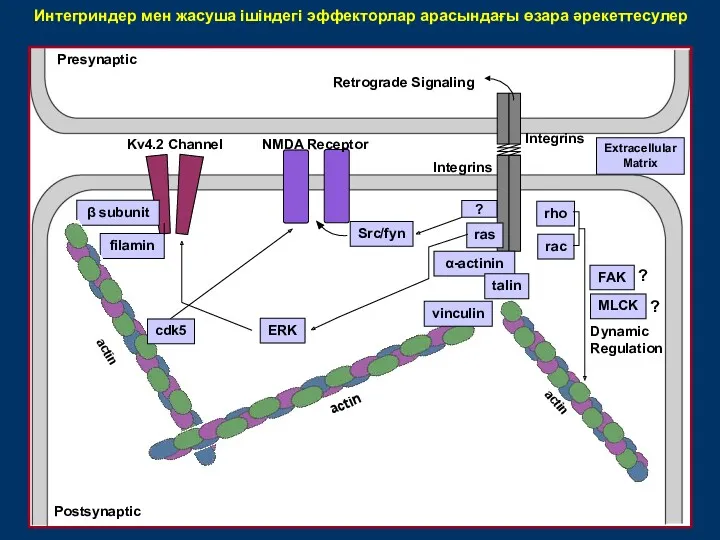
Интегриндер мен жасуша ішіндегі эффекторлар арасындағы өзара әрекеттесулер
Интегриндер мен жасуша ішіндегі эффекторлар арасындағы өзара әрекеттесулер
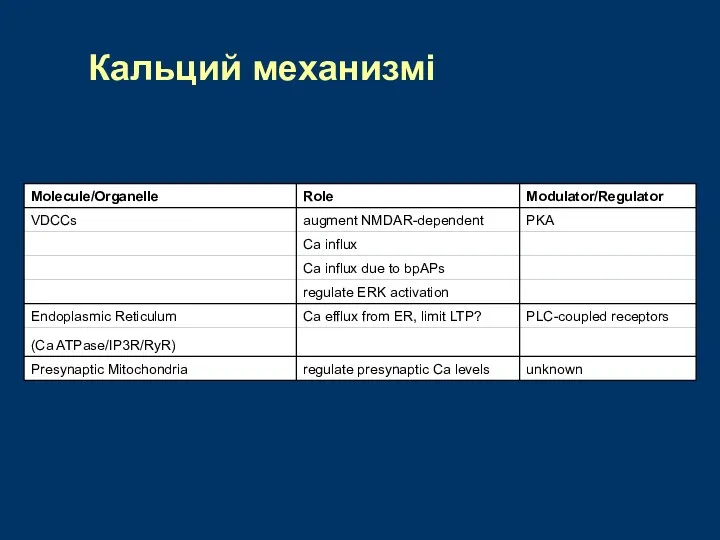
Кальций механизмі
Кальций механизмі

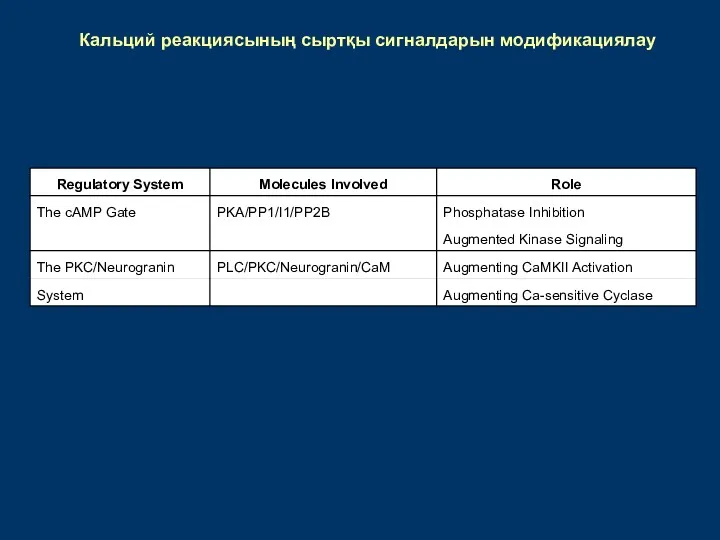
Кальций реакциясының сыртқы сигналдарын модификациялау
Кальций реакциясының сыртқы сигналдарын модификациялау
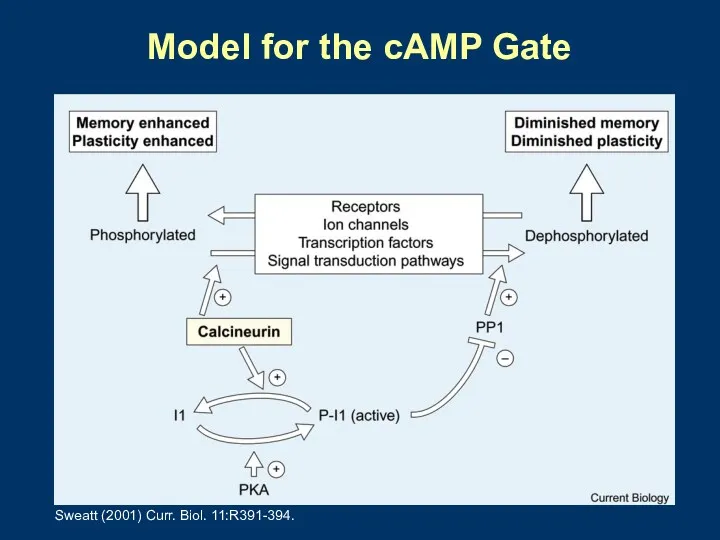
Model for the cAMP Gate
Sweatt (2001) Curr. Biol. 11:R391-394.
Model for the cAMP Gate
Sweatt (2001) Curr. Biol. 11:R391-394.
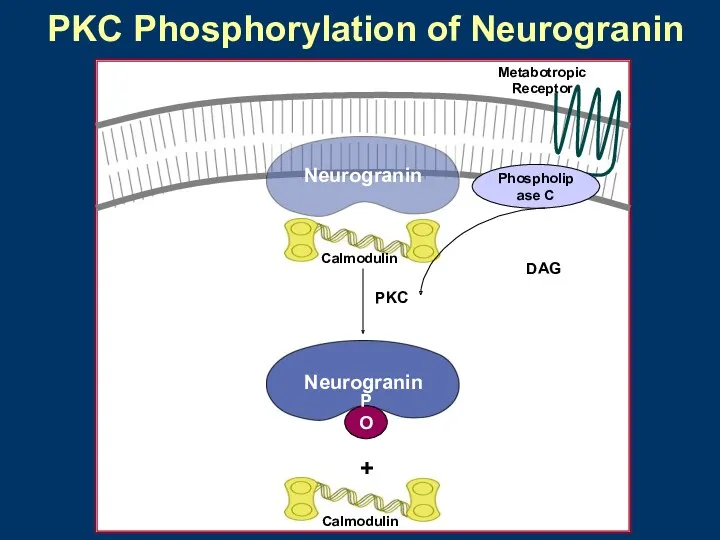
Phospholipase C
Neurogranin
Neurogranin
PO4
+
DAG
PKC
Calmodulin
Calmodulin
Metabotropic
Receptor
PKC Phosphorylation of Neurogranin
Phospholipase C
Neurogranin
Neurogranin
PO4
+
DAG
PKC
Calmodulin
Calmodulin
Metabotropic
Receptor
PKC Phosphorylation of Neurogranin
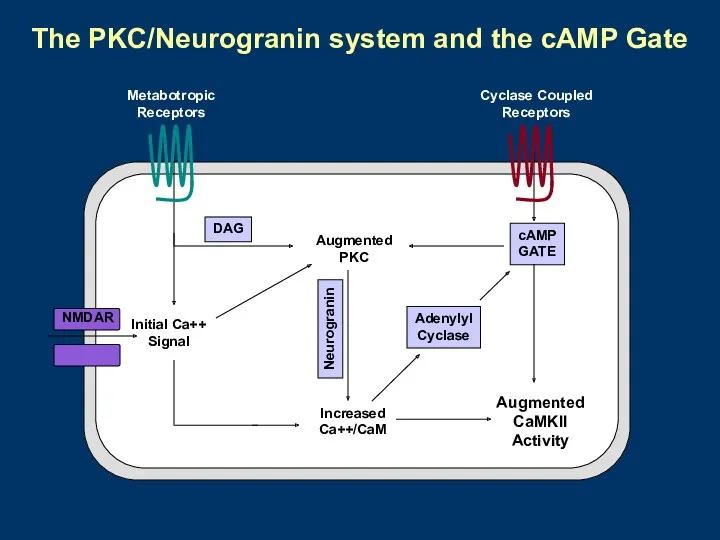
Augmented
PKC
cAMP
GATE
Initial Ca++
Signal
Increased
Ca++/CaM
Augmented
CaMKII
Activity
Adenylyl
Cyclase
NMDAR
Neurogranin
DAG
Cyclase Coupled
Receptors
Metabotropic
Receptors
The PKC/Neurogranin system and the cAMP Gate
Augmented
PKC
cAMP
GATE
Initial Ca++
Signal
Increased
Ca++/CaM
Augmented
CaMKII
Activity
Adenylyl
Cyclase
NMDAR
Neurogranin
DAG
Cyclase Coupled
Receptors
Metabotropic
Receptors
The PKC/Neurogranin system and the cAMP Gate
 Неклеточные формы жизни. Вирусы
Неклеточные формы жизни. Вирусы Кожа. Строение и функции кожи
Кожа. Строение и функции кожи Қос мекенділердің сыртқы және ішкі органдарының құрылысы
Қос мекенділердің сыртқы және ішкі органдарының құрылысы 20190315_tip_ploskie_chervi
20190315_tip_ploskie_chervi Высшие жирные кислоты. Липиды
Высшие жирные кислоты. Липиды Система растений и животных — отображение эволюции
Система растений и животных — отображение эволюции Взаємодія алельних генів
Взаємодія алельних генів Обмен веществ и энергии
Обмен веществ и энергии Викторина Животные живого уголка
Викторина Животные живого уголка Редуценты и их экологическая роль
Редуценты и их экологическая роль чому скисає молоко
чому скисає молоко Жылқыларды ұстауға арналған ғимараттарға қойылатын санитарлық-гигиеналық талаптар
Жылқыларды ұстауға арналған ғимараттарға қойылатын санитарлық-гигиеналық талаптар Женская половая система
Женская половая система Птицы - наши друзья
Птицы - наши друзья Явища природи. (1 класс)
Явища природи. (1 класс) Интегративная деятельность организма. Рефлексы
Интегративная деятельность организма. Рефлексы Мочеточники. Мочевой пузырь. Мочеиспускательный канал. Надпочечники
Мочеточники. Мочевой пузырь. Мочеиспускательный канал. Надпочечники презентация Пушистый доктор
презентация Пушистый доктор царство грибы
царство грибы Экологические группы птиц 3
Экологические группы птиц 3 Порівняння будови скелетів хребетних тварин
Порівняння будови скелетів хребетних тварин Человек. Пирамида потребностей человека (по А. Маслоу)
Человек. Пирамида потребностей человека (по А. Маслоу) Размножение многоклеточных животных
Размножение многоклеточных животных Возникновение и систематика растений
Возникновение и систематика растений Биология клетки в культуре. Оборудование и среды для работы с клеточными культурами
Биология клетки в культуре. Оборудование и среды для работы с клеточными культурами Презентация к уроку биологии в 5 классе Методы изучения природы
Презентация к уроку биологии в 5 классе Методы изучения природы Физиология микроорганизмов: дыхание, размножение. Микробиологические исследования: выделение чистой культуры аэробов, анаэробов
Физиология микроорганизмов: дыхание, размножение. Микробиологические исследования: выделение чистой культуры аэробов, анаэробов Угольная рыба
Угольная рыба