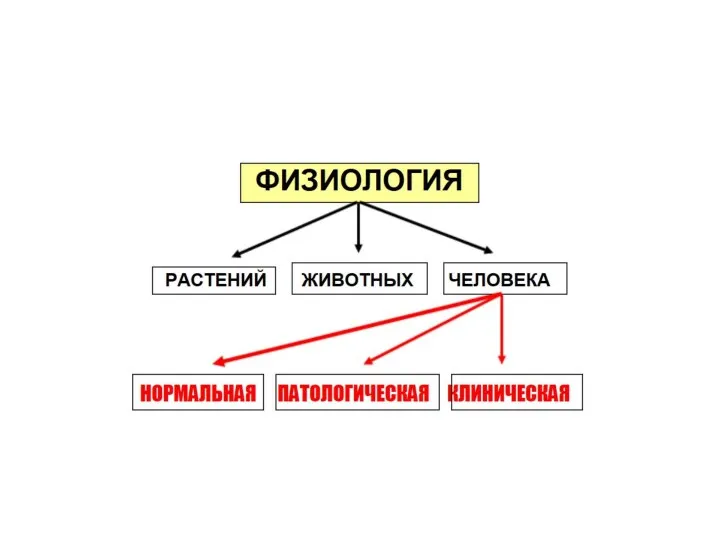
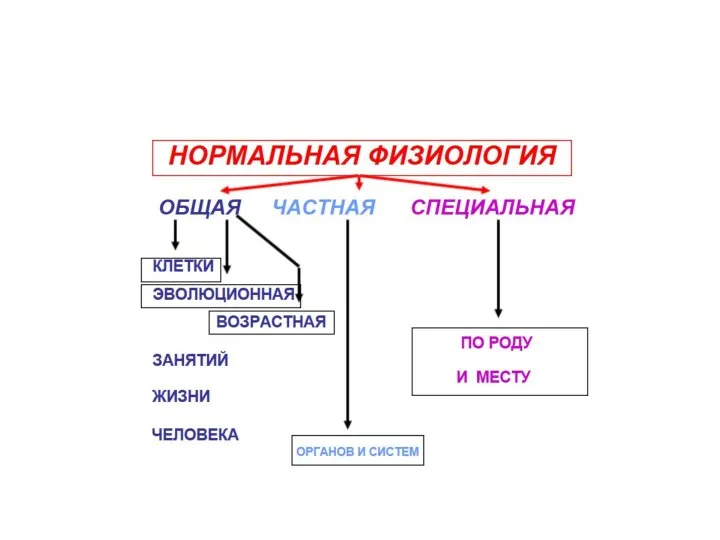
- Нормальная физиология

Содержание
- 14. Возбудимые ткани
- 15. Возбуждение – формирование специфической ответной реакции ткани на раздражение К возбудимым тканям относят: нервную, мышечную, секреторную
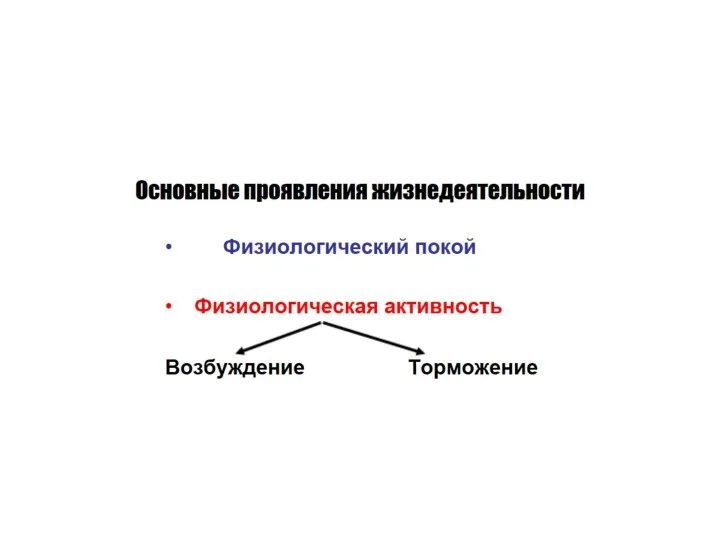
- 16. ВОЗБУДИМОСТЬ Это способность ткани отвечать на раздражение возбуждением (генерацией потенциала действия – ПД)
- 17. В основе всех реакций организма лежит раздражимость - способность реагировать на действие различных факторов изменением функции.
- 18. Свойства возбудимых тканей 1. Возбудимость – способность ткани отвечать возбуждением на раздражение 2. Проводимость – способность
- 19. Строение биологической мембраны В основе возбуждения лежит способность ткани изменять проницаемость мембран своих клеток для ионов,
- 20. поляризация Наличие разных зарядов по обе стороны мембраны: Снаружи + Внутри – Клетка представляет собой «диполь»
- 21. +
- 22. Величина МП плазмолеммы нервных клеток и кардиомиоцитов варьирует от –60 мВ до –90 мВ плазмолеммы скелетного
- 23. Градиент Это вектор, показывающий разницу между наибольшим и наименьшим значением какой-либо величины в разных точках пространства,
- 24. ФАКТОРЫ, ФОРМИРУЮЩИЕ МП ИОННАЯ АСИМЕТРИЯ Концентрационный градиент калия [Kin ] [Kex] Концентрационный градиент натрия [Naex] [Nain]
- 25. 2.Полупроницаемость мембраны K+ Na+ Cl- Белок- + + + + + + + + + +
- 26. «Электрический градиент» Это сила, создаваемая электрическим полем трансмембранной разности потенциалов Выход калия наружу уменьшает концентрационный градиент,
- 27. «Электрический градиент» Трансмембранная разность потенциалов создает электрическое поле, а следовательно и электрический градиент По мере выхода
- 28. Виды транспорта через биологическую мембрану Диффузия и облегченная диффузия идут по градиенту концентрации без затраты энергии,
- 29. Облегченная диффузия с участием белка-переносчика Белок-переносчик пребывает попеременно в одном из двух состояний – «пинг» и
- 30. Ионные каналы Это пути с воротами, которые могут находиться в открытом или закрытом состоянии и регулировать
- 31. Конформационные состояния ионного канала Ионный канал может находиться в состоянии покоя, активации и инактивации. Состояние покоя
- 32. Интегральный белок Лиганд-зависимый канал (калиевый, кальциевый) имеющий одни (активационные) ворота
- 33. Причины наличия мембранного потенциала 1 – концентрация ионов калия и натрия по обе стороны мембраны различна:
- 34. Работа натрий-калиевого насоса
- 35. Поляризация – поддержание разницы потенциалов по обе стороны мембраны Деполяризация – уменьшение величины мембранного потенциала Гиперполяризация
- 36. деполяризация Возникает при открытии натриевых каналов Натрий входит в клетку: уменьшает отрицательный заряд на внутренней поверхности
- 37. КРИТИЧЕСКИЙ УРОВЕНЬ ДЕПОЛЯРИЗАЦИИИ Екр Уровень деполяризации, при котором открывается максимально возможное количество натриевых каналов (все каналы
- 38. Порог деполяризации Разность между величиной исходной поляризации мембраны (Е0) и критическим уровнем деполяризации (Екр) Δ V=
- 39. Локальный ответ –местное возбуждение возникающее на подпороговый стимул
- 40. Свойства ЛО Не подчиняется закону «все или ничего» Амплитуда ЛО зависит от силы стимула Распространяется по
- 41. Потенциал действия (ПД) Это разность потенциалов между возбужденным и невозбужденным участками мембраны, которая возникает в результате
- 42. Потенциал действия (ПД) – кратковременное изменение разности потенциалов между наружной и внутренней поверхностями мембраны, возникающее в
- 43. Функциональные изменения натриевого канала при развитии ПД У натриевого канала два типа ворот: активационные и инактивационные.
- 44. Состояние проницаемости мембраны к ионам при развитии потенциала действия При действии раздражителя быстро открываются натриевые каналы.
- 45. Условия возникновения ПД Деполяризация должна достигнуть критического уровня деполяризации Ток натрия в клетку должен превышать ток
- 46. Фазы возбудимость Супернормальность первичная-локальный ответ; Абсолютная рефрактерность – отсутствие возбудимости регенеративная деполяризация и реверсия; Относительная рефрактерность
- 47. Фазы ПД Медленная деполяризация Быстрая деполяризация Инверсия Реверсия Быстрая реполяризация Медленная реполяризация Гиперполяризация
- 48. Первые электрофизиологические опыты. Луиджи Гальвани Он обратил внимание на сокращение препарата задних лапок лягушки при прикосновении
- 49. Первый «балконный» опыт Л.Гальвани (1786 г). Гальвани повторил этот опыт в условиях лаборатории, прикасаясь к препаратам
- 50. Алессандро Вольта А.Вольта повторил опыт Гальвани, но объяснил, полученные результаты тем, что в цепи из двух
- 51. Гальванический элемент. Вольтов столб В 1800 году А.Вольта опустил в банку с кислотой две пластинки —
- 52. Второй опыт Л. Гальвани Для того, чтобы доказать, что ткани животных тоже способны генерировать электрический ток
- 53. Дальнейшие исследования XIX век В 1838 г. Карло Маттеуччи доказал наличие разности электрических потенциалов между поврежденной
- 55. Скачать презентацию








Возбудимые ткани
Возбудимые ткани

Возбуждение – формирование специфической ответной реакции ткани на раздражение
К возбудимым тканям
К возбудимым тканям

ВОЗБУДИМОСТЬ
Это способность ткани отвечать на раздражение возбуждением (генерацией потенциала действия –
ВОЗБУДИМОСТЬ
Это способность ткани отвечать на раздражение возбуждением (генерацией потенциала действия –

В основе всех реакций организма лежит раздражимость - способность реагировать на
В основе всех реакций организма лежит раздражимость - способность реагировать на

Свойства возбудимых тканей
1. Возбудимость – способность ткани отвечать возбуждением на раздражение
2.
Свойства возбудимых тканей
1. Возбудимость – способность ткани отвечать возбуждением на раздражение
2.
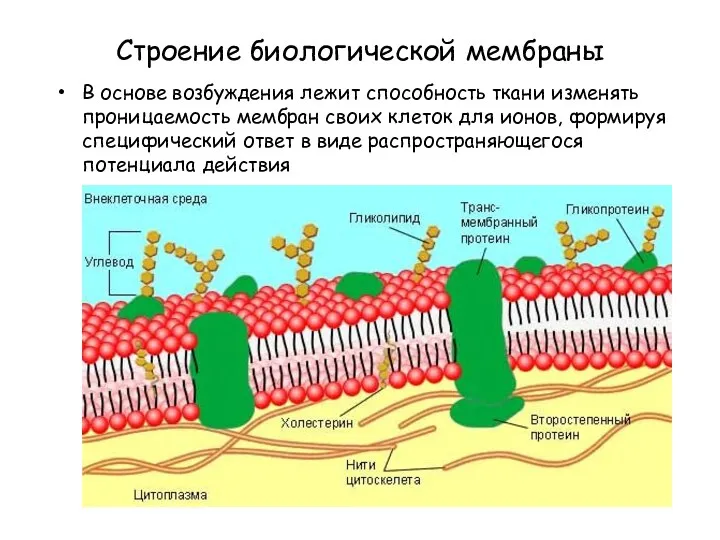
Строение биологической мембраны
В основе возбуждения лежит способность ткани изменять проницаемость мембран
Строение биологической мембраны
В основе возбуждения лежит способность ткани изменять проницаемость мембран

поляризация
Наличие разных зарядов по обе стороны
мембраны:
Снаружи +
Внутри –
Клетка представляет собой
поляризация
Наличие разных зарядов по обе стороны
мембраны:
Снаружи +
Внутри –
Клетка представляет собой
+
+

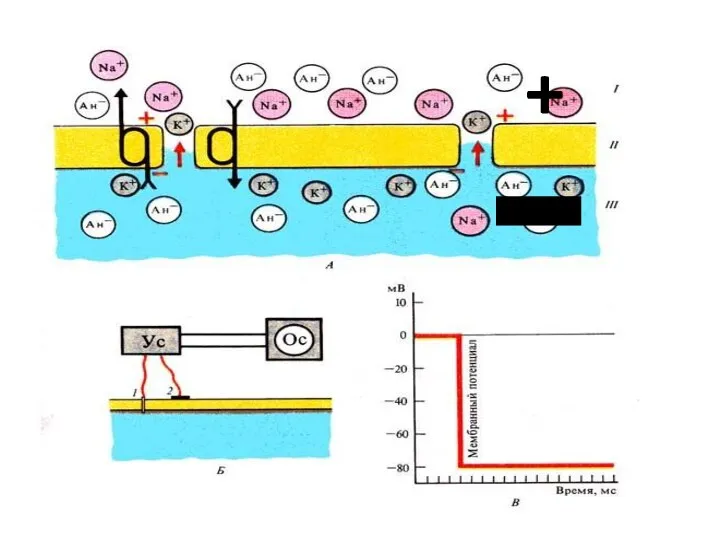
Величина МП
плазмолеммы нервных клеток и кардиомиоцитов варьирует от –60 мВ
Величина МП
плазмолеммы нервных клеток и кардиомиоцитов варьирует от –60 мВ

Градиент
Это вектор, показывающий разницу между наибольшим и наименьшим значением какой-либо величины
Градиент
Это вектор, показывающий разницу между наибольшим и наименьшим значением какой-либо величины
![ФАКТОРЫ, ФОРМИРУЮЩИЕ МП ИОННАЯ АСИМЕТРИЯ Концентрационный градиент калия [Kin ]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/364207/slide-23.jpg)
ФАКТОРЫ, ФОРМИРУЮЩИЕ МП
ИОННАЯ АСИМЕТРИЯ
Концентрационный градиент калия
[Kin ]
[Kex]
Концентрационный градиент натрия
[Naex]
ФАКТОРЫ, ФОРМИРУЮЩИЕ МП
ИОННАЯ АСИМЕТРИЯ
Концентрационный градиент калия
[Kin ]
[Kex]
Концентрационный градиент натрия
[Naex]
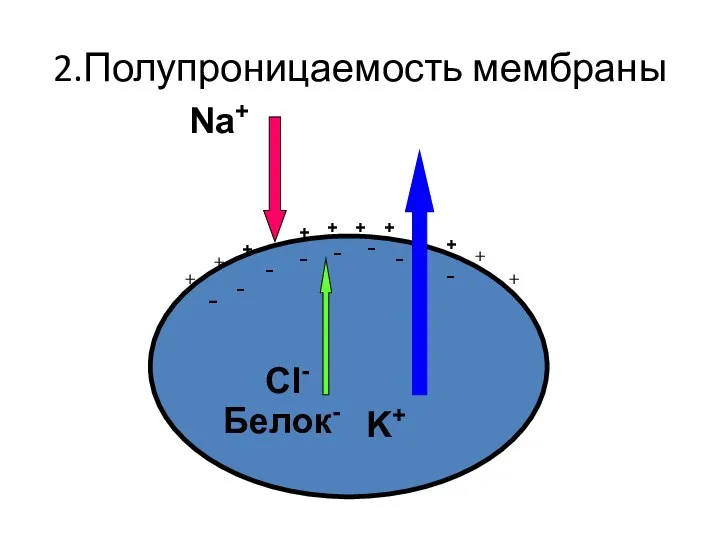
2.Полупроницаемость мембраны
K+
Na+
Cl-
Белок-
+
+
+
+
+
+
+
+
+
+
-
-
-
-
-
-
-
-
2.Полупроницаемость мембраны
K+
Na+
Cl-
Белок-
+
+
+
+
+
+
+
+
+
+
-
-
-
-
-
-
-
-

«Электрический градиент»
Это сила, создаваемая электрическим полем трансмембранной разности потенциалов
Выход
«Электрический градиент»
Это сила, создаваемая электрическим полем трансмембранной разности потенциалов
Выход

«Электрический градиент»
Трансмембранная разность потенциалов создает электрическое поле, а следовательно и
«Электрический градиент»
Трансмембранная разность потенциалов создает электрическое поле, а следовательно и
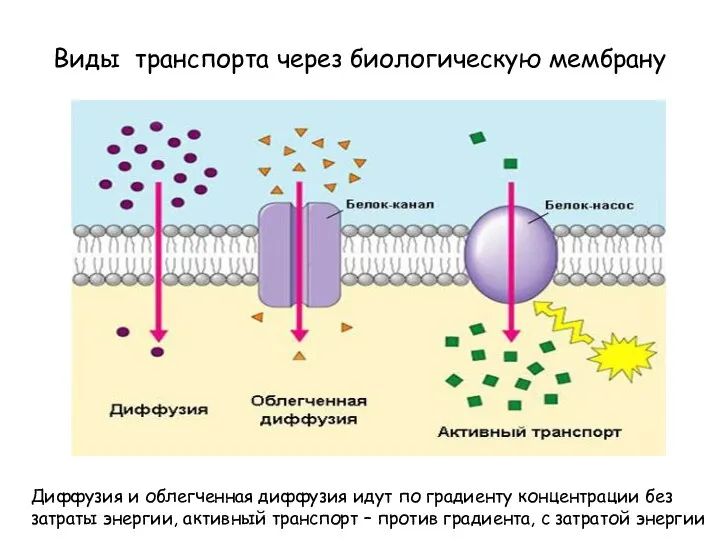
Виды транспорта через биологическую мембрану
Диффузия и облегченная диффузия идут по градиенту
Виды транспорта через биологическую мембрану
Диффузия и облегченная диффузия идут по градиенту
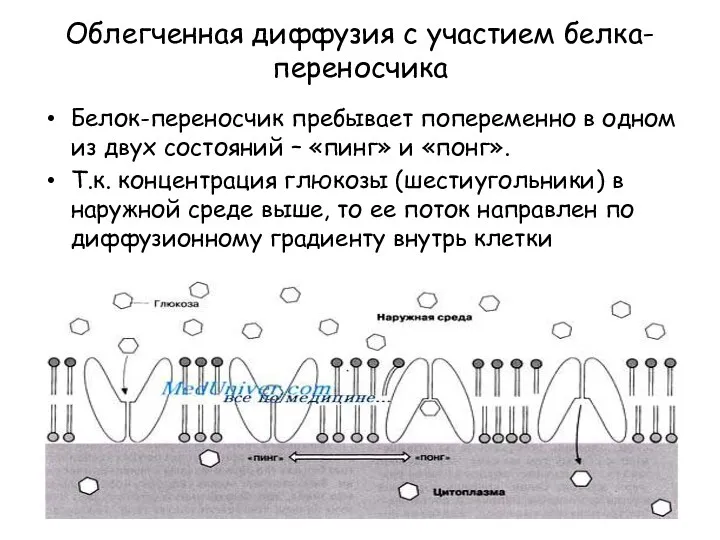
Облегченная диффузия с участием белка-переносчика
Белок-переносчик пребывает попеременно в одном из двух
Облегченная диффузия с участием белка-переносчика
Белок-переносчик пребывает попеременно в одном из двух

Ионные каналы
Это пути с воротами, которые могут находиться в открытом или
Ионные каналы
Это пути с воротами, которые могут находиться в открытом или
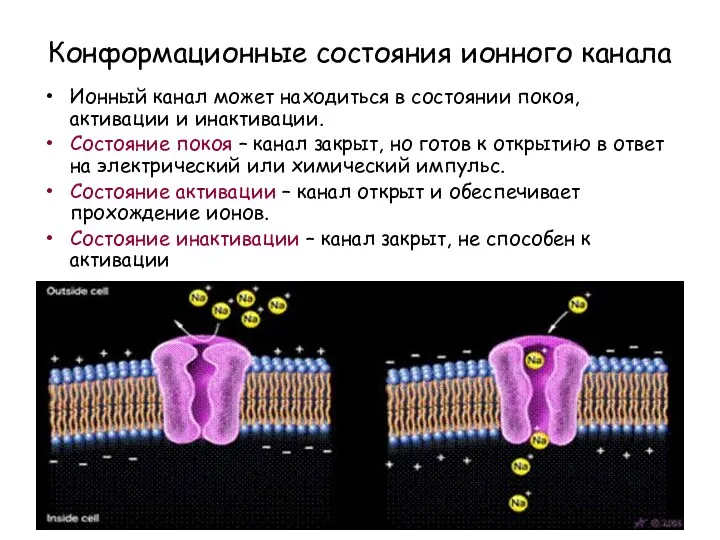
Конформационные состояния ионного канала
Ионный канал может находиться в состоянии покоя, активации
Конформационные состояния ионного канала
Ионный канал может находиться в состоянии покоя, активации
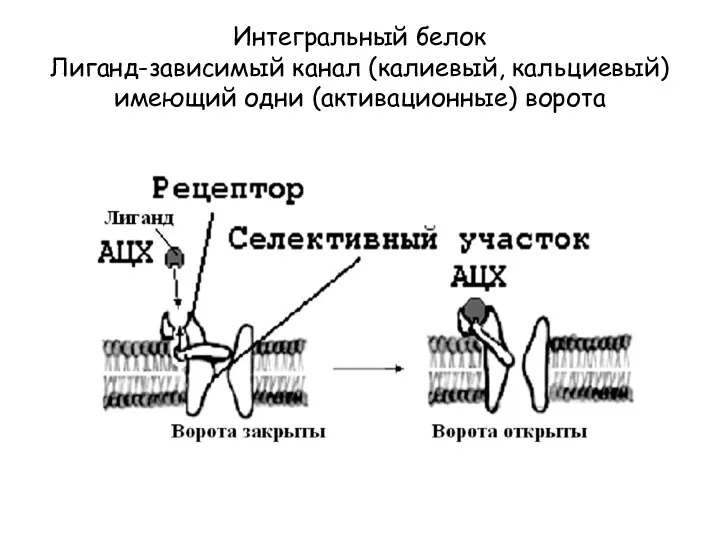
Интегральный белок
Лиганд-зависимый канал (калиевый, кальциевый) имеющий одни (активационные) ворота
Интегральный белок
Лиганд-зависимый канал (калиевый, кальциевый) имеющий одни (активационные) ворота

Причины наличия мембранного потенциала
1 – концентрация ионов калия и натрия по
Причины наличия мембранного потенциала
1 – концентрация ионов калия и натрия по
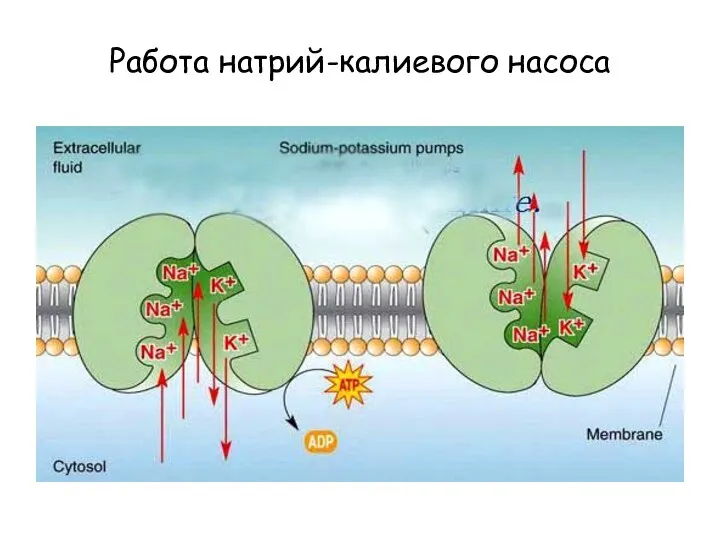
Работа натрий-калиевого насоса
Работа натрий-калиевого насоса

Поляризация – поддержание разницы потенциалов по обе стороны мембраны
Деполяризация – уменьшение
Поляризация – поддержание разницы потенциалов по обе стороны мембраны
Деполяризация – уменьшение

деполяризация
Возникает при открытии натриевых каналов
Натрий входит в клетку:
уменьшает отрицательный заряд
деполяризация
Возникает при открытии натриевых каналов
Натрий входит в клетку:
уменьшает отрицательный заряд

КРИТИЧЕСКИЙ УРОВЕНЬ ДЕПОЛЯРИЗАЦИИИ Екр
Уровень деполяризации, при котором открывается максимально возможное количество
КРИТИЧЕСКИЙ УРОВЕНЬ ДЕПОЛЯРИЗАЦИИИ Екр
Уровень деполяризации, при котором открывается максимально возможное количество

Порог деполяризации
Разность между величиной исходной поляризации мембраны (Е0) и критическим уровнем
Порог деполяризации
Разность между величиной исходной поляризации мембраны (Е0) и критическим уровнем

Локальный ответ –местное возбуждение возникающее на подпороговый стимул
Локальный ответ –местное возбуждение возникающее на подпороговый стимул

Свойства ЛО
Не подчиняется закону «все или ничего»
Амплитуда ЛО зависит от силы
Свойства ЛО
Не подчиняется закону «все или ничего»
Амплитуда ЛО зависит от силы

Потенциал действия (ПД)
Это разность потенциалов между возбужденным и невозбужденным участками мембраны,
Потенциал действия (ПД)
Это разность потенциалов между возбужденным и невозбужденным участками мембраны,
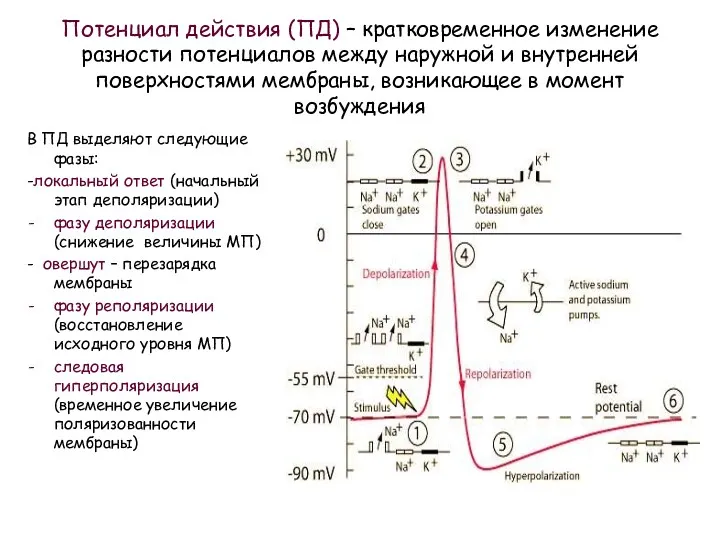
Потенциал действия (ПД) – кратковременное изменение разности потенциалов между наружной и
Потенциал действия (ПД) – кратковременное изменение разности потенциалов между наружной и

Функциональные изменения натриевого канала при развитии ПД
У натриевого канала два типа
Функциональные изменения натриевого канала при развитии ПД
У натриевого канала два типа
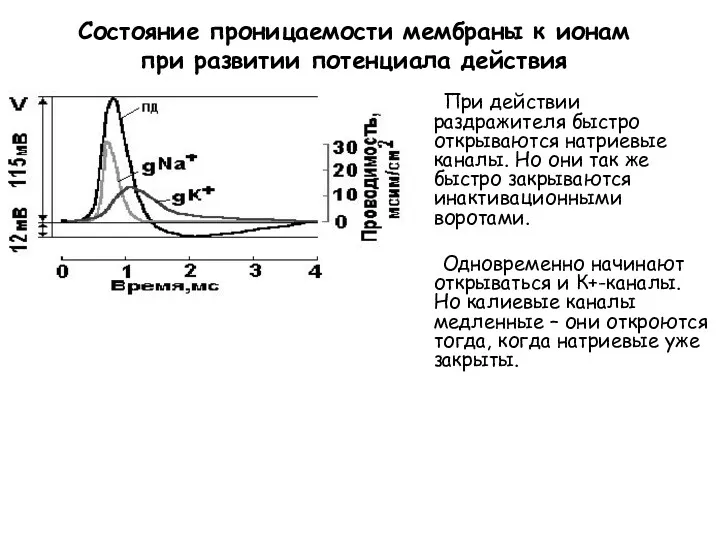
Состояние проницаемости мембраны к ионам при развитии потенциала действия
При действии раздражителя
Состояние проницаемости мембраны к ионам при развитии потенциала действия
При действии раздражителя

Условия возникновения ПД
Деполяризация должна достигнуть критического уровня деполяризации
Ток натрия в клетку
Условия возникновения ПД
Деполяризация должна достигнуть критического уровня деполяризации
Ток натрия в клетку

Фазы возбудимость
Супернормальность первичная-локальный ответ;
Абсолютная рефрактерность – отсутствие возбудимости регенеративная деполяризация и
Фазы возбудимость
Супернормальность первичная-локальный ответ;
Абсолютная рефрактерность – отсутствие возбудимости регенеративная деполяризация и

Фазы ПД
Медленная деполяризация
Быстрая деполяризация
Инверсия
Реверсия
Быстрая реполяризация
Медленная реполяризация
Гиперполяризация
Фазы ПД
Медленная деполяризация
Быстрая деполяризация
Инверсия
Реверсия
Быстрая реполяризация
Медленная реполяризация
Гиперполяризация

Первые электрофизиологические опыты.
Луиджи Гальвани
Он обратил внимание на сокращение препарата задних
Первые электрофизиологические опыты.
Луиджи Гальвани
Он обратил внимание на сокращение препарата задних
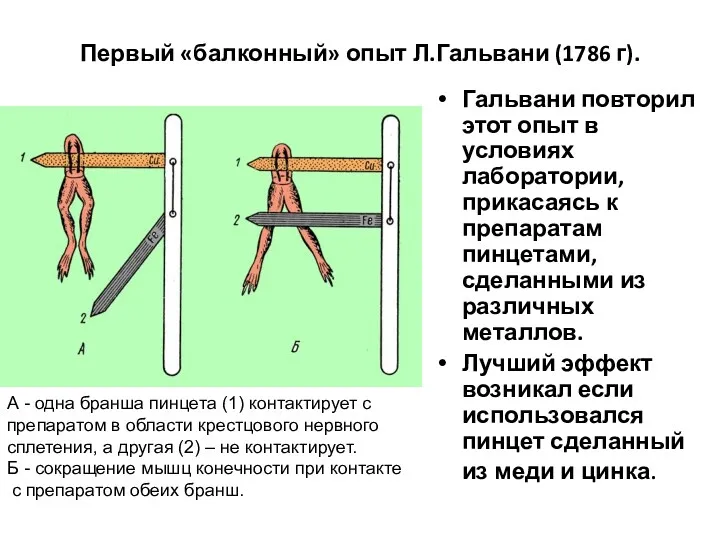
Первый «балконный» опыт Л.Гальвани (1786 г).
Гальвани повторил этот опыт в условиях
Первый «балконный» опыт Л.Гальвани (1786 г).
Гальвани повторил этот опыт в условиях

Алессандро Вольта
А.Вольта повторил опыт Гальвани, но объяснил, полученные результаты тем, что
Алессандро Вольта
А.Вольта повторил опыт Гальвани, но объяснил, полученные результаты тем, что

Гальванический элемент. Вольтов столб
В 1800 году А.Вольта опустил в банку с
Гальванический элемент. Вольтов столб
В 1800 году А.Вольта опустил в банку с
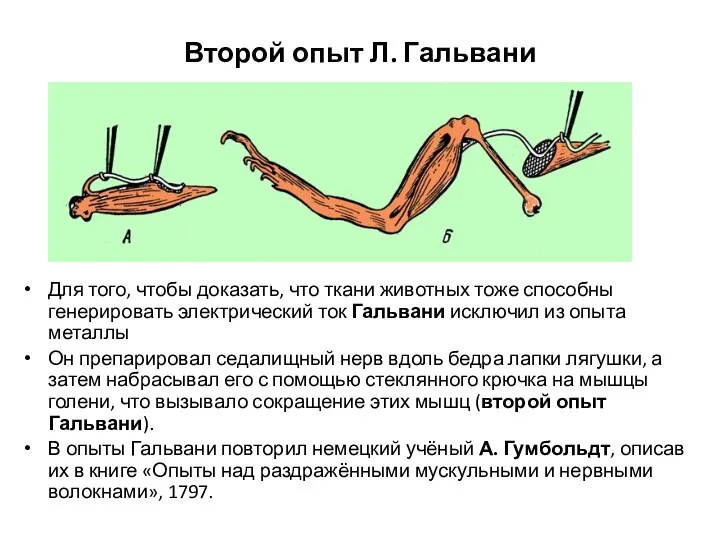
Второй опыт Л. Гальвани
Для того, чтобы доказать, что ткани животных тоже
Второй опыт Л. Гальвани
Для того, чтобы доказать, что ткани животных тоже
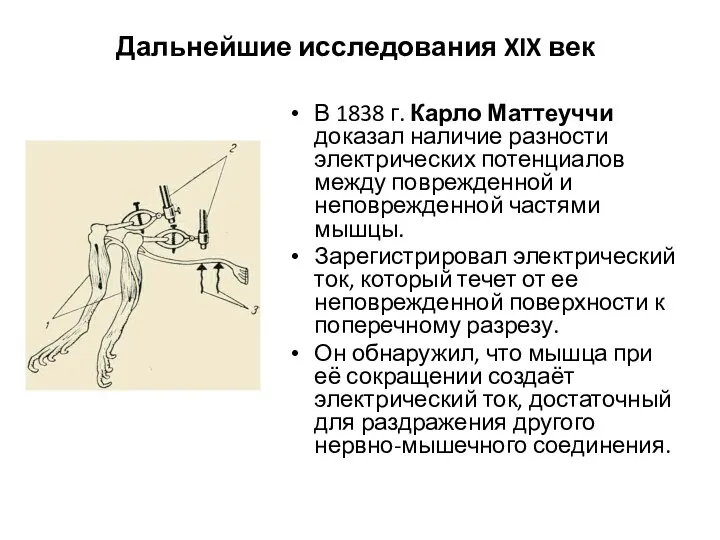
Дальнейшие исследования XIX век
В 1838 г. Карло Маттеуччи доказал наличие разности
Дальнейшие исследования XIX век
В 1838 г. Карло Маттеуччи доказал наличие разности
 Классификация вирусов по Балтимору
Классификация вирусов по Балтимору Гистология и цитология, их содержание, задачи, связь с другими науками, значение
Гистология и цитология, их содержание, задачи, связь с другими науками, значение Решение задач по молекулярной биологии. Подготовка к ЕГЭ (11 класс). Тренинг-практикум
Решение задач по молекулярной биологии. Подготовка к ЕГЭ (11 класс). Тренинг-практикум жизнь в морях и океанах(1)
жизнь в морях и океанах(1) Ткани человека. Гуморальная система. Нервная система
Ткани человека. Гуморальная система. Нервная система Отряды парнокопытные и непарнокопытные
Отряды парнокопытные и непарнокопытные Вкусовой и обонятельный анализаторы. Боль
Вкусовой и обонятельный анализаторы. Боль Генотоксиканты. Классификации мутагенов. Генотоксические поражения и болезни
Генотоксиканты. Классификации мутагенов. Генотоксические поражения и болезни Как сделать транскриптон? Алгоритм действий
Как сделать транскриптон? Алгоритм действий Анатомия и физиология человека
Анатомия и физиология человека Общий путь катаболизма. (Лекция 9)
Общий путь катаболизма. (Лекция 9) Электронные версии гистологических препаратов для сдачи ОСПЭ
Электронные версии гистологических препаратов для сдачи ОСПЭ Параретровіруси. Pararetroviruses vs. Retroviruses
Параретровіруси. Pararetroviruses vs. Retroviruses Презентация по биологии на тему: Обобщающий урок по темам: Прокариоты. Грибы. Растения. 7 класс.
Презентация по биологии на тему: Обобщающий урок по темам: Прокариоты. Грибы. Растения. 7 класс. Хрящевые рыбы
Хрящевые рыбы Растительноядные, хищные, паразиты и сверх паразиты среди представителей насекомых
Растительноядные, хищные, паразиты и сверх паразиты среди представителей насекомых Восприятие. Отличия восприятие и ощущения
Восприятие. Отличия восприятие и ощущения Митоз и мейоз. Митоз, или непрямое деление
Митоз и мейоз. Митоз, или непрямое деление Экология микроорганизмов. Микроэкология полости рта
Экология микроорганизмов. Микроэкология полости рта Проверочная работа по теме Систематика растений 6 класс
Проверочная работа по теме Систематика растений 6 класс Дикие животные
Дикие животные Черепно-мозговые нервы
Черепно-мозговые нервы Съедобные и несъедобные грибы
Съедобные и несъедобные грибы Презентация Птицы - наши друзья!
Презентация Птицы - наши друзья! Внутрішня будова ссавців
Внутрішня будова ссавців Анатомия головного мозга
Анатомия головного мозга Кто такие рыбы (окружающий мир, 1 класс)
Кто такие рыбы (окружающий мир, 1 класс) Russian animals in danger
Russian animals in danger